
Influence of Neonatal Sex on Breast Milk Protein and Antioxidant Content in Spanish Women in the First Month of Lactation

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population of Study
2.2. Collection and Processing of Breast Milk Samples
2.3. Fat and Protein Content in Breast Milk
2.4. Breast Milk Antioxidants
2.5. Oxidative Damage of Breast Milk Lipids and Proteins
2.6. Statistical Analysis
3. Results
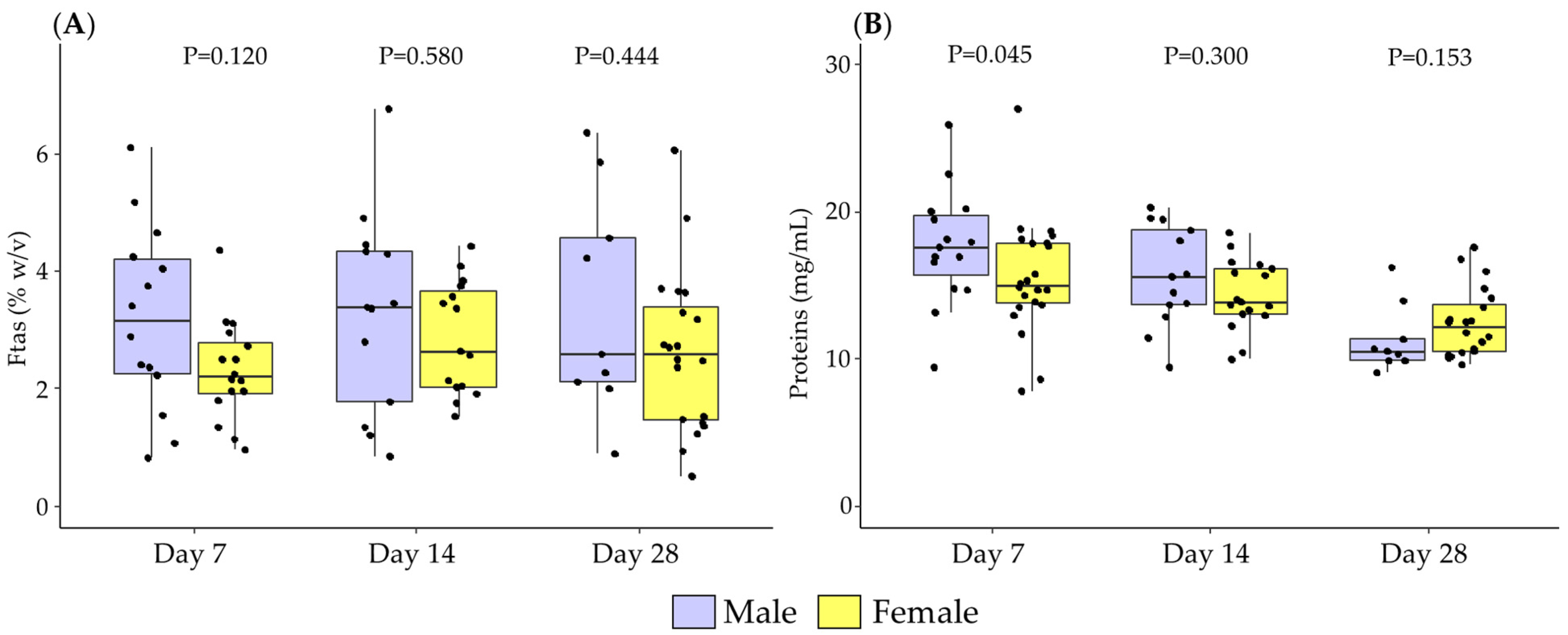
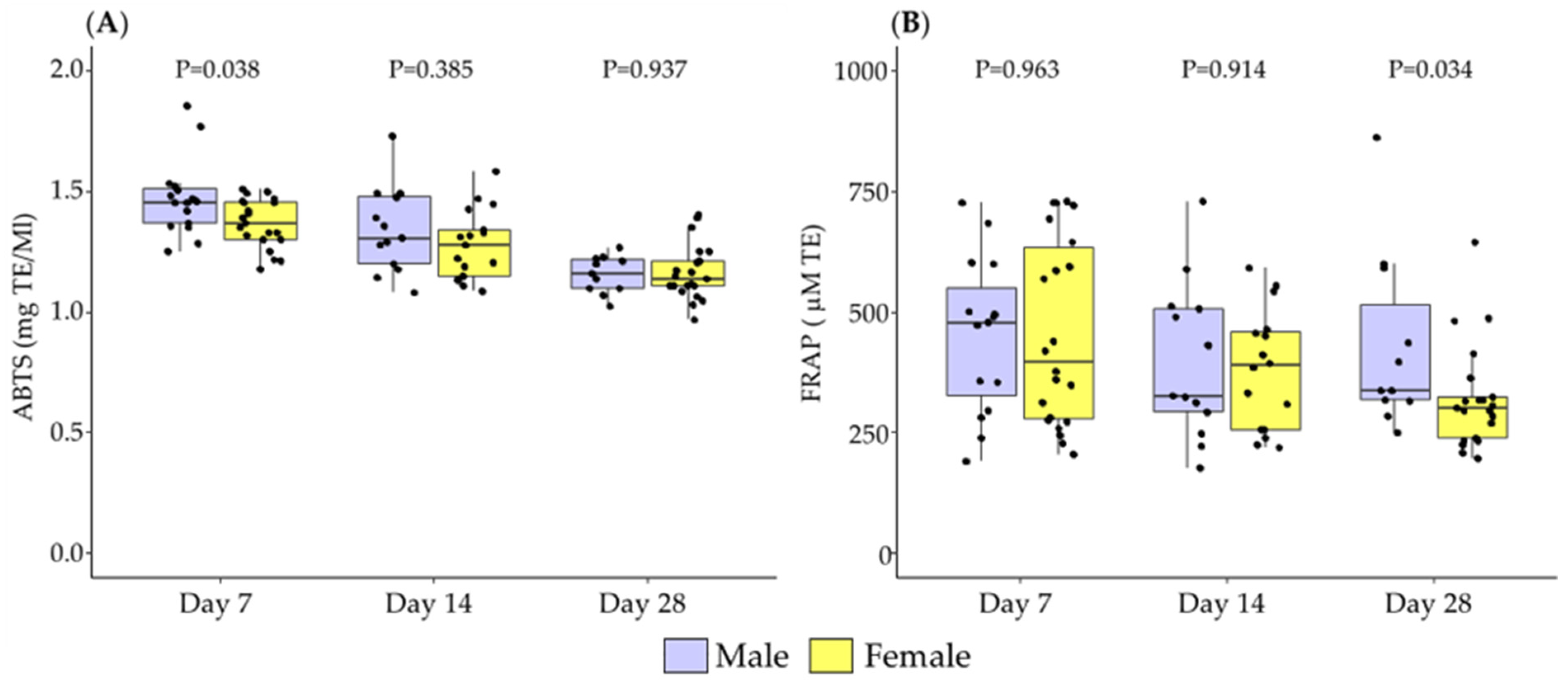
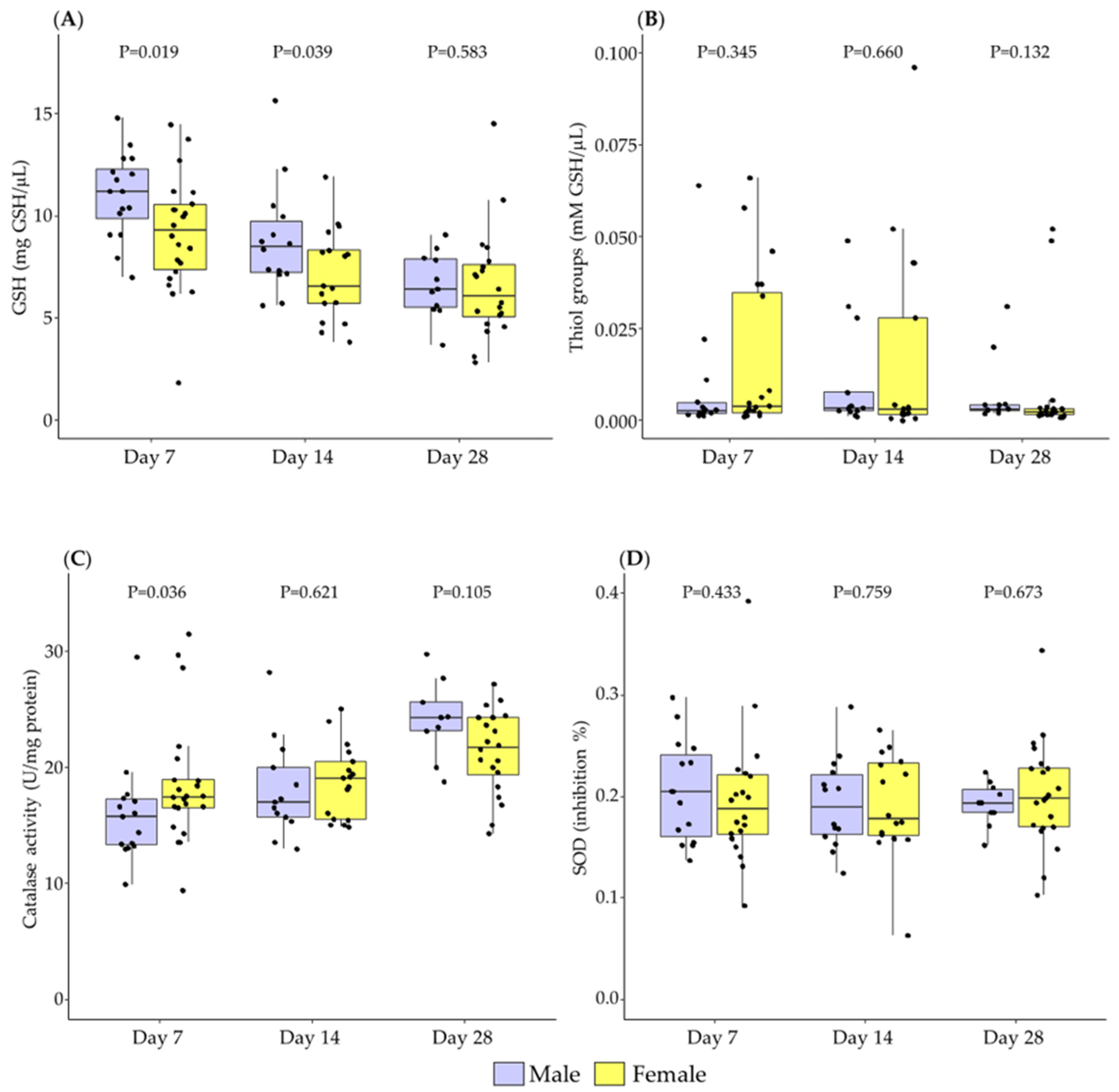
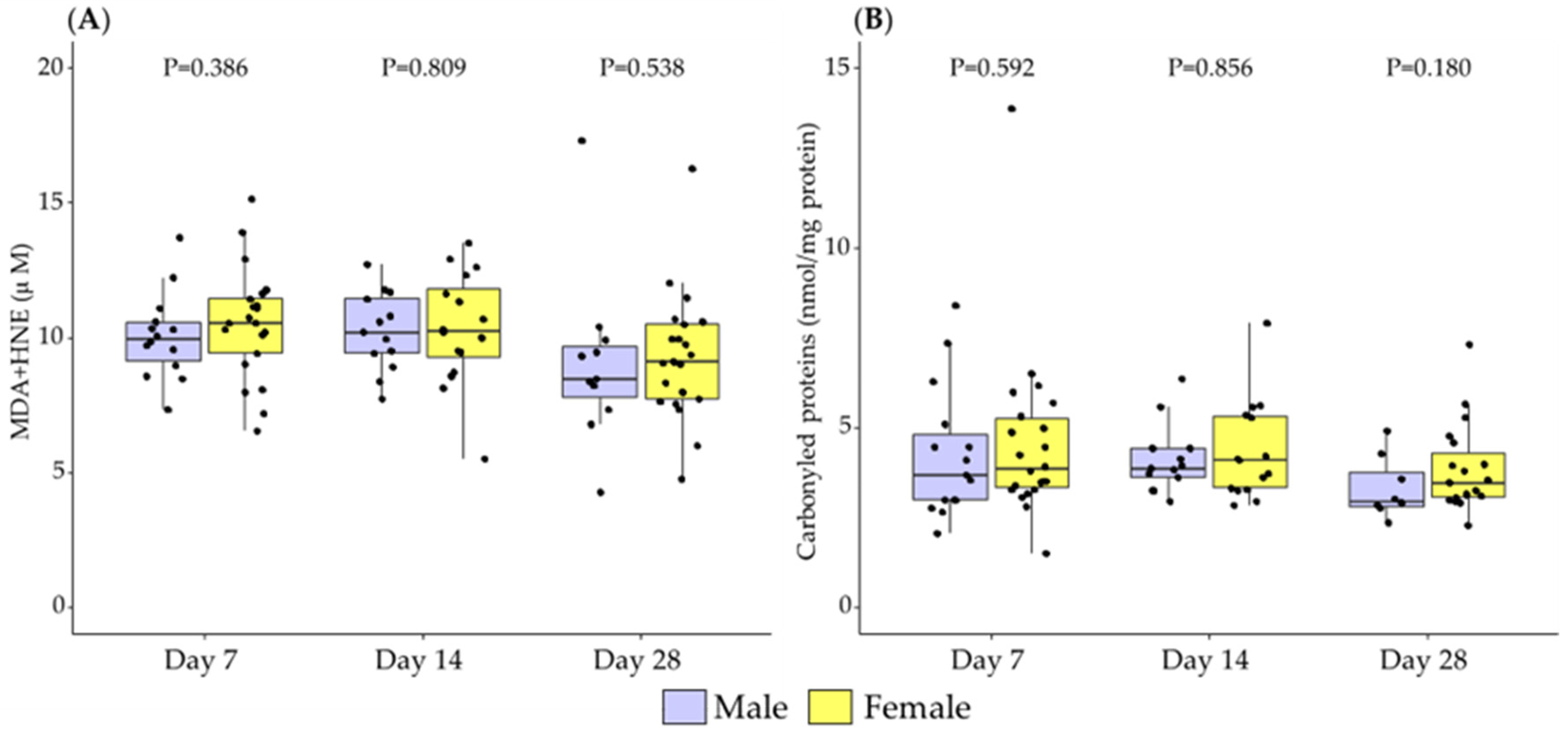
3.1. Differences in Breast Milk Levels According to Newborn Sex
3.2. Association between Newborn Sex and Lactation Period in Breast Milk Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duijts, L.; Jaddoe, V.W.V.; Hofman, A.; Moll, H.A. Prolonged and Exclusive Breastfeeding Reduces the Risk of Infectious Diseases in Infancy. Pediatrics 2010, 126, e18–e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deoni, S.C.L.; Dean, D.C.; Piryatinsky, I.; O’Muircheartaigh, J.; Waskiewicz, N.; Lehman, K.; Han, M.; Dirks, H. Breastfeeding and Early White Matter Development: A Cross-Sectional Study. Neuroimage 2013, 82, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paine, B.J.; Makrides, M.; Gibson, R.A. Duration of Breast-Feeding and Bayley’s Mental Developmental Index at 1 Year of Age. J. Paediatr. Child Health 1999, 35, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Quigley, M.A.; Hockley, C.; Carson, C.; Kelly, Y.; Renfrew, M.J.; Sacker, A. Breastfeeding Is Associated with Improved Child Cognitive Development: A Population-Based Cohort Study. J. Pediatr. 2012, 160, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; de Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J. Maternal and Child Undernutrition Study Group Maternal and Child Undernutrition: Global and Regional Exposures and Health Consequences. Lancet 2008, 371, 243–260. [Google Scholar] [CrossRef]
- Hauck, F.R.; Thompson, J.M.D.; Tanabe, K.O.; Moon, R.Y.; Vennemann, M.M. Breastfeeding and Reduced Risk of Sudden Infant Death Syndrome: A Meta-Analysis. Pediatrics 2011, 128, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ip, S.; Chung, M.; Raman, G.; Chew, P.; Magula, N.; DeVine, D.; Trikalinos, T.; Lau, J. Breastfeeding and Maternal and Infant Health Outcomes in Developed Countries. Evid. Rep. Technol. Assess (Full Rep.) 2007, 153, 1–186. [Google Scholar]
