
Basic Cognition of Melatonin Regulation of Plant Growth under Salt Stress: A Meta-Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
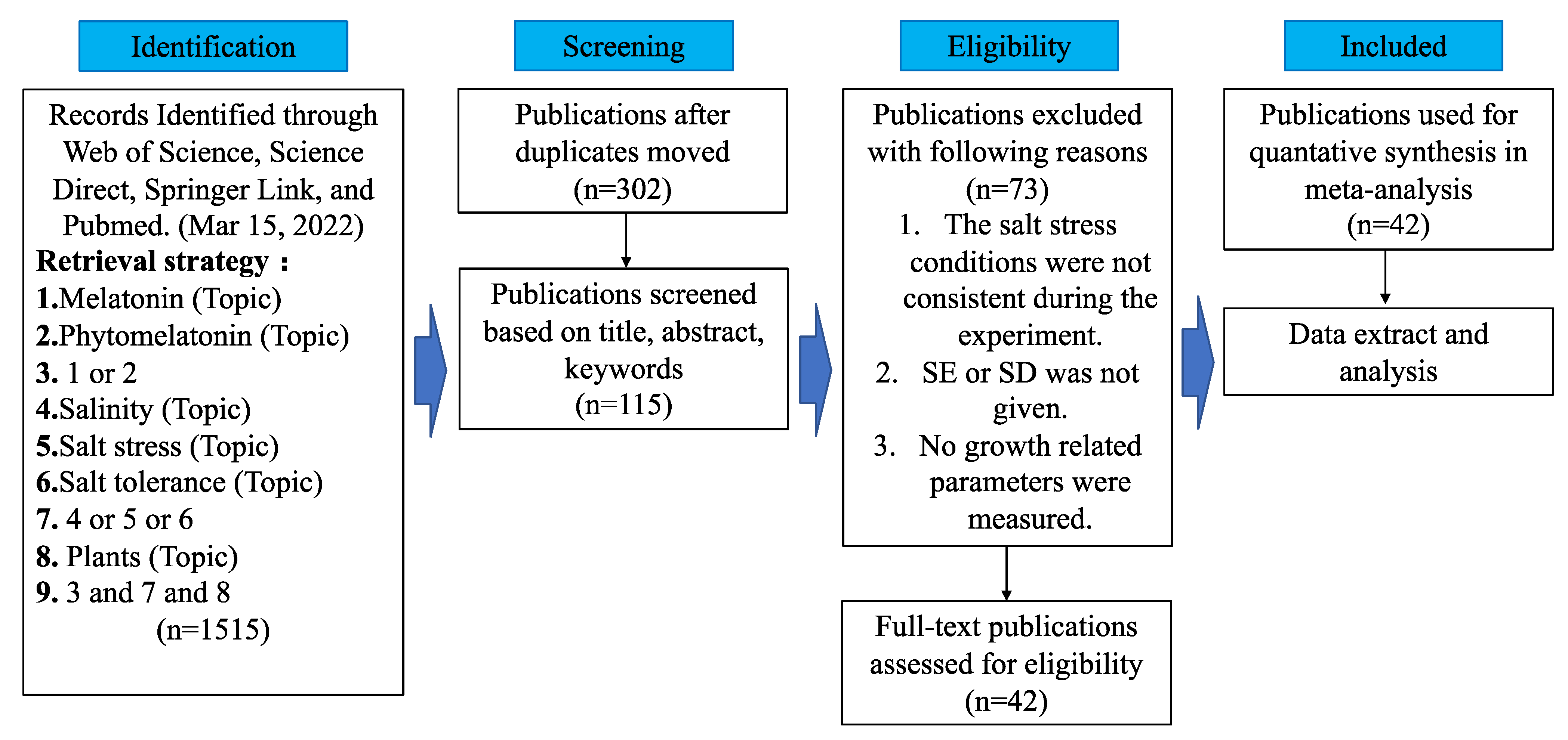
2.1. Literature Survey and Selection
2.2. Data Extraction
2.3. Meta-Analysis
2.3.1. Effect Sizes
2.3.2. Heterogeneity Test
2.3.3. Meta Subgroup Analysis
2.3.4. Publication Bias Test
2.4. Statistics
3. Results
3.1. Overall Response of Plant Growth to Exogenous MT
3.2. The Difference in Response of Growth Parameters to Exogenous MT
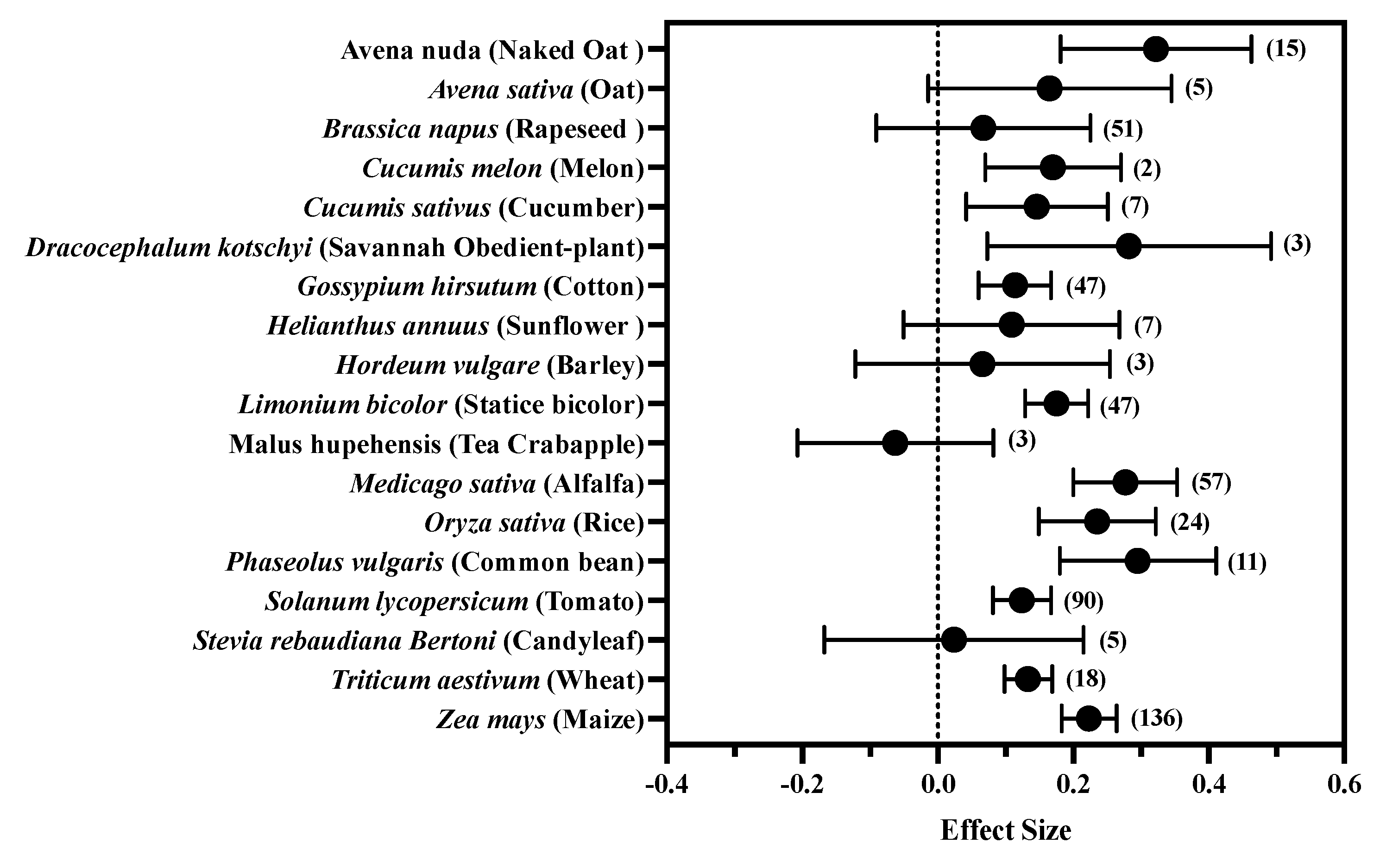
3.3. The Response of Plant Species to Exogenous MT
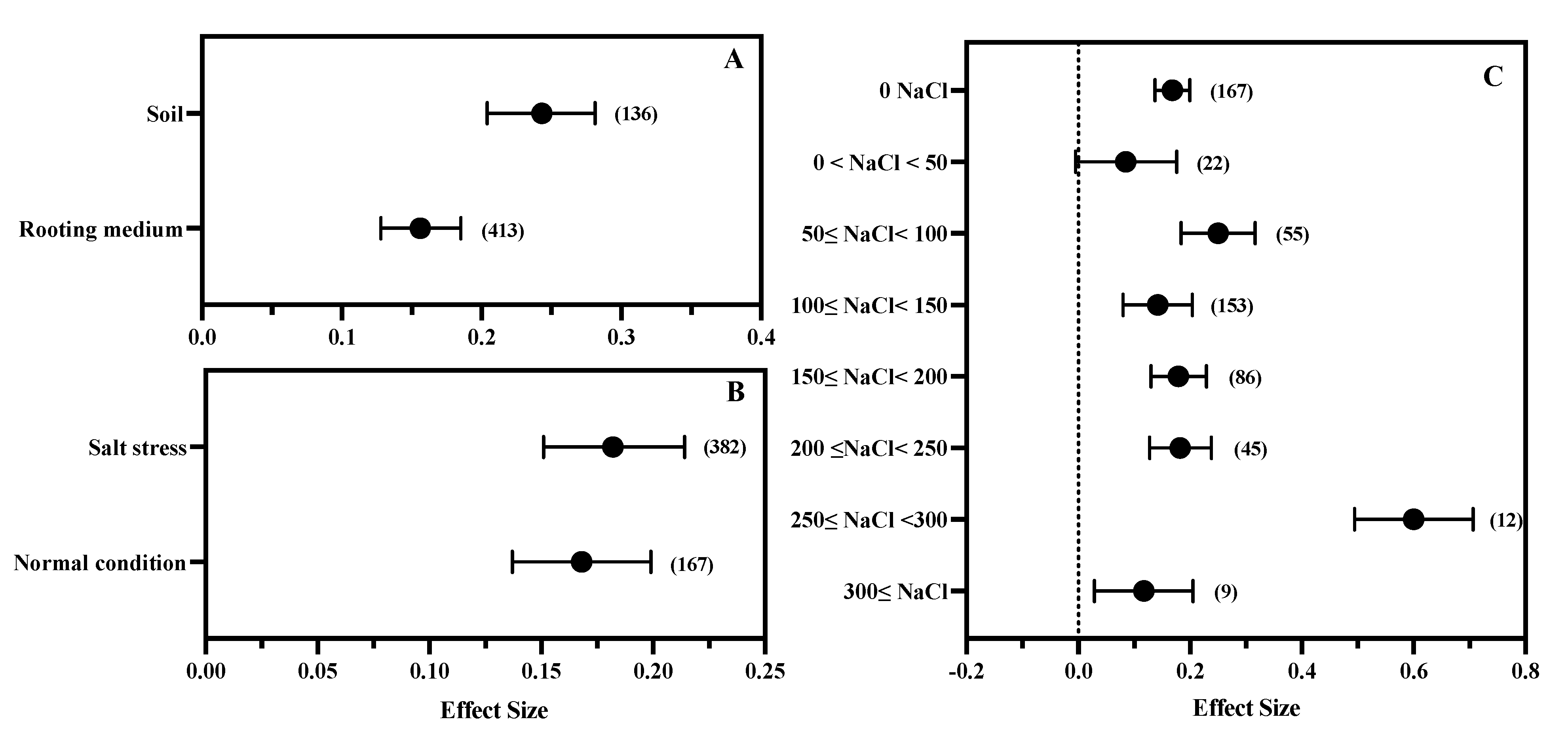
3.4. The Effect of Salt Stress Intensity and Rooting Environment
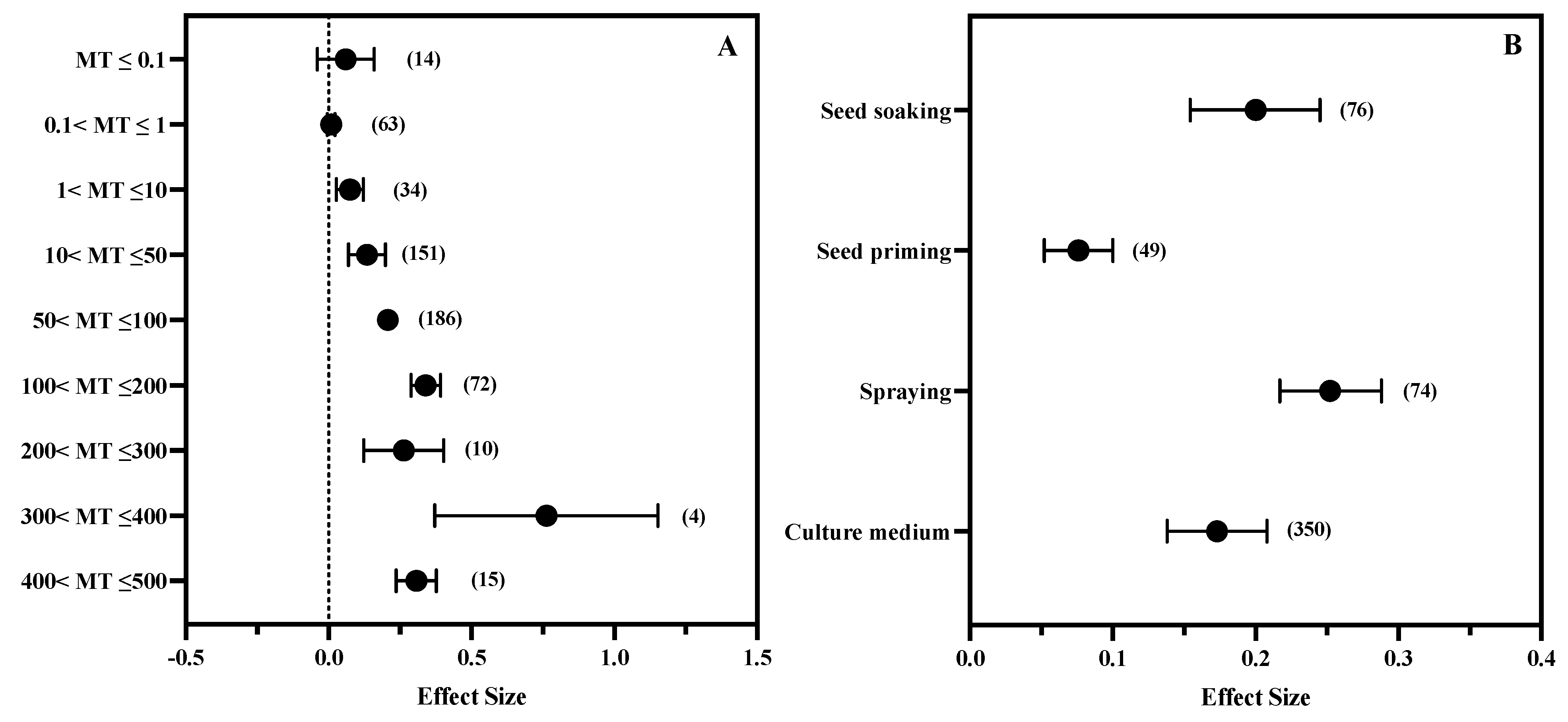
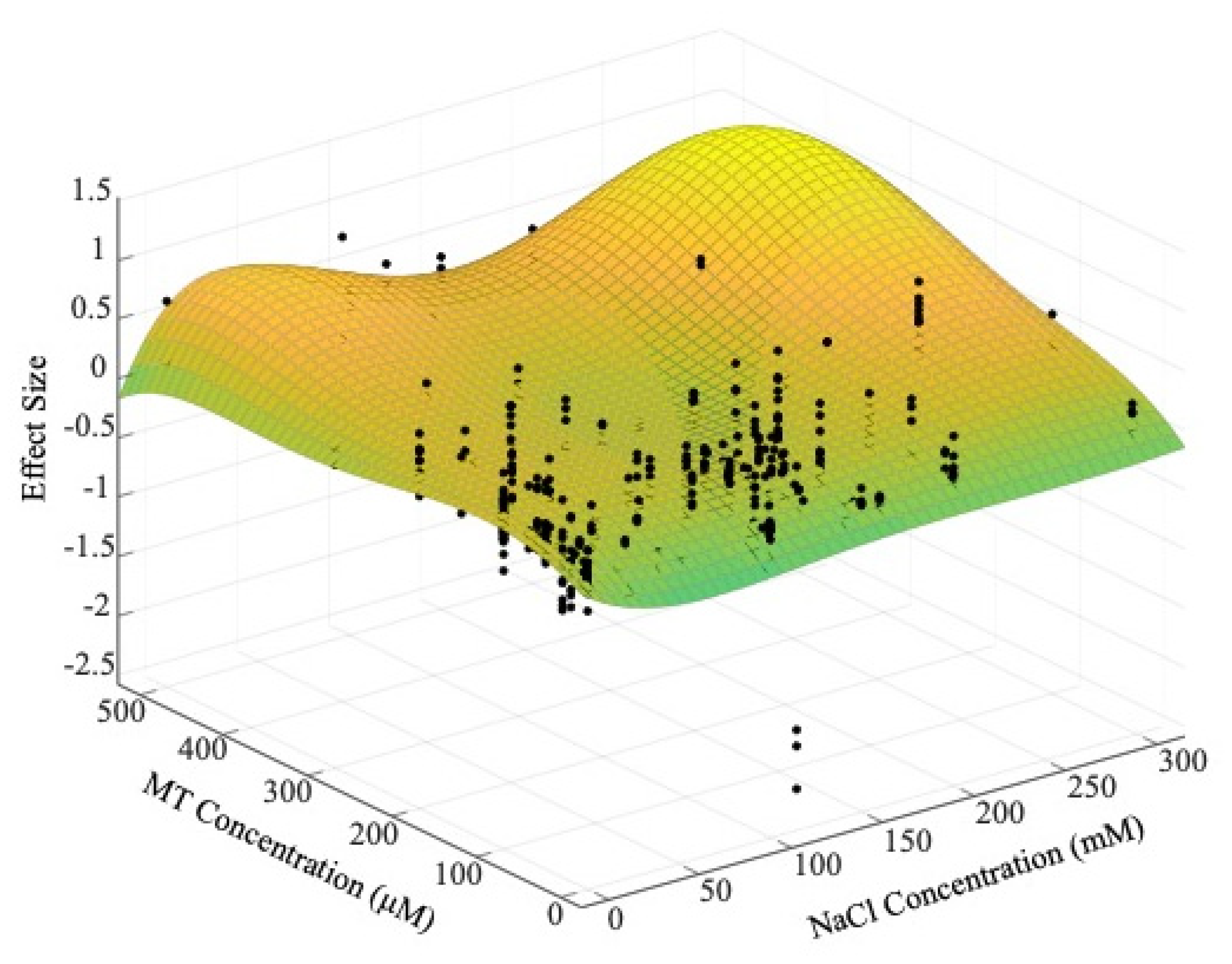
3.5. The Effect of MT Concentration and Application Method
4. Discussion
4.1. Effect of MT on Plant Growth under Salt Stress
4.2. Effects of Salt Stress Characteristics on MT Regulation of Plant Growth under Salt Stress
4.3. Suitable Concentration Range and Application Method of MT in Regulating Plant Growth under Salt Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Number | Plant Species | Plant Sample Planting Medium | Salt Stress Concentration | Melatonin Concentration | Melatonin Treatment | Measurement Index |
|---|---|---|---|---|---|---|
| 1 | Avena nuda | Nutrition liquid | 0; 150 | 50; 100 | Added to the rooting medium | Plant height; dry weight; fresh weight; stem diameter |
| 2 | Avena sativa | Nutrition liquid | 0; 300 | 100 | Added to the rooting medium | Fresh weight; plant height; root length |
| 3 | Brassica napus | Nutrition liquid | 125 | 50 | Added to the rooting medium | Plant height; fresh weight |
| 4 | Brassica napus | Nutrition liquid | 0; 200 | 0.01; 0.1; 1; 10; 100 | Added to the rooting medium | Root fresh weight; root length |
| 5 | Brassica napus | Nutrition liquid | 128 | 30; 45; 60; 75; 100 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height; root length; leaf area |
| 6 | Cucumis melon | Nutrition liquid | 100 | 100 | Added to the rooting medium | Shoot dry weight; root dry weight; plant height |
| 7 | Cucumis sativus | Nutrition liquid | 0; 200 | 100 | Added to the rooting medium | Plant height; leaf area; dry weight; fresh weight |
| 8 | Dracocephalum kotschyi | Soil | 0; 30 | 100; 200 | Added to the rooting medium | Dry weight |
| 9 | Gossypium hirsutum | Nutrition liquid | 150 | 50; 100; 200; 500 | Foliar application | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight |
| 10 | Gossypium hirsutum | Nutrition liquid | 100 | 25; 50; 75 | Seed priming | Root length; leaf area |
| 11 | Gossypium hirsutum | Nutrition liquid | 0; 100 | 0.1; 0.5; 1; 10; 50 | Added to the rooting medium | Plant height; root length; shoot fresh weight; root fresh weight |
| 12 | Gossypium hirsutum | Nutrition liquid | 150 | 20 | Added to the rooting medium | germination percentage; plant height |
| 13 | Gossypium hirsutum | Nutrition liquid | 150 | 10; 20; 50; 100 | Seed priming | germination percentage |
| 14 | Helianthus annuus | Nutrition liquid | 0; 120 | 5; 15; 30; 45 | Added to the rooting medium | fresh weight |
| 15 | Hordeum vulgare | Nutrition liquid | 0; 60 | 20 | Seed soaking | Plant height; fresh weight |
