
Exploration of Patient-Derived Pancreatic Ductal Adenocarcinoma Ex Vivo Tissue for Treatment Response

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Analysis of Patient-Derived Samples
2.2. RNA Extraction and Transcriptome Library Preparation
2.3. Illumina Sequencing and Gene Expression Analysis
2.4. PDAC Cell Line Culturing Condition
2.5. Cell Viability Assay
2.6. RNA Extraction and Two-Step Real-Time PCR
2.7. Statistical Evaluation
3. Results
3.1. Effect of IPA on Ex Vivo Tissue Culture
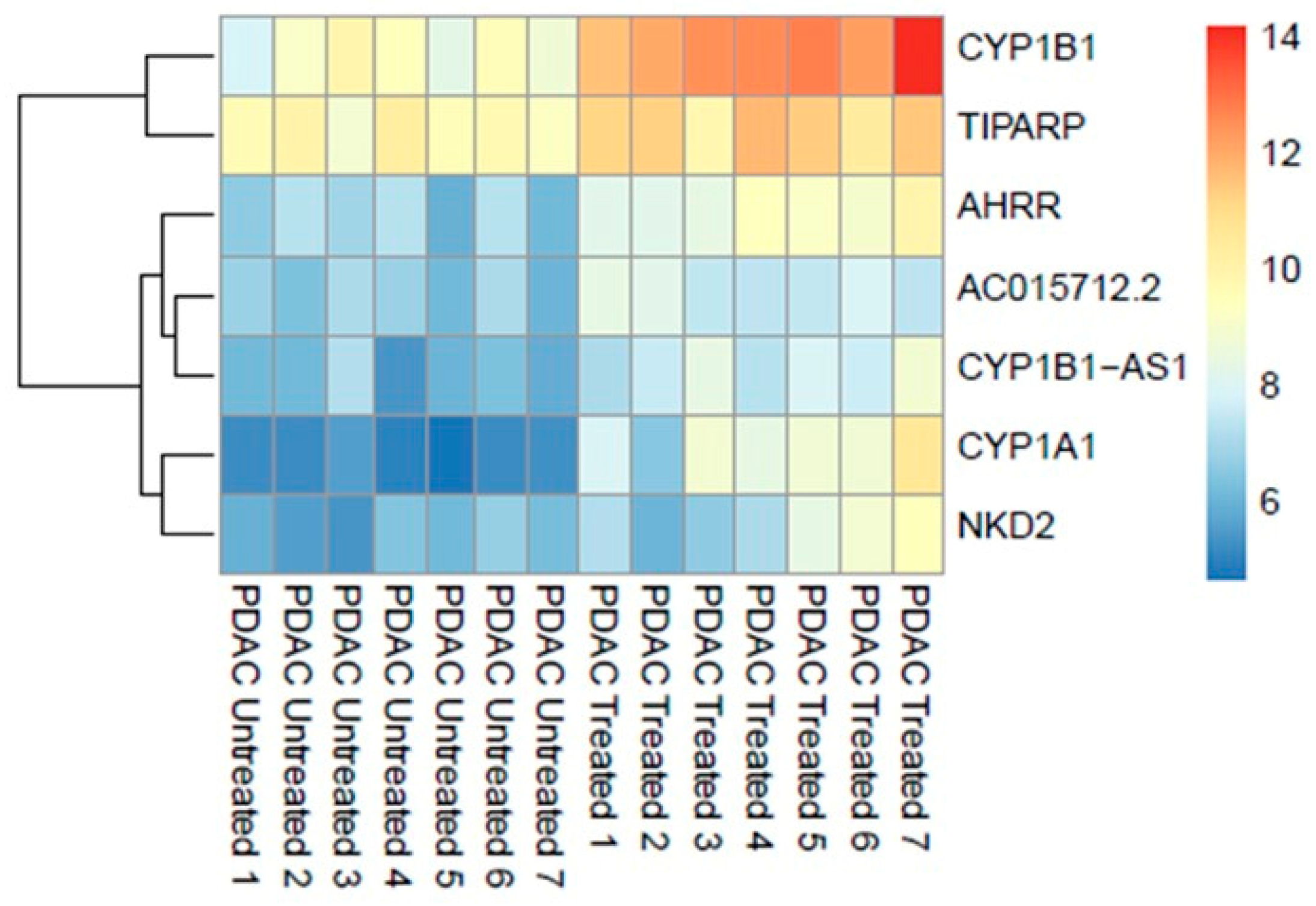
3.1.1. RNA Expression
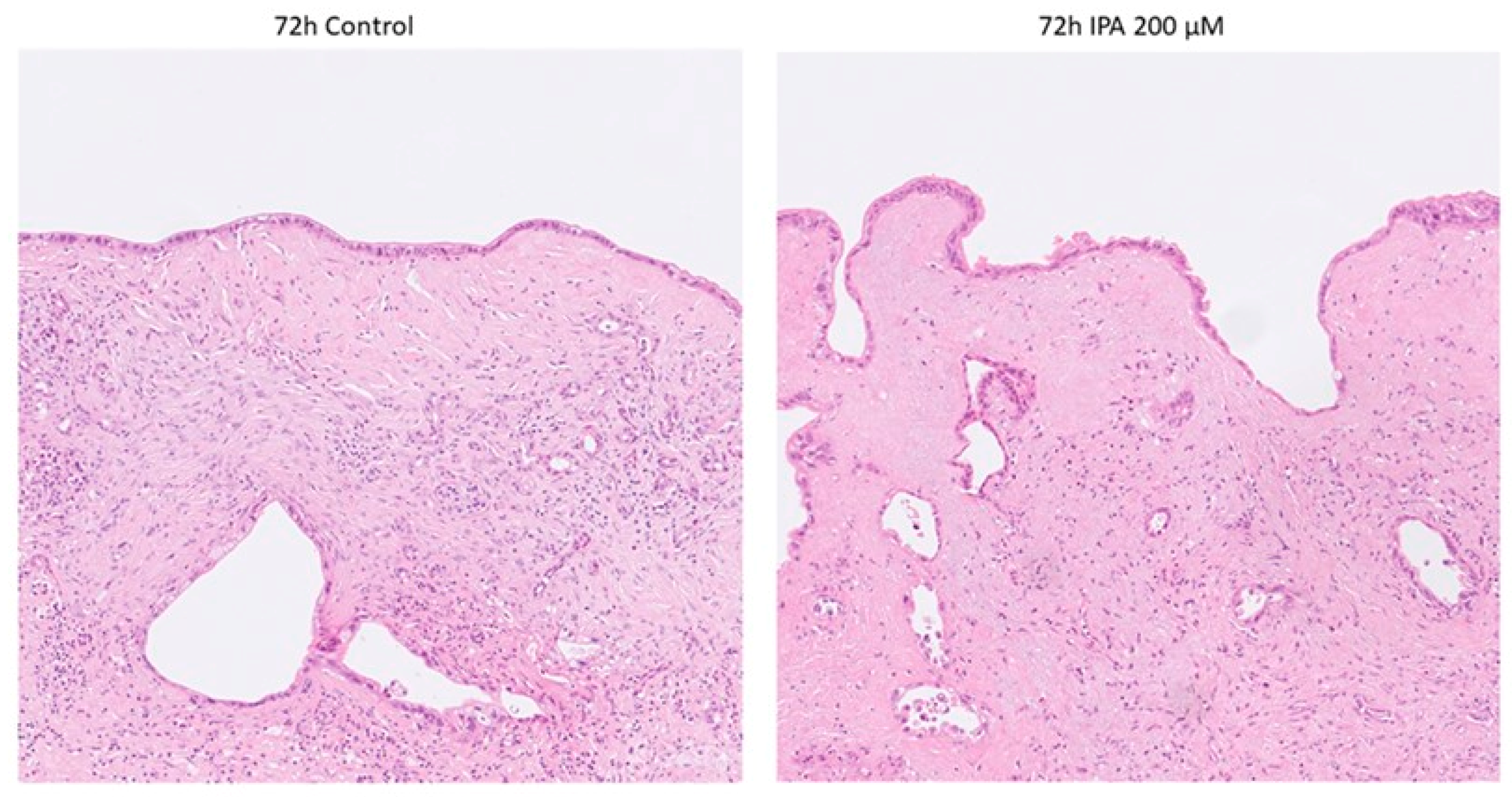
3.1.2. Pathological Examination of FFPE Tissue
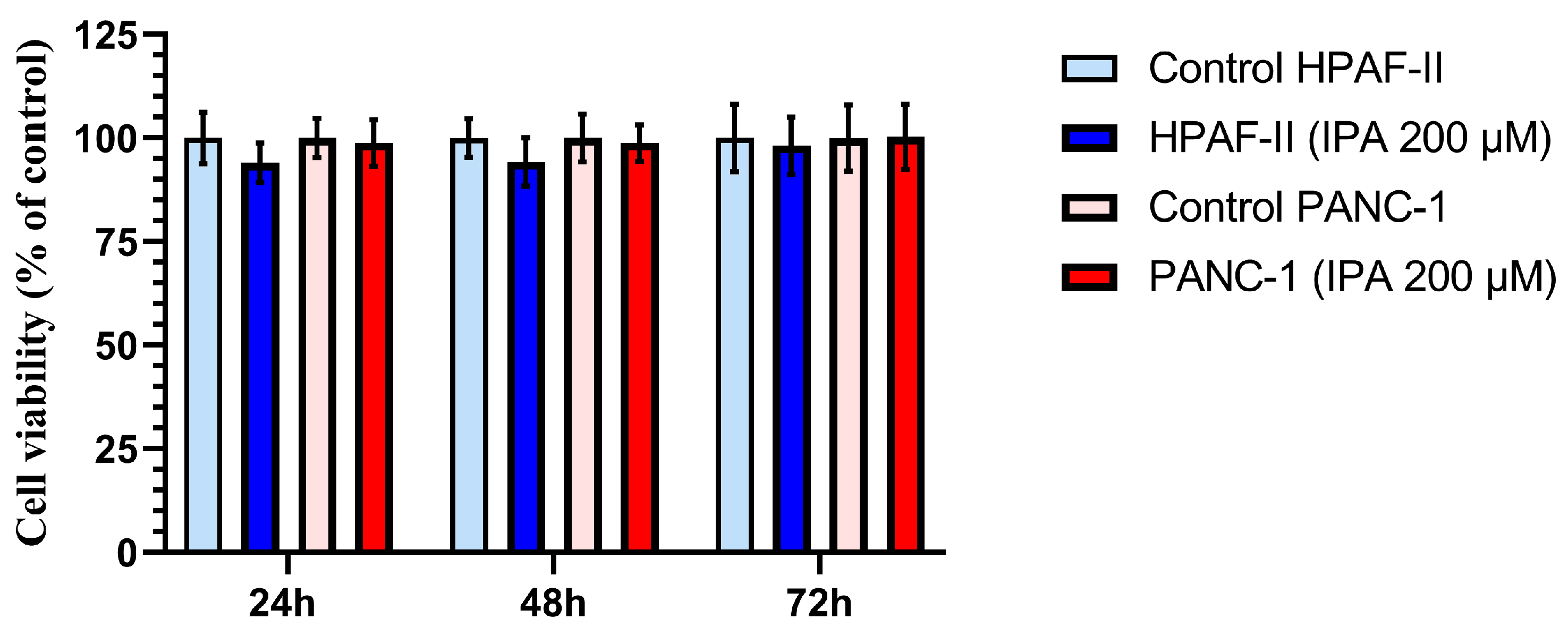
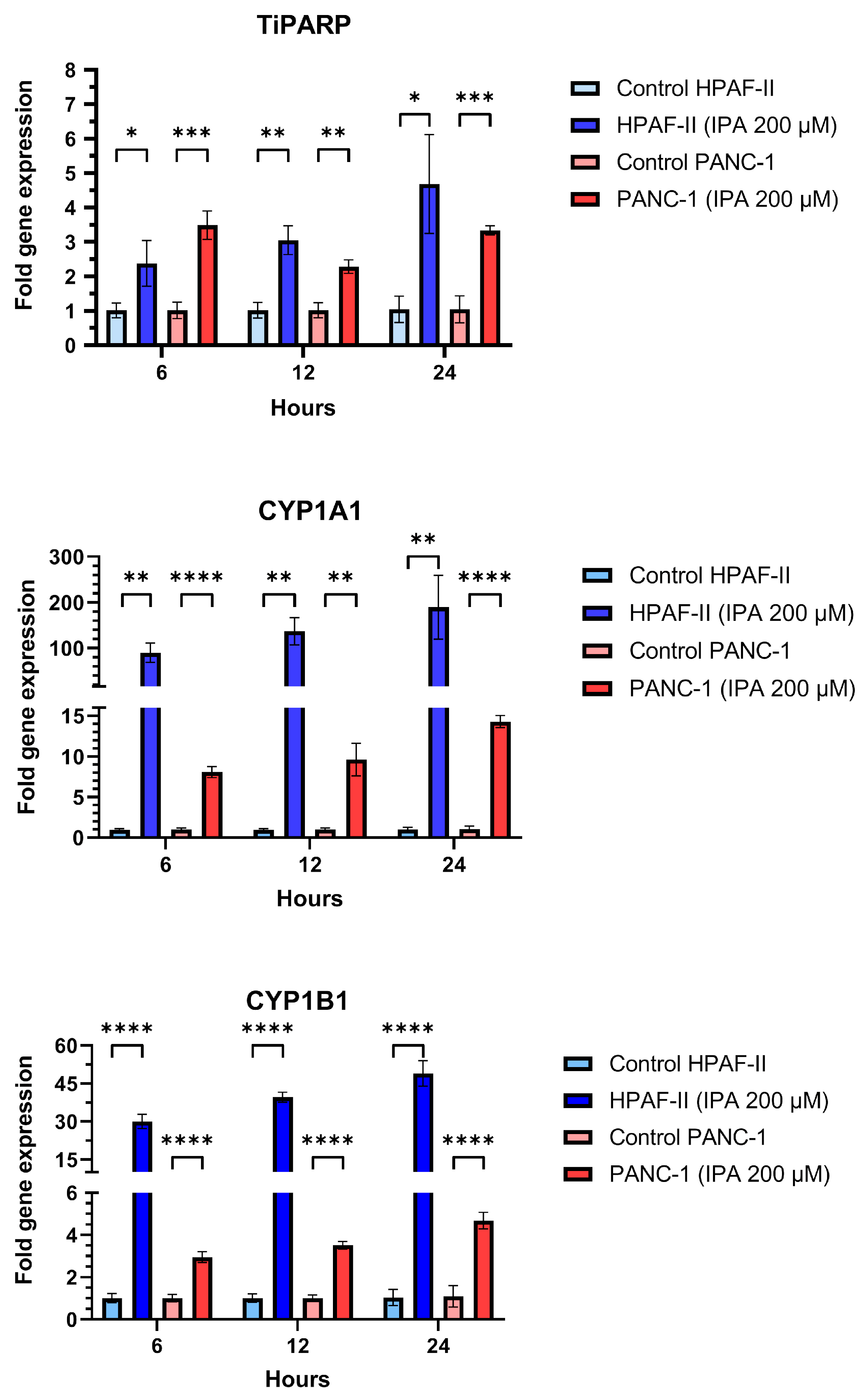
3.2. Effect of IPA on Pancreatic Cell Lines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ushio, J.; Kanno, A.; Ikeda, E.; Ando, K.; Nagai, H.; Miwata, T.; Kawasaki, Y.; Tada, Y.; Yokoyama, K.; Numao, N.; et al. Pancreatic Ductal Adenocarcinoma: Epidemiology and Risk Factors. Diagnostics 2021, 11, 562. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principe, D.R.; Underwood, P.W.; Korc, M.; Trevino, J.G.; Munshi, H.G.; Rana, A. The Current Treatment Paradigm for Pancreatic Ductal Adenocarcinoma and Barriers to Therapeutic Efficacy. Front. Oncol. 2021, 11, 2773. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Seo, Y.D.; Chang, J.H.; Coveler, A.; Nigjeh, E.N.; Pan, S.; Jalikis, F.; Yeung, R.S.; Crispe, I.N.; Pillarisetty, V.G. Long-lived pancreatic ductal adenocarcinoma slice cultures enable precise study of the immune microenvironment. Oncoimmunology 2017, 6, e1333210. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Wei, X.; Xu, X. Patient-derived xenograft models in hepatopancreatobiliary cancer. Cancer Cell Int. 2022, 22, 41. [Google Scholar] [CrossRef] [PubMed]
- Närhi, K.; Nagaraj, A.S.; Parri, E.; Turkki, R.; van Duijn, P.W.; Hemmes, A.; Lahtela, J.; Uotinen, V.; Mäyränpää, M.I.; Salmenkivi, K.; et al. Spatial aspects of oncogenic signalling determine the response to combination therapy in slice explants from Kras-driven lung tumours. J. Pathol. 2018, 245, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Suri, R.; Zimmerman, J.W.; Burkhart, R.A. Modeling human pancreatic ductal adenocarcinoma for translational research: Current options, challenges, and prospective directions. Ann. Pancreat. Cancer 2020, 3, 17. [Google Scholar] [CrossRef]
