
Genetic Factors Associated with Response to Vitamin E Treatment in NAFLD

Abstract
:1. Introduction
2. Genetic Factors Associated with Treatment Response
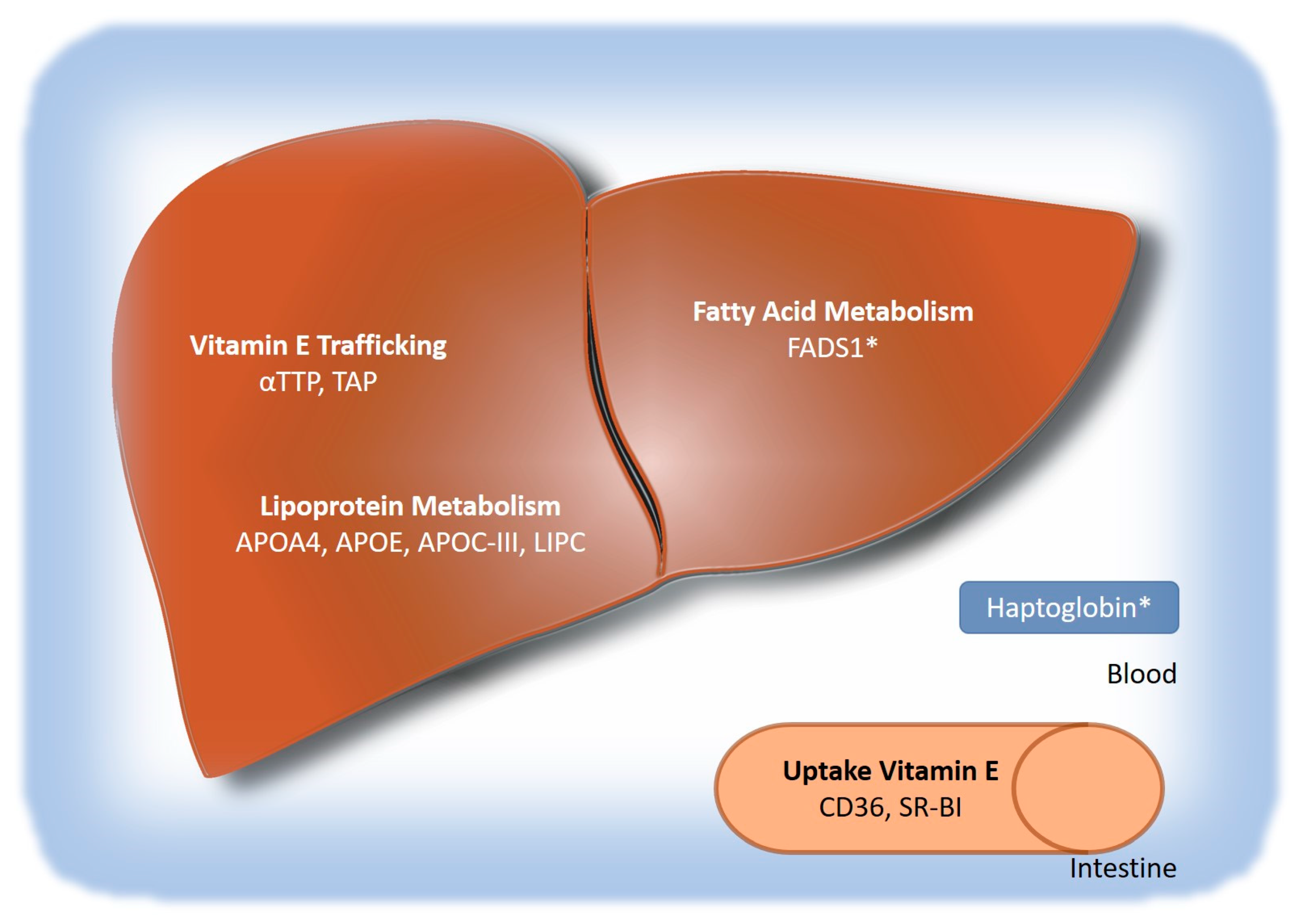
2.1. Genetic Factors Influencing Vitamin E Levels
2.1.1. Genes Involved in Vitamin E Absorption and Hepatic Uptake
2.1.2. Genes Involved in Hepatic Transport
2.1.3. Genes Involved in Extrahepatic Transport
2.1.4. Genes Involved in vitamin E metabolism
2.2. Genetic Factors Associated with NAFLD
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Numbers | Dose/Duration | Patient Characteristics | ALT Change or % Reduction from Baseline | Steatosis | Lobular Inflammation | Ballooning | NAS Score | Fibrosis | Ref |
|---|---|---|---|---|---|---|---|---|---|
| 80 placebo, 80 vitamin E (PIVENS) | 800 IU/d 96 weeks | Adults without diabetes, biopsy confirmed NASH (NAS score of at least 4, presence of ballooning) | −37 ** | −0.7 *** | −0.6 ** | −0.5 * | −1.9 *** | −0.3 | [22] |
| 58 placebo, 58 vitamin E (TONIC) | 800 IU/d 96 weeks | Children (aged 8–17 years), biopsy confirmed NAFLD (>5% of hepatocytes showed macrovesicular fat) | Week 48: −44.5% *; Week 72: −44.2%; Week 96: −48.3% | −0.8 | −0.4 | −0.5 ** | −1.8 * | −0.3 | [23] |
| 32 placebo/36 vitamin E | 800 IU/d 72 weeks | Adults with diabetes, biopsy confirmed NASH (zone 3 macrovesicular steatosis with ballooning and lobular inflammation) | −20 *+ | −1.0 * | −0.4 | −0.5 | not listed | −0.6 | [16] |
| 20 placebo/20 DHA-VE-CHO | 250 mg of DHA, 39 UI of vitamin E, and 201 mg of choline; 48 weeks | Children (aged 4–16 years) with liver biopsy-proven NASH | −18.2 * | −0.8 * | −0.15 | −0.75 ** | −1.7 *** | 0.17 | [87] |
| MAF (Allele) | Effect of MA on αT Plasma/Serum Levels | References | ||
|---|---|---|---|---|
| Polymorphisms associated with αT serum or plasma levels | ||||
| APOA4 | rs675 | 0.17 (A) | A lower | [37] |
| APOE | rs662799 | 0.09 (C) | T/T lower; T/C higher | [50,51] |
| Apo C-III | ApoC-III S1/S2 | 0.2 (C) | in women G/G lower | [53] |
| α-TTP | rs6994076 | 0.45 (A) | A/A higher; A/A lower | [44,45] |
| BUD13 * | rs28927680 | 0.06 (G) | not specified | [40] |
| rs12292921 | 0.07 (A) | A higher | [36] | |
| CD36 | rs1527479 | 0.49 (T) | T/T lower | [38] |
| CYP4F2 | rs2108622 | 0.29 (T) | T/T higher | [30,36] |
| LIPC | rs1800588 | 0.22 (T) | T higher | [40] |
| PNPLA3 | rs738409 | 0.21 (G) | no association | [72] |
| SR-BI | rs11057830 | 0.15 (A) | A/A lower | [36] |
| SR-BI | exon 8 | 0.13 (C) | C/C higher | [37] |
| SLC30A8 | rs13266634 | 0.29 (T) | not specified | [40] |
| SUGP1 * | rs10401969 | 0.08 (C) | not specified | [75] |
| TAP | rs2299826 | 0.11 (T) | T/T higher | [44] |
| TM6SF2 | rs58542926 | 0.07 (T) | T lower | [75] |
| ZPR1 * | rs964184 | 0.15 (G) | G/G higher | [36,75] |
| Polymorphisms associated with αT levels after supplementation | ||||
| α-TTP | rs6994076 | 0.45 (A) | A/A higher on treatment | [44] |
| CYP4F2 | rs2108622 | 0.29 (T) | T/T higher on treatment | [30,36] |
| Polymorphisms and copy number variations associated with treatment response in NAFLD | ||||
