
Alopecia Areata: A Review of the Role of Oxidative Stress, Possible Biomarkers, and Potential Novel Therapeutic Approaches
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
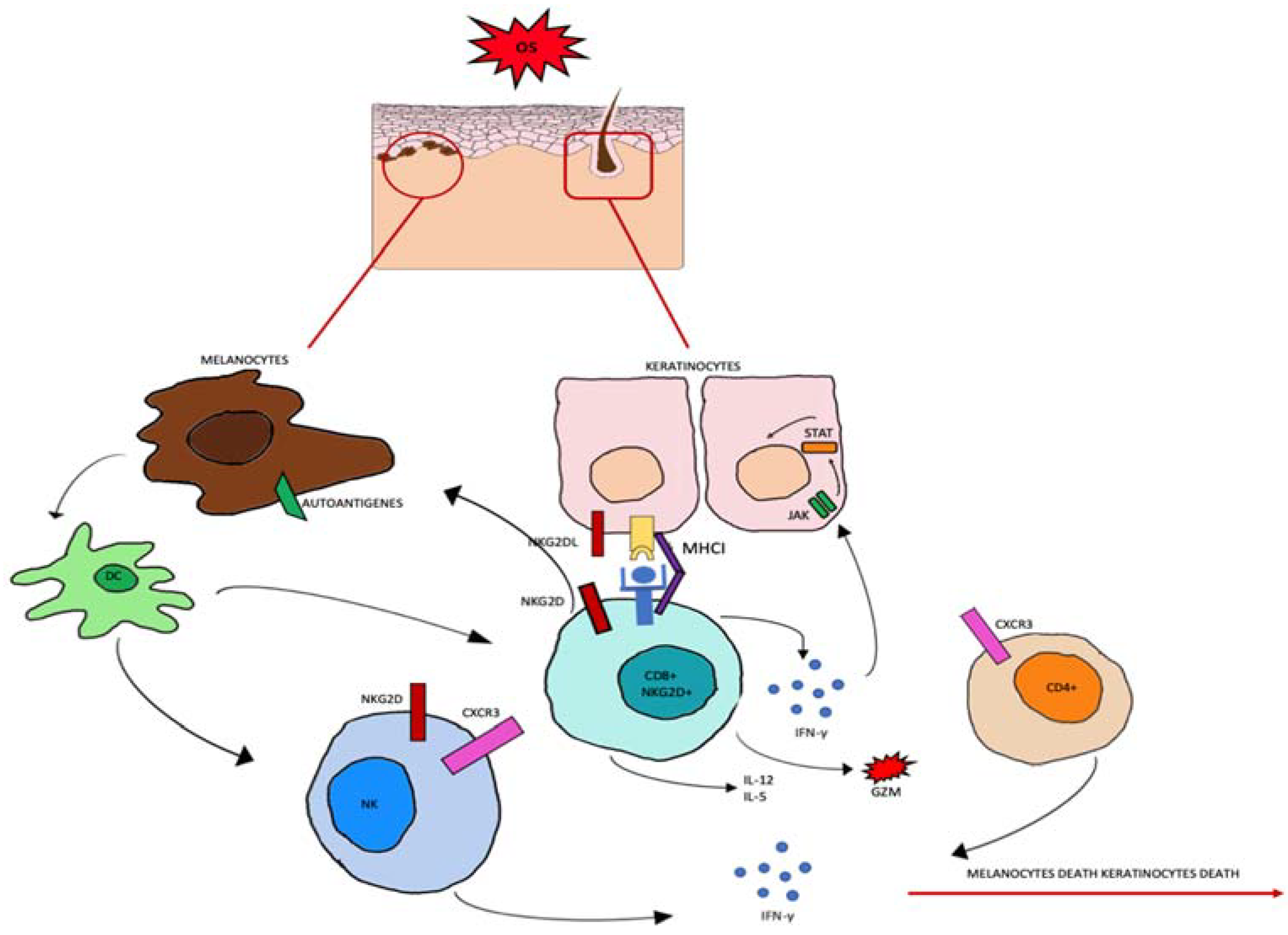
3.1. OS Role in Pathogenesis of AA
3.2. Autophagy Role in Pathogenesis of AA
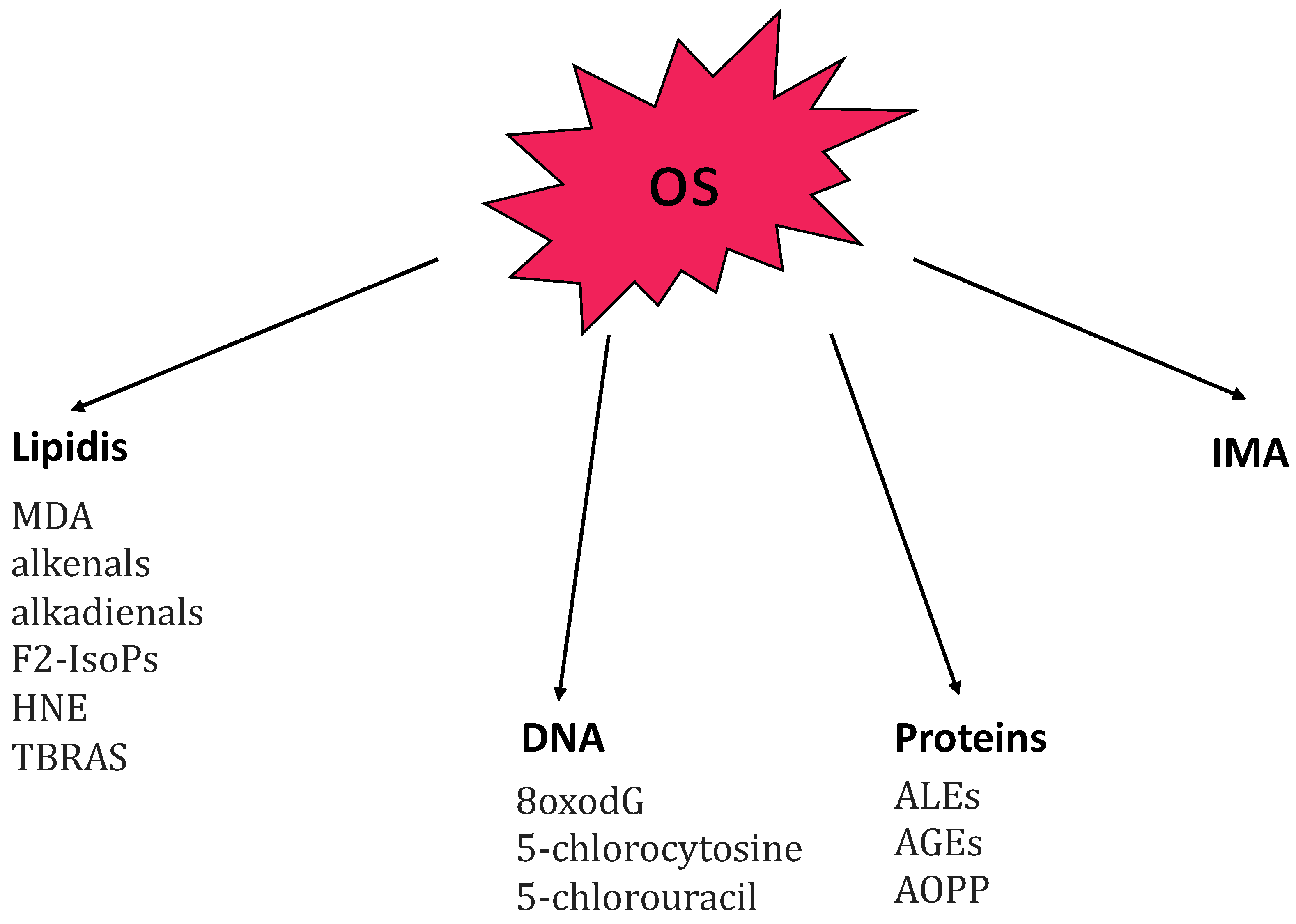
3.3. OS Biomarkers in AA
- -
- -
- advanced glycation end-products (AGEs) [46];
- -
3.4. Potential Therapeutic Target in AA
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Villasante Fricke, A.C.; Miteva, M. Epidemiology and burden of alopecia areata: A systematic review. Clin. Cosmet. Investig. Dermatol. 2015, 8, 397–403. [Google Scholar] [PubMed] [Green Version]
- Finner, A.M. Alopecia areata: Clinical presentation, diagnosis, and unusual cases. Dermatol. Ther. 2011, 24, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Mane, M.; Nath, A.K.; Thappa, D.M. Utility of dermoscopy in alopecia areata. Indian J. Dermatol. 2011, 56, 407–411. [Google Scholar]
- Chelidze, K.; Lipner, S.R. Nail changes in alopecia areata: An update and review. Int. J. Dermatol. 2018, 57, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Esmer, O.; Karadag, R.; Cakici, O.; Bilgili, S.G.; Demircan, Y.T.; Bayramlar, H.; Karadag, A.S. Ocular findings in patients with alopecia areata. Int. J. Dermatol. 2016, 55, 814–818. [Google Scholar] [CrossRef]
- Spano, F.; Donovan, J.C. Alopecia areata: Part 1: Pathogenesis, diagnosis, and prognosis. Can. Fam. Physician 2015, 61, 751–755. [Google Scholar]
- Trüeb, R.M.; Dias, M.F.R.G. Alopecia Areata: A Comprehensive Review of Pathogenesis and Management. Clin. Rev. Allergy Immunol. 2018, 54, 68–87. [Google Scholar] [CrossRef]
- Manolache, L.; Benea, V. Stress in patients with alopecia areata and vitiligo. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 921–928. [Google Scholar] [CrossRef]
- Bertolini, M.; McElwee, K.; Gilhar, A.; Bulfone-Paus, S.; Paus, R. Hair follicle immune privilege and its collapse in alopecia areata. Exp. Dermatol. 2020, 29, 703–725. [Google Scholar] [CrossRef]
- Strazzulla, L.C.; Wang, E.H.C.; Avila, L.; Lo Sicco, K.; Brinster, N.; Christiano, A.M.; Shapiro, J. Alopecia areata: Disease characteristics, clinical evaluation, and new perspectives on pathogenesis. J. Am. Acad. Dermatol. 2018, 78, 1–12. [Google Scholar] [CrossRef]
- Rajabi, F.; Drake, L.A.; Senna, M.M.; Rezaei, N. Alopecia areata: A review of disease pathogenesis. Br. J. Dermatol. 2018, 179, 1033–1048. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, M.; Pretzlaff, M.; Sulk, M.; Bähr, M.; Gherardini, J.; Uchida, Y.; Reibelt, M.; Kinori, M.; Rossi, A.; Bíró, T.; et al. Vasoactive intestinal peptide, whose receptor-mediated signalling may be defective in alopecia areata, provides protection from hair follicle immune privilege collapse. Br. J. Dermatol. 2016, 175, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.H.C.; Yu, M.; Breitkopf, T.; Akhoundsadegh, N.; Wang, X.; Shi, F.T.; Leung, G.; Dutz, J.P.; Shapiro, J.; McElwee, K.J. Identification of Autoantigen Epitopes in Alopecia Areata. J. Investig. Dermatol. 2016, 136, 1617–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renert-Yuval, Y.; Guttman-Yassky, E. The Changing Landscape of Alopecia Areata: The Therapeutic Paradigm. Adv. Ther. 2017, 34, 1594–1609. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Meng, X.; Song, Z. Vitamin D and alopecia areata: Possible roles in pathogenesis and potential implications for therapy. Am. J. Transl. Res. 2019, 11, 5285–5300. [Google Scholar] [PubMed]