- Amitay, E.L.; Keinan-Boker, L. Breastfeeding and Childhood Leukemia Incidence: A Meta-Analysis and Systematic Review. JAMA Pediatr. 2015, 169, e151025. [Google Scholar] [CrossRef] [Green Version]
- Singhal, A.; Farooqi, I.S.; O’Rahilly, S.; Cole, T.J.; Fewtrell, M.; Lucas, A. Early Nutrition and Leptin Concentrations in Later Life. Am. J. Clin. Nutr. 2002, 75, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Singhal, A.; Cole, T.J.; Lucas, A. Early Nutrition in Preterm Infants and Later Blood Pressure: Two Cohorts after Randomised Trials. Lancet 2001, 357, 413–419. [Google Scholar] [CrossRef]
- Wagner, C.L.; Taylor, S.N.; Johnson, D. Host Factors in Amniotic Fluid and Breast Milk That Contribute to Gut Maturation. Clin. Rev. Allergy Immunol. 2008, 34, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Castellote, C.; Casillas, R.; Ramírez-Santana, C.; Pérez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, A. Premature Delivery Influences the Immunological Composition of Colostrum and Transitional and Mature Human Milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulski, J.K.; Hartmann, P.E. Changes in Human Milk Composition during the Initiation of Lactation. Aust. J. Exp. Biol. Med. Sci. 1981, 59, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Gila-Díaz, A.; Herranz Carrillo, G.; Cañas, S.; Saenz de Pipaón, M.; Martínez-Orgado, J.A.; Rodríguez-Rodríguez, P.; López de Pablo, Á.L.; Martin-Cabrejas, M.A.; Ramiro-Cortijo, D.; Arribas, S.M. Influence of Maternal Age and Gestational Age on Breast Milk Antioxidants During the First Month of Lactation. Nutrients 2020, 12, 2569. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.J.; Ejima, K.; Li, P.; Demerath, E.W.; Allison, D.B.; Fields, D.A. Associations of Breastfeeding or Formula Feeding with Infant Anthropometry and Body Composition at 6 Months. Matern. Child Nutr. 2021, 17, e13105. [Google Scholar] [CrossRef] [PubMed]
- Clifton, V.L. Review: Sex and the Human Placenta: Mediating Differential Strategies of Fetal Growth and Survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef] [PubMed]
