
Anti-Neuroinflammatory Potential of a Nectandra angustifolia (Laurel Amarillo) Ethanolic Extract

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Plant Material, Preparation of Ethanol Extract and Chemical Characterization
2.3. Cell Culture
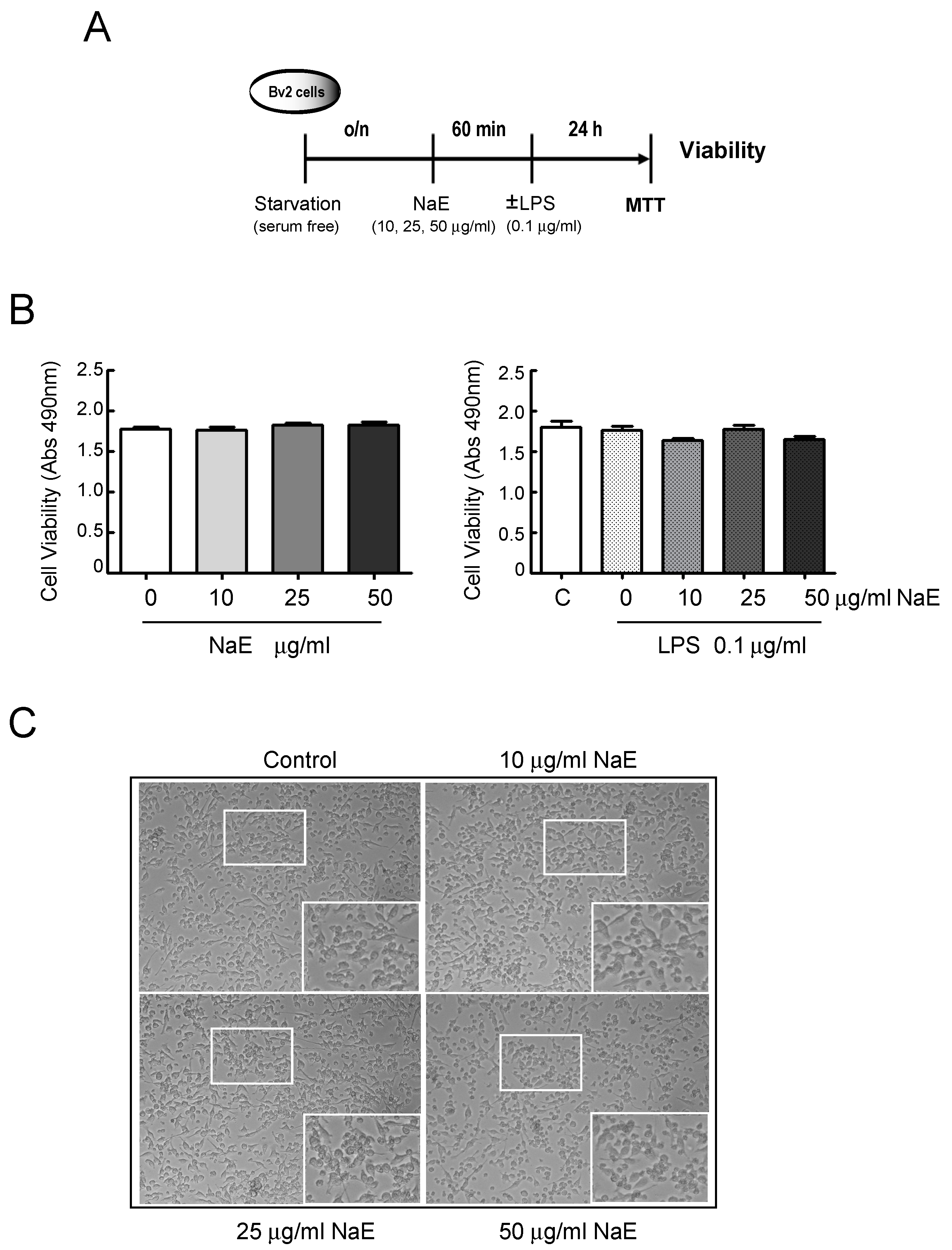
2.4. Viability Assay
2.5. Flow Cytometric (FC) Analysis
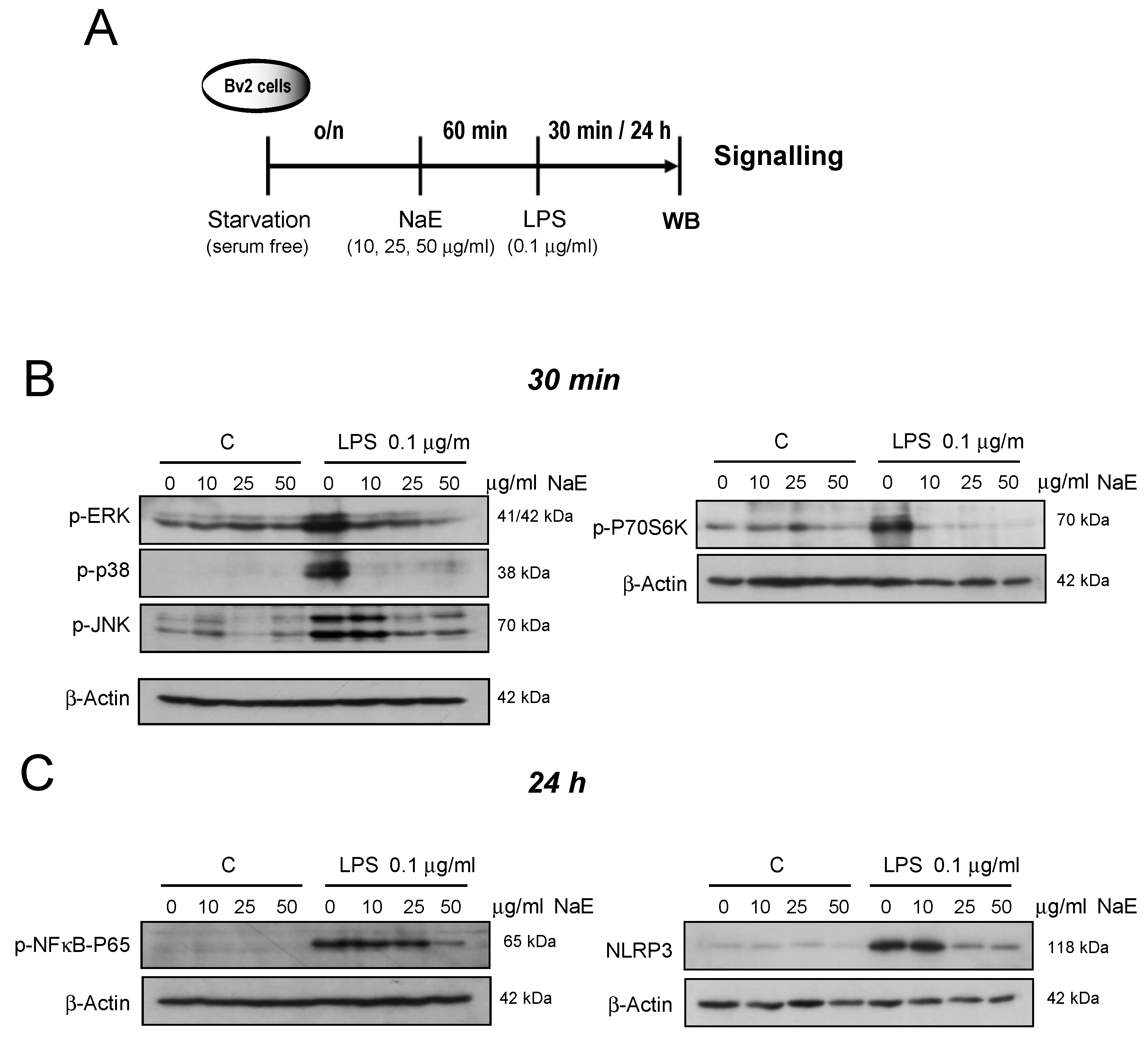
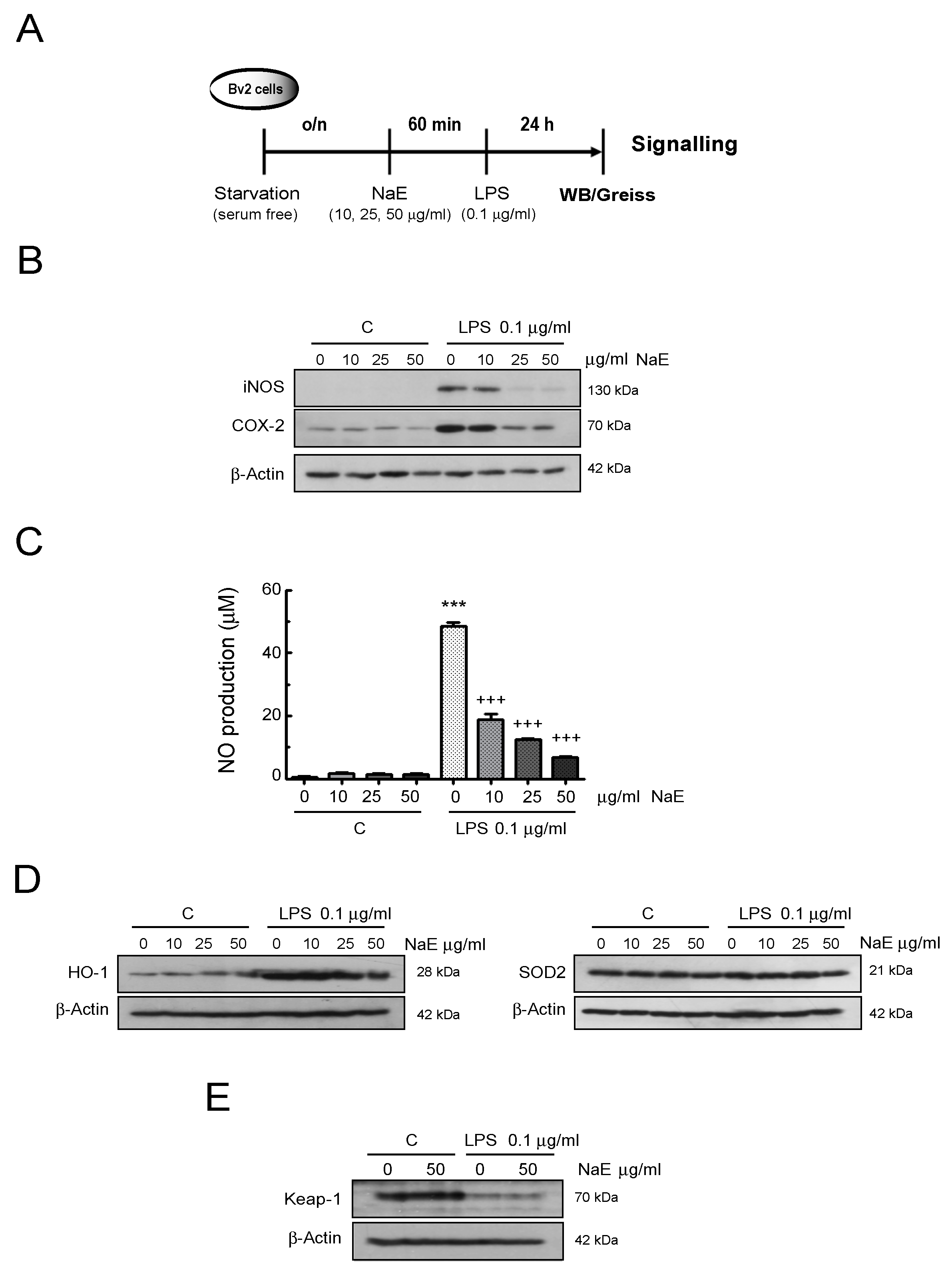
2.6. Western Blot (WB) Analysis
2.7. Measurement of Intracellular Superoxide Radicals
2.8. Measurement of Lipid Peroxidation
2.9. NO Assay
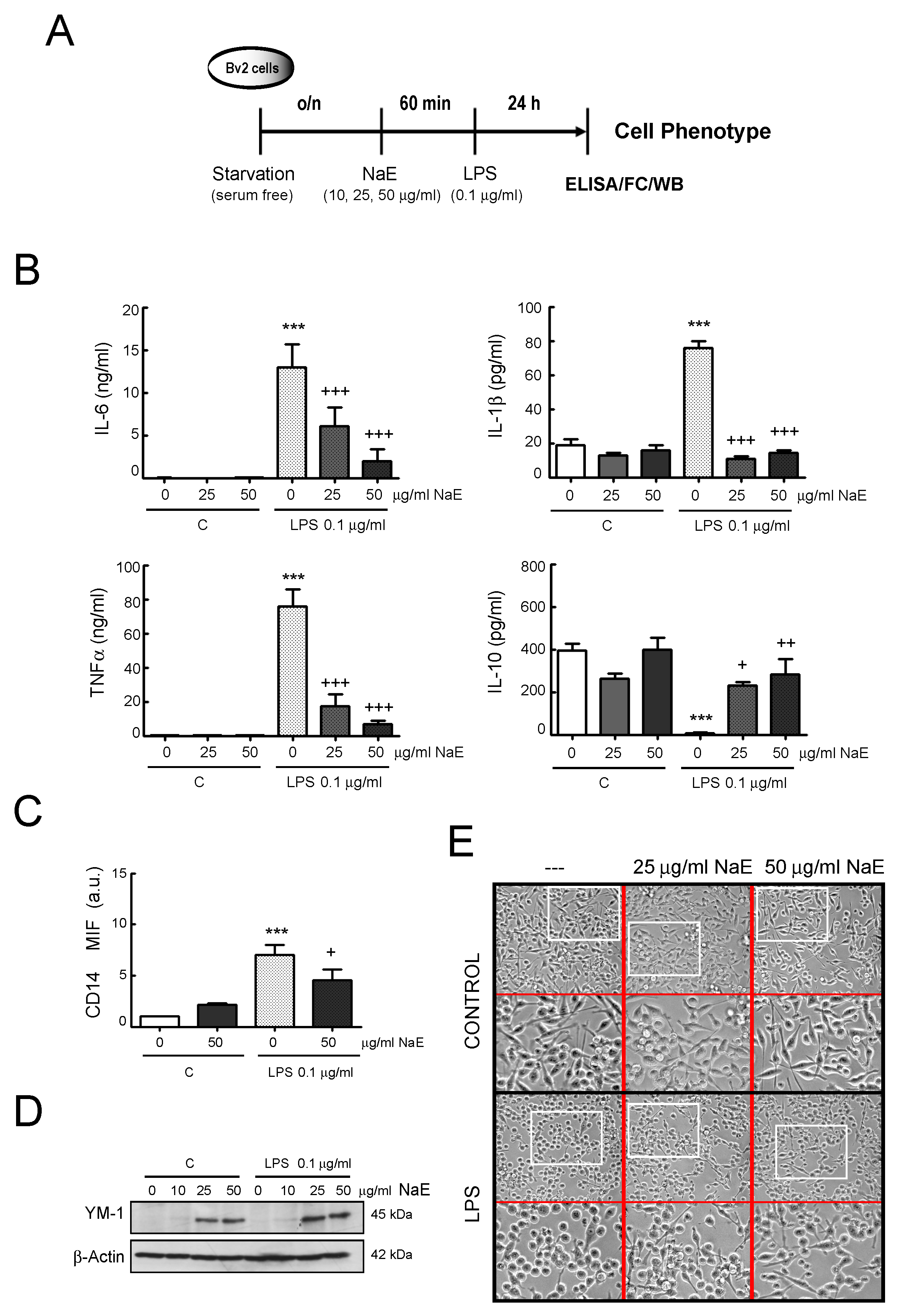
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Scratch Test Assay
2.12. Phagocytosis Assay
2.13. Animals
2.14. Real-Time Polymerase Chain Reaction Analysis
2.15. Statistical Analysis
3. Results
3.1. Effects of NaE on the Inflammatory Response Induced by LPS on BV2 Cells
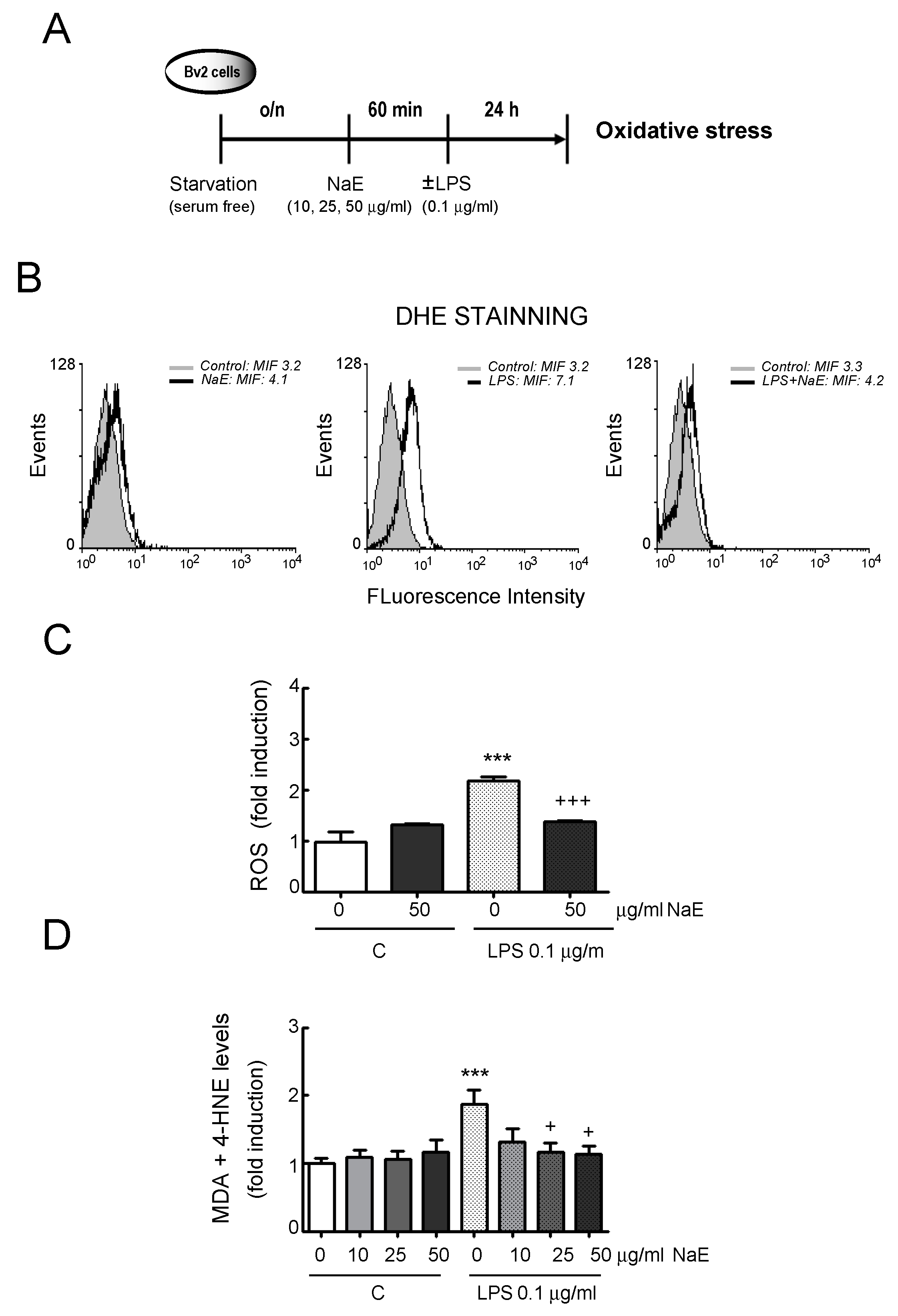
3.2. NaE Modulates LPS-Mediated BV2 Cells’ Oxidative Stress
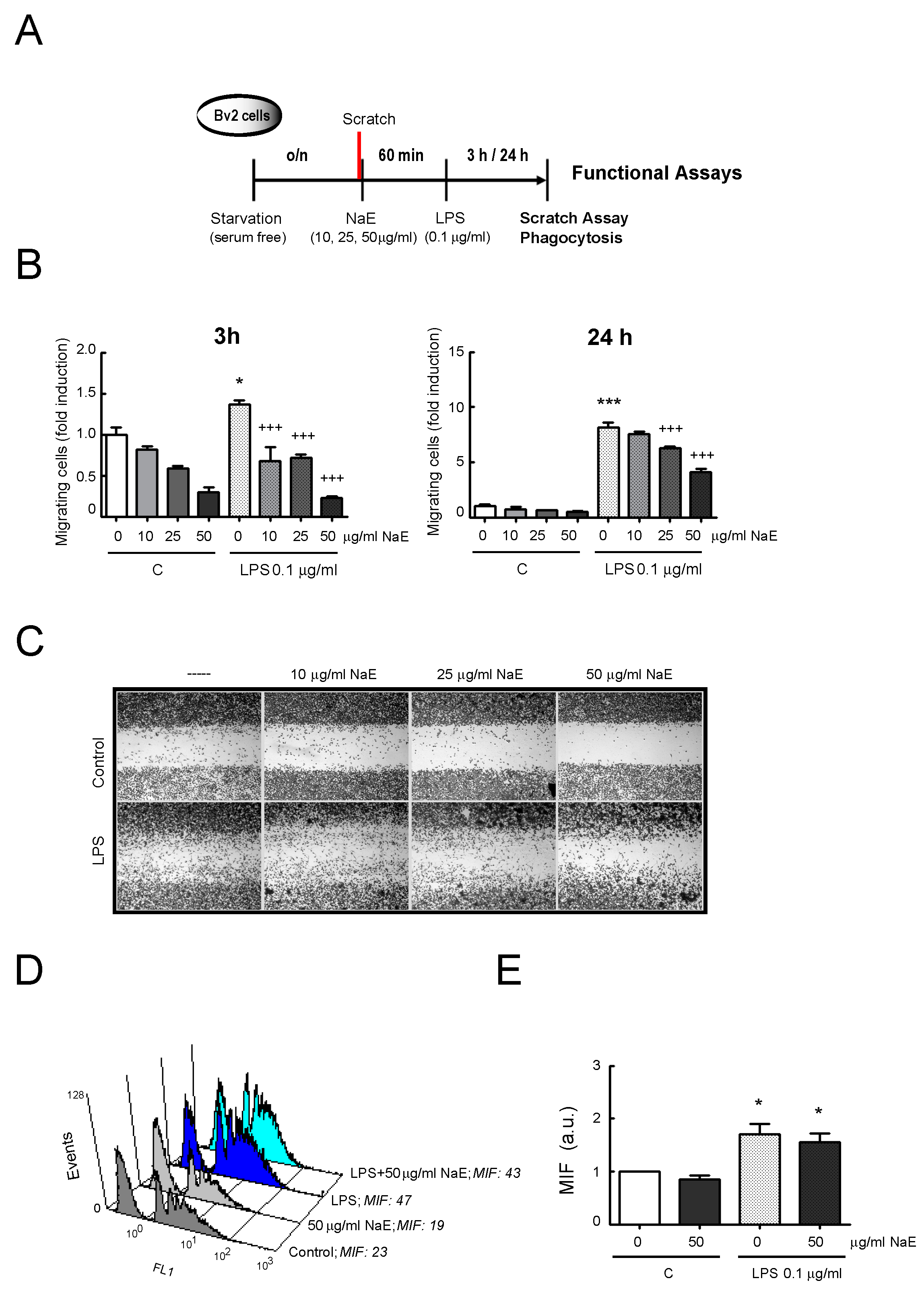
3.3. NaE Modulates LPS-Mediated BV2 Cell Migration
3.4. Effect of NaE on BV2 Cell Phagocytosis
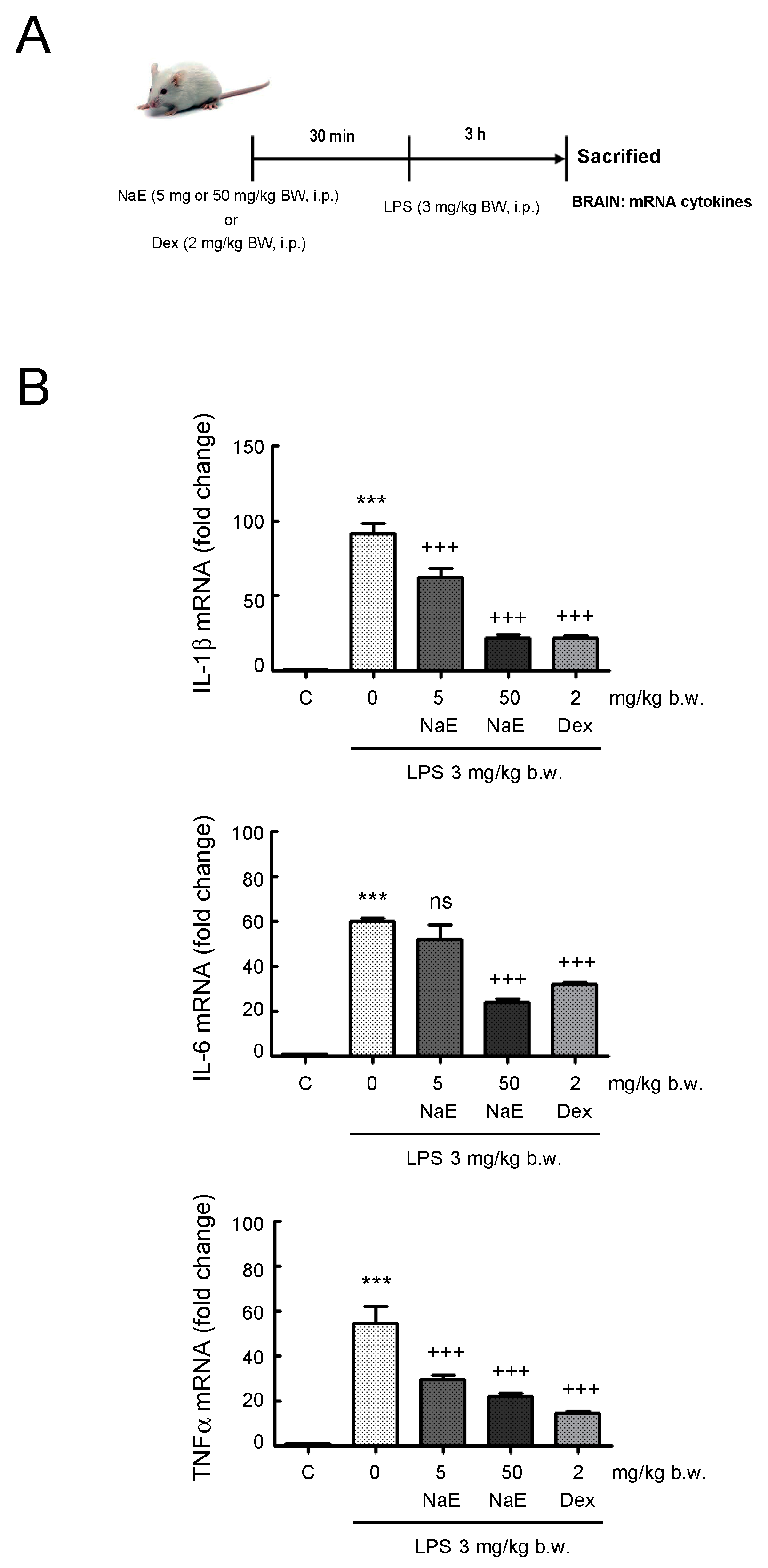
3.5. NaE Modulates In Vivo Neuroinflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graeber, M.B.; Li, W.; Rodriguez, M.L. Role of microglia in CNS inflammation. FEBS Lett. 2011, 585, 3798–3805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.Q.; Zhou, J.W. Neuroinflammation in the central nervous system: Symphony of glial cells. Glia 2019, 67, 1017–1035. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Perry, V.H. Microglial physiology: Unique stimuli, specialized responses. Annu. Rev. Immunol. 2009, 27, 119–145. [Google Scholar] [CrossRef]
