
Two Argan Oil Phytosterols, Schottenol and Spinasterol, Attenuate Oxidative Stress and Restore LPS-Dysregulated Peroxisomal Functions in Acox1−/− and Wild-Type BV-2 Microglial Cells

 , ,
, ,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Treatments
2.2. MTT Test
2.3. Griess Test
2.4. Measurement of Intracellular ROS
2.5. Cell Homogenate Preparation
2.6. Catalase Activity Measurement
2.7. Acyl-CoA Oxidase 1 Activity Measurement
2.8. Immunoblotting
2.9. Quantitative Reverse-Transcription-PCR
2.10. Data Analysis
3. Results
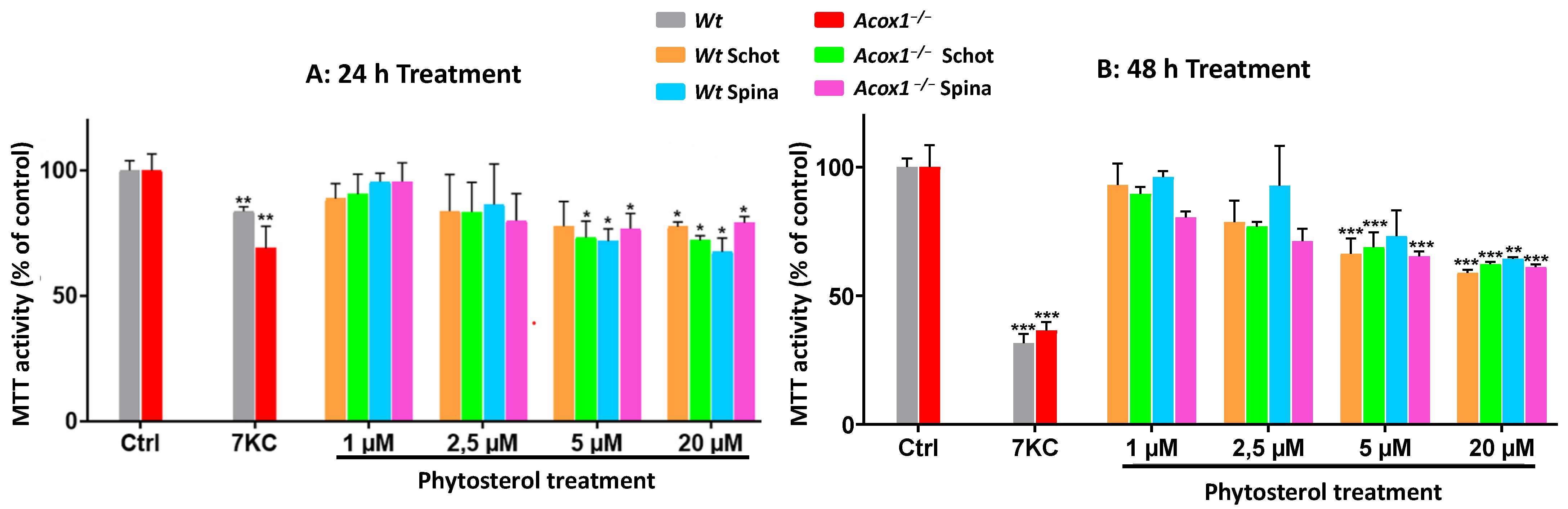
3.1. Effects of Schot and Spina on Cell Viability
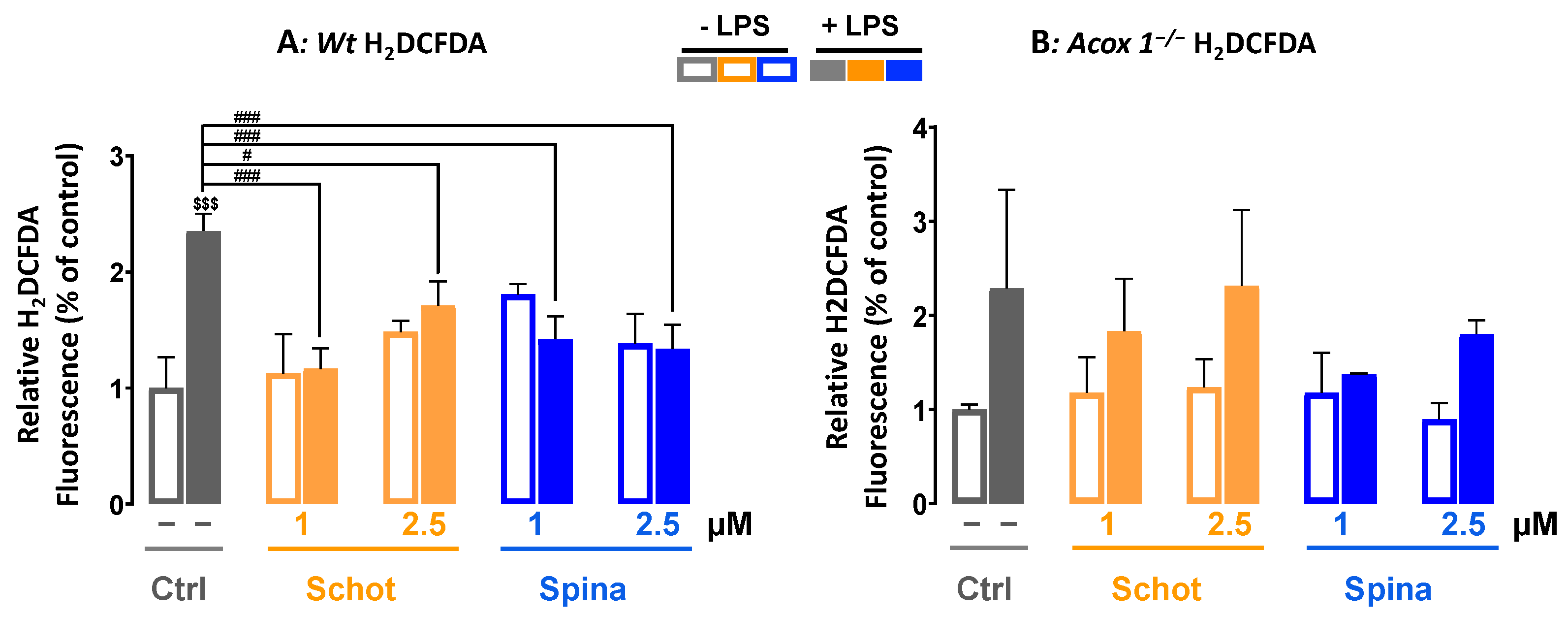
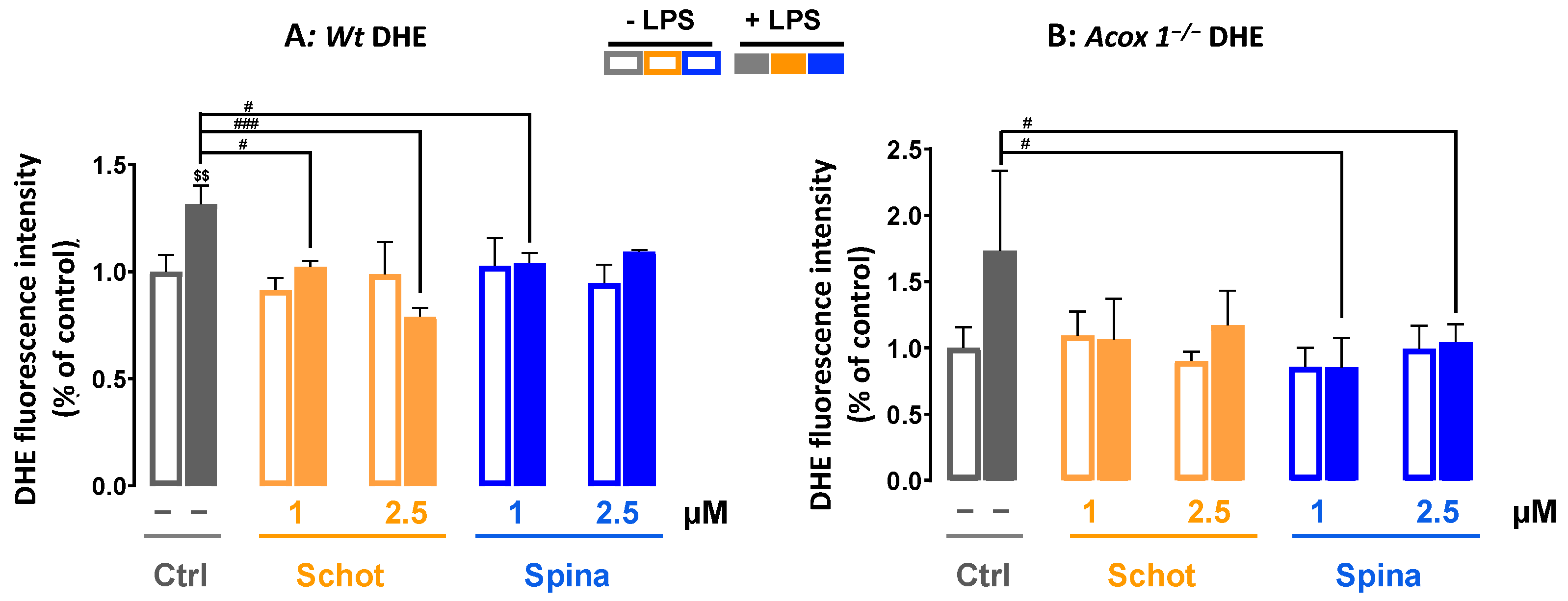
3.2. Schot and Spina Effects on LPS-Induced Intracellular ROS Accumulation
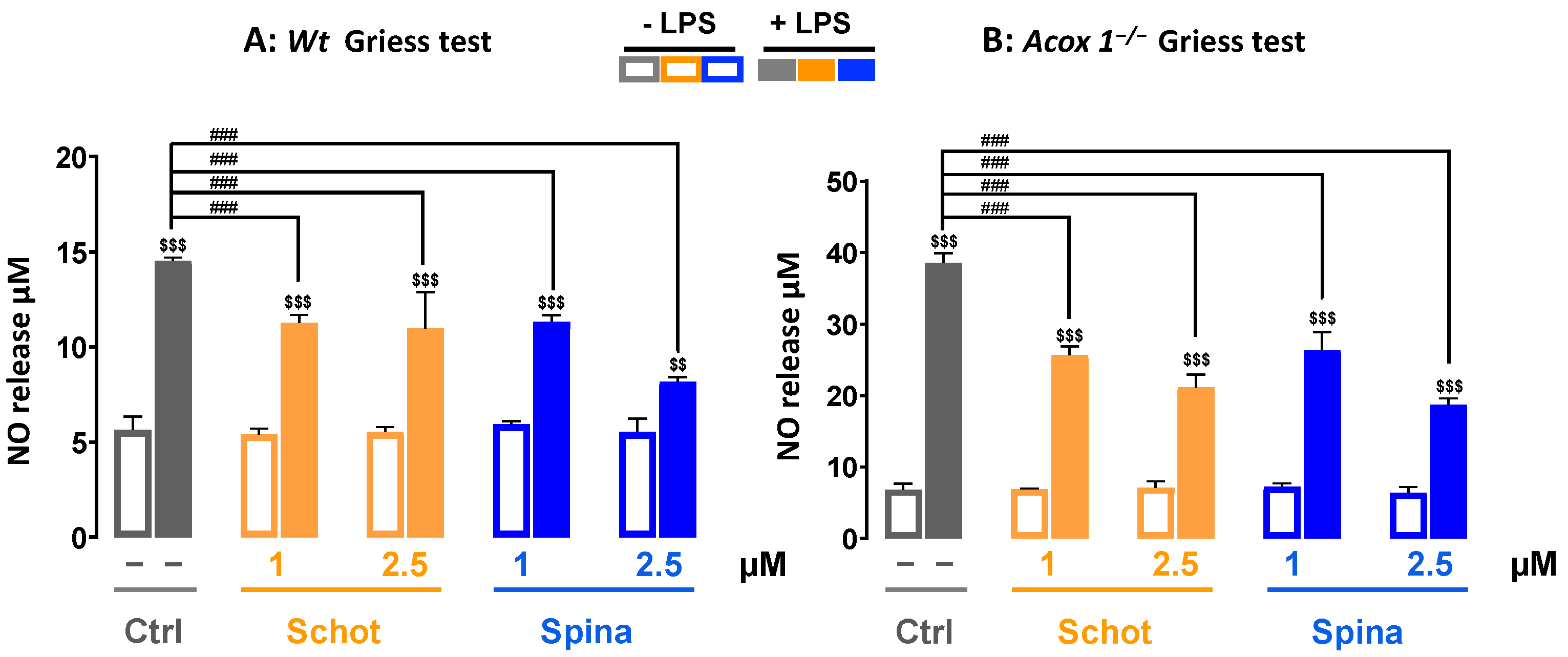
3.3. Schot and Spina Effects on LPS-Induced Nitric Oxide (NO) Generation
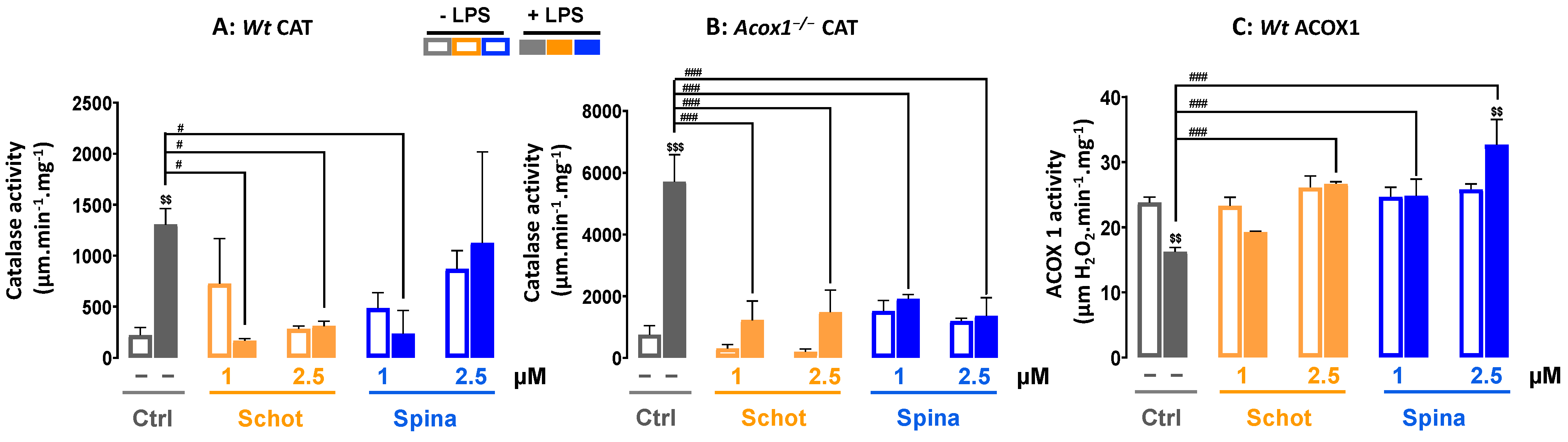
3.4. Effect of Schot and Spina on Peroxisomal Catalase and ACOX1 Activities
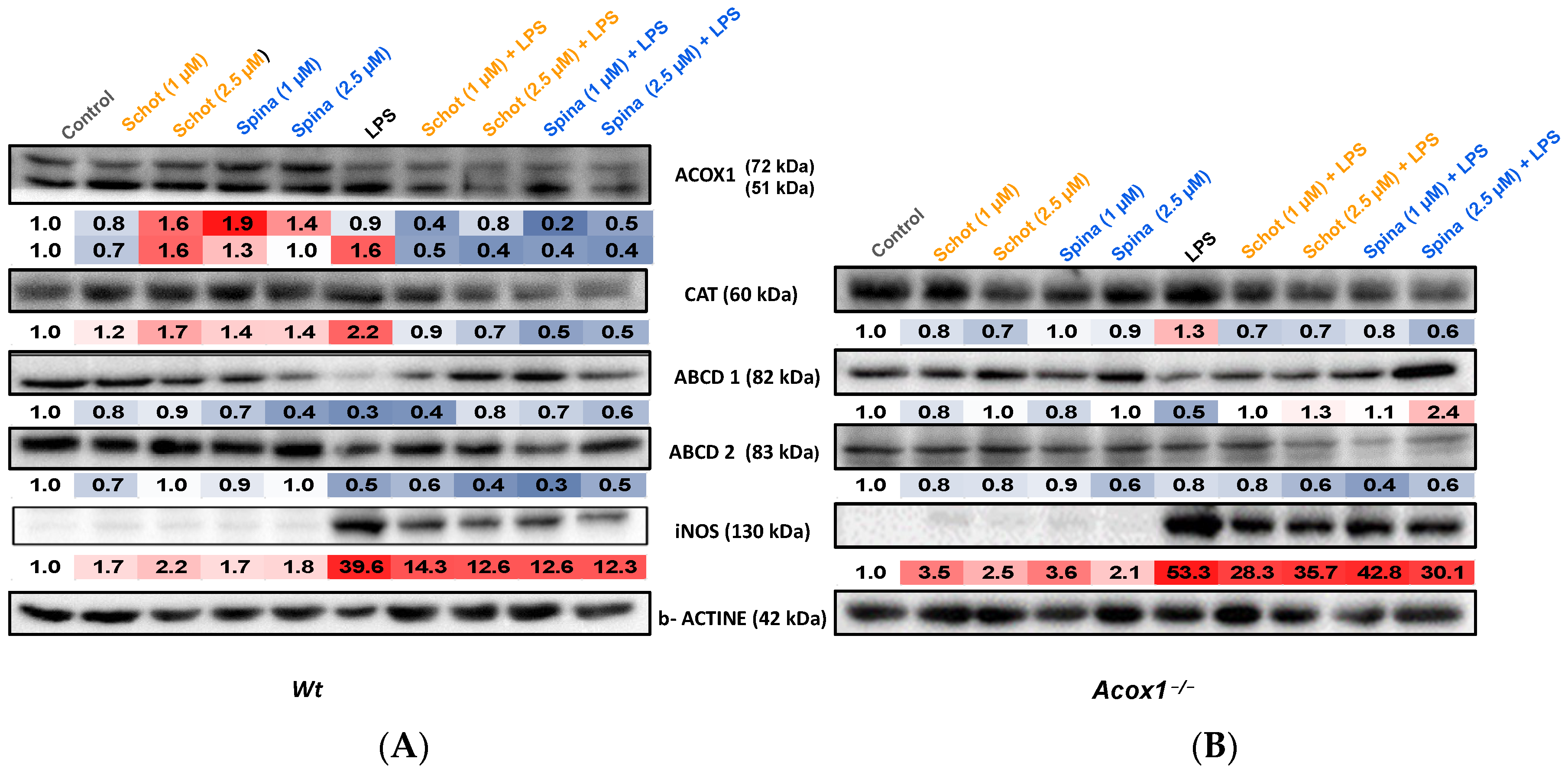
3.5. Effects of Schot and Spina on Peroxisomal Protein Expression
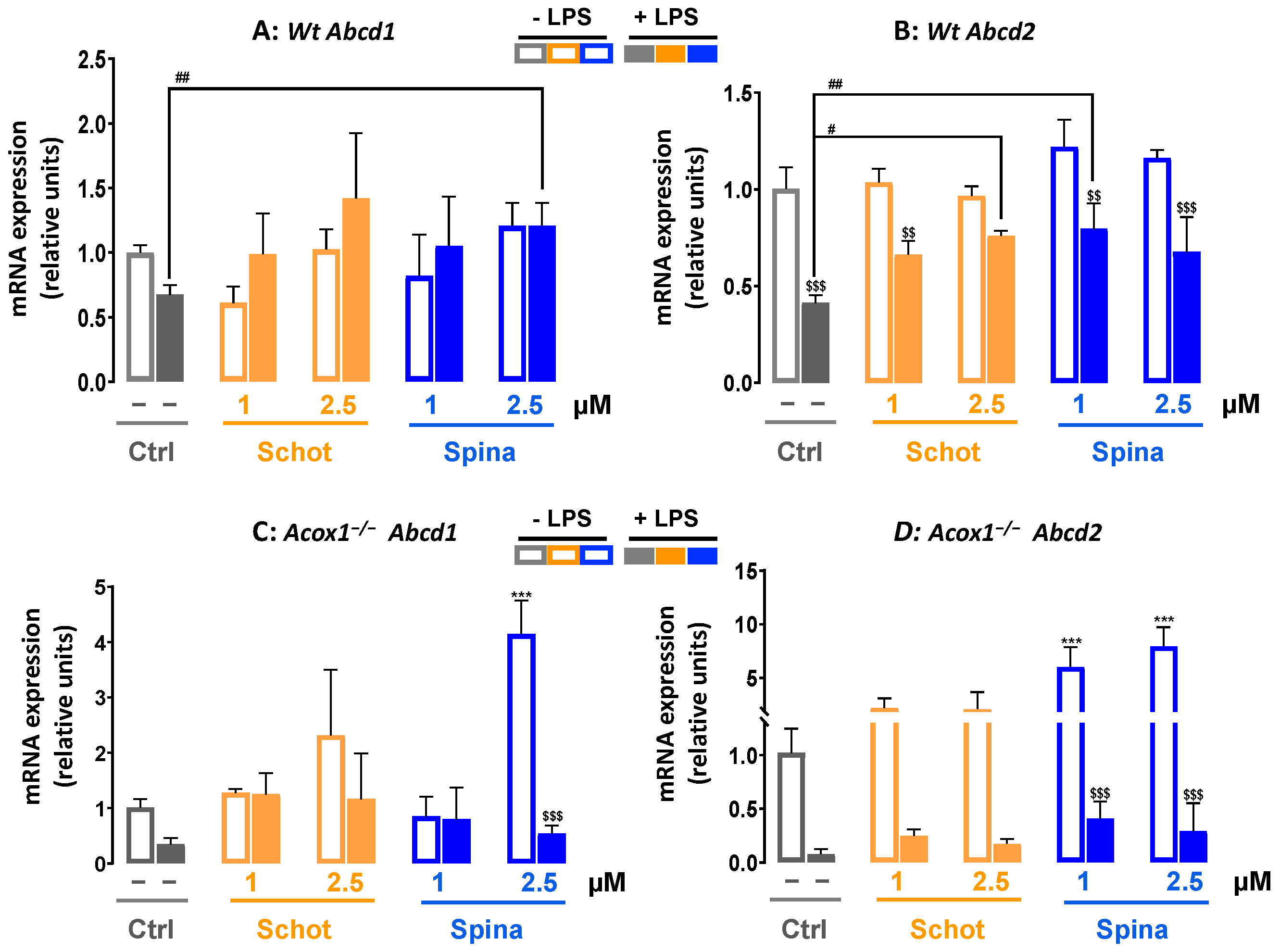
3.6. Shot and Spina Effect on the Expression of Peroxisomal Protein-Encoding Genes
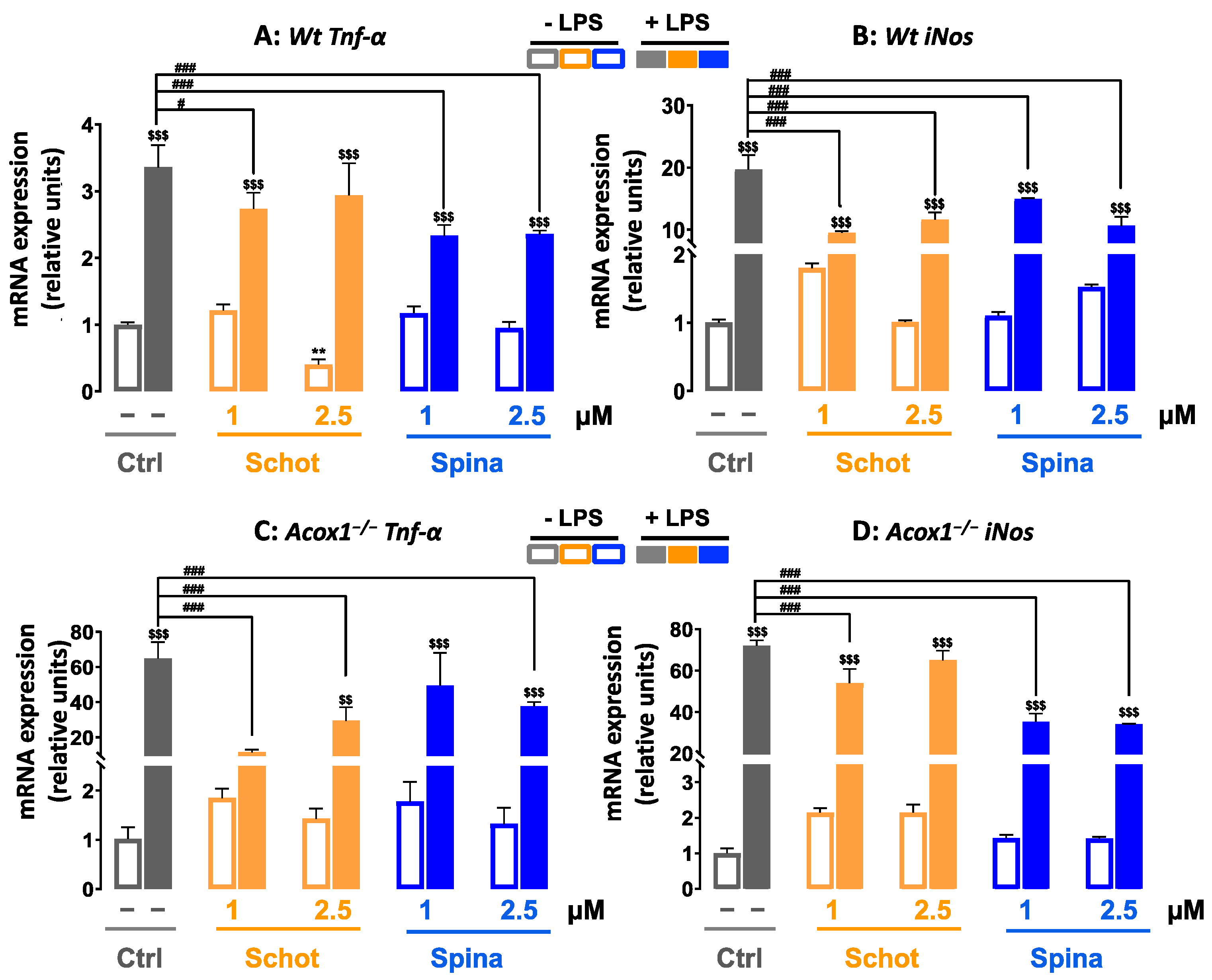
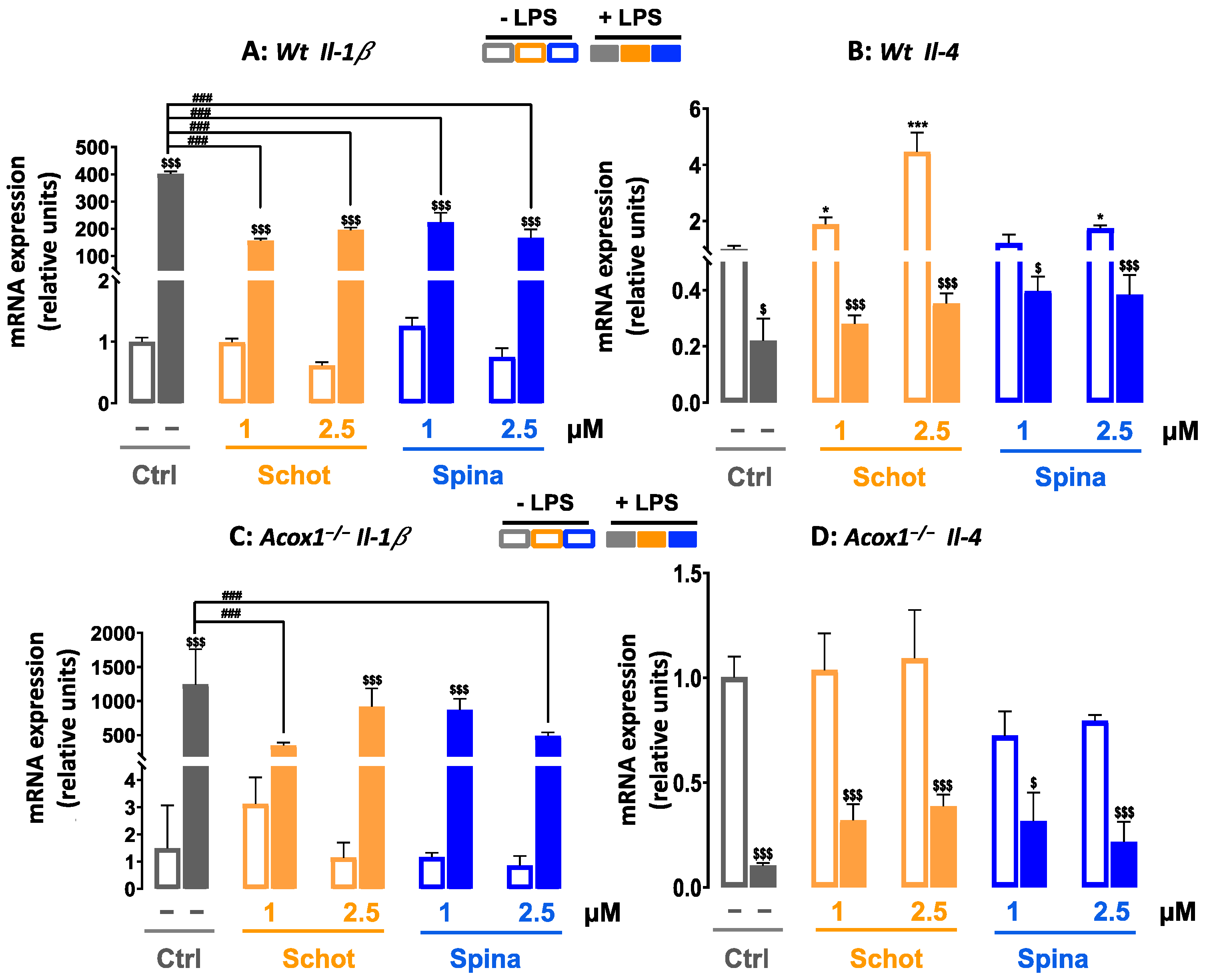
3.7. Effect of Schot and Spina on the Expression of Inflammation-Marker-Encoding Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salvi, J.; Andreoletti, P.; Audinat, E.; Balland, E.; Ben Fradj, S.; Cherkaoui-Malki, M.; Heurtaux, T.; Liénard, F.; Nédélec, E.; Rovère, C.; et al. Microgliosis: A Double-Edged Sword in the Control of Food Intake. FEBS J. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Javanmehr, N.; Saleki, K.; Alijanizadeh, P.; Rezaei, N. Microglia Dynamics in Aging-Related Neurobehavioral and Neuroinflammatory Diseases. J. Neuroinflamm. 2022, 19, 273. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.M.; DeCiero, D.P.; Ito, M.; Moser, A.B.; Moser, H.W. Adrenomyeloneuropathy: A Neuropathologic Review Featuring Its Noninflammatory Myelopathy. J. Neuropathol. Exp. Neurol. 2000, 59, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Von Bernhardi, R.; Eugenín-Von Bernhardi, L.; Eugenín, J. Microglial Cell Dysregulation in Brain Aging and Neurodegeneration. Front. Aging Neurosci. 2015, 7, 124. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C. Microglia and Neurodegeneration: The Role of Systemic Inflammation. Glia 2013, 61, 71–90. [Google Scholar] [CrossRef]
- Pawate, S.; Shen, Q.; Fan, F.; Bhat, N.R. Redox Regulation of Glial Inflammatory Response to Lipopolysaccharide and Interferongamma. J. Neurosci. Res. 2004, 77, 540–551. [Google Scholar] [CrossRef]
- Perry, V.H.; Teeling, J. Microglia and Macrophages of the Central Nervous System: The Contribution of Microglia Priming and Systemic Inflammation to Chronic Neurodegeneration. Semin. Immunopathol. 2013, 35, 601–612. [Google Scholar] [CrossRef]
- Konsman, J.P.; Parnet, P.; Dantzer, R. Cytokine-Induced Sickness Behaviour: Mechanisms and Implications. Trends Neurosci. 2002, 25, 154–159. [Google Scholar] [CrossRef]
- Vilhardt, F.; Haslund-Vinding, J.; Jaquet, V.; McBean, G. Microglia Antioxidant Systems and Redox Signalling. Br. J. Pharmacol. 2017, 174, 1719–1732. [Google Scholar] [CrossRef]