- Baek, J.; Lee, M.-G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative stress in the skin: Impact and related protection. Int. J. Cosmet. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef]
- Rusanova, I.; Martínez-Ruiz, L.; Florido, J.; Rodríguez-Santana, C.; Guerra-Librero, A.; Acuña-Castroviejo, D.; Escames, G. Protective Effects of Melatonin on the Skin: Future Perspectives. Int. J. Mol. Sci. 2019, 20, 4948. [Google Scholar] [CrossRef] [Green Version]
- Tampa, M.; Mitran, C.I.; Mitran, M.I.; Amuzescu, A.; Matei, C.; Georgescu, S.R. Ischemia-Modified Albumin-A Potential New Marker of Oxidative Stress in Dermatological Diseases. Medicina 2022, 58, 669. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and Clinical Significance of Biomarkers of Oxidative Stress in Humans. Oxidative Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef] [PubMed]
- Akar, A.; Arca, E.; Erbil, H.; Akay, C.; Sayal, A.; Gür, A.R. Antioxidant enzymes and lipid peroxidation in the scalp of patients with alopecia areata. J. Dermatol. Sci. 2002, 29, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Koca, R.; Armutcu, F.; Altinyazar, C.; Gürel, A. Evaluation of lipid peroxidation, oxidant/antioxidant status, and serum nitric oxide levels in alopecia areata. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2005, 11, CR296–CR299. [Google Scholar]
- Kim, S.W.; Kim, B.J.; Youn, S.W.; Park, K.C.; Huh, C.H. Evaluation of free oxygen radical and antioxidant capacity in alopecia areata. J. Dermatol. 2010, 37, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Abdel Fattah, N.S.; Ebrahim, A.A.; El Okda, E.S. Lipid peroxidation/antioxidant activity in patients with alopecia areata. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, R.; Tawdy, A.; Hay, R.A.; Rashed, L.; Tawfik, D. The antioxidant role of paraoxonase 1 and vitamin E in three autoimmune diseases. Skin Pharmacol. Physiol. 2013, 26, 2–7. [Google Scholar] [CrossRef]
- Bilgili, S.G.; Ozkol, H.; Karadag, A.S.; Ozkol, H.U.; Seker, A.; Calka, O.; Aslan, M. Serum paraoxonase activity and oxidative status in subjects with alopecia areata. Cutan. Ocul. Toxicol. 2013, 32, 290–293. [Google Scholar] [CrossRef]
- Motor, S.; Ozturk, S.; Ozcan, O.; Gurpinar, A.B.; Can, Y.; Yuksel, R.; Yenin, J.Z.; Seraslan, G.; Ozturk, O.H. Evaluation of total antioxidant status, total oxidant status and oxidative stress index in patients with alopecia areata. Int. J. Clin. Exp. Med. 2014, 7, 1089–1093. [Google Scholar]
- Bakry, A.; Elshazly, R.M.A.; Shoeib, M.A.M.; Gooda, A. Oxidative stress in alopecia areata: A case-control study. Am. J. Clin. Dermatol. 2014, 15, 57–64. [Google Scholar] [CrossRef]
- Alzolibani, A.A. Preferential recognition of hydroxyl radical-modified superoxide dismutase by circulating autoantibodies in patients with alopecia areata. Ann. Dermatol. 2014, 26, 576–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, Z.; Alzolibani, A.A.; Al-Shobaili, H.A.; Saif, G.B.; Al Robaee, A.A. Biochemical and immunological studies on erythrocytes superoxide dismutase modified by nitric oxide in patients with alopecia areata: Implications in alopecia patchy persistent and alopecia universalis. Immunol. Lett. 2014, 160, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, G.; Seçkin, H.Y.; Benli, İ.; Akbaş, A.; Baş, Y.; Karakus, N.; Bütün, İ.; Özyurt, H. Relationship between manganese superoxide dismutase (MnSODAla-9Val) and glutathione peroxidase (GPx1 Pro 197 Leu) gene polymorphisms and alopecia areata. Int. J. Clin. Exp. Med. 2015, 8, 21533–21540. [Google Scholar] [PubMed]