- Kirkley, K.S.; Popichak, K.A.; Afzali, M.F.; Legare, M.E.; Tjalkens, R.B. Microglia amplify inflammatory activation of astrocytes in manganese neurotoxicity. J. Neuroinflammation. 2017, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Vonkeman, H.E.; van de Laar, M.A.F.J. Nonsteroidal anti-inflammatory drugs: Adverse effects and their prevention. Semin. Arthritis Rheum. 2010, 39, 294–312. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.M.; Woessner, K.M. An Overview of Nonsteroidal Antiinflammatory Drug Reactions. Immunol. Allergy Clin. North. Am. 2016, 36, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Yoldi, M.J. Anti-Inflammatory and Antioxidant Properties of Plant Extracts. Antioxidants 2021, 10, 921. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Antony, P.J.; Lana, M.J.M.P.; da Silva, B.F.X.; Oliveira, R.V.; Jothi, G.; Hariharan, G.; Mohana, T.; Gan, R.Y.; Gurgel, R.Q.; et al. Natural products modulating interleukins and other inflammatory mediators in tumor-bearing animals: A systematic review. Phytomedicine 2022, 100, 154038. [Google Scholar] [CrossRef]
- Luthra, R.; Roy, A. Role of Medicinal Plants against Neurodegenerative Diseases. Curr. Pharm. Biotechnol. 2022, 23, 123–139. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, C.D.R.; Barreto Arantes, M.; Menezes de Faria Pereira, S.; da Cruz, L.L.; de Souza Passos, M.; Pereira de Moraes, L.; Curcino Vieira, I.J.; Barros de Oliveira, D. Plants as Sources of Anti-Inflammatory Agents. Molecules 2020, 25, 3726. [Google Scholar] [CrossRef] [PubMed]
- Zuloaga, F.O.; Morrone, O.; Rodríguez, D. Análisis de la biodiversidad de plantas vasculares de la Argentina. Kurtziana 1999, 27, 17–167. [Google Scholar]