| 16 | Limonium bicolor | Nutrition liquid | 0; 300 | 5 | Added to the rooting medium | Fresh weight; leaf area; number of blades; plant height; root length |
| 17 | Limonium bicolor | Nutrition liquid | 0; 300 | 5 | Added to the rooting medium | Shoot dry weight |
| 18 | Limonium bicolor | Nutrition liquid | 0; 100; 200 | 1; 50; 100; 200; 300; 500 | Seed priming | Germination percentage; germination potential |
| 19 | Malus hupehensis | Nutrition liquid | 0; 200 | 0.1 | Added to the rooting medium | Plant height; dry weight |
| 20 | Medicago sativa | Nutrition liquid | 250 | 150 | Added to the rooting medium | Germination percentage |
| 21 | Medicago sativa | Nutrition liquid | 200 | 1; 10; 50; 100; 500 | Added to the rooting medium | Germination percentage; fresh weight; root length |
| 22 | Oryza sativa | Nutrition liquid | 150 | 50; 100; 200 | Foliar application | Dry weight |
| 23 | Oryza sativa | Nutrition liquid | 150 | 200 | Foliar application | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight |
| 24 | Oryza sativa | Nutrition liquid | 50; 100 | 25; 50; 100; 200; 300; 400 | Foliar application | Dry weight; plant height |
| 25 | Oryza sativa | Nutrition liquid | 0; 150 | 50; 100; 200 | Foliar application | dry weight; fresh weight |
| 26 | Oryza sativa | Nutrition liquid | 120 | 10; 50; 100; 200; 500 | Added to the rooting medium | Germination potential; fresh weight |
| 27 | Oryza sativa | Nutrition liquid | 100 | 5 | Seed soaking | Germination percentage |
| 28 | Phaseolus vulgaris | Nutrition liquid | 0; 68.38 | 100 | Seed soaking | Root length; root surface area |
| 29 | Phaseolus vulgaris | Nutrition liquid | 0; 200 | 150 | Added to the rooting medium | Dry weight; fresh weight |
| 30 | Solanum lycopersicum | Nutrition liquid | 150 | 1; 50; 100; 150; 200 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height |
| 31 | Solanum lycopersicum | Nutrition liquid | 0; 100 | 20; 50 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height; leaf area; root length |
| 32 | Solanum lycopersicum | Nutrition liquid | 0; 120 | 30 | Added to the rooting medium | Plant height; root length |
| 33 | Solanum lycopersicum | Nutrition liquid | 0; 150 | 100 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height |
| 34 | Solanum lycopersicum | Nutrition liquid | 0; 160 | 1 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height; root length |
| 35 | Stevia rebaudiana Bertoni | Nutrition liquid | 25 | 5; 10; 20 | Added to the rooting medium | Dry weight; fresh weight |
| 36 | Triticum aestivum | Nutrition liquid | 0; 100 | 1 | Added to the rooting medium | Shoot dry weight; root dry weight |
| 37 | Triticum aestivum | Soil | 0; 60; 120 | 70 | Foliar application | Shoot dry weight; root dry weight; plant height; root length; leaf area |
| 38 | Zea mays | Nutrition liquid | 0; 100 | 100 | Added to the rooting medium | Plant height; shoot dry weight; root dry weight; shoot fresh weight; root fresh weight |
| 39 | Zea mays | Nutrition liquid | 0; 100 | 1; 2 | Added to the rooting medium | Shoot dry weight; root dry weight; leaf area |
| 40 | Zea mays | Nutrition liquid | 0; 100 | 1 | Added to the rooting medium | Shoot dry weight; root dry weight; |
| 41 | Zea mays | Soil | 0; 25; 50; 80 | 50; 100; 200 | Seed soaking | Plant height; shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; root length; stem diameter |