- Yenin, J.Z.; Serarslan, G.; Yönden, Z.; Ulutaş, K.T. Investigation of oxidative stress in patients with alopecia areata and its relationship with disease severity, duration, recurrence and pattern. Clin. Exp. Dermatol. 2015, 40, 617–621. [Google Scholar] [CrossRef]
- Öztürk, P.; Arıcan, Ö.; Kurutaş, E.B.; Mülayim, K. Oxidative Stress Biomarkers and Adenosine Deaminase over the Alopecic Area of the Patients with Alopecia Areata. Balkan Med. J. 2016, 33, 188–192. [Google Scholar] [CrossRef]
- Cwynar, A.; Olszewska-Słonina, D.; Czajkowski, R.; Zegarska, B.; Białecka, A.; Męcińska-Jundziłł, K.; Piskorska, E.; Lampka, M. Investigation of oxidative stress in patients with alopecia areata by measuring the levels of malondialdehyde and ceruloplasmin in the blood. Postepy Dermatol. Alergol. 2018, 35, 572–576. [Google Scholar] [CrossRef] [Green Version]
- Ataş, H.; Gönül, M.; Öztürk, Y.; Kavutçu, M. Ischemic modified albumin as a new biomarker in predicting oxidative stress in alopecia areata. Turk. J. Med. Sci. 2019, 49, 129–138. [Google Scholar] [CrossRef]
- Cwynar, A.; Olszewska-Słonina, D.; Czajkowski, R.; Zegarska, B.; Wybranowski, T.; Hołyńska-Iwan, I.; Piskorska, E.; Kaczorowski, P.; Woźniak, A. Evaluation of selected parameters of oxidative stress in patients with alopecia areata. Postepy Dermatol. Alergol. 2019, 36, 115–116. [Google Scholar] [CrossRef]
- Incel-Uysal, P.; Akdogan, N.; Alli, N.; Oktem, A.; Candar, T.; Topcuoglu, C.; Turhan, T. Assessment of Metabolic Profile and Ischemia-modified Albumin Level in Patients with Alopecia Areata: A Case-Control Study. Indian J. Dermatol. 2019, 64, 12–18. [Google Scholar] [CrossRef]
- Dizen-Namdar, N.; Emel Kocak, F.; Kidir, M.; Sarici, G.; Tak, H.; Altuntas, I. Evaluation of Serum Paraoxonase, Arylesterase, Prolidase Activities and Oxidative Stress in Patients with Alopecia Areata. Skin Pharmacol. Physiol. 2019, 32, 59–64. [Google Scholar] [CrossRef]
- Abbas, A.N. Ginger (Zingiber officinale (L.) Rosc) improves oxidative stress and trace elements status in patients with alopecia areata. Niger. J. Clin. Pract. 2020, 23, 1555–1560. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, K.; Kozłowska, M.; Kaszuba, A.; Lesiak, A.; Narbutt, J.; Zalewska-Janowska, A. Increased Serum Levels of IFN-γ, IL-1β, and IL-6 in Patients with Alopecia Areata and Nonsegmental Vitiligo. Oxidative Med. Cell. Longev. 2020, 2020, 5693572. [Google Scholar] [CrossRef] [PubMed]
- Khaki, L.; Vaezi, G.; Ayatollahi, A.; Firooz, A. Evaluation of Glutathione Reductase and Glutathione Peroxidase in the Serum of Iranian Patients with Alopecia Areata: A Case-control Study. Iran J. Allergy Asthma Immunol. 2020, 19, 676–678. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.I.; Khashaba, R.A.; Fawzy, E.; Baghdady, S.M.A.; Rezk, S.M. Cross talk between oxidative stress and inflammation in alopecia areata. J. Cosmet. Dermatol. 2021, 20, 2305–2310. [Google Scholar] [CrossRef]