- Campos-Navarro, R.; Scarpa, G.F. The cultural-bound disease "empacho" in Argentina. A comprehensive botanico-historical and ethnopharmacological review. J. Ethnopharmacol. 2013, 148, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Haretche, F.; Olivaro, C.; Vázquez, A. Prospección química del bosque de galería del río Uruguay. Rev. Bras. Farmacogn. 2008, 18, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Alves Conserva, G.A.; Quirós-Guerrero, L.M.; Costa-Silva, T.A.; Marcourt, L.; Pinto, E.G.; Tempone, A.G.; Fernandes, J.P.S.; Wolfender, J.L.; Queiroz, E.F.; Lago, J.H.G. Metabolite profile of Nectandra oppositifolia Nees & Mart. and assessment of antitrypanosomal activity of bioactive compounds through efficiency analyses. PLoS ONE 2021, 16, e0247334. [Google Scholar]
- González Torres, D.M. Catálogo de Plantas Medicinales (y Alimenticias y Ứtiles) Usadas en Paraguay; Servilibro: Asunción, Paraguay, 1992. [Google Scholar]
- Macías-Villamizar, V.E.; Cuca-Suárez, L.E.; Coy-Barrera, E.D. Genus Nectandra: Phytochemistry and biological activity. Bol. Latinoam. Caribe Plantas Med. Aromát. 2015, 14, 317–342. [Google Scholar]
- Ferrini, L.; Rodríguez, J.P.; Melana Colavita, J.P.; Olea, G.; Ojeda, G.A.; Ricciardi, G.; Torres, A.M.; Aguirre, M.V. Anti-inflammatory activity of Nectandra angustifolia (Laurel Amarillo) ethanolic extract. J. Ethnopharmacol. 2021, 272, 113937. [Google Scholar] [CrossRef]
