
Purification, Identification, and Properties of a Novel Carotenoid Produced by Arthrobacter sp. QL17 Isolated from Mount Qomolangma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Pigment-Producing Bacteria
2.2. Hydrogen Peroxide (H2O2) Tolerance Assay
2.3. Purification and Structural Identification of the Bioactive Compound
2.4. Assessment of Radical Scavenging Activity
2.5. In vitro Cytotoxic Assay
2.6. Data Processing and Statistical Analysis
3. Results and Discussion
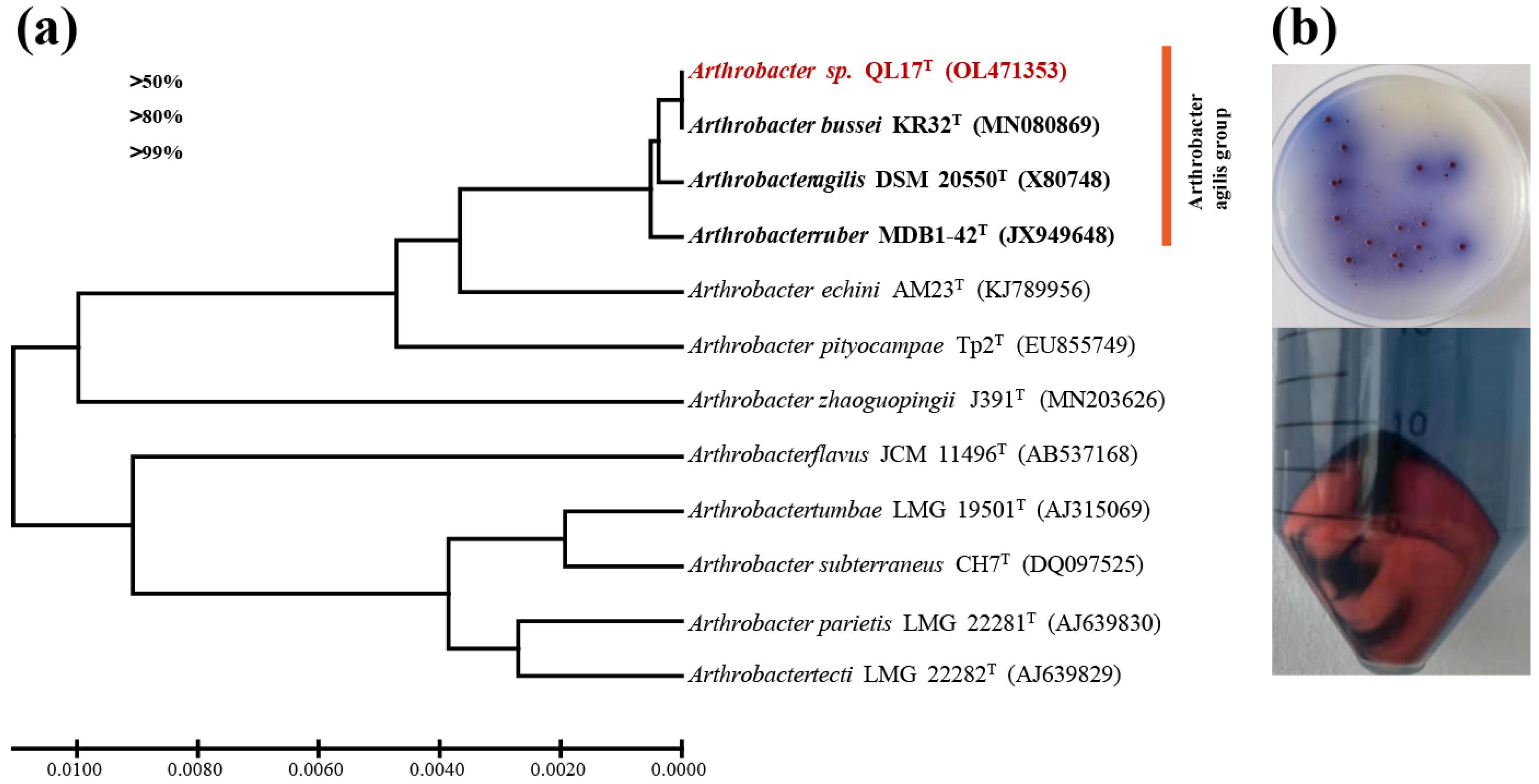
3.1. Growth and Characterization of Arthrobacter sp. QL17
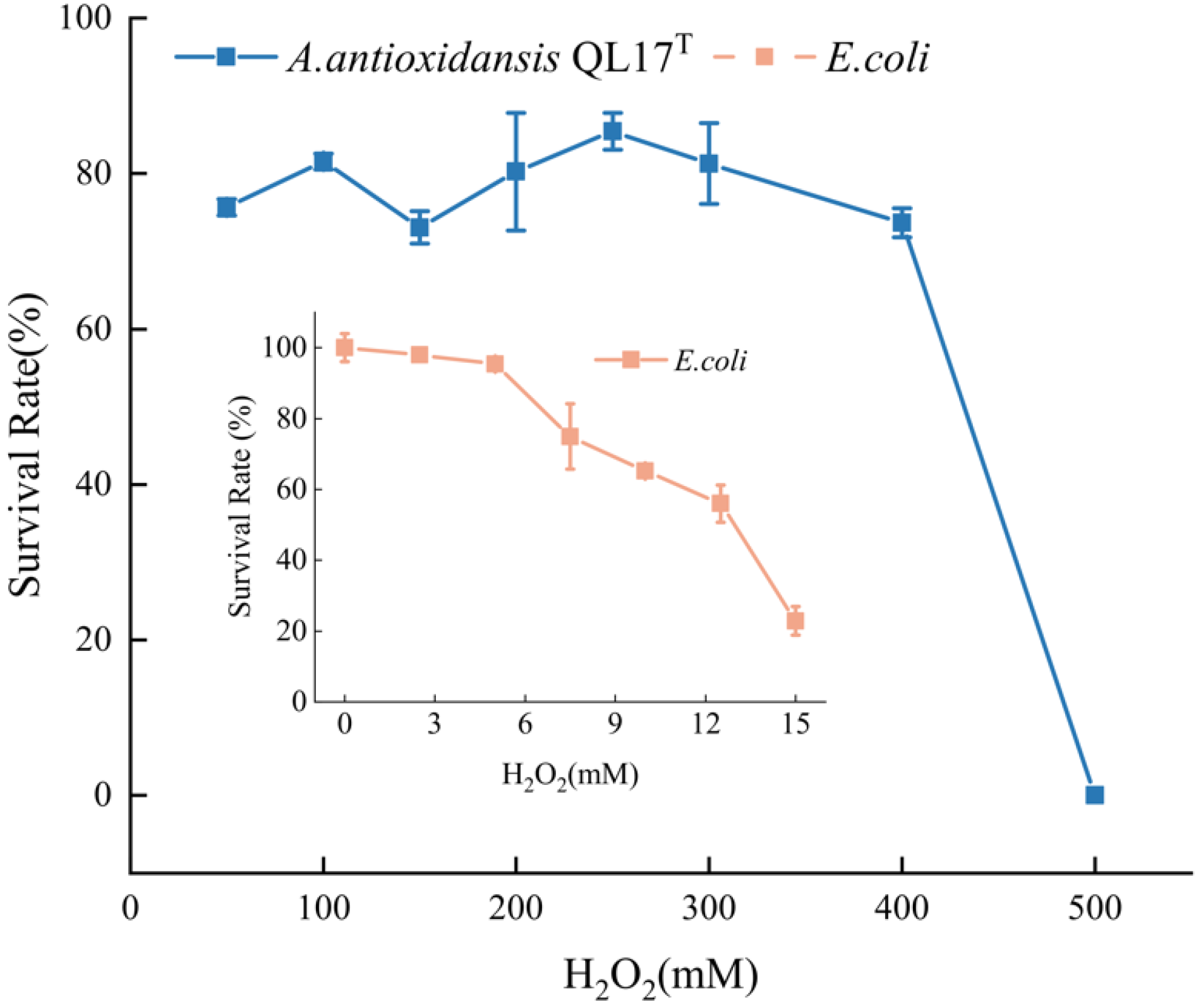
3.2. Hydrogen Peroxide (H2O2) Stress Tolerance of A. sp. QL17
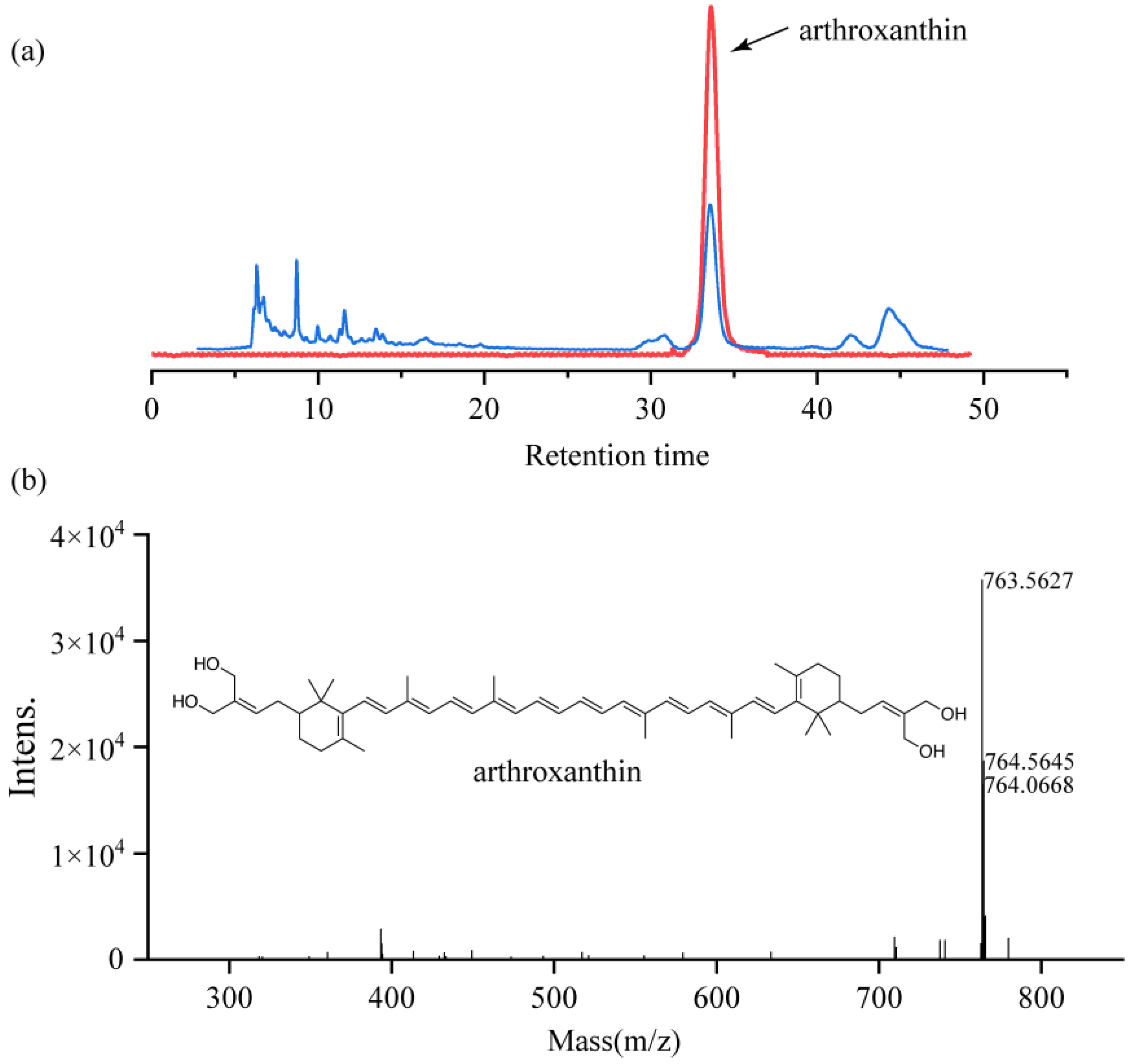
3.3. Purification and Structural Identification of Antioxidants Produced by Strain QL17
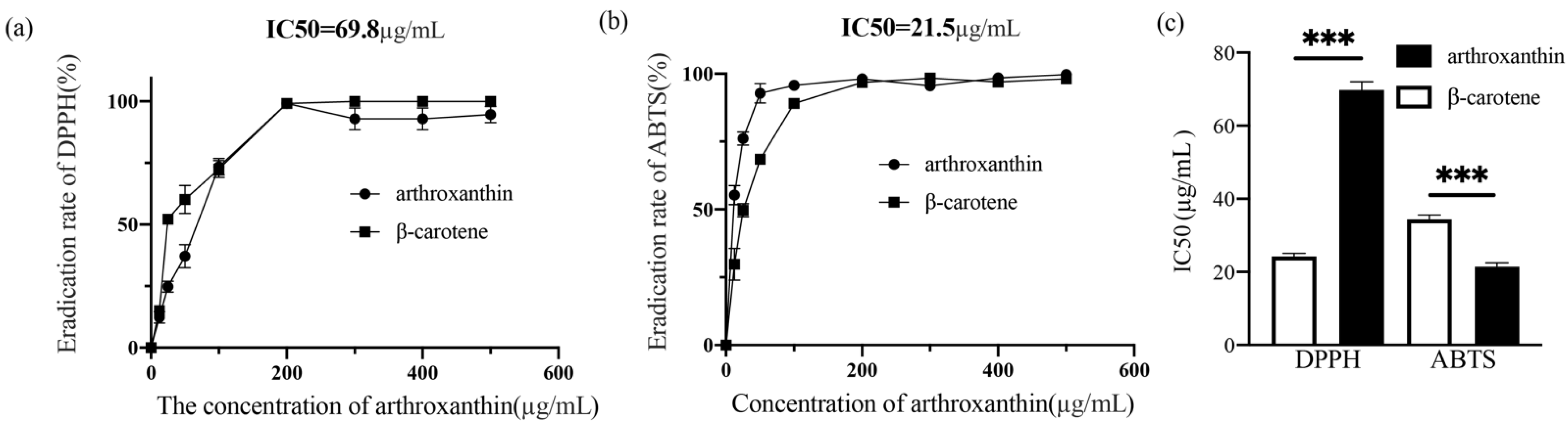
3.4. The Radical-Scavenging Activity of Arthroxanthin
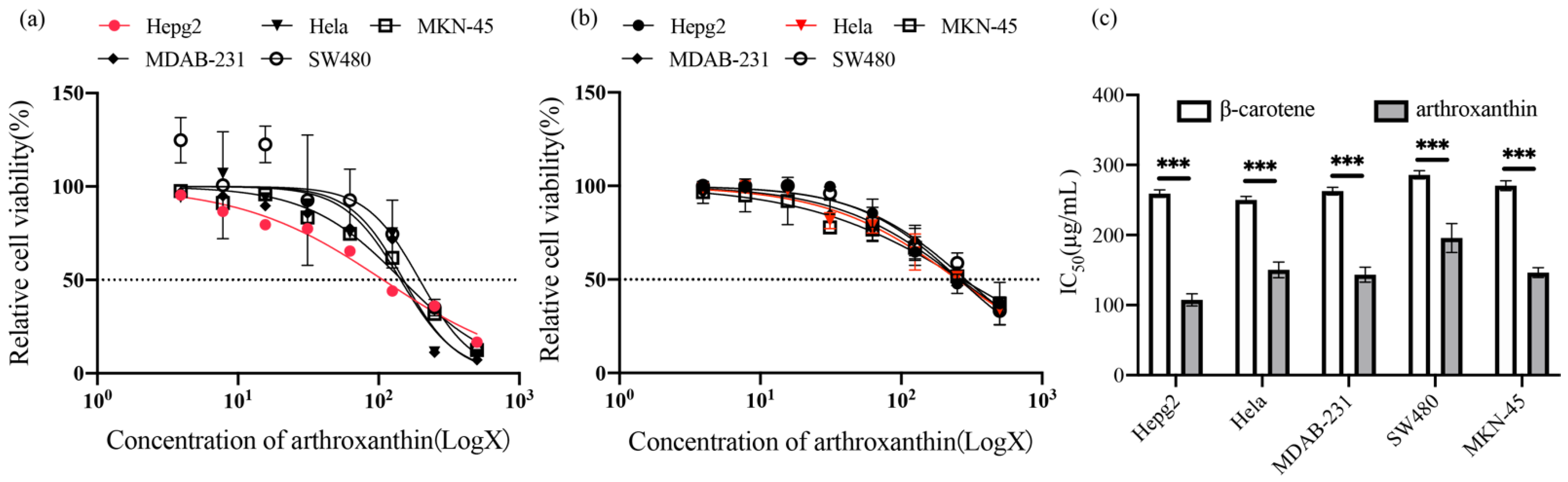
3.5. Cytotoxic Activities of Arthroxanthin
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, M.P.N.; Xiao, M.; Li, W.J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef]
- Phadwal, K. Carotenoid biosynthetic pathway: Molecular phylogenies and evolutionary behavior of crt genes in eubacteria. Gene 2005, 345, 35–43. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [Green Version]