- Bresnahan, E.; Ramadori, P.; Heikenwalder, M.; Zender, L.; Lujambio, A. Novel patient-derived preclinical models of liver cancer. J. Hepatol. 2020, 72, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Templeton, A.R.; Jeffery, P.L.; Thomas, P.B.; Perera, M.P.J.; Ng, G.; Calabrese, A.R.; Nicholls, C.; Mackenzie, N.J.; Wood, J.; Bray, L.J.; et al. Patient-Derived Explants as a Precision Medicine Patient-Proximal Testing Platform Informing Cancer Management. Front. Oncol. 2021, 11, 767697. [Google Scholar] [CrossRef]
- Hynds, R.E.; Frese, K.K.; Pearce, D.R.; Grönroos, E.; Dive, C.; Swanton, C. Progress towards non-small-cell lung cancer models that represent clinical evolutionary trajectories. Open Biol. 2021, 11, 200247. [Google Scholar] [CrossRef]
- Ghaderi, M.; Fernández Moro, C.; Pouso Elduayen, S.; Hultin, E.; Verbeke, C.S.; Björnstedt, M.; Dillner, J. Genome-wide transcriptome profiling of ex-vivo precision-cut slices from human pancreatic ductal adenocarcinoma. Sci. Rep. 2020, 10, 9070. [Google Scholar] [CrossRef] [PubMed]