- Oliveira de Melo, J.; da Conceição Torrado Truiti, M.; Muscará, M.N.; Bolonheis, S.M.; Dantas, J.A.; Caparroz-Assef, S.M.; Cuman, R.K.; Bersani-Amado, C.A. Anti-inflammatory activity of crude extract and fractions of nectandra falcifolia leaves. Biol. Pharm. Bull. 2006, 29, 2241–2245. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.M.; Ojeda, G.A.; Angelina, E.; Bustillo, S.; Peruchena, N.; Tonidandel, L.; Larcher, R.; Nardin, T.; Dellacassa, E. The anti-snake activity of Nectandra angustifolia flavonoids on phospholipase A2: In vitro and in silico evaluation. J. Ethnopharmacol. 2023, 302, 115889. [Google Scholar] [CrossRef]
- Bocchini, V.; Mazzolla, R.; Barluzzi, R.; Blasi, E.; Sick, P.; Kettenmann, H. An immortalized cell line expresses properties of activated microglial cells. J. Neurosci. Res. 1992, 31, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Ibeas, E.; Fuentes, L.; Martín, R.; Hernández, M.; Nieto, M.L. Secreted phospholipase A2 type IIA as a mediator connecting innate and adaptive immunity: New role in atherosclerosis. Cardiovas. Res. 2009, 81, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Liu, T.; Liu, Y.; Jiang, Y.; Seshadri, V.D.D.; Mohan, S.M.; Ling, L. Neuroprotective effect of biosynthesised gold nanoparticles synthesised from root extract of Paeonia moutan against Parkinson disease—In vitro &in vivo model. J. Photochem. Photobiol. B 2019, 200, 111635. [Google Scholar] [PubMed]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepeda, S.B.; Sandoval, M.J.; Crescitelli, M.C.; Rauschemberger, M.B.; Massheimer, V.L. The isoflavone genistein enhances osteoblastogenesis: Signaling pathways involved. J. Physiol. Biochem. 2020, 76, 9–110. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Castaño, A.; Herrera, A.J.; Cano, J.; Machado, A. The degenerative effect of a single intranigral injection of LPS on the dopaminergic system is prevented by dexamethasone, and not mimicked by rh-TNF-alpha, IL-1beta and IFN-gamma. J. Neurochem. 