| CYP4F2 | rs2108622 | 0.29 (T) | no association with treatment response | [30] |
| FADS1 | rs174576 | 0.34 (A) | A associated with treatment response | [79] |
| Hp | CNV | 0.16 (1/1) | Hp-2 allele associated with treatment response | [84] |
References
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2017, 15, 11–20. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef] [PubMed]
- Rowe, I.A.; Wong, V.W.-S.; Loomba, R. Treatment Candidacy for Pharmacologic Therapies for NASH. Clin. Gastroenterol. Hepatol. 2022, 20, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Arab, J.P.; Arrese, M.; Trauner, M. Recent Insights into the Pathogenesis of Nonalcoholic Fatty Liver Disease. Annu. Rev. Pathol. 2018, 13, 321–350. [Google Scholar] [CrossRef]
- McPherson, S.; Hardy, T.; Henderson, E.; Burt, A.D.; Day, C.P.; Anstee, Q.M. Evidence of NAFLD progression from steatosis to fibrosing-steatohepatitis using paired biopsies: Implications for prognosis and clinical management. J. Hepatol. 2015, 62, 1148–1155. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Wilson, L.A.; Behling, C.; Guy, C.; Contos, M.; Cummings, O.; Yeh, M.; Gill, R.; Chalasani, N.; et al. Association of Histologic Disease Activity with Progression of Nonalcoholic Fatty Liver Disease. JAMA Netw. Open 2019, 2, e1912565. [Google Scholar] [CrossRef] [Green Version]
- Anstee, Q.M.; Targher, G.; Day, C.P. Progression of NAFLD to diabetes mellitus, cardiovascular disease or cirrhosis. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 330–344. [Google Scholar] [CrossRef]
- Houttu, V.; Csader, S.; Nieuwdorp, M.; Holleboom, A.G.; Schwab, U. Dietary Interventions in Patients with Non-alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. Front. Nutr. 2021, 8, 716783. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Lopez, R.; Tamimi, T.A.; Yerian, L.; Chung, Y.M.; Berk, M.; Zhang, R.; McIntyre, T.M.; Hazen, S.L. Mass spectrometric profiling of oxidized lipid products in human nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. J. Lipid Res. 2010, 51, 3046–3054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanuri, G.; Ladurner, R.; Skibovskaya, J.; Spruss, A.; Königsrainer, A.; Bischoff, S.C.; Bergheim, I. Expression of toll-like receptors 1-5 but not TLR 6-10 is elevated in livers of patients with non-alcoholic fatty liver disease. Liver Int. 2013, 35, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Kitada, T.; Yamada, T.; Sakaguchi, H.; Nakatani, K.; Wakasa, K. In situ detection of lipid peroxidation and oxidative DNA damage in non-alcoholic fatty liver diseases. J. Hepatol. 2002, 37, 56–62. [Google Scholar] [CrossRef]
- Podszun, M.C.; Chung, J.-Y.; Ylaya, K.; Kleiner, D.E.; Hewitt, S.M.; Rotman, Y. 4-HNE Immunohistochemistry and Image Analysis for Detection of Lipid Peroxidation in Human Liver Samples Using Vitamin E Treatment in NAFLD as a Proof of Concept. J. Histochem. Cytochem. 2020, 68, 635–643. [Google Scholar] [CrossRef]