- Sachdeva, S.; Khurana, A.; Goyal, P.; Sardana, K. Does oxidative stress correlate with disease activity and severity in alopecia areata? An analytical study. J. Cosmet. Dermatol. 2022, 21, 1629–1634. [Google Scholar] [CrossRef]
- Shakoei, S.; Mirmiranpoor, H.; Nakhjavani, M.; Nasimi, M.; Bakhshi, G.; Azizpour, A. Oxidative stress and antioxidant markers in patients with alopecia areata: A comparative cross-sectional study. Indian J. Dermatol. Venereol. Leprol. 2022, 1–5. [Google Scholar] [CrossRef]
- Taskin, S.; Celik, H.; Cakirca, G.; Manav, V.; Taskin, A. Nitric oxide synthase activity: A novel potential biomarker for predicting Alopecia areata. J. Cosmet. Dermatol. 2022, 21, 7075–7080. [Google Scholar] [CrossRef]
- Juárez-Rendón, K.J.; Rivera Sánchez, G.; Reyes-López, M.Á.; García-Ortiz, J.E.; Bocanegra-García, V.; Guardiola-Avila, I.; Altamirano-García, M.L. Alopecia Areata. Current situation and perspectives. Arch. Argent Pediatr. 2017, 115, e404–e411, (In English, Spanish). [Google Scholar]
- Cannavò, S.P.; Riso, G.; Casciaro, M.; Di Salvo, E.; Gangemi, S. Oxidative stress involvement in psoriasis: A systematic review. Free Radic. Res. 2019, 53, 829–840. [Google Scholar] [CrossRef]
- Jung, J.M.; Yang, H.J.; Lee, W.J.; Won, C.H.; Lee, M.W.; Chang, S.E. Association between psoriasis and alopecia areata: A systematic review and meta-analysis. J. Dermatol. 2022, 49, 912–915. [Google Scholar] [CrossRef]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, P.; Mathur, M.C. Oxidative stress in alopecia areata: A systematic review and meta-analysis. Int. J. Dermatol. 2020, 59, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Alzolibani, A.A.; Rasheed, Z.; Al Robaee, A.A. Acquired immunogenicity of DNA after modification with malondialdehyde in patients with alopecia areata. Scand. J. Clin. Lab. Investig. 2014, 74, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, V.; Singh, C. IFN-γ regulates xanthine oxidase-mediated iNOS-independent oxidative stress in maneb- and paraquat-treated rat polymorphonuclear leukocytes. Mol. Cell. Biochem. 2017, 427, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Shi, Y.; Huang, W.; Lao, C.; Zou, Z.; Pan, S.; Huang, Z. Theobromine mitigates IL-1β-induced oxidative stress, inflammatory response, and degradation of type II collagen in human chondrocytes. Int. Immunopharmacol. 2020, 82, 106226. [Google Scholar] [CrossRef]
- Liu, C.W.; Lee, T.L.; Chen, Y.C.; Liang, C.J.; Wang, S.H.; Lue, J.H.; Tsai, J.S.; Lee, S.W.; Chen, S.H.; Yang, Y.F.; et al. PM2.5-induced oxidative stress increases intercellular adhesion molecule-1 expression in lung epithelial cells through the IL-6/AKT/STAT3/NF-κB-dependent pathway. Part. Fibre Toxicol. 2018, 15, 4. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, S.; Li, C. Perspectives of New Advances in the Pathogenesis of Vitiligo: From Oxidative Stress to Autoimmunity. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 1017–1023. [Google Scholar] [CrossRef]
- Dell’Anna, M.L.; Maresca, V.; Briganti, S.; Camera, E.; Falchi, M.; Picardo, M. Mitochondrial impairment in peripheral blood mononuclear cells during the active phase of vitiligo. J. Investig. Dermatol. 2001, 117, 908–913. [Google Scholar] [CrossRef] [Green Version]