2002, 81, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Overbergh, L.; Giulietti, A.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. The use of RT-qPCR for the quantification of cytokine gene expression. J. Biomol. Tech. 2003, 14, 33–43. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔ Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Motterlini, R.; Green, C.J.; Foresti, R. Regulation of heme oxygenase-1 by redox signals involving nitric oxide. Antioxid. Redox Signal. 2002, 4, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Obuobi, S.; Karatayev, S.; Chai, C.L.; Ee, P.L.R.; Mátyus, P. The role of modulation of antioxidant enzyme systems in the treatment of neurodegenerative diseases. J. Enzyme Inhib. Med. Chem. 2016, 31, 194–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, A.; Takasu, K.; Sawada, M.; Suzuki, H. Hemokinin-1 gene expression is upregulated in microglia activated by lipopolysaccharide through NF-κB and p38 MAPK signaling pathways. PLoS ONE 2012, 7, e32268. [Google Scholar] [CrossRef]
- Schlegelmilch, T.; Henke, K.; Peri, F. Microglia in the developing brain: From immunity to behavior. Curr. Opin. Neurobiol. 2011, 21, 5–10. [Google Scholar] [CrossRef]
- Neumann, H.; Kotter, M.R.; Franklin, R.J. Debris clearance by microglia: An essential link between degeneration and regeneration. Brain 2009, 132, 288–295. [Google Scholar] [CrossRef]