- Bazou, D.; Maimon, N.; Gruionu, G.; Grahovac, J.; Seano, G.; Liu, H.; Evans, C.L.; Munn, L.L. Vascular beds maintain pancreatic tumor explants for ex vivo drug screening. J. Tissue Eng. Regen. Med. 2018, 12, e318–e322. [Google Scholar] [CrossRef] [PubMed]
- Aoki, R.; Aoki-Yoshida, A.; Suzuki, C.; Takayama, Y. Indole-3-Pyruvic Acid, an Aryl Hydrocarbon Receptor Activator, Suppresses Experimental Colitis in Mice. J. Immunol. 2018, 201, 3683–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Moro, C.F.; del Chiaro, M.; Pouso, S.; Sebestyén, A.; Löhr, M.; Björnstedt, M.; Verbeke, C.S. Ex vivo organotypic culture system of precision-cut slices of human pancreatic ductal adenocarcinoma. Sci. Rep. 2019, 9, 2133. [Google Scholar] [CrossRef] [Green Version]
- Bostick, M.; Bolduc, N.; Lehman, A.; Farmer, A. Strand-Specific Transcriptome Sequencing Using SMART Technology. Curr. Protoc. Mol. Biol. 2016, 116, 4.27.1–4.27.18. [Google Scholar] [CrossRef] [PubMed]