- Sato, K.; Gosho, M.; Yamamoto, T.; Kobayashi, Y.; Ishii, N.; Ohashi, T.; Nakade, Y.; Ito, K.; Fukuzawa, Y.; Yoneda, M. Vitamin E has a beneficial effect on nonalcoholic fatty liver disease: A meta-analysis of randomized controlled trials. Nutrition 2015, 31, 923–930. [Google Scholar] [CrossRef]
- Bril, F.; Biernacki, D.M.; Kalavalapalli, S.; Lomonaco, R.; Subbarayan, S.K.; Lai, J.; Tio, F.; Suman, A.; Orsak, B.K.; Hecht, J.; et al. Role of Vitamin E for Nonalcoholic Steatohepatitis in Patients with Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2019, 42, 1481–1488. [Google Scholar] [CrossRef]
- Vilar-Gomez, E.; Vuppalanchi, R.; Gawrieh, S.; Ghabril, M.; Saxena, R.; Cummings, O.W.; Chalasani, N. Vitamin E Improves Transplant-Free Survival and Hepatic Decompensation Among Patients with Nonalcoholic Steatohepatitis and Advanced Fibrosis. Hepatology 2018, 71, 495–509. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.M.; Bishop, K.S. On the existence of a hitherto unrecognized dietary factor essential for reproduction. Science 1922, 56, 650–651. [Google Scholar] [CrossRef] [Green Version]
- Kayden, H.; Traber, M. Absorption, lipoprotein transport, and regulation of plasma concentrations of vitamin E in humans. J. Lipid Res. 1993, 34, 343–358. [Google Scholar] [CrossRef]
- Erhardt, A.; Stahl, W.; Sies, H.; Lirussi, F.; Donner, A.; Häussinger, D. Plasma levels of vitamin e and carotenoids are decreased in patients with nonalcoholic steatohepatitis (nash). Eur. J. Med Res. 2011, 16, 76–78. [Google Scholar] [CrossRef]
- Violet, P.; Traber, M.G.; Levine, M. Vitamin E Sequestration by Liver Fat in Humans. JCI Insight 2020, 5, e133309. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Chalasani, N.; Kowdley, K.V.; McCullough, A.; Diehl, A.M.; Bass, N.M.; Neuschwander-Tetri, B.A.; Lavine, J.E.; Tonascia, J.; Unalp, A.; et al. Pioglitazone, Vitamin E, or Placebo for Nonalcoholic Steatohepatitis. N. Engl. J. Med. 2010, 362, 1675–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavine, J.E.; Schwimmer, J.B.; Van Natta, M.L.; Molleston, J.P.; Murray, K.F.; Rosenthal, P.; Abrams, S.H.; Scheimann, A.O.; Sanyal, A.J.; Chalasani, N.; et al. Effect of Vitamin E or Metformin for Treatment of Nonalcoholic Fatty Liver Disease in Children and Adolescents: The TONIC randomized controlled trial. JAMA: J. Am. Med. Assoc. 2011, 305, 1659–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noureddin, N.; Han, M.A.T.; Alkhouri, N.; Noureddin, M. Accounting for the Placebo Effect and Optimizing Outcomes in Clinical Trials of Nonalcoholic Steatohepatitis (NASH). Curr. Hepatol. Rep. 2020, 19, 63–69. [Google Scholar] [CrossRef]
- Miller, E.R., III.; Pastor-Barriuso, R.; Dalal, D.; Riemersma, R.A.; Appel, L.J.; Guallar, E. Meta-Analysis: High-Dosage Vitamin E Supplementation May Increase All-Cause Mortality. Ann. Intern. Med. 2005, 142, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R.; Cohen, D. Proximity to urban parks and mental health. J. Ment. Health Policy Econ. 2014, 17, 280. [Google Scholar] [CrossRef]
- Köpcke, W. Vitamin E and Mortality: A Critical Perspective of the Conflicting Meta-analysis Outcomes. In Vitamin E in Human Health; Humana Press: Cham, Switzerland, 2019; pp. 235–245. [Google Scholar] [CrossRef]