- Ishijima, T.; Nakajima, K. Inflammatory cytokines TNFα, IL-1β, and IL-6 are induced in endotoxin- stimulated microglia through different signaling cascades. Sci. Prog. 2021, 104, 368504211054985. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Park, S.M.; Choi, M.S.; Sohn, N.W.; Shin, J.W. Ginsenoside Rg3 attenuates microglia activation following systemic lipopolysaccharide treatment in mice. Biol. Pharm. Bull. 2012, 35, 1546–1552. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro Alvares Batista, C.; Freitas Gomes, G.; Candelario-Jalil, E.; Fiebich, B.L.; Pinheiro de Oliveira, A.C. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [Green Version]
- Grubić Kezele, T.; Ćurko-Cofek, B. Neuroprotective Panel of Olive Polyphenols: Mechanisms of Action, Anti-Demyelination, and Anti-Stroke Properties. Nutrients 2022, 14, 4533. [Google Scholar] [CrossRef] [PubMed]
- Dheen, S.T.; Kaur, C.; Ling, E. Microglial activation and its implications in the brain diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Moore, P.K. Dexamethasone inhibits lipopolysaccharide-induced hydrogen sulphide biosynthesis in intact cells and in an animal model of endotoxic shock. J. Cell. Mol. Med. 2009, 13, 2684–2692. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Watanabe, S. Differential modulation of lipopolysaccharide- and zymosan-induced hypophagia by dexamethasone treatment. Pharmacol. Biochem. Behav. 2008, 90, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid permeability across an in situ model of the blood-brain barrier. Free Radical Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols journey through blood-brain barrier towards neuronal protection. Sci. Rep. 2017, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization from M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Ye, X.; Zhu, M.; Che, X.; Wang, H.; Liang, X.