- Li, J.; Baud, O.; Vartanian, T.; Volpe, J.J.; Rosenberg, P.A. Peroxynitrite Generated by Inducible Nitric Oxide Synthase and NADPH Oxidase Mediates Microglial Toxicity to Oligodendrocytes. Proc. Natl. Acad. Sci. USA 2005, 102, 9936–9941. [Google Scholar] [CrossRef]
- Liu, B.; Du, L.; Hong, J.S. Naloxone Protects Rat Dopaminergic Neurons against Inflammatory Damage through Inhibition of Microglia Activation and Superoxide Generation. J. Pharmacol. Exp. Ther. 2000, 293, 607–617. [Google Scholar] [PubMed]
- Wang, T.; Liu, B.; Qin, L.; Wilson, B.; Hong, J.S. Protective Effect of the SOD/Catalase Mimetic MnTMPyP on Inflammation-Mediated Dopaminergic Neurodegeneration in Mesencephalic Neuronal-Glial Cultures. J. Neuroimmunol. 2004, 147, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Nell, H.J.; Au, J.L.; Giordano, C.R.; Terlecky, S.R.; Walton, P.A.; Whitehead, S.N.; Cechetto, D.F. Targeted Antioxidant, Catalase-SKL, Reduces Beta-Amyloid Toxicity in the Rat Brain. Brain Pathol. 2017, 27, 86–94. [Google Scholar] [CrossRef]
- Wanders, R.J.A. Peroxisomal Disorders: Improved Laboratory Diagnosis, New Defects and the Complicated Route to Treatment. Mol. Cell Probes 2018, 40, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, Y.; Abe, Y.; Imoto, Y.; Tanaka, A.J.; Okumoto, K.; Honsho, M.; Tamura, S.; Miyata, N.; Yamashita, T.; Chung, W.K.; et al. Recent Insights into Peroxisome Biogenesis and Associated Diseases. J. Cell Sci. 2020, 133, jcs236943. [Google Scholar] [CrossRef] [PubMed]
- Tahri-Joutey, M.; Andreoletti, P.; Surapureddi, S.; Nasser, B.; Cherkaoui-Malki, M.; Latruffe, N. Mechanisms Mediating the Regulation of Peroxisomal Fatty Acid Beta-Oxidation by PPARα. Int. J. Mol. Sci. 2021, 22, 8969. [Google Scholar] [CrossRef]
- Singh, I.; Pujol, A. Pathomechanisms Underlying X-Adrenoleukodystrophy: A Three-Hit Hypothesis. Brain Pathol. 2010, 20, 838–844. [Google Scholar] [CrossRef]
- Raas, Q.; Gondcaille, C.; Hamon, Y.; Leoni, V.; Caccia, C.; Menetrier, F.; Lizard, G.; Trompier, D.; Savary, S. CRISPR/Cas9-Mediated Knockout of Abcd1 and Abcd2 Genes in BV-2 Cells: Novel Microglial Models for X-Linked Adrenoleukodystrophy. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 704–714. [Google Scholar] [CrossRef]
- Raas, Q.; Saih, F.E.; Gondcaille, C.; Trompier, D.; Hamon, Y.; Leoni, V.; Caccia, C.; Nasser, B.; Jadot, M.; Menetrier, F.; et al. A Microglial Cell Model for Acyl-CoA Oxidase 1 Deficiency. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 567–576. [Google Scholar] [CrossRef]
- Essadek, S.; Bouchab, H.; El Kebbaj, R.; Gondcaille, C.; El Kamouni, S.; Savary, S.; Vamecq, J.; Essamadi, A.; Cherkaoui-Malki, M.; Nasser, B.; et al. Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-Oxidative Functions: Protective Action of Argan Oil. Pharmaceuticals 2022, 15, 465. [Google Scholar] [CrossRef]
- Badreddine, A.; Zarrouk, A.; Karym, E.M.; Debbabi, M.; Nury, T.; Meddeb, W.; Sghaier, R.; Bezine, M.; Vejux, A.; Martine, L.; et al. Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N. Int. J. Mol. Sci. 2017, 18, 2220. [Google Scholar] [CrossRef] [PubMed]