- Liede, K.E.; Haukka, J.K.; Saxén, L.M.; Heinonen, O.P. Increased tendency towards gingival bleeding caused by joint effect of α-tocopherol supplementation and acetylsalicylic acid. Ann. Med. 1998, 30, 542–546. [Google Scholar] [CrossRef]
- Sesso, H.D.; Buring, J.E.; Christen, W.G.; Kurth, T.; Belanger, C.; MacFadyen, J.; Bubes, V.; Manson, J.E.; Glynn, R.J.; Gaziano, J.M. Vitamins E and C in the Prevention of Cardiovascular Disease in Men. JAMA 2008, 300, 2123–2133. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.A.; Thompson, I.; Tangen, C.M.; John, J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.; Ford, G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the Risk of Prostate Cancer: Updated Results of The Selenium and Vitamin E Cancer Prevention Trial (SE-LECT). J. Am. Med. Assoc. 2011, 306, 1549–1556. [Google Scholar] [CrossRef]
- Athinarayanan, S.; Wei, R.; Zhang, M.; Bai, S.; Traber, M.G.; Yates, K.; Cummings, O.W.; Molleston, J.; Liu, W.; Chalasani, N. Genetic Polymorphism of Cytochrome P450 4F2, Vitamin E Level and Histological Response in Adults and Children with Nonalcoholic Fatty Liver Disease Who Participated in PIVENS and TONIC Clinical Trials. PLoS ONE 2014, 9, e95366. [Google Scholar] [CrossRef] [Green Version]
- Nagita, A.; Ando, M. Assessment of hepatic vitamin E status in adult patients with liver disease. Hepatology 1997, 26, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E.; Klein, A.; Bietrix, F.; Gleize, B.; Malezet-Desmoulins, C.; Schneider, M.; Margotat, A.; Lagrost, L.; Collet, X.; Borel, P. Scavenger Receptor Class B Type I (SR-BI) Is Involved in Vitamin E Transport across the Enterocyte. J. Biol. Chem. 2006, 281, 4739–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, A.; Roi, S.; Nowicki, M.; Niot, I.; Reboul, E. Cluster-determinant 36 (CD36) impacts on vitamin E postprandial response. Mol. Nutr. Food Res. 2014, 58, 2297–2306. [Google Scholar] [CrossRef]
- Narushima, K.; Takada, T.; Yamanashi, Y.; Suzuki, H. Niemann-Pick C1-Like 1 Mediates α-Tocopherol Transport. Mol. Pharmacol. 2008, 74, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Major, J.M.; Yu, K.; Wheeler, W.; Zhang, H.; Cornelis, M.C.; Wright, M.E.; Yeager, M.; Snyder, K.; Weinstein, S.J.; Mondul, A.; et al. Genome-wide association study identifies common variants associated with circulating vitamin E levels. Hum. Mol. Genet. 2011, 20, 3876–3883. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Moussa, M.; Reboul, E.; Lyan, B.; Defoort, C.; Vincent-Baudry, S.; Maillot, M.; Gastaldi, M.; Darmon, M.; Portugal, H.; et al. Human Plasma Levels of Vitamin E and Carotenoids Are Associated with Genetic Polymorphisms in Genes Involved in Lipid Metabolism. J. Nutr. 2007, 137, 2653–2659. [Google Scholar] [CrossRef] [Green Version]