- Maresca, V.; Roccella, M.; Roccella, F.; Camera, E.; Del Porto, G.; Passi, S.; Grammatico, P.; Picardo, M. Increased sensitivity to peroxidative agents as a possible pathogenic factor of melanocyte damage in vitiligo. J. Investig. Dermatol. 1997, 109, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Jimbow, K.; Chen, H.; Park, J.S.; Thomas, P.D. Increased sensitivity of melanocytes to oxidative stress and abnormal expression of tyrosinase-related protein in vitiligo. Br. J. Dermatol. 2001, 144, 55–65. [Google Scholar] [CrossRef]
- Speeckaert, R.; Dugardin, J.; Lambert, J.; Lapeere, H.; Verhaeghe, E.; Speeckaert, M.M.; van Geel, N. Critical appraisal of the oxidative stress pathway in vitiligo: A systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, M.; Conic, R.R.Z.; Atanaskova Mesinkovska, N. Hair Follicle Melanogenesis Reflected in Hair Pigmentation as a Developmental Factor in Alopecia Areata. Skin Appendage Disord. 2021, 7, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, S.; Li, C. Mechanisms of melanocyte death in vitiligo. Med. Res. Rev. 2021, 41, 1138–1166. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Hatanaka, Y.; Sutoh, Y.; Suzuki, Y.; Oba, K.; Hatanaka, K.C.; Mitsuhashi, T.; Otsuka, N.; Fugo, K.; Kasahara, M.; et al. Immunohistochemical validation and expression profiling of NKG2D ligands in a wide spectrum of human epithelial neoplasms. J. Histochem. Cytochem. 2015, 63, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Gilhar, A.; Etzioni, A.; Paus, R. Alopecia areata. N. Engl. J. Med. 2012, 366, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Gilhar, A. Collapse of immune privilege in alopecia areata: Coincidental or substantial? J. Investig. Dermatol. 2010, 130, 2535–2537. [Google Scholar] [CrossRef] [Green Version]
- Kwon, K.W.; Kim, S.J.; Kim, H.; Kim, W.S.; Kang, S.M.; Choi, E.; Ha, S.J.; Yoon, J.H.; Shin, S.J. IL-15 Generates IFN-γ-producing Cells Reciprocally Expressing Lymphoid-Myeloid Markers during Dendritic Cell Differentiation. Int. J. Biol. Sci. 2019, 15, 464–480. [Google Scholar] [CrossRef]
- Edamitsu, T.; Taguchi, K.; Okuyama, R.; Yamamoto, M. AHR and NRF2 in Skin Homeostasis and Atopic Dermatitis. Antioxidants 2022, 11, 227. [Google Scholar] [CrossRef]
- Panieri, E.; Telkoparan-Akillilar, P.; Saso, L. NRF2, a crucial modulator of skin cells protection against vitiligo, psoriasis, and cancer. Biofactors 2022, 1–23. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B. Autophagy in Human Diseases. N. Engl. J. Med. 2020, 383, 1564–1576. [Google Scholar] [CrossRef]
- Albano, G.D.; Gagliardo, R.P.; Montalbano, A.M.; Profita, M. Overview of the Mechanisms of Oxidative Stress: Impact in Inflammation of the Airway Diseases. Antioxidants 2022, 11, 2237. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.; Kim, S.K.; Park, H.; Kim, J.E.; Lee, S. Autophagy: Guardian of Skin Barrier. Biomedicines 2022, 10, 1817. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Sun, J.; Song, X. Hair Follicle Melanocytes Initiate Autoimmunity in Alopecia Areata: A Trigger Point. Clin. Rev. Allergy Immunol. 2022, 63, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [Green Version]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Khan, A.Q.; Agha, M.V.; Sheikhan, K.S.A.M.; Younis, S.M.; Tamimi, M.A.; Alam, M.; Ahmad, A.; Uddin, S.; Buddenkotte, J.; Steinhoff, M. Targeting deregulated oxidative stress in skin inflammatory diseases: An update on clinical importance. Biomed. Pharmacother. 2022, 154, 113601. [Google Scholar] [CrossRef]