- El Kharrassi, Y.; Samadi, M.; Lopez, T.; Nury, T.; El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; Vamecq, J.; Moustaid, K.; Latruffe, N.; et al. Biological Activities of Schottenol and Spinasterol, Two Natural Phytosterols Present in Argan Oil and in Cactus Pear Seed Oil, on Murine Miroglial BV2 Cells. Biochem. Biophys. Res. Commun. 2014, 446, 798–804. [Google Scholar] [CrossRef]
- Lin, Y.; Knol, D.; Trautwein, E.A. Phytosterol Oxidation Products (POP) in Foods with Added Phytosterols and Estimation of Their Daily Intake: A Literature Review. Eur. J. Lipid Sci. Technol. 2016, 118, 1423–1438. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.; MacDougall, D.E.; Ntanios, F.; Vanstone, C.A. Dietary Phytosterols as Cholesterol-Lowering Agents in Humans. Can J. Physiol. Pharm. 1997, 75, 217–227. [Google Scholar] [CrossRef]
- El Kharrassi, Y.; Maata, N.; Mazri, M.A.; El Kamouni, S.; Talbi, M.; El Kebbaj, R.; Moustaid, K.; Essamadi, A.K.; Andreoletti, P.; El Mzouri, E.H.; et al. Chemical and Phytochemical Characterizations of Argan Oil (Argania spinosa L. Skeels), Olive Oil (Olea europaea L. Cv. Moroccan Picholine), Cactus Pear (Opuntia Megacantha Salm-Dyck) Seed Oil and Cactus Cladode Essential Oil. J. Food Meas. Charact. 2018, 12, 747–754. [Google Scholar] [CrossRef]
- Mukherjee, P.K. Chapter 20—Phyto-Pharmaceuticals, Nutraceuticals and Their Evaluation. In Quality Control and Evaluation of Herbal Drugs; Mukherjee, P.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 707–722. ISBN 978-0-12-813374-3. [Google Scholar]
- Yuan, L.; Zhang, F.; Shen, M.; Jia, S.; Xie, J. Phytosterols Suppress Phagocytosis and Inhibit Inflammatory Mediators via ERK Pathway on LPS-Triggered Inflammatory Responses in RAW264.7 Macrophages and the Correlation with Their Structure. Foods 2019, 8, 582. [Google Scholar] [CrossRef]
- Hayes, K.C.; Pronczuk, A.; Perlman, D. Nonesterified Phytosterols Dissolved and Recrystallized in Oil Reduce Plasma Cholesterol in Gerbils and Humans. J. Nutr. 2004, 134, 1395–1399. [Google Scholar] [CrossRef]
- Varady, K.A.; Houweling, A.H.; Jones, P.J.H. Effect of Plant Sterols and Exercise Training on Cholesterol Absorption and Synthesis in Previously Sedentary Hypercholesterolemic Subjects. Transl. Res. 2007, 149, 22–30. [Google Scholar] [CrossRef]
- Moghadasian, M.H. Pharmacological Properties of Plant Sterols in Vivo and in Vitro Observations. Life Sci. 2000, 67, 605–615. [Google Scholar] [CrossRef]
- Arisawa, M.; Kinghorn, A.D.; Cordell, G.A.; Phoebe, C.H.; Fansworth, N.R. Plant Anticancer Agents. XXXVI. Schottenol Glucoside from Baccharis Coridifolia and Ipomopsis Aggregata. Planta Med. 1985, 6, 544–545. [Google Scholar] [CrossRef]
- Berger, A.; Jones, P.J.H.; Abumweis, S.S. Plant Sterols: Factors Affecting Their Efficacy and Safety as Functional Food Ingredients. Lipids Health Dis. 2004, 3, 5. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Wen, Z.-H.; Wang, S.-K.; Hsu, C.-H.; Duh, C.-Y. New Anti-Inflammatory 4-Methylated Steroids from the Formosan Soft Coral Nephthea Chabroli. Steroids 2008, 73, 1181–1186. [Google Scholar] [CrossRef]
- Blasi, E.; Barluzzi, R.; Bocchini, V.; Mazzolla, R.; Bistoni, F. Immortalization of Murine Microglial Cells by a V-Raf/v-Myc Carrying Retrovirus. J. Neuroimmunol. 1990, 27, 229–237. [Google Scholar] [CrossRef]
- Badreddine, A.; Karym, E.M.; Zarrouk, A.; Nury, T.; El Kharrassi, Y.; Nasser, B.; Cherkaoui Malki, M.; Lizard, G.; Samadi, M. An Expeditious Synthesis of Spinasterol and Schottenol, Two Phytosterols Present in Argan Oil and in Cactus Pear Seed Oil, and Evaluation of Their Biological Activities on Cells of the Central Nervous System. Steroids 2015, 99, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of Nitrate, Nitrite, and [15N]Nitrate in Biological Fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Yazdani, M. Concerns in the Application of Fluorescent Probes DCDHF-DA, DHR 123 and DHE to Measure Reactive Oxygen Species in Vitro. Toxicol. In Vitro 2015, 30, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Sasaki, Y.; Tokuyama, S.; Sogabe, A.; Tahara, Y. Conversion of a Typical Catalase from Bacillus Sp. TE124 to a Catalase-Peroxidase by Directed Evolution. J. Biosci. Bioeng. 2002, 93, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Oaxaca-Castillo, D.; Andreoletti, P.; Vluggens, A.; Yu, S.; Van Veldhoven, P.P.; Reddy, J.K.; Cherkaoui-Malki, M. Biochemical Characterization of Two Functional Human Liver Acyl-CoA Oxidase Isoforms 1a and 1b Encoded by a Single Gene. Biochem. Biophys. Res. Commun. 2007, 360, 314–319. [Google Scholar] [CrossRef]
- Remans, T.; Keunen, E.; Bex, G.J.; Smeets, K.; Vangronsveld, J.; Cuypers, A. Reliable Gene Expression Analysis by Reverse Transcription-Quantitative PCR: Reporting and Minimizing the Uncertainty in Data Accuracy. Plant Cell 2014, 26, 3829–3837. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, L.; Zhang, H.; Shimoda, L.A.; DeBerardinis, R.J.; Semenza, G.L. Analysis of Hypoxia-Induced Metabolic Reprogramming. Methods Enzymol. 2014, 542, 425–455. [Google Scholar] [CrossRef]
- Laroux, F.S.; Pavlick, K.P.; Hines, I.N.; Kawachi, S.; Harada, H.; Bharwani, S.; Hoffman, J.M.; Grisham, M.B. Role of Nitric Oxide in Inflammation. Acta Physiol. Scand. 2001, 173, 113–118. [Google Scholar] [CrossRef]
- Graeber, M.B. Microglia. In Encyclopedia of the Neurological Sciences; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: New York, NY, USA, 2003; pp. 137–140. ISBN 978-0-12-226870-0. [Google Scholar]
- Saliba, S.W.; Jauch, H.; Gargouri, B.; Keil, A.; Hurrle, T.; Volz, N.; Mohr, F.; Van Der Stelt, M.; Bräse, S.; Fiebich, B.L. Anti-Neuroinflammatory Effects of GPR55 Antagonists in LPS-Activated Primary Microglial Cells. J. Neuroinflamm. 2018, 15, 322. [Google Scholar] [CrossRef] [PubMed]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-Mediated Neuroinflammation in Neurodegenerative Diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Yammine, A.; Ghzaiel, I.; Sassi, K.; Zarrouk, A.; Brahmi, F.; Samadi, M.; Rup-Jacques, S.; Vervandier-Fasseur, D.; Pais de Barros, J.P.; et al. Attenuation of 7-Ketocholesterol- and 7β-Hydroxycholesterol-Induced Oxiapoptophagy by Nutrients, Synthetic Molecules and Oils: Potential for the Prevention of Age-Related Diseases. Ageing Res. Rev. 2021, 68, 101324. [Google Scholar] [CrossRef]
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of Reactive Oxygen Species Generating and Scavenging System in Salt Sensitive and Salt Tolerant Cultivars of Rice. BMC Plant Biol. 2016, 16, 131. [Google Scholar] [CrossRef]
- Jelinek, M.; Jurajda, M.; Duris, K. Oxidative Stress in the Brain: Basic Concepts and Treatment Strategies in Stroke. Antioxidants 2021, 10, 1886. [Google Scholar] [CrossRef]
- More, G.K.; Makola, R.T. In-Vitro Analysis of Free Radical Scavenging Activities and Suppression of LPS-Induced ROS Production in Macrophage Cells by Solanum Sisymbriifolium Extracts. Sci. Rep. 2020, 10, 6493. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Chen, W.; Hu, R.; Feng, H. The Injury and Therapy of Reactive Oxygen Species in Intracerebral Hemorrhage Looking at Mitochondria. Oxid. Med. Cell Longev. 2016, 2016, 2592935. [Google Scholar] [CrossRef]
- Andersen, J.K. Oxidative Stress in Neurodegeneration: Cause or Consequence? Nat. Med. 2004, 10, S18–S25. [Google Scholar] [CrossRef]
- Cenini, G.; Lloret, A.; Cascella, R. Oxidative Stress in Neurodegenerative Diseases: From a Mitochondrial Point of View. Oxid. Med. Cell Longev. 2019, 2019, 2105607. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative Stress and Neurodegeneration: Where Are We Now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Niki, E. Antioxidant Effects of Phytosterol and Its Components. J. Nutr. Sci. Vitaminol. 2003, 49, 277–280. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [PubMed]
- Rafikov, R.; Kumar, S.; Aggarwal, S.; Hou, Y.; Kangath, A.; Pardo, D.; Fineman, J.R.; Black, S.M. Endothelin-1 Stimulates Catalase Activity through the PKCδ-Mediated Phosphorylation of Serine 167. Free Radic. Biol. Med. 2014, 67, 255–264. [Google Scholar] [CrossRef]
- Cao, C.; Leng, Y.; Kufe, D. Catalase Activity Is Regulated by C-Abl and Arg in the Oxidative Stress Response. J. Biol. Chem. 2003, 278, 29667–29675. [Google Scholar] [CrossRef]
- Baskar, A.A.; Al Numair, K.S.; Gabriel Paulraj, M.; Alsaif, M.A.; Muamar, M.A.; Ignacimuthu, S. β-Sitosterol Prevents Lipid Peroxidation and Improves Antioxidant Status and Histoarchitecture in Rats with 1,2-Dimethylhydrazine-Induced Colon Cancer. J. Med. Food 2012, 15, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, M.; Moreno, J.J. Beta-Sitosterol Modulates Antioxidant Enzyme Response in RAW 264.7 Macrophages. Free Radic. Biol. Med. 2005, 39, 91–97. [Google Scholar] [CrossRef]
- Lefebvre, P.; Chinetti, G.; Fruchart, J.-C.; Staels, B. Sorting out the Roles of PPAR Alpha in Energy Metabolism and Vascular Homeostasis. J. Clin. Invest. 2006, 116, 571–580. [Google Scholar] [CrossRef]
- Zhang, N.; Chu, E.S.H.; Zhang, J.; Li, X.; Liang, Q.; Chen, J.; Chen, M.; Teoh, N.; Farrell, G.; Sung, J.J.Y.; et al. Peroxisome Proliferator Activated Receptor Alpha Inhibits Hepatocarcinogenesis through Mediating NF-ΚB Signaling Pathway. Oncotarget 2014, 5, 8330–8340. [Google Scholar] [CrossRef]
- Shi, X.; Chen, Y.; Nadeem, L.; Xu, G. Beneficial Effect of TNF-α Inhibition on Diabetic Peripheral Neuropathy. J. Neuroinflamm. 2013, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Zelcer, N.; Tontonoz, P. Liver X Receptors as Integrators of Metabolic and Inflammatory Signaling. J. Clin. Investig. 2006, 116, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Di Cara, F.; Savary, S.; Kovacs, W.J.; Kim, P.; Rachubinski, R.A. The Peroxisome: An up-and-Coming Organelle in Immunometabolism. Trends Cell Biol. 2022, 33, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Jo, D.S.; Cho, D.-H. Peroxisomal Dysfunction in Neurodegenerative Diseases. Arch. Pharm. Res. 2019, 42, 393–406. [Google Scholar] [CrossRef]
- Lizard, G.; Rouaud, O.; Demarquoy, J.; Cherkaoui-Malki, M.; Iuliano, L. Potential Roles of Peroxisomes in Alzheimer’s Disease and in Dementia of the Alzheimer’s Type. J. Alzheimer’s Dis. 2012, 29, 241–254. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Wu, J.-S.; Tsai, H.-D.; Huang, C.-Y.; Chen, J.-J.; Sun, G.Y.; Lin, T.-N. Peroxisome Proliferator-Activated Receptor Gamma (PPAR-γ) and Neurodegenerative Disorders. Mol. Neurobiol. 2012, 46, 114–124. [Google Scholar] [CrossRef]
- Trompier, D.; Vejux, A.; Zarrouk, A.; Gondcaille, C.; Geillon, F.; Nury, T.; Savary, S.; Lizard, G. Brain Peroxisomes. Biochimie 2014, 98, 102–110. [Google Scholar] [CrossRef]
- Ferdinandusse, S.; Denis, S.; Hogenhout, E.M.; Koster, J.; Van Roermund, C.W.; IJlst, L.; Moser, A.B.; Wanders, R.J.; Waterham, H.R. Clinical, Biochemical, and Mutational Spectrum of Peroxisomal Acyl-Coenzyme A Oxidase Deficiency. Hum. Mutat. 2007, 28, 904–912. [Google Scholar] [CrossRef]
- Griffin, E.N.; Ackerman, S.L. Lipid Metabolism and Axon Degeneration: An ACOX1 Balancing Act. Neuron 2020, 106, 551–553. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Bronfman, M.; Leighton, F. Detection of Peroxisomal Fatty Acyl-Coenzyme A Oxidase Activity. Biochem. J. 1979, 182, 779–788. [Google Scholar] [CrossRef]
- El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; El Kharrassi, Y.; Vamecq, J.; Mandard, S.; Saih, F.E.; Latruffe, N.; El Kebbaj, M.S.; Lizard, G.; et al. Argan Oil Prevents Down-Regulation Induced by Endotoxin on Liver Fatty Acid Oxidation and Gluconeogenesis and on Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1α, (PGC-1α), Peroxisome Proliferator-Activated Receptor α (PPARα) and Estrogen r. Biochimie Open 2015, 1, 51–59. [Google Scholar] [CrossRef]
- Roy, B.; Jacobson, A. The Intimate Relationships of MRNA Decay and Translation. Trends Genet. 2013, 29, 691–699. [Google Scholar] [CrossRef]
- Guan, B.-J.; Krokowski, D.; Majumder, M.; Schmotzer, C.L.; Kimball, S.R.; Merrick, W.C.; Koromilas, A.E.; Hatzoglou, M. Translational Control during Endoplasmic Reticulum Stress beyond Phosphorylation of the Translation Initiation Factor EIF2α. J. Biol. Chem. 2014, 289, 12593–12611. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, W.; Wang, M.; Zhu, J.; Xia, L. TLR4-SIRT3 Mechanism Modulates Mitochondrial and Redox Homeostasis and Promotes EPCs Recruitment and Survival. Oxid. Med. Cell Longev. 2022, 2022, 1282362. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-F.; Zheng, H.; Luo, Z.; Hogstrand, C.; Bai, Z.-Y.; Wei, X.-L. Dietary Choline Alleviates High-Fat Diet-Induced Hepatic Lipid Dysregulation via UPRmt Modulated by SIRT3-Mediated MtHSP70 Deacetylation. Int. J. Mol. Sci. 2022, 23, 4204. [Google Scholar] [CrossRef] [PubMed]
- Weinhofer, I.; Kunze, M.; Rampler, H.; Bookout, A.L.; Forss-Petter, S.; Berger, J. Liver X Receptor Alpha Interferes with SREBP1c-Mediated Abcd2 Expression. Novel Cross-Talk in Gene Regulation. J. Biol. Chem. 2005, 280, 41243–41251. [Google Scholar] [CrossRef]
- Kemp, S.; Wei, H.M.; Lu, J.F.; Braiterman, L.T.; McGuinness, M.C.; Moser, A.B.; Watkins, P.A.; Smith, K.D. Gene Redundancy and Pharmacological Gene Therapy: Implications for X-Linked Adrenoleukodystrophy. Nat. Med. 1998, 4, 1261–1268. [Google Scholar] [CrossRef]
- Netik, A.; Forss-Petter, S.; Holzinger, A.; Molzer, B.; Unterrainer, G.; Berger, J. Adrenoleukodystrophy-Related Protein Can Compensate Functionally for Adrenoleukodystrophy Protein Deficiency (X-ALD): Implications for Therapy. Hum. Mol. Genet. 1999, 8, 907–913. [Google Scholar] [CrossRef]
- Tawbeh, A.; Gondcaille, C.; Trompier, D. Peroxisomal ABC Transporters: An Update. Int. J. Mol. Sci. 2021, 22, 6093. [Google Scholar] [CrossRef]
- Trompier, D.; Gondcaille, C.; Lizard, G.; Savary, S. Regulation of the Adrenoleukodystrophy-Related Gene (ABCD2): Focus on Oxysterols and LXR Antagonists. Biochem. Biophys. Res. Commun. 2014, 446, 651–655. [Google Scholar] [CrossRef]
- Nomaguchi, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Toida, T.; Iwatsuki, K.; Goto, T.; Kawada, T. Aloe Vera Phytosterols Act as Ligands for PPAR and Improve the Expression Levels of PPAR Target Genes in the Livers of Mice with Diet-Induced Obesity. Obes. Res. Clin. Pract. 2011, 5, e190–e201. [Google Scholar] [CrossRef]
- Contreras, A.V.; Torres, N.; Tovar, A.R. PPAR-α as a Key Nutritional and Environmental Sensor for Metabolic Adaptation. Adv. Nutr. 2013, 4, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Fantacuzzi, M.; De Filippis, B.; Amoroso, R.; Giampietro, L. PPAR Ligands Containing Stilbene Scaffold. Mini Rev. Med. Chem. 2019, 19, 1599–1610. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Konno, R.; Shimizu, T.; Ide, T.; Takahashi, N.; Kawada, T.; Nagao, K.; Inoue, N.; Yanagita, T.; Hamada, T.; et al. Campest-5-En-3-One, an Oxidized Derivative of Campesterol, Activates PPARalpha, Promotes Energy Consumption and Reduces Visceral Fat Deposition in Rats. Biochim. Biophys. Acta 2006, 1760, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, J.-Y.; Kwon, H.C.; Kwon, J.; Jang, D.S.; Kang, K.S. Dual Beneficial Effects of α-Spinasterol Isolated from Aster Pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells. Plants 2022, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; He, L.; Zhong, Z.; Zhao, R.; Weng, S.; Mi, H.; Liu, F. Stigmasterol Restores the Balance of Treg/Th17 Cells by Activating the Butyrate-PPARγ Axis in Colitis. Front Immunol. 2021, 12, 741934. [Google Scholar] [CrossRef]
- Wilms, H.; Sievers, J.; Rickert, U.; Rostami-Yazdi, M.; Mrowietz, U.; Lucius, R. Dimethylfumarate Inhibits Microglial and Astrocytic Inflammation by Suppressing the Synthesis of Nitric Oxide, IL-1β, TNF-α and IL-6 in an in-Vitro Model of Brain Inflammation. J. Neuroinflamm. 2010, 7, 1–8. [Google Scholar] [CrossRef]
- Hoogland, I.C.M.; Houbolt, C.; Van Westerloo, D.J.; Van Gool, W.A.; Van de Beek, D. Systemic Inflammation and Microglial Activation: Systematic Review of Animal Experiments. J. Neuroinflamm. 2015, 12, 1–13. [Google Scholar] [CrossRef]
- Valerio, M.S.; Minderman, H.; Mace, T.; Awad, A.B. β-Sitosterol Modulates TLR4 Receptor Expression and Intracellular MyD88-Dependent Pathway Activation in J774A.1 Murine Macrophages. Cell Immunol. 2013, 285, 76–83. [Google Scholar] [CrossRef]
- Kurano, M.; Hasegawa, K.; Kunimi, M.; Hara, M.; Yatomi, Y.; Teramoto, T.; Tsukamoto, K. Sitosterol Prevents Obesity-Related Chronic Inflammation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 191–198. [Google Scholar] [CrossRef]
- Walsh, J.T.; Hendrix, S.; Boato, F.; Smirnov, I.; Zheng, J.; Lukens, J.R.; Gadani, S.; Hechler, D.; Gölz, G.; Rosenberger, K.; et al. MHCII-Independent CD4+ T Cells Protect Injured CNS Neurons via IL-4. J. Clin. Investig. 2015, 125, 2547. [Google Scholar] [CrossRef] [PubMed]
- Hühner, L.; Rilka, J.; Gilsbach, R.; Zhou, X.; Machado, V.; Spittau, B. Interleukin-4 Protects Dopaminergic Neurons In Vitro but Is Dispensable for MPTP-Induced Neurodegeneration In Vivo. Front Mol. Neurosci. 2017, 10, 62. [Google Scholar] [CrossRef]
- Park, K.W.; Lee, D.Y.; Joe, E.H.; Kim, S.U.; Jin, B.K. Neuroprotective Role of Microglia Expressing Interleukin-4. J. Neurosci. Res. 2005, 81, 397–402. [Google Scholar] [CrossRef]
- Casella, G.; Garzetti, L.; Gatta, A.T.; Finardi, A.; Maiorino, C.; Ruffini, F.; Martino, G.; Muzio, L.; Furlan, R. IL4 Induces IL6-Producing M2 Macrophages Associated to Inhibition of Neuroinflammation in Vitro and in Vivo. J. Neuroinflamm. 2016, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Spittau, B. Interleukin 4-Induced Neuroprotection and Regulation of Microglia Activation as a Therapeutic Approach in the MPTP Model of Parkinson’s Disease. Neural Regen. Res. 2017, 12, 1433–1434. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Primer Sequences |
|---|---|---|
| Abcd1-F Abcd1-R | NM_007435.2 | 5′GCCAAGTTGTGGATGTGGAG3′ 5′TTCCGCAGAGTCGGGATAGA3′ |
| Abcd2-F Abcd2-R | NM_011994.4 | 5′TAGAACGCATCCTGCACAGC3′ 5′CTCCTTCGCCATCGAATTGT3′ |
| Acox1-F Acox1-R | NM_001377522.1 | 5′TCGAAGCCAGCGTTACGAG3′ 5′GGTCTGCGATGCCAAATTCC3′ |
| Cat-F Cat-R | NM_009804.2 | 5′AGCGACCAGATGAAGCAGTG3′ 5′TCCGCTCTCTGTCAAAGTGTG3′ |
| Il-1β-F Il-1β-R | NM_008361.4 | 5′GAGATTGAGCTGTCTGCTCA 3′ 5′AAGGAGAACCAAGCAACGAC 3′ |
| Il-4-F IL-4-R | NM_021283.2 | 5′CCATATCCACGGATGCGACAA3′ 5′CCTCGTTCAAAATGCCGATGAT3′ |
| iNos-F iNos-R | NM_010927.4 | 5′CCTAGTCAACTGCAAGAGAA3′ 5′TTTCAGGTCACTTTGGTAGG3′ |
| Tnf-α-F Tnf-α-R | NM_013693.3 | 5′CCCTCACACTCAGATCATCTTCT3′ 5′GCTACGACGTGGGCTACAG3′ |
| 36b4-F 36b4-R | NM_007475.5 | 5′CGACCTGGAAGTCCAACTAC3′ 5′ATCTGCTGCATCTGCTTG3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essadek, S.; Gondcaille, C.; Savary, S.; Samadi, M.; Vamecq, J.; Lizard, G.; El Kebbaj, R.; Latruffe, N.; Benani, A.; Nasser, B.; et al. Two Argan Oil Phytosterols, Schottenol and Spinasterol, Attenuate Oxidative Stress and Restore LPS-Dysregulated Peroxisomal Functions in Acox1−/− and Wild-Type BV-2 Microglial Cells. Antioxidants 2023, 12, 168. https://doi.org/10.3390/antiox12010168
Essadek S, Gondcaille C, Savary S, Samadi M, Vamecq J, Lizard G, El Kebbaj R, Latruffe N, Benani A, Nasser B, et al. Two Argan Oil Phytosterols, Schottenol and Spinasterol, Attenuate Oxidative Stress and Restore LPS-Dysregulated Peroxisomal Functions in Acox1−/− and Wild-Type BV-2 Microglial Cells. Antioxidants. 2023; 12(1):168. https://doi.org/10.3390/antiox12010168
Chicago/Turabian StyleEssadek, Soukaina, Catherine Gondcaille, Stéphane Savary, Mohammad Samadi, Joseph Vamecq, Gérard Lizard, Riad El Kebbaj, Norbert Latruffe, Alexandre Benani, Boubker Nasser, and et al. 2023. "Two Argan Oil Phytosterols, Schottenol and Spinasterol, Attenuate Oxidative Stress and Restore LPS-Dysregulated Peroxisomal Functions in Acox1−/− and Wild-Type BV-2 Microglial Cells" Antioxidants 12, no. 1: 168. https://doi.org/10.3390/antiox12010168