| 42 | Zea mays | Soil | 0; 105 | 30; 60; 90 | Added to the rooting medium | Shoot dry weight; root dry weight; shoot fresh weight; root fresh weight; plant height; leaf area; root length; stem diameter; root diameter |
| Std-Eff | Coefficient | Std.Err. | t | P > |t| | 95% Conf. Intervall | |
|---|---|---|---|---|---|---|
| Slope | 0.0728124 | 0.012001 | 6.07 | 0.000 | 0.0492328 | 0.096392 |
| Bias | 3.420367 | 0.08347499 | 4.10 | 0.000 | 1.780659 | 5.060074 |
| Studies | Effect Size | 95% Conf. Interval | |
|---|---|---|---|
| Observed | 0.178 | 0.154 | 0.202 |
| Observed + Imputed | 0.255 | 0.228 | 0.281 |
References
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Reddy, I.N.B.L.; Kim, B.-K.; Yoon, I.-S.; Kim, K.-H.; Kwon, T.-R. Salt Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; de Sousa, L. Global mapping of soil salinity change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Dietz, K.-J. Tuning of Redox Regulatory Mechanisms, Reactive Oxygen Species and Redox Homeostasis under Salinity Stress. Front. Plant Sci. 2016, 7, 548. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.H.; Foster, K.J.; et al. Energy costs of salt tolerance in crop plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2020, 225, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Lu, B.; Liu, L.T.; Duan, W.J.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.C.; Zhang, Y.J.; Li, C.D.; et al. Melatonin promotes seed germination under salt stress by regulating ABA and GA3 in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2021, 162, 506–516. [Google Scholar] [CrossRef]
- Khan, R.W.A.; Awan, F.S.; Iqbal, R.K. Evaluation and inditification of salt tolerance wheat through in vitro salinitu induction in seeds. Pak. J. Bot. 2022, 54, 1987–1993. [Google Scholar] [CrossRef]
- Nguyen, V.Q.; Sreewongchai, T.; Siangliw, M.; Roytrakul, S.; Yokthongwattana, C. Comparative proteomic analysis of chromosome segment substitution lines of Thai jasmine rice KDML105 under short-term salinity stress. Planta 2022, 256, 12. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.Y.; Dong, J.X.; Song, X.; Jiang, J.; Li, H.Y. Overexpression of Tamarix hispida ThTrx5 Confers Salt Tolerance to Arabidopsis by Activating Stress Response Signals. Int. J. Mol. Sci. 2020, 21, 1165. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, C.; Hussain, S.; Huang, J.; Liang, Q.; Zhu, L.; Cao, X.; Kong, Y.; Li, Y.; Wang, L.; et al. Effects of nitric oxide on nitrogen metabolism and the salt resistance of rice (Oryza sativa L.) seedlings with different salt tolerances. Plant Physiol. Biochem. 2020, 155, 374–383. [Google Scholar] [CrossRef]
- Zushi, K.; Higashijima, M. Correlation network analysis identified the key interactions of texture profiles with the sensory, physical, and organoleptic properties of cherry tomato cultivars grown under salt stress. Sci. Hortic. 2022, 293, 110754. [Google Scholar] [CrossRef]
- Al Rubaye, O.A.M.; Yetisir, H.; Ulas, F.; Ulas, A. Enhancing Salt Stress Tolerance of Different Pepper (Capsicum annuum L.) Inbred Line Genotypes by Rootstock with Vigorous Root System. Gesunde Pflanz. 2021, 73, 375–389. [Google Scholar] [CrossRef]