- Rezaeeyan, Z.; Safarpour, A.; Amoozegar, M.A.; Babavalian, H.; Tebyanian, H.; Shakeri, F. High carotenoid production by a halotolerant bacterium, Kocuria sp. strain QWT-12 and anticancer activity of its carotenoid. Excli. J. 2017, 16, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In vitro dual (anticancer and antiviral) activity of the carotenoids produced by haloalkaliphilic archaeon Natrialba sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef] [PubMed]
- Sutthiwong, N.; Fouillaud, M.; Valla, A.; Caro, Y.; Dufossé, L. Bacteria belonging to the extremely versatile genus Arthrobacter as novel source of natural pigments with extended hue range. Food Res. Int. 2014, 65, 156–162. [Google Scholar] [CrossRef]
- Reddy, G.S.; Aggarwal, R.K.; Matsumoto, G.I.; Shivaji, S. Arthrobacter flavus sp. nov., a psychrophilic bacterium isolated from a pond in McMurdo Dry Valley, Antarctica. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 4, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Galaup, P.; Gautier, A.; Piriou, Y.; Villeblanche, A.D.; Valla, A.; Dufossé, L. First pigment fingerprints from the rind of French PDO red-smear ripened soft cheeses Epoisses, Mont d’Or and Maroilles. Innov. Food Sci. Emerg. Technol. 2007, 8, 373–378. [Google Scholar] [CrossRef]