- Lecompte, S.; de Edelenyi, F.S.; Goumidi, L.; Maiani, G.; Moschonis, G.; Widhalm, K.; Molnár, D.; Kafatos, A.; Spinneker, A.; Breidenassel, C.; et al. Polymorphisms in the CD36/FAT gene are associated with plasma vitamin E concentrations in humans. Am. J. Clin. Nutr. 2011, 93, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Krapp, A.; Ahle, S.; Kersting, S.; Hua, Y.; Kneser, K.; Nielsen, M.S.; Gliemann, J.; Beisiegel, U. Hepatic lipase mediates the uptake of chylomicrons and beta-VLDL into cells via the LDL receptor-related protein (LRP). J. Lipid Res. 1996, 37, 926–936. [Google Scholar] [CrossRef]
- Hall, M.A.; Verma, A.; Brown-Gentry, K.D.; Goodloe, R.; Boston, J.; Wilson, S.; McClellan, B.; Sutcliffe, C.; Dilks, H.H.; Gillani, N.B.; et al. Detection of Pleiotropy through a Phenome-Wide Association Study (PheWAS) of Epidemiologic Data as Part of the Environmental Architecture for Genes Linked to Environment (EAGLE) Study. PLoS Genet. 2014, 10, e1004678. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Ouahchi, K.; Arita, M.; Kayden, H.; Hentati, F.; Hamida, M.B.; Sokol, R.; Arai, H.; Inoue, K.; Mandel, J.L.; Koenig, M. Ataxia with Isolated Vitamin E Protein. Nat. Genet. 1995, 9, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Ulatowski, L.; Dreussi, C.; Noy, N.; Barnholtz-Sloan, J.; Klein, E.; Manor, D. Expression of the α-tocopherol transfer protein gene is regulated by oxidative stress and common single-nucleotide polymorphisms. Free Radic. Biol. Med. 2012, 53, 2318–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, M.E.; Peters, U.; Gunter, M.J.; Moore, S.C.; Lawson, K.A.; Yeager, M.; Weinstein, S.J.; Snyder, K.; Virtamo, J.; Albanes, D. Association of Variants in Two Vitamin E Transport Genes with Circulating Vitamin E Concentrations and Prostate Cancer Risk. Cancer Res. 2009, 69, 1429–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanon-Moreno, V.; Asensio-Marquez, E.M.; Ciancotti-Oliver, L.; Garcia-Medina, J.J.; Sanz, P.; Ortega-Azorin, C.; Pinazo-Duran, M.D.; Ordovás, J.M.; Corella, D. Effects of Polymorphisms in Vitamin E-, Vitamin C-, and Glutathione Perox-idase-Related Genes on Serum Biomarkers and Associations with Glaucoma. Mol. Vis. 2013, 19, 231–242. [Google Scholar] [PubMed]
- Yamauchi, J.; Iwamoto, T.; Kida, S.; Masushige, S.; Yamada, K.; Esashi, T. Tocopherol-Associated Protein Is a Ligand-Dependent Transcriptional Activator. Biochem. Biophys. Res. Commun. 2001, 285, 295–299. [Google Scholar] [CrossRef]
- Kempná, P.; Zingg, J.-M.; Ricciarelli, R.; Hierl, M.; Saxena, S.; Azzi, A. Cloning of novel human SEC14p-like proteins: Ligand binding and functional properties. Free Radic. Biol. Med. 2003, 34, 1458–1472. [Google Scholar] [CrossRef]
- Podszun, M.C.; Alawad, A.S.; Lingala, S.; Morris, N.; Huang, W.-C.A.; Yang, S.; Schoenfeld, M.; Rolt, A.; Ouwerkerk, R.; Valdez, K.; et al. Vitamin E treatment in NAFLD patients demonstrates that oxidative stress drives steatosis through upregulation of de-novo lipogenesis. Redox Biol. 2020, 37, 101710. [Google Scholar] [CrossRef]
- Weinberg, R.B.; Cook, V.R.; Beckstead, J.A.; Martin, D.D.O.; Gallagher, J.W.; Shelness, G.S.; Ryan, R.O. Structure and Interfacial Properties of Human Apolipoprotein A-V. J. Biol. Chem. 2003, 278, 34438–34444. [Google Scholar] [CrossRef] [Green Version]