- Hussain, S.; Zhang, J.-H.; Zhong, C.; Zhu, L.-F.; Cao, X.-C.; Yu, S.-M.; Allen Bohr, J.; Hu, J.-J.; Jin, Q.-Y. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 4, 96. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Zheng, Y.; Tong, C.L.; Zhang, D.J. Effects of exogenous melatonin on plant growth, root hormones and photosynthetic characteristics of trifoliate orange subjected to salt stress. Plant Growth Regul. 2022, 97, 551–558. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin in flowering, fruit set and fruit ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef] [PubMed]
- García, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.-X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin Against Environmental Plant Stressors: A Review. Curr. Protein Pept. Sci. 2021, 22, 413–429. [Google Scholar] [CrossRef]
- Li, J.P.; Liu, J.; Zhu, T.T.; Zhao, C.; Li, L.Y.; Chen, M. The Role of Melatonin in Salt Stress Responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef]
- Fan, J.B.; Xie, Y.; Zhang, Z.C.; Chen, L. Melatonin: A Multifunctional Factor in Plants. Int. J. Mol. Sci. 2018, 19, 1528. [Google Scholar] [CrossRef]
- Shen, J.; Chen, D.D.; Zhang, X.P.; Song, L.R.; Dong, J.; Xu, Q.J.; Hu, M.J.; Cheng, Y.Y.; Shen, F.F.; Wang, W. Mitigation of salt stress response in upland cotton (Gossypium hirsutum) by exogenous melatonin. J. Plant Res. 2021, 134, 857–871. [Google Scholar] [CrossRef]
- Gu, Q.; Xiao, Q.Q.; Chen, Z.P.; Han, Y. Crosstalk between Melatonin and Reactive Oxygen Species in Plant Abiotic Stress Responses: An Update. Int. J. Mol. Sci. 2022, 23, 5666. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Gao, H.; Lu, M.X.; Hao, C.Y.; Pu, Z.Q.; Guo, M.J.; Hou, D.R.; Chen, L.Y.; Huang, X. Melatonin-Nitric Oxide Crosstalk and Their Roles in the Redox Network in Plants. Int. J. Mol. Sci. 2019, 20, 6200. [Google Scholar] [CrossRef]
- Zeng, W.; Mostafa, S.; Lu, Z.G.; Jin, B. Melatonin-Mediated Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2022, 13, 847175. [Google Scholar] [CrossRef] [PubMed]
- Madigan, A.P.; Egidi, E.; Bedon, F.; Franks, A.E.; Plummer, K.M. Bacterial and Fungal Communities Are Differentially Modified by Melatonin in Agricultural Soils Under Abiotic Stress. Front. Microbiol. 2019, 10, 2616. [Google Scholar] [CrossRef]
- Liang, C.Z.; Zheng, G.Y.; Li, W.Z.; Wang, Y.Q.; Hu, B.; Wang, H.R.; Wu, H.K.; Qian, Y.W.; Zhu, X.G.; Tan, D.X.; et al. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Zhang, T.G.; Shi, Z.F.; Zhang, X.H.; Zheng, S.; Wang, J.; Mo, J.N. Alleviating effects of exogenous melatonin on salt stress in cucumber. Sci. Hortic. 2020, 262, 109070. [Google Scholar] [CrossRef]
- Zhao, G.; Zhao, Y.Y.; Yu, X.L.; Kiprotich, F.; Han, H.; Guan, R.Z.; Wang, R.; Shen, W.B. Nitric Oxide Is Required for Melatonin-Enhanced Tolerance against Salinity Stress in Rapeseed (Brassica napus L.) Seedlings. Int. J. Mol. Sci. 2018, 19, 1912. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.Q.; Zhang, H.J.; Cao, Y.Y.; Weeda, S.; Ren, S.X.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Wei, L.; Zhao, H.Y.; Wang, B.X.; Wu, X.Y.; Lan, R.J.; Huang, X.; Chen, B.; Chen, G.; Jiang, C.Q.; Wang, J.L.; et al. Exogenous Melatonin Improves the Growth of Rice Seedlings by Regulating Redox Balance and Ion Homeostasis Under Salt Stress. J. Plant Growth Regul. 2021, 41, 2108–2121. [Google Scholar] [CrossRef]