- Silva, T.R.; Tavares, R.S.N.; Canela-Garayoa, R.; Eras, J.; Rodrigues, M.V.N.; Neri-Numa, I.A.; Pastore, G.M.; Rosa, L.H.; Schultz, J.A.A.; Debonsi, H.M.; et al. Chemical Characterization and Biotechnological Applicability of Pigments Isolated from Antarctic Bacteria. Mar. Biotechnol. 2019, 21, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Flegler, A.; Runzheimer, K.; Kombeitz, V.; Mänz, A.T.; Heidler von Heilborn, D.; Etzbach, L.; Schieber, A.; Hölzl, G.; Hüttel, B.; Woehle, C.; et al. Arthrobacter bussei sp. nov., a pink-coloured organism isolated from cheese made of cow’s milk. Int. J. Syst. Evol. Microbiol. 2020, 70, 3027–3036. [Google Scholar] [CrossRef]
- Fong, N.J.; Burgess, M.L.; Barrow, K.D.; Glenn, D.R. Carotenoid accumulation in the psychrotrophic bacterium Arthrobacter agilis in response to thermal and salt stress. Appl. Microbiol. Biotechnol. 2001, 56, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid pigmentation in Antarctic heterotrophic bacteria as a strategy to withstand environmental stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Jagannadham, M.V.; Chattopadhyay, M.K.; Subbalakshmi, C.; Vairamani, M.; Narayanan, K.; Rao, C.M.; Shivaji, S. Carotenoids of an Antarctic psychrotolerant bacterium, Sphingobacterium antarcticus, and a mesophilic bacterium, Sphingobacterium multivorum. Arch. Microbiol. 2000, 173, 418–424. [Google Scholar] [CrossRef]
- Malvick, D.; Syverson, R.; Mollov, D.; Ishimaru, C.A. Goss’s Bacterial Blight and Wilt of Corn Caused by Clavibacter michiganensis subsp. nebraskensis Occurs in Minnesota. Plant Dis. 2010, 94, 1064. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Li, T.X.; Liu, R.H.; Wang, X.B.; Luo, J.; Luo, J.G.; Kong, L.Y.; Yang, M.H. Hypoxia-Protective Azaphilone Adducts from Peyronellaea glomerata. J. Nat. Prod. 2018, 81, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Shang, X.; Yang, L.; Zheng, S.; Liu, K.; Li, X. Purification, identification and properties of a new blue pigment produced from Streptomyces sp. A1013Y. Food Chem. 2020, 308, 125600. [Google Scholar] [CrossRef]
- Lee, C.C.; Houghton, P. Cytotoxicity of plants from Malaysia and Thailand used traditionally to treat cancer. J. Ethnopharmacol. 2005, 100, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.L. Carotenoids-Antioxidant Properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Tobias, A.V.; Arnold, F.H. Biosynthesis of novel carotenoid families based on unnatural carbon backbones: A model for diversification of natural product pathways. Biochim. Biophys. Acta 2006, 1761, 235–246. [Google Scholar] [CrossRef]
- Mojib, N.; Farhoomand, A.; Andersen, D.T.; Bej, A.K. UV and cold tolerance of a pigment-producing Antarctic Janthinobacterium sp. Ant5-2. Extremophiles 2013, 17, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xue, F.; Yu, X. GC-MS, FTIR and Raman Analysis of Antioxidant Components of Red Pigments from Stemphylium lycopersici. Curr. Microbiol. 2017, 74, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R. Mechanisms of genomic and non-genomic actions of carotenoids. Biochim. Biophys. Acta 2005, 1740, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Bonete, M.J.; León, R.; Kharroub, K. Characterization and biological activities of carotenoids produced by three haloarchaeal strains isolated from Algerian salt lakes. Arch. Microbiol. 2021, 204, 6. [Google Scholar] [CrossRef] [PubMed]
- Al-Dabbagh, B.; Elhaty, I.A.; Elhaw, M.; Murali, C.; Al Mansoori, A.; Awad, B.; Amin, A. Antioxidant and anticancer activities of chamomile (Matricaria recutita L.). BMC Res. Notes 2019, 12, 3. [Google Scholar] [CrossRef]
- Jayappriyan, K.R.; Rajkumar, R.; Venkatakrishnan, V.; Nagaraj, S.; Rengasamy, R. In vitro anticancer activity of natural β-carotene from Dunaliella salina EU5891199 in PC-3 cells. Biomed. Prev. Nutr. 2013, 3, 99–105. [Google Scholar] [CrossRef]
- Velmurugan, B.; Mani, A.; Nagini, S. Combination of S-allylcysteine and lycopene induces apoptosis Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Boneteby modulating Bcl-2, Bax, Bim and caspases during experimental gastric carcinogenesis. Eur. J. Cancer Prev. 2005, 14, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Seren, S.; Lieberman, R.; Bayraktar, U.D.; Heath, E.; Sahin, K.; Andic, F.; Kucuk, O. Lycopene in cancer prevention and treatment. Am. J. Ther. 2008, 15, 66–81. [Google Scholar] [CrossRef]
- Kotake-Nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids affect proliferation of human prostate cancer cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef]
- Faraone, I.; Sinisgalli, C.; Ostuni, A.; Armentano, M.F.; Carmosino, M.; Milella, L.; Russo, D.; Labanca, F.; Khan, H. Astaxanthin anticancer effects are mediated through multiple molecular mechanisms: A systematic review. Pharmacol. Res. 2020, 155, 104689. [Google Scholar] [CrossRef]
- Leone, A.; Zefferino, R.; Longo, C.; Leo, L.; Zacheo, G. Supercritical CO(2)-extracted tomato Oleoresins enhance gap junction intercellular communications and recover from mercury chloride inhibition in keratinocytes. J. Agric. Food Chem. 2010, 58, 4769–4778. [Google Scholar] [CrossRef] [PubMed]
- Udayawara Rudresh, D.; Maradagi, T.; Stephen, N.M.; Niraikulam, A.; Nambi Ramudu, K.; Ponesakki, G. Neoxanthin prevents H(2)O(2)-induced cytotoxicity in HepG2 cells by activating endogenous antioxidant signals and suppressing apoptosis signals. Mol. Biol. Rep. 2021, 48, 6923–6934. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH | δC | Position | δH | δC |
|---|---|---|---|---|---|
| 1,1′ | 3.62 * | 71.2,72.9 | 14,14′ | 6.68 | 131.6 |
| 2,2′ | 138.9 | 15,15′ | 6.68 | 129.6 | |
| 3,3′ | 5.48 | 124.6 | 16,16′ | 138.3 | |