-J.; Wu, C.; Xue, X.; Yang, J. Lipopolysaccharide induces neuroinflammation in microglia by activating the MTOR pathway and downregulating Vps34 to inhibit autophagosome formation. J. Neuroinflammation 2020, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- García-Aguilar, A.; Palomino, O.; Benito, M.; Guillén, C. Dietary Polyphenols in Metabolic and Neurodegenerative Diseases: Molecular Targets in Autophagy and Biological Effects. Antioxidants 2021, 10, 142. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Duan, H.; Li, R.; Peng, W.; Wu, C. Activation of Nrf2/HO-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress. J. Adv. Res. 2021, 34, 43–63. [Google Scholar] [CrossRef]
- Zhao, F.; Ci, X.; Man, X.; Li, J.; Wei, Z.; Zhang, S. Food-Derived Pharmacological Modulators of the Nrf2/ARE Pathway: Their Role in the Treatment of Diseases. Molecules 2021, 26, 1016. [Google Scholar] [CrossRef]
- Yin, S.; Cao, W. Toll-Like Receptor Signaling Induces Nrf2 Pathway Activation through p62-Triggered Keap1 Degradation. Mol. Cell. Biol. 2015, 35, 2673–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Hou, D.H. Multiple regulations of Keap1/Nrf2 system by dietary phytochemicals. Mol. Nutr. Food Res. 2016, 60, 1731–1755. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [Green Version]
- Do, H.T.T.; Bui, B.P.; Sim, S.; Jung, J.K.; Lee, H.; Cho, J. Anti-Inflammatory and Anti-Migratory Activities of Isoquinoline-1-Carboxamide Derivatives in LPS-Treated BV2 Microglial Cells via Inhibition of MAPKs/NF-κB Pathway. Int. J. Mol. Sci. 2020, 21, 2319. [Google Scholar] [CrossRef] [Green Version]