- Li, J.P.; Zhao, C.; Zhang, M.J.; Yuan, F.; Chen, M. Exogenous melatonin improves seed germination in Limonium bicolor under salt stress. Plant Signal. Behav. 2019, 14, 1659705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.J.; Sun, Q.Q.; Cao, Y.Y.; Li, X.S.; Zhao, B.; Wu, P.; Guo, Y.D. Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production. Sci. Rep. 2017, 7, 503. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Liu, J.L.; Wang, W.X.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Liu, L.Y.; Li, H.Y.; Zhu, S.H.; Gao, Y.; Zheng, X.Q.; Xu, Y. The response of agronomic characters and rice yield to organic fertilization in subtropical China: A three-level meta-analysis. Field Crops Res. 2021, 263, 108049. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Burda, B.U.; O’Connor, E.A.; Webber, E.M.; Redmond, N.; Perdue, L.A. Estimating data from figures with a Web-based program: Considerations for a systematic review. Res Synth Methods 2017, 8, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Zhou, B.; Xu, J.; Ioannou, A.; Feng, Z.; Saitanis, C.J.; Frei, M.; Calabrese, E.J.; Fotopoulos, V. Exogenous application of melatonin to plants, algae, and harvested products to sustain agricultural productivity and enhance nutritional and nutraceutical value: A meta-analysis. Environ. Res. 2021, 200, 111746. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought effect on plant biomass allocation: A meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef]
- Wang, L.L.; Li, Q.; Coulter, J.A.; Xie, J.H.; Luo, Z.Z.; Zhang, R.Z.; Deng, X.P.; Li, L.L. Winter wheat yield and water use efficiency response to organic fertilization in northern China: A meta-analysis. Agric. Water Manag. 2020, 229, 105934. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Mosaad, I.S.M.; Al-Ghamdi, A.A.; Abbasi, A.M.; Zhou, X.-B. Melatonin Application Alleviates Stress-Induced Photosynthetic Inhibition and Oxidative Damage by Regulating Antioxidant Defense System of Maize: A Meta-Analysis. Antioxidants 2022, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Deng, Q.; Duan, H.; Guo, Y. Effects of biochar application on root traits: A meta-analysis. GCB Bioenergy 2017, 9, 1563–1572. [Google Scholar] [CrossRef]
- Ke, Q.B.; Ye, J.; Wang, B.M.; Ren, J.H.; Yin, L.N.; Deng, X.P.; Wang, S.W. Melatonin Mitigates Salt Stress in Wheat Seedlings by Modulating Polyamine Metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.M.; Liu, M.; Zhou, R.; Jiang, F.L.; Sun, M.T.; Wen, J.Q.; Zhu, Z.H.; Wu, Z. Relationship between melatonin and abscisic acid in response to salt stress of tomato. Sci. Hortic. 2021, 285, 110176. [Google Scholar] [CrossRef]
- Zhao, B.Q.; Liu, Q.Y.; Wang, B.S.; Yuan, F. Roles of Phytohormones and Their Signaling Pathways in Leaf Development and Stress Responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef]
- Läuchli, A.; Epstein, E. Plant responses to saline and sodic conditions. In Agricultural Salinity Assessment and Management; Wallendar, W.W., Tanji, K.K., Eds.; American Society of Civil Engineers: New York, NY, USA, 1990; Volume 71, pp. 113–137. [Google Scholar]