- Cho, S.J.; Roman, G.; Yeboah, F.; Konishi, Y. The road to advanced glycation end products: A mechanistic perspective. Curr. Med. Chem. 2007, 14, 1653–1671. [Google Scholar] [CrossRef]
- Hansel, B.; Kontush, A.; Bonnefont-Rousselot, D.; Bruckert, E.; Chapman, M.J. Alterations in lipoprotein defense against oxidative stress in metabolic syndrome. Curr. Atheroscler. Rep. 2006, 8, 501–509. [Google Scholar] [CrossRef]
- Yan, S.F.; D’Agati, V.; Schmidt, A.M.; Ramasamy, R. Receptor for Advanced Glycation Endproducts (RAGE): A formidable force in the pathogenesis of the cardiovascular complications of diabetes & aging. Curr. Mol. Med. 2007, 7, 699–710. [Google Scholar]
- Ozgor, O.; Akoglu, G.; Sungu, N.; Karaismailoglu, E.; Aktas, A. Receptor for advanced glycation end products is overexpressed in psoriatic plaques independent of disease severity. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 556–560. [Google Scholar]
- Gupta, S.C.; Kismali, G.; Aggarwal, B.B. Curcumin, a component of turmeric: From farm to pharmacy. Biofactors 2013, 39, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Soboleva, A.G.; Bruskin, S.A.; Nikolaev, A.A.; Sobolev, V.V.; Mezentsev, A.V. [Role of receptor for advanced glycation end-products in pathgenesis of psoriasis]. Mol. Biol. 2013, 47, 743–753. (In Russian) [Google Scholar] [CrossRef]
- Haslam, I.S.; Jadkauskaite, L.; Szabó, I.L.; Staege, S.; Hesebeck-Brinckmann, J.; Jenkins, G.; Bhogal, R.K.; Lim, F.L.; Farjo, N.; Farjo, B.; et al. Oxidative Damage Control in a Human (Mini-) Organ: Nrf2 Activation Protects against Oxidative Stress-Induced Hair Growth Inhibition. J. Investig. Dermatol. 2017, 137, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadkauskaite, L.; Coulombe, P.A.; Schäfer, M.; Dinkova-Kostova, A.T.; Paus, R.; Haslam, I.S. Oxidative stress management in the hair follicle: Could targeting NRF2 counter age-related hair disorders and beyond? Bioessays 2017, 39. [Google Scholar] [CrossRef]
- Simakou, T.; Butcher, J.P.; Reid, S.; Henriquez, F.L. Alopecia areata: A multifactorial autoimmune condition. J. Autoimmun. 2019, 98, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.A.; Kim, Y.E.; Kwon, O.; Park, H. Treatment outcome of oral tofacitinib and ruxolitinib in patients with alopecia areata: A systematic review and meta-analysis. Indian J. Dermatol. Venereol. Leprol. 2021, 87, 621–627. [Google Scholar] [CrossRef]
- Behrangi, E.; Barough, M.S.; Khoramdad, M.; Hejazi, P.; Koltapeh, M.P.; Goodarzi, A. Efficacy and safety of tofacitinib for treatment of alopecia areata in children: A systematic review and meta-analysis. J. Cosmet. Dermatol. 2022, 21, 6644–6652. [Google Scholar] [CrossRef]
- Abdel Fattah, N.S.; Atef, M.M.; Al-Qaradaghi, S.M. Evaluation of serum zinc level in patients with newly diagnosed and resistant alopecia areata. Int. J. Dermatol. 2016, 55, 24–29. [Google Scholar] [CrossRef]
- Kil, M.S.; Kim, C.W.; Kim, S.S. Analysis of serum zinc and copper concentrations in hair loss. Ann. Dermatol. 2013, 25, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Dastgheib, L.; Mostafavi-Pour, Z.; Abdorazagh, A.A.; Khoshdel, Z.; Sadati, M.S.; Ahrari, I.; Ahrari, S.; Ghavipisheh, M. Comparison of zn, cu, and fe content in hair and serum in alopecia areata patients with normal group. Dermatol. Res. Pract. 2014, 2014, 784863. [Google Scholar] [CrossRef]