- Meakin, A.S.; Cuffe, J.S.M.; Darby, J.R.T.; Morrison, J.L.; Clifton, V.L. Let’s Talk about Placental Sex, Baby: Understanding Mechanisms That Drive Female- and Male-Specific Fetal Growth and Developmental Outcomes. Int. J. Mol. Sci. 2021, 22, 6386. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.L.; Wright, I.M.R.; Abdel-Latif, M.E. New South Wales and Australian Capital Territory Neonatal Intensive Care Units Audit Group Mortality and Adverse Neurologic Outcomes Are Greater in Preterm Male Infants. Pediatrics 2012, 129, 124–131. [Google Scholar] [CrossRef]
- Bucher, M.; Kadam, L.; Ahuna, K.; Myatt, L. Differences in Glycolysis and Mitochondrial Respiration between Cytotrophoblast and Syncytiotrophoblast In-Vitro: Evidence for Sexual Dimorphism. Int. J. Mol. Sci. 2021, 22, 10875. [Google Scholar] [CrossRef]
- Phuthong, S.; Reyes-Hernández, C.G.; Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Gil-Ortega, M.; González-Blázquez, R.; González, M.C.; López de Pablo, A.L.; Arribas, S.M. Sex Differences in Placental Protein Expression and Efficiency in a Rat Model of Fetal Programming Induced by Maternal Undernutrition. Int. J. Mol. Sci. 2020, 22, 237. [Google Scholar] [CrossRef]
- Broere-Brown, Z.A.; Adank, M.C.; Benschop, L.; Tielemans, M.; Muka, T.; Gonçalves, R.; Bramer, W.M.; Schoufour, J.D.; Voortman, T.; Steegers, E.A.P.; et al. Fetal Sex and Maternal Pregnancy Outcomes: A Systematic Review and Meta-Analysis. Biol. Sex Differ. 2020, 11, 26. [Google Scholar] [CrossRef]
- Lorente-Pozo, S.; Parra-Llorca, A.; Torres, B.; Torres-Cuevas, I.; Nuñez-Ramiro, A.; Cernada, M.; García-Robles, A.; Vento, M. Influence of Sex on Gestational Complications, Fetal-to-Neonatal Transition, and Postnatal Adaptation. Front. Pediatr. 2018, 6, 63. [Google Scholar] [CrossRef]
- Naeye, R.L.; Burt, L.S.; Wright, D.L.; Blanc, W.A.; Tatter, D. Neonatal Mortality, the Male Disadvantage. Pediatrics 1971, 48, 902–906. [Google Scholar] [CrossRef]
- Tottman, A.C.; Oliver, C.J.; Alsweiler, J.M.; Cormack, B.E. Do Preterm Girls Need Different Nutrition to Preterm Boys? Sex-Specific Nutrition for the Preterm Infant. Pediatr. Res. 2021, 89, 313–317. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; de la Calle, M.; Böger, R.; Hannemann, J.; Lüneburg, N.; López-Giménez, M.R.; Rodríguez-Rodríguez, P.; Martín-Cabrejas, M.Á.; Benítez, V.; de Pablo, Á.L.L.; et al. Male Fetal Sex Is Associated with Low Maternal Plasma Anti-Inflammatory Cytokine Profile in the First Trimester of Healthy Pregnancies. Cytokine 2020, 136, 155290. [Google Scholar] [CrossRef]