- Chen, J.; Li, H.; Yang, K.; Wang, Y.Z.; Yang, L.F.; Hu, L.B.; Liu, R.X.; Shi, Z.Q. Melatonin facilitates lateral root development by coordinating PAO-derived hydrogen peroxide and Rboh-derived superoxide radical. Free Radic. Biol. Med. 2019, 143, 534–544. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Altaf, M.M.; Khan, L.U.; Shahid, S.; Jahan, M.S. Melatonin alleviates salt damage in tomato seedling: A root architecture system, photosynthetic capacity, ion homeostasis, and antioxidant enzymes analysis. Sci. Hortic. 2021, 285, 110145. [Google Scholar] [CrossRef]
- Rewald, B.; Rachmilevitch, S.; Ephrath, J.E. Salt Stress Effects on Root Systems of Two Mature Olive Cultivars. In Proceedings of the International Symposium on Olive Irrigation and Oil Quality, Nazareth, Israel, 6–10 December 2009; pp. 109–118. [Google Scholar]
- Xu, C.; Ling, F.-L.; Xu, K.-Z.; Wu, Z.-H.; Liu, X.-L.; An, J.-H.; Zhao, L.-P. Effect of Salt Stress on Photosynthetic Characteristics and Physiological and Biochemical Traits of Different Rice Varieties. Chin. J. Rice Sci. 2013, 27, 280–286. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of Salt Tolerant PGPR in Growth and Yield Augmentation of Wheat (Triticum aestivum L.) Under Saline Conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef]
- Hu, D.D.; Dong, S.T.; Zhang, J.W.; Zhao, B.; Ren, B.Z.; Liu, P. Endogenous hormones improve the salt tolerance of maize (Zea mays L.) by inducing root architecture and ion balance optimizations. J. Agron. Crop Sci. 2022, 1–13. [Google Scholar] [CrossRef]
- Pelagio-Flores, R.; Munoz-Parra, E.; Ortiz-Castro, R.; Lopez-Bucio, J. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 2012, 53, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol. 2019, 19, 414. [Google Scholar] [CrossRef]
- Yan, F.Y.; Wei, H.M.; Li, W.W.; Liu, Z.H.; Tang, S.; Chen, L.; Ding, C.Q.; Jiang, Y.; Ding, Y.F.; Li, G.H. Melatonin improves K+ and Na+ homeostasis in rice under salt stress by mediated nitric oxide. Ecotoxicol. Environ. Saf. 2020, 206, 111358. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wei, H.; Ding, Y.; Li, W.; Liu, Z.; Chen, L.; Tang, S.; Ding, C.; Jiang, Y.; Li, G. Melatonin regulates antioxidant strategy in response to continuous salt stress in rice seedlings. Plant Physiol. Biochem. 2021, 165, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.Q.; Gao, Y.M.; Li, M.; Du, Y.L.; Zhang, Y.X.; Zhang, W.H.; Du, J.D. The molecular events underpinning cultivar differences in melatonin counteracting salt damage in Phaseolus vulgaris. Funct. Plant Biol. 2022, 49, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Jalili, S.; Ehsanpour, A.A.; Javadirad, S.M. The role of melatonin on caspase-3-like activity and expression of the genes involved in programmed cell death (PCD) induced by in vitro salt stress in alfalfa (Medicago sativa L.) roots. Bot. Stud. 2022, 63, 19. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bhatla, S.C. Exogenous Melatonin Modulates Endogenous H2S Homeostasis and L-Cysteine Desulfhydrase Activity in Salt-Stressed Tomato (Solanum lycopersicum L. var. cherry) Seedling Cotyledons. J. Plant Growth Regul. 2021, 40, 2502–2514. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Pre-sowing seed treatment—A shotgun approach to improve germination, plant growth, and crop yield under saline and non-saline conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; da Silva, E.C. Physiology and Biochemistry of Salt Stress Tolerance in Plants. In Abiotic Stresses in Crop Plants; Chakraborty, U., Chakraborty, B., Eds.; CABI: Wallingford, UK, 2015; pp. 81–101. [Google Scholar]