| 4,4′ | 1.92 | 27.2 | 17,17′ | 6.41 | 125.3 |
| 5,5′ | 1.77 | 42.7 | 18,18′ | 6.68 | 133.1 |
| 6,6′ | 1.55 | 29.7 | 19,19′ | 6.68 | 130.5 |
| 7,7′ | 1.99 | 29.9 | 20,20′ | 138.2 | |
| 8,8′ | 134.7 | 21,21′ | 6.29 | 125.3 | |
| 9,9′ | 133.3 | 22,22′ | 6.29 | 131.6 | |
| 10,10′ | 29.4 | 23,23′ | 6.29 | 129.6 | |
| 11,11′ | 1.21 | 24.5 | 24,24′ | 1.98 | 13.1 |
| 12,12′ | 1.21 | 24.5 | 25,25′ | 1.98 | 13.0 |
| 13,13′ | 1.93 | 13.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Jiang, K.; Zhang, W.; Dong, S.; Wu, Y.; Zhang, G.; Wu, S.; Chen, T.; Liu, G. Purification, Identification, and Properties of a Novel Carotenoid Produced by Arthrobacter sp. QL17 Isolated from Mount Qomolangma. Antioxidants 2022, 11, 1493. https://doi.org/10.3390/antiox11081493
Yu X, Jiang K, Zhang W, Dong S, Wu Y, Zhang G, Wu S, Chen T, Liu G. Purification, Identification, and Properties of a Novel Carotenoid Produced by Arthrobacter sp. QL17 Isolated from Mount Qomolangma. Antioxidants. 2022; 11(8):1493. https://doi.org/10.3390/antiox11081493
Chicago/Turabian StyleYu, Xue, Kan Jiang, Wei Zhang, Shuqing Dong, Yujie Wu, Gaosen Zhang, Shiyu Wu, Tuo Chen, and Guangxiu Liu. 2022. "Purification, Identification, and Properties of a Novel Carotenoid Produced by Arthrobacter sp. QL17 Isolated from Mount Qomolangma" Antioxidants 11, no. 8: 1493. https://doi.org/10.3390/antiox11081493