- Hardman, J.A.; Nicu, C.; Tai, C.; Harries, M.; Mironov, A.; Purba, T.S.; Paus, R. Does dysfunctional autophagy contribute to immune privilege collapse and alopecia areata pathogenesis? J. Dermatol. Sci. 2020, 100, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Wikramanayake, T.C.; Villasante, A.C.; Mauro, L.M.; Perez, C.I.; Schachner, L.A.; Jimenez, J.J. Prevention and treatment of alopecia areata with quercetin in the C3H/HeJ mouse model. Cell Stress Chaperones 2012, 17, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.M.; Mirza, M.A.; Park, M.K.; Qureshi, A.A.; Cho, E. The Role of Micronutrients in Alopecia Areata: A Review. Am. J. Clin. Dermatol. 2017, 18, 663–679. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Authors (Reference) | Year | Number of Patients | Tissues | Markers of Oxidative/Stress | Oxidative Stress Main Outcome(s) |
|---|---|---|---|---|---|
| Akar A. [23] | 2002 | 10 AA 10 HC | Scalp samples | TBARS, SOD, GSH-Px | TBARS, SOD, and GSH-Px were significantly higher than those of controls levels of TBARS, SOD, and GSH-Px in early phase of disease |
| Koka [24] | 2005 | 24 AA 20HC | Blood samples | MDA, NO (nitrite/nitrate), XO and SOD activity | MDA, NO (nitrite/nitrate), and XO activity levels were higher in AA patients than in controls; SOD activity was lower |
| Kim S. W. [25] | 2010 | 16 pz AA_ 16 HC_ | Blood samples_ | ROM, AC | ROM were increased and AC was reduced versus controls |
| Fattah N. S. A. [26] | 2011 | 50 AA 50 HC | Punch biopsies | MDA, SOD | MDA levels were high in AA patients and SOD activity was low |
| Ramadan S. [27] | 2012 | 15 AA 15 HC | Punch biopsies Blood samples | PON1, Vitamin E | Lower tissue and serum PON1 and Vitamin E levels in the patients than in the controls |
| Bilgili S. G. [28] | 2013 | 39 AA 39 HC | Blood samples | PON1, TAC, TOS, OSI |
TAC levels and PON1 activity were lower in AA patients than in controls; TOS levels and OSI were significantly higher
|
| Sedat Motor [29] | 2014 | 46 AA 36 hc | Blood samples | TAS, TOS | TAS, TOS, and OSİ levels showed no significant difference between the control and AA groups |
| Bakry O. A. [30] | 2014 | 35 AA 30 HC | Blood samples | OSI, TOC, TAC, MDA | TOC, MDA, and OSI were high in AA patients and TAC value was low; higher MDA and OSI values and lower TAC values were found in severe AA than in mild or moderate AA |
| Alzolibani A. A. [31] | 2014 | 26 AA_ 30 HC_ | Blood samples | SOD | SOD activity was reduced in AA patients compared to controls |
| Rasheed Z. [32] | 2014 | 26 AA_ 30 HC_ | Blood samples | NO, SOD | NO was elevated and SOD activity was reduced in AA patients compared to controls |
| Kalkan G. [33] | 2015 | 119 AA 104 HC | nSOD Ala-9Val and GPx1 Pro 198 Leu polymorphisms and AA susceptibility | nSOD Ala-9Val SNP genotype distributions and allele frequencies of the AA patients and the control group | |
| Yenin J. Z. [34] | 2015 | 62 AA 62 HC | Blood samples | MD and CAT, SOD, GSH-Px | No statistically significant difference in patient plasma MDA levels, CAT, GSH-Px, or SOD activities with regard to AA severity, duration, recurrence, or pattern |
| Perihan Öztürk [35] | 2016 | 30 AA 30 HC | Scalp-scrapes | ADA | Factors associated with oxidative stress were elevated in AA patients |
| Cwynar A. [36] | 2018 |
24 AA
22 HC | Blood samples | MDA, CER | MDA high in AA patients |
| Ataş H. [37] | 2019 | Blood samples | IMA | IMA levels increased in AA patients | |
| Cwynar A. [38] | 2019 | 30 AA 30 HC | Blood samples | PON1, MDA, AOPPs | |