- Kamiab, F. Exogenous melatonin mitigates the salinity damages and improves the growth of pistachio under salinity stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Abadia, J.; Marjani, M. Melatonin foliar sprays elicit salinity stress tolerance and enhance fruit yield and quality in strawberry (Fragaria × ananassa Duch.). Plant Physiol. Biochem. 2020, 149, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Dolferus, R. To grow or not to grow: A stressful decision for plants. Plant Sci. 2014, 229, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Dsouza, A.; Price, G.W.; Dixon, M.; Graham, T. A Conceptual Framework for Incorporation of Composting in Closed-Loop Urban Controlled Environment Agriculture. Sustainability 2021, 13, 2471. [Google Scholar] [CrossRef]
- Moreno, J.E.; Campos, M.L. Waking up for defense! Melatonin as a regulator of stomatal immunity in plants. Plant Physiol. 2022, 188, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Xie, C.; Zhang, H.Q.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P.; et al. Melatonin and Its Effects on Plant Systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef]
- Park, W.J. Melatonin as an Endogenous Plant Regulatory Signal: Debates and Perspectives. J. Plant Biol. 2011, 54, 143–149. [Google Scholar] [CrossRef]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.T.; Lu, X.C.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D.L. Melatonin Mediates Enhancement of Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef]
- Rehaman, A.; Mishra, A.K.; Ferdose, A.; Per, T.S.; Hanief, M.; Jan, A.T.; Asgher, M. Melatonin in Plant Defense against Abiotic Stress. Forests 2021, 12, 1404. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.H.; Rizwan, M.; Fahad, S.; Xu, Z.H.; Hu, L.Y. Seed priming with melatonin coping drought stress in rapeseed by regulating reactive oxygen species detoxification: Antioxidant defense system, osmotic adjustment, stomatal traits and chloroplast ultrastructure perseveration. Ind. Crops Prod. 2019, 140, 111597. [Google Scholar] [CrossRef]
- Kolodziejczyk, I.; Dzitko, K.; Szewczyk, R.; Posmyk, M.M. Exogenous melatonin improves corn (Zea mays L.) embryo proteome in seeds subjected to chilling stress. J. Plant Physiol. 2016, 193, 47–56. [Google Scholar] [CrossRef]
- Jiang, D.; Lu, B.; Liu, L.T.; Duan, W.J.; Chen, L.; Li, J.; Zhang, K.; Sun, H.C.; Zhang, Y.J.; Dong, H.Z.; et al. Exogenous melatonin improves salt stress adaptation of cotton seedlings by regulating active oxygen metabolism. PeerJ 2020, 8, e10486. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F. Effect of seed priming on horticultural crops. Sci. Hortic. 2021, 286, 1101970. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.J.; Dong, Y.T.; Zhang, F.; He, Q.L.; Chen, J.H.; Zhu, S.J.; Zhao, T.L. Seed priming with melatonin improves salt tolerance in cotton through regulating photosynthesis, scavenging reactive oxygen species and coordinating with phytohormone signal pathways. Ind. Crops Prod. 2021, 169, 113671. [Google Scholar] [CrossRef]