- Moro, C.F.; Selvam, A.K.; Ghaderi, M.; Pimenoff, V.N.; Gerling, M.; Bozóky, B.; Elduayen, S.P.; Dillner, J.; Björnstedt, M. Drug-induced tumor-specific cytotoxicity in a whole tissue ex vivo model of human pancreatic ductal adenocarcinoma. Front. Oncol. 2022, 12, 965182. [Google Scholar] [CrossRef] [PubMed]
- Kallio, M.A.; Tuimala, J.T.; Hupponen, T.; Klemelä, P.; Gentile, M.; Scheinin, I.; Koski, M.; Käki, J.; Korpelainen, E.I. Chipster: User-friendly analysis software for microarray and other high-throughput data. BMC Genom. 2011, 12, 507. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Herwig, R.; Hardt, C.; Lienhard, M.; Kamburov, A. Analyzing and interpreting genome data at the network level with ConsensusPathDB. Nat. Protoc. 2016, 11, 1889–1907. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Giari, L.; Guerranti, C.; Tognon, M.; Castaldelli, G.; Fano, E.A.; Martini, F. Environmental doses of perfluorooctanoic acid change the expression of genes in target tissues of common carp. Environ. Toxicol. Chem. 2018, 37, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Beischlag, T.V.; Morales, J.L.; Hollingshead, B.D.; Perdew, G.H. The Aryl Hydrocarbon Receptor Complex and the Control of Gene Expression. Crit