| Incel-Uysal P. [39] | 2019 | 35 AA 35 HC | Blood samples | IMA, sd-LDL, and visfatin levels | IMA levels increased in AA patients |
| Nazli Dizen-Namdar [40] | 2019 |
60 AA,
50 HC | Blood samples | Serum PON1, prolidase, arylesterase activities, TOS, TAS, OSI | TOS and OSI levels and prolidase were high in AA patients; PON1 and arylesterase activities were low; no difference in serum TAS levels between the two groups |
| Abbas A. N. [41] | 2020 | 20 AA | Blood samples | GSH, MDA, TAS | Improvement of the antioxidant/oxidant balance of the erythrocytes and lymphocytes |
| Tomaszewska K. [42] | 2020 | 30 AA 30 Vit 30 HC | Blood samples | IFN-γ, IL-1β, IL-6 | Oxidative stress may play a significant role in promoting and amplifying the inflammatory process both in AA and vitiligo |
| Khaki L. [43] | 2020 | 56 AA 19 HC | Blood samples | GSH-Px, GSH-Rx | No differences in serum levels of glutathione reductase and glutathione peroxidase between the two groups |
| Mustafa A. I. [44] | 2021 | 49 AA 49 HC | Serum | 8-OHdG, HMBG1, CRP | High levels correlated with disease gravity |
| Sachdeva S. [45] | 2022 | 40 AA, 40 HC | Blood samples | MDA, SOD, TAS | TAS and SOD were lower in AA patients, MDA was higher |
| Shakoei S. [46] | 2022 | 40 AA, 40 HC | Blood samples |
Blood sugar, C-reactive protein, lipid profile, and
AOPPs, AGEs, PON1, lecithin-cholesterol acyltransferase and serum ferric-reducing antioxidant power | Advanced glycation end-products and advanced oxidation protein-products were significantly higher in patients with alopecia areata |
| Taskin S. [47] | 2022 | 30 AA 10 HC | Blood samples | NO, ONOO−, NOS activity | NO, ONOO−, and NOS activity were significantly higher in AA patients with than in the control group |
| Difference in Redox Balance between AA Patients and Healthy Controls | No Difference in Redox Balance between AA Patients and Healthy Controls | ||
|---|---|---|---|
| Authors | Markers of OS | Authors | Markers of OS |
| Bakry et al., 2014 | OSI, TOC | Motor et al., 2014 | TAS, TOS, OSI |
| Cwynar et al., 2018 | MDA, CER | Khaki et al., 2020 | GSH-Px, GSH-Rx |
| Bilgili et al., 2013 | TAC, TOS, OSI, PON1 | ||
| Nazli Dizen-Namdar et al., 2019 | TAS, TOS, OSI, PON1, ARE, prolidase enzyme activity | ||
| Akar et al., 2001 | TBRAS, SOD, GSH-Px | ||
| Tomaszewska. et al., 2020 | iNOS | ||
| Taskin et al., 2022 | NO, ONOO−, iNOS | ||
| Rasheed. et al., 2014 | NO, SOD | ||
| Kim et al., 2010 | ROM, AC | ||
| Koca et al., 2005 | MDA, NO, SOD, XO | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterle, L.; Sanfilippo, S.; Borgia, F.; Cicero, N.; Gangemi, S. Alopecia Areata: A Review of the Role of Oxidative Stress, Possible Biomarkers, and Potential Novel Therapeutic Approaches. Antioxidants 2023, 12, 135. https://doi.org/10.3390/antiox12010135
Peterle L, Sanfilippo S, Borgia F, Cicero N, Gangemi S. Alopecia Areata: A Review of the Role of Oxidative Stress, Possible Biomarkers, and Potential Novel Therapeutic Approaches. Antioxidants. 2023; 12(1):135. https://doi.org/10.3390/antiox12010135
Chicago/Turabian StylePeterle, Lucia, Serena Sanfilippo, Francesco Borgia, Nicola Cicero, and Sebastiano Gangemi. 2023. "Alopecia Areata: A Review of the Role of Oxidative Stress, Possible Biomarkers, and Potential Novel Therapeutic Approaches" Antioxidants 12, no. 1: 135. https://doi.org/10.3390/antiox12010135