- Rajora, N.; Vats, S.; Raturi, G.; Thakral, V.; Kaur, S.; Rachappanavar, V.; Kumar, M.; Kesarwani, A.K.; Sonah, H.; Sharma, T.R.; et al. Seed priming with melatonin: A promising approach to combat abiotic stress in plants. Plant Stress 2022, 4, 100071. [Google Scholar] [CrossRef]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of Melatonin on Anti-oxidative Systems and Photosystem II in Cold-Stressed Rice Seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]






| Influence Factors | Classification of Subgroup | |
|---|---|---|
| Plant properties | Plant species | Avena nuda (naked oat), Avena sativa (oat), Brassica napus (rapeseed), Cucumis melon (melon), Cucumis sativus (cucumber), Dracocephalum kotschyi (Savannah obedient-plant), Gossypium hirsutum (cotton), Helianthus annuus (sunflower), Hordeum vulgare (barley), Limonium bicolor (statice bicolor), Malus hupehensis (tea crabapple), Medicago sativa (alfalfa), Oryza sativa (rice), Phaseolus vulgaris (common bean), Solanum lycopersicum (tomato), Stevia rebaudiana Bertoni (candyleaf), Triticum aestivum (wheat), Zea mays (maize) |
| Growth parameters | Stem diameter, Root length, Root diameter, Root surface area, Root fresh weight, Root dry weight, Plant height, Number of blades, Leaf area, Shoot fresh weight, Shoot dry weight, Germination potential, Germination percentage, Fresh weight, Dry weight. | |
| Types of growth parameters | Whole plant index, Germination index, Stem index, Root index. | |
| Salt stress properties | Salt stress or not | Salt stress, Normal condition |
| Salt stress (mM NaCl) concentration | 0 = NaCl, 0 < NaCl < 50, 50 ≤ NaCl < 100, 100 ≤ NaCl < 150, 150 ≤ NaCl < 200, 200 ≤ NaCl < 250, 250 ≤ NaCl < 300, 300 ≤ NaCl. | |
| Ways of applying salt stress (rooting environment) | Rooting medium, Soil. | |
| Meltonin properties | MT concentration (μM) | MT ≤ 0.1, 0.1 < MT ≤ 1, 1 < MT ≤ 10, 0 < MT ≤ 50, 50 < MT ≤ 100, 100 < MT ≤ 200, 200 < MT ≤ 300, 300 < MT ≤ 400, 400 < MT ≤ 500. |
| Mode of exogenous application | Rooting medium, Foliar application, Seed priming, Seed soaking |
| Categorical Variable | QB | df | p-Value |
|---|---|---|---|
| Plants | 54.83 | 17 | 0.000 |
| Salt stress or not | 0.40 | 1 | 0.526 |
| Type of rooting environment | 12.48 | 1 | 0.000 |
| Salt concentration | 72.62 | 7 | 0.000 |
| MT concentration | 335.08 | 8 | 0.000 |
| MT treatment type | 74.42 | 3 | 0.000 |
| Growth parameters | 56.00 | 14 | 0.000 |
| Types of growth parameters | 7.15 | 3 | 0.067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, F.; Zhao, H.; Wu, L.; Huang, Z.; Niu, Y.; Qi, B.; Zhang, L.; Fan, S.; Ding, Y.; Li, G.; et al. Basic Cognition of Melatonin Regulation of Plant Growth under Salt Stress: A Meta-Analysis. Antioxidants 2022, 11, 1610. https://doi.org/10.3390/antiox11081610
Yan F, Zhao H, Wu L, Huang Z, Niu Y, Qi B, Zhang L, Fan S, Ding Y, Li G, et al. Basic Cognition of Melatonin Regulation of Plant Growth under Salt Stress: A Meta-Analysis. Antioxidants. 2022; 11(8):1610. https://doi.org/10.3390/antiox11081610
Chicago/Turabian StyleYan, Feiyu, Hongliang Zhao, Longmei Wu, Zhiwei Huang, Yuan Niu, Bo Qi, Linqing Zhang, Song Fan, Yanfeng Ding, Ganghua Li, and et al. 2022. "Basic Cognition of Melatonin Regulation of Plant Growth under Salt Stress: A Meta-Analysis" Antioxidants 11, no. 8: 1610. https://doi.org/10.3390/antiox11081610