- Galante, L.; Milan, A.M.; Reynolds, C.M.; Cameron-Smith, D.; Vickers, M.H.; Pundir, S. Sex-Specific Human Milk Composition: The Role of Infant Sex in Determining Early Life Nutrition. Nutrients 2018, 10, 1194. [Google Scholar] [CrossRef] [Green Version]
- Herreid, E.O.; Harmon, C. A Study of Methods of Obtaining Milk Samples for Estimating Milk Fat by the Mojonnier Method. J. Dairy Sci. 1944, 27, 33–38. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Martinez-Saez, N.; García, A.T.; Pérez, I.D.; Rebollo-Hernanz, M.; Mesías, M.; Morales, F.J.; Martín-Cabrejas, M.A.; del Castillo, M.D. Use of Spent Coffee Grounds as Food Ingredient in Bakery Products. Food Chem. 2017, 216, 114–122. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of Protein Modification by Oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; de la Calle, M.; Rodríguez-Rodríguez, P.; de Pablo, Á.L.L.; López-Giménez, M.R.; Aguilera, Y.; Martín-Cabrejas, M.A.; González, M.D.C.; Arribas, S.M. Maternal Antioxidant Status in Early Pregnancy and Development of Fetal Complications in Twin Pregnancies: A Pilot Study. Antioxidants 2020, 9, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennet, L. Sex, Drugs and Rock and Roll: Tales from Preterm Fetal Life. J. Physiol. 2017, 595, 1865–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powe, C.E.; Knott, C.D.; Conklin-Brittain, N. Infant Sex Predicts Breast Milk Energy Content. Am. J. Hum. Biol. 2010, 22, 50–54. [Google Scholar] [CrossRef]
- Hahn, W.-H.; Song, J.-H.; Song, S.; Kang, N.M. Do Gender and Birth Height of Infant Affect Calorie of Human Milk? An Association Study between Human Milk Macronutrient and Various Birth Factors. J. Matern. Fetal Neonatal Med. 2017, 30, 1608–1612. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.C.; Mitoulas, L.R.; Cregan, M.D.; Ramsay, D.T.; Doherty, D.A.; Hartmann, P.E. Volume and Frequency of Breastfeedings and Fat Content of Breast Milk throughout the Day. Pediatrics 2006, 117, e387–e395. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Roth, E.; Lo, Y.-J.; Hurst, C.; Vollner, J.; Kendell, A. In Poor Families, Mothers’ Milk Is Richer for Daughters than Sons: A Test of Trivers-Willard Hypothesis in Agropastoral Settlements in Northern Kenya. Am. J. Phys. Anthropol. 2012, 149, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L.; Willard, D.E. Natural Selection of Parental Ability to Vary the Sex Ratio of Offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef] [Green Version]
- Tamimi, R.M.; Lagiou, P.; Mucci, L.A.; Hsieh, C.-C.; Adami, H.-O.; Trichopoulos, D. Average Energy Intake among Pregnant Women Carrying a Boy Compared with a Girl. BMJ 2003, 326, 1245–1246. [Google Scholar] [CrossRef] [Green Version]
- Fields, D.A.; George, B.; Williams, M.; Whitaker, K.; Allison, D.B.; Teague, A.; Demerath, E.W. Associations between Human Breast Milk Hormones and Adipocytokines and Infant Growth and Body Composition in the First 6 Months of Life. Pediatr. Obes. 2017, 12 (Suppl. S1), 78–85. [Google Scholar] [CrossRef] [Green Version]
- Chang, N.; Jung, J.A.; Kim, H.; Jo, A.; Kang, S.; Lee, S.-W.; Yi, H.; Kim, J.; Yim, J.-G.; Jung, B.-M. Macronutrient Composition of Human Milk from Korean Mothers of Full Term Infants Born at 37–42 Gestational Weeks. Nutr. Res. Pract. 2015, 9, 433–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, S.; Ozdemir, T.; Katipoglu, N.; Akcan, A.B.; Kaynak Turkmen, M. Comparison of Changes in Breast Milk Macronutrient Content During the First Month in Preterm and Term Infants. Breastfeed Med. 2020, 15, 56–62. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, L.; Bao, W.; Rong, S. Nutritional Composition of Breast Milk in Chinese Women: A Systematic Review. Asia Pac. J. Clin. Nutr. 2018, 27, 491–502. [Google Scholar] [CrossRef]
- Alexander, T.; Conlon, C.A.; Gamble, G.; von Hurst, P.R.; van Dorp, L.; Ichhpuniani, B.; Housh, R.; Ke, P.; Bloomfield, F.H. Body Composition of New Zealand-Born Term Babies Differs by Ethnicity, Gestational Age and Sex. Early Hum. Dev. 2019, 140, 104924. [Google Scholar] [CrossRef] [PubMed]
- Ramel, S.E.; Gray, H.L.; Christiansen, E.; Boys, C.; Georgieff, M.K.; Demerath, E.W. Greater Early Gains in Fat-Free Mass, but Not Fat Mass, Are Associated with Improved Neurodevelopment at 1 Year Corrected Age for Prematurity in Very Low Birth Weight Preterm Infants. J. Pediatr. 2016, 173, 108–115. [Google Scholar] [CrossRef]
- Panfoli, I.; Candiano, G.; Malova, M.; de Angelis, L.; Cardiello, V.; Buonocore, G.; Ramenghi, L.A. Oxidative Stress as a Primary Risk Factor for Brain Damage in Preterm Newborns. Front. Pediatr. 2018, 6, 369. [Google Scholar] [CrossRef] [Green Version]
- Lorenzetti, S.; Plösch, T.; Teller, I.C. Antioxidative Molecules in Human Milk and Environmental Contaminants. Antioxidants 2021, 10, 550. [Google Scholar] [CrossRef] [PubMed]
- Torres-Cuevas, I.; Parra-Llorca, A.; Sánchez-Illana, A.; Nuñez-Ramiro, A.; Kuligowski, J.; Cháfer-Pericás, C.; Cernada, M.; Escobar, J.; Vento, M. Oxygen and Oxidative Stress in the Perinatal Period. Redox Biol. 2017, 12, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martín-Cabrejas, M.A.; López de Pablo, Á.L.; Sáenz de Pipaón, M.; Ramiro-Cortijo, D. A Review of Bioactive Factors in Human Breastmilk: A Focus on Prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiles, J.L.; Ochoa, J.J.; Ramirez-Tortosa, M.C.; Linde, J.; Bompadre, S.; Battino, M.; Narbona, E.; Maldonado, J.; Mataix, J. Coenzyme Q Concentration and Total Antioxidant Capacity of Human Milk at Different Stages of Lactation in Mothers of Preterm and Full-Term Infants. Free Radic. Res. 2006, 40, 199–206. [Google Scholar] [CrossRef]
- Zarban, A.; Taheri, F.; Chahkandi, T.; Sharifzadeh, G.; Khorashadizadeh, M. Antioxidant and Radical Scavenging Activity of Human Colostrum, Transitional and Mature Milk. J. Clin. Biochem. Nutr. 2009, 45, 150–154. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Castañeda, P.C.; García-González, A.; Bencomo-Alvarez, A.E.; Barros-Nuñez, P.; Gaxiola-Robles, R.; Méndez-Rodríguez, L.C.; Zenteno-Savín, T. Micronutrient Content and Antioxidant Enzyme Activities in Human Breast Milk. J. Trace Elem. Med. Biol. 2019, 51, 36–41. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Kang, H.-M.; Saltveit, M.E. Antioxidant Capacity of Lettuce Leaf Tissue Increases after Wounding. J. Agric. Food Chem. 2002, 50, 7536–7541. [Google Scholar] [CrossRef] [PubMed]
- Brink, L.R.; Lönnerdal, B. Milk Fat Globule Membrane: The Role of Its Various Components in Infant Health and Development. J. Nutr. Biochem. 2020, 85, 108465. [Google Scholar] [CrossRef]
- Saraymen, R.; Kilic, E.; Yazar, S.; Cetin, M. Influence of Sex and Age on the Activity of Antioxidant Enzymes of Polymorphonuclear Leukocytes in Healthy Subjects. Yonsei Med. J. 2003, 44, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giergiel, M.; Lopucki, M.; Stachowicz, N.; Kankofer, M. The Influence of Age and Gender on Antioxidant Enzyme Activities in Humans and Laboratory Animals. Aging Clin. Exp. Res. 2012, 24, 561–569. [Google Scholar] [CrossRef]
- Friel, J.K.; Martin, S.M.; Langdon, M.; Herzberg, G.R.; Buettner, G.R. Milk from Mothers of Both Premature and Full-Term Infants Provides Better Antioxidant Protection than Does Infant Formula. Pediatr. Res. 2002, 51, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Cieslak, M.; Ferreira, C.H.F.; Shifrin, Y.; Pan, J.; Belik, J. Human Milk H2O2 Content: Does It Benefit Preterm Infants? Pediatr. Res. 2018, 83, 687–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gough, D.R.; Cotter, T.G. Hydrogen Peroxide: A Jekyll and Hyde Signalling Molecule. Cell Death Dis. 2011, 2, e213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shehri, S.S.; Knox, C.L.; Liley, H.G.; Cowley, D.M.; Wright, J.R.; Henman, M.G.; Hewavitharana, A.K.; Charles, B.G.; Shaw, P.N.; Sweeney, E.L.; et al. Breastmilk-Saliva Interactions Boost Innate Immunity by Regulating the Oral Microbiome in Early Infancy. PLoS ONE 2015, 10, e0135047. [Google Scholar] [CrossRef] [Green Version]
- Al-Kerwi, E.A.A.; Al-Hashimi, A.H.; Salman, A.M. Mother’s Milk and Hydrogen Peroxide. Asia Pac. J. Clin. Nutr. 2005, 14, 428–431. [Google Scholar]
- Walters, K.A.; Simanainen, U.; Gibson, D.A. Androgen Action in Female Reproductive Physiology. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hoover, K.L.; Barbalinardo, L.H.; Platia, M.P. Delayed Lactogenesis II Secondary to Gestational Ovarian Theca Lutein Cysts in Two Normal Singleton Pregnancies. J. Hum. Lact. 2002, 18, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Baratta, M.; Grolli, S.; Poletti, A.; Ramoni, R.; Motta, M.; Tamanini, C. Role of Androgens in Proliferation and Differentiation of Mouse Mammary Epithelial Cell Line HC11. J. Endocrinol. 2000, 167, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male (n = 24) | Female (n = 31) | p-Value | |
|---|---|---|---|
| Maternal age (years) | 34.0 [30.2; 36.2] | 35.0 [33.0; 37.0] | 0.213 |
| Gestational age (weeks) | 34.5 [28.7; 38.3] | 38.0 [36.2; 39.6] | 0.032 |
| Gravida | 2.00 [1.00; 2.00] | 2.00 [2.00; 2.00] | 0.265 |
| Previous abortion | 6 (25.0%) | 11 (35.5%) | 0.589 |
| Maternal weight (kg) | 70.0 [63.1; 81.4] | 68.8 [57.1; 76.2] | 0.346 |
| Maternal height (cm) | 162 [160; 166] | 160 [157; 163] | 0.178 |
| Body fat (%) | 38.2 [36.0; 42.2] | 37.8 [32.2; 42.6] | 0.651 |
| Muscle (%) | 26.1 [24.4; 27.1] | 26.3 [25.0; 28.1] | 0.492 |
| Twin | 4 (16.7%) | 3 (9.7%) | 0.686 |
| TRA | 0 (0.0%) | 3 (10.3%) | 0.254 |
| C-section | 5 (20.8%) | 11 (35.5%) | 0.375 |
| Fetal growth restriction | 2 (9.1%) | 4 (12.9%) | 1.000 |
| Birth weight (g) | 2060 [1280; 3248] | 2850 [1735; 3210] | 0.594 |
| Birth weight Z-score | −0.71 [−1.36; −0.32] | −0.66 [−1.38; −0.22] | 0.999 |
| Birth length (cm) | 41.0 [37.4; 49.6] | 48.0 [42.5; 48.5] | 0.405 |
| Birth length Z-score | −0.99 [−1.28; −0.62] | −0.27 [−1.41; 0.35] | 0.396 |
| Birth head circumference (cm) | 29.5 [27.0; 34.6] | 33.5 [30.1; 34.5] | 0.553 |
| Birth head circumference Z-score | −0.39 [−0.80; 0.19] | −0.47 [−1.57; 0.06] | 0.686 |
| Apgar at 5 min | 10.0 [9.00; 10.0] | 10.0 [9.00; 10.0] | 0.765 |
| Main Effects | Proteins | p | ABTS | p | FRAP | p | GSH | p | Catalase Activity | p |
|---|---|---|---|---|---|---|---|---|---|---|
| Male | 2.70 ± 1.20 | 0.048 | 0.11 ± 0.06 | 0.056 | 24.82 ± 74.90 | 0.743 | 1.82 ± 0.94 | 0.064 | −3.99 ± 2.17 | 0.078 |
| Day 14 | −1.09 ± 1.22 | 0.383 | −0.10 ± 0.05 | 0.045 | −26.87 ± 56.29 | 0.637 | −2.17 ± 0.87 | 0.019 | −0.17 ± 1.50 | 0.913 |
| Day 28 | −3.42 ± 1.40 | 0.022 | −0.20 ± 0.04 | <0.001 | −62.85 ± 58.03 | 0.289 | −2.53 ± 0.89 | 0.008 | 3.19 ± 1.98 | 0.119 |
| Male: Day 14 | −2.39 ± 2.08 | 0.263 | −0.06 ± 0.08 | 0.410 | 4.81 ± 89.65 | 0.958 | −3.37 ± 1.41 | 0.799 | 3.53 ± 2.56 | 0.181 |
| Male: Day 28 | −0.36 ± 2.48 | 0.887 | −0.12 ± 0.07 | 0.103 | 44.91 ± 94.76 | 0.640 | −1.43 ± 1.44 | 0.332 | 0.71 ± 3.49 | 0.841 |
| AIC/BIC | 332.2/368.5 | 6.1/43.5 | 762.0/799.5 | 318.8/356.6 | 372.8/409.2 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramiro-Cortijo, D.; Gila-Diaz, A.; Herranz Carrillo, G.; Cañas, S.; Gil-Ramírez, A.; Ruvira, S.; Martin-Cabrejas, M.A.; Arribas, S.M. Influence of Neonatal Sex on Breast Milk Protein and Antioxidant Content in Spanish Women in the First Month of Lactation. Antioxidants 2022, 11, 1472. https://doi.org/10.3390/antiox11081472
Ramiro-Cortijo D, Gila-Diaz A, Herranz Carrillo G, Cañas S, Gil-Ramírez A, Ruvira S, Martin-Cabrejas MA, Arribas SM. Influence of Neonatal Sex on Breast Milk Protein and Antioxidant Content in Spanish Women in the First Month of Lactation. Antioxidants. 2022; 11(8):1472. https://doi.org/10.3390/antiox11081472
Chicago/Turabian StyleRamiro-Cortijo, David, Andrea Gila-Diaz, Gloria Herranz Carrillo, Silvia Cañas, Alicia Gil-Ramírez, Santiago Ruvira, María A. Martin-Cabrejas, and Silvia M. Arribas. 2022. "Influence of Neonatal Sex on Breast Milk Protein and Antioxidant Content in Spanish Women in the First Month of Lactation" Antioxidants 11, no. 8: 1472. https://doi.org/10.3390/antiox11081472