- Girona, J.; Guardiola, M.; Cabré, A.; Manzanares, J.M.; Heras, M.; Ribalta, J.; Masana, L.; Masana, L. The apolipoprotein A5 gene –1131T→C polymorphism affects vitamin E plasma concentrations in type 2 diabetic patients. Clin. Chem. Lab. Med. 2008, 46, 453–457. [Google Scholar] [CrossRef]
- Sundl, I.; Guardiola, M.; Khoschsorur, G.; Solà, R.; Vallve, J.-C.; Godàs, G.; Masana, L.; Maritschnegg, M.; Meinitzer, A.; Cardinault, N.; et al. Increased concentrations of circulating vitamin E in carriers of the apolipoprotein A5 gene −1131T>C variant and associations with plasma lipids and lipid peroxidation. J. Lipid Res. 2007, 48, 2506–2513. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, C.; Harry, P.; Lagrost, L.; White, S.; Shah, N.; North, J.; Gilligan, M.; Gambert, P.; Ball, M. Variation at the apo AI/CIII/A1V gene complex is associated with elevated plasma levels of apo CIII. Atherosclerosis 1991, 87, 239–247. [Google Scholar] [CrossRef]
- Borel, P.; Moussa, M.; Reboul, E.; Lyan, B.; Defoort, C.; Vincent-Baudry, S.; Maillot, M.; Gastaldi, M.; Darmon, M.; Portugal, H.; et al. Human fasting plasma concentrations of vitamin E and carotenoids, and their association with genetic variants in apo C-III, cholesteryl ester transfer protein, hepatic lipase, intestinal fatty acid binding protein and microsomal triacylglycerol transfer protein. Br. J. Nutr. 2008, 101, 680–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardowell, S.A.; Ding, X.; Parker, R.S. Disruption of P450-mediated vitamin E hydroxylase activities alters vitamin E status in tocopherol supplemented mice and reveals extra-hepatic vitamin E metabolism. J. Lipid Res. 2012, 53, 2667–2676. [Google Scholar] [CrossRef] [Green Version]
- Schmolz, L.; Birringer, M.; Lorkowski, S.; Wallert, M. Complexity of vitamin e metabolism. World J. Biol. Chem. 2016, 7, 14–43. [Google Scholar] [CrossRef]
- Sontag, T.J.; Parker, R.S. Influence of major structural features of tocopherols and tocotrienols on their ω-oxidation by tocopherol-ω-hydroxylase. J. Lipid Res. 2007, 48, 1090–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Joyce, A.; Yates, K.; Aouizerat, B.; Sanyal, A.J. Metabolomic Profiling to Identify Predictors of Response to Vitamin E for Non-Alcoholic Steatohepatitis (NASH). PLoS ONE 2012, 7, e44106. [Google Scholar] [CrossRef]
- Bardowell, S.A.; Stec, D.E.; Parker, R.S. Common Variants of Cytochrome P450 4F2 Exhibit Altered Vitamin E-ω-Hydroxylase Specific Activity. J. Nutr. 2010, 140, 1901–1906. [Google Scholar] [CrossRef] [Green Version]
- Anstee, Q.M.; Day, C.P. The genetics of NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 645–655. [Google Scholar] [CrossRef]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2008, 40, 1461–1465. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, C.; Yerges-Armstrong, L.M.; Wu, J.; Hernaez, R.; Kim, L.J.; Palmer, L.; Gudnason, V.; Eiriksdottir, G.; Garcia, M.E.; Launer, L.J.; et al. Genome-Wide Association Analysis Identifies Variants Associated with Nonalcoholic Fatty Liver Disease That Have Distinct Effects on Metabolic Traits. PLoS Genet. 2011, 7, e1001324. [Google Scholar] [CrossRef]