Rev. Eukaryot. Gene Expr. 2008, 18, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, E.A.; Mezrich, J.D.; Bradfield, C.A. The aryl hydrocarbon receptor: A perspective on potential roles in the immune system. Immunology 2009, 127, 299. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Snyder, M.; Kenison, J.E.; Yang, K.; Lara, B.; Lydell, E.; Bennani, K.; Novikov, O.; Federico, A.; Monti, S.; et al. How the AHR Became Important in Cancer: The Role of Chronically Active AHR in Cancer Aggression. Int. J. Mol. Sci. 2021, 22, 387. [Google Scholar] [CrossRef] [PubMed]
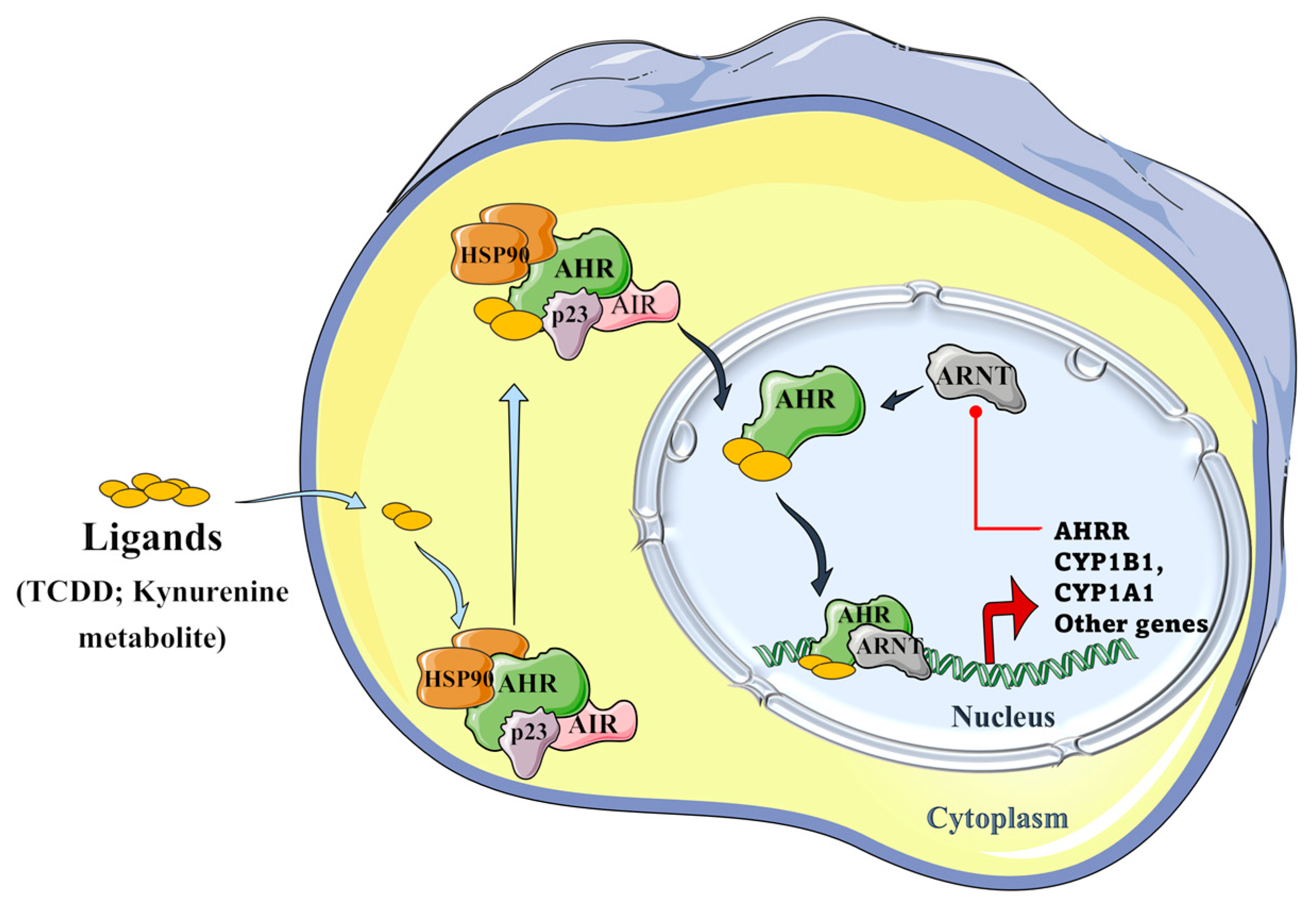
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture ID | Gender | Preoperative Chemotherapy | Histological Type | Grade of Differentiation | Stage * |
|---|---|---|---|---|---|
| 1 | Female | No | PDAC | Moderate | pT3 N2 |
| 2 | Male | No | PDAC | Poor | pT2 N1 |
| 3 | Male | No | PDAC | Moderate | pT2 N0 |
| 4 | Female | No | PDAC | Poor | pT3 N2 |
| 5 | Male | No | PDAC | Moderate–poor | pT3 N2 |
| 6 | Male | No | PDAC | Moderate | pT3 N2 |
| 7 | Female | No | PDAC | Moderate–poor | pT3 N2 |
| p-Value | Pathway | Database | Genes Mapped |
|---|---|---|---|
| 1.66 × 10−7 | Aryl Hydrocarbon Receptor | Wikipathways | AHRR; CYP1A1; CYP1B1 |
| 2.13 × 10−7 | Amodiaquine Pathway Pharmacokinetics | PharmGKB | CYP1A1; CYP1B1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szekerczés, T.; Selvam, A.K.; Moro, C.F.; Elduayen, S.P.; Dillner, J.; Björnstedt, M.; Ghaderi, M. Exploration of Patient-Derived Pancreatic Ductal Adenocarcinoma Ex Vivo Tissue for Treatment Response. Antioxidants 2023, 12, 167. https://doi.org/10.3390/antiox12010167
Szekerczés T, Selvam AK, Moro CF, Elduayen SP, Dillner J, Björnstedt M, Ghaderi M. Exploration of Patient-Derived Pancreatic Ductal Adenocarcinoma Ex Vivo Tissue for Treatment Response. Antioxidants. 2023; 12(1):167. https://doi.org/10.3390/antiox12010167
Chicago/Turabian StyleSzekerczés, Tímea, Arun Kumar Selvam, Carlos Fernández Moro, Soledad Pouso Elduayen, Joakim Dillner, Mikael Björnstedt, and Mehran Ghaderi. 2023. "Exploration of Patient-Derived Pancreatic Ductal Adenocarcinoma Ex Vivo Tissue for Treatment Response" Antioxidants 12, no. 1: 167. https://doi.org/10.3390/antiox12010167