- De Caris, M.G.; Grieco, M.; Maggi, E.; Francioso, A.; Armeli, F.; Mosca, L.; Pinto, A.; D’Erme, M.; Mancini, P.; Businaro, R. Blueberry Counteracts BV-2 Microglia Morphological and Functional Switch after LPS Challenge. Nutrients 2020, 12, 1830. [Google Scholar] [CrossRef]
- Uhlemann, R.; Gertz, K.; Boehmerle, W.; Schwarz, T.; Nolte, C.; Freyer, D.; Kettenmann, H.; Endres, M.; Kronenberg, G. Actin dynamics shape microglia effector functions. Brain Struct. Funct. 2016, 221, 2717–2734. [Google Scholar] [CrossRef]
- Bettegazzi, B.; Bellani, S.; Cattaneo, S.; Codazzi, F.; Grohovaz, F.; Zacchetti, D. Gα13 Contributes to LPS-Induced Morphological Alterations and Affects Migration of Microglia. Mol. Neurobiol. 2021, 58, 6397–6414. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crescitelli, M.C.; Simon, I.; Ferrini, L.; Calvo, H.; Torres, A.M.; Cabero, I.; Panedas, M.M.; Rauschemberger, M.B.; Aguirre, M.V.; Rodríguez, J.P.; et al. Anti-Neuroinflammatory Potential of a Nectandra angustifolia (Laurel Amarillo) Ethanolic Extract. Antioxidants 2023, 12, 232. https://doi.org/10.3390/antiox12020232
Crescitelli MC, Simon I, Ferrini L, Calvo H, Torres AM, Cabero I, Panedas MM, Rauschemberger MB, Aguirre MV, Rodríguez JP, et al. Anti-Neuroinflammatory Potential of a Nectandra angustifolia (Laurel Amarillo) Ethanolic Extract. Antioxidants. 2023; 12(2):232. https://doi.org/10.3390/antiox12020232
Chicago/Turabian StyleCrescitelli, María Carla, Inmaculada Simon, Leandro Ferrini, Hugo Calvo, Ana M. Torres, Isabel Cabero, Mónica Macías Panedas, Maria B. Rauschemberger, Maria V. Aguirre, Juan Pablo Rodríguez, and et al. 2023. "Anti-Neuroinflammatory Potential of a Nectandra angustifolia (Laurel Amarillo) Ethanolic Extract" Antioxidants 12, no. 2: 232. https://doi.org/10.3390/antiox12020232