- Chambers, J.C.; Alcohol Genome-wide Association (AlcGen) Consortium; Zhang, W.; Sehmi, J.; Li, X.; Wass, M.; Van Der Harst, P.; Holm, H.; Sanna, S.; Kavousi, M.; et al. Genome-wide association study identifies loci influencing concentrations of liver enzymes in plasma. Nat. Genet. 2011, 43, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Anstee, Q.M.; Darlay, R.; Cockell, S.; Meroni, M.; Govaere, O.; Tiniakos, D.; Burt, A.D.; Bedossa, P.; Palmer, J.; Liu, Y.-L.; et al. Genome-wide association study of non-alcoholic fatty liver and steatohepatitis in a histologically characterised cohort☆. J. Hepatol. 2020, 73, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Conti, D.V.; Bogumil, D.; Sheng, X.; Noureddin, M.; Wilkens, L.R.; Le Marchand, L.; Rosen, H.R.; Haiman, C.A.; Setiawan, V.W. Association of Genetic Risk Score with NAFLD in An Ethnically Diverse Cohort. Hepatol. Commun. 2021, 5, 1689–1703. [Google Scholar] [CrossRef] [PubMed]
- Parisinos, C.A.; Wilman, H.R.; Thomas, E.L.; Kelly, M.; Nicholls, R.C.; McGonigle, J.; Neubauer, S.; Hingorani, A.D.; Patel, R.S.; Hemingway, H.; et al. Genome-wide and Mendelian randomisation studies of liver MRI yield insights into the pathogenesis of steatohepatitis. J. Hepatol. 2020, 73, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Targher, G.; Mantovani, A.; Alisi, A.; Mosca, A.; Panera, N.; Byrne, C.D.; Nobili, V. Relationship Between PNPLA3 rs738409 Polymorphism and Decreased Kidney Function in Children With NAFLD. Hepatology 2019, 70, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Salari, N.; Darvishi, N.; Mansouri, K.; Ghasemi, H.; Hosseinian-Far, M.; Darvishi, F.; Mohammadi, M. Association between PNPLA3 rs738409 polymorphism and nonalcoholic fatty liver disease: A systematic review and meta-analysis. BMC Endocr. Disord. 2021, 21, 125. [Google Scholar] [CrossRef]
- Trépo, E.; Romeo, S.; Zucman-Rossi, J.; Nahon, P. PNPLA3 gene in liver diseases. J. Hepatol. 2016, 65, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Rotman, Y.; Koh, C.; Zmuda, J.M.; Kleiner, D.E.; Liang, T.J.; NASH CRN. The association of genetic variability in patatin-like phospholipase domain-containing protein 3 (PNPLA3) with histological severity of nonalcoholic fatty liver disease. Hepatology 2010, 52, 894–903. [Google Scholar] [CrossRef] [Green Version]
- Fukui, A.; Kawabe, N.; Hashimoto, S.; Murao, M.; Nakano, T.; Shimazaki, H.; Kan, T.; Nakaoka, K.; Ohki, M.; Takagawa, Y.; et al. Vitamin E reduces liver stiffness in nonalcoholic fatty liver disease. World J. Hepatol. 2015, 7, 2749–2756. [Google Scholar] [CrossRef] [Green Version]
- Mondul, A.; Mancina, R.M.; Merlo, A.; Dongiovanni, P.; Rametta, R.; Montalcini, T.; Valenti, L.; Albanes, D.; Romeo, S. PNPLA3 I148M Variant Influences Circulating Retinol in Adults with Nonalcoholic Fatty Liver Disease or Obesity. J. Nutr. 2015, 145, 1687–1691. [Google Scholar] [CrossRef] [Green Version]
- Kozlitina, J.; Smagris, E.; Stender, S.; Nordestgaard, B.G.; Zhou, H.H.; Tybjærg-Hansen, A.; Vogt, T.F.; Hobbs, H.H.; Cohen, J.C. Exome-wide association study identifies a TM6SF2 variant that confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2014, 46, 352–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdessian, H.; Taxiarchis, A.; Popov, S.; Silveira, A.; Franco-Cereceda, A.; Hamsten, A.; Eriksson, P.; Hooft, F.V. TM6SF2 is a regulator of liver fat metabolism influencing triglyceride secretion and hepatic lipid droplet content. Proc. Natl. Acad. Sci. USA 2014, 111, 8913–8918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, A.R.; Perry, J.R.B.; Tanaka, T.; Hernandez, D.G.; Zheng, H.-F.; Melzer, D.; Gibbs, J.R.; Nalls, M.A.; Weedon, M.N.; Spector, T.D.; et al. Imputation of Variants from the 1000 Genomes Project Modestly Improves Known Associations and Can Identify Low-frequency Variant-Phenotype Associations Undetected by HapMap Based Imputation. PLoS ONE 2013, 8, e64343. [Google Scholar] [CrossRef] [PubMed]
- Dallio, M.; Masarone, M.; Romeo, M.; Tuccillo, C.; Morisco, F.; Persico, M.; Loguercio, C.; Federico, A. PNPLA3, TM6SF2, and MBOAT7 Influence on Nutraceutical Therapy Response for Non-alcoholic Fatty Liver Disease: A Randomized Controlled Trial. Front. Med. 2021, 8, 734847. [Google Scholar] [CrossRef]
- Athinarayanan, S.; Fan, Y.; Wang, X.; Callaway, E.; Cai, D.; Chalasani, N.; Chapkin, R.S.; Liu, W. Fatty Acid Desaturase 1 Influences Hepatic Lipid Homeostasis by Modulating the PPARα-FGF21 Axis. Hepatol. Commun. 2020, 5, 461–477. [Google Scholar] [CrossRef]
- Mirkov, S.; Myers, J.L.; Ramírez, J.; Liu, W. SNPs affecting serum metabolomic traits may regulate gene transcription and lipid accumulation in the liver. Metabolism 2012, 61, 1523–1527. [Google Scholar] [CrossRef] [Green Version]
- Nobili, V.; Alisi, A.; Liu, Z.; Liang, T.; Crudele, A.; Raponi, M.; Lin, J.; Chalasani, N.P.; Liu, W. In a pilot study, reduced fatty acid desaturase 1 function was associated with nonalcoholic fatty liver disease and response to treatment in children. Pediatr. Res. 2018, 84, 696–703. [Google Scholar] [CrossRef]
- Yang, F.; Ghio, A.J.; Herbert, D.C.; Weaker, F.J.; Walter, C.A.; Coalson, J.J. Pulmonary Expression of the Human Haptoglobin Gene. Am. J. Respir. Cell Mol. Biol. 2000, 23, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.E.; Schaer, M.J.; Buehler, P.W.; Wilson, M.T.; Reeder, B.J.; Silkstone, G.; Svistunenko, D.A.; Bülow, L.; Alayash, A.I. Haptoglobin Binding Stabilizes Hemoglobin Ferryl Iron and the Globin Radical on Tyrosine β145. Antioxid. Redox Signal. 2013, 18, 2264–2273. [Google Scholar] [CrossRef]
- Galicia, G.; Ceuppens, J.L. Haptoglobin Function and Regulation in Autoimmune Diseases. In Acute Phase Proteins: Regulation and Functions of Acute Phase Proteins; Intechopen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- MacKellar, M.; Vigerust, D.J. Role of Haptoglobin in Health and Disease: A Focus on Diabetes. Clin. Diabetes 2016, 34, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Banini, B.A.; Cazanave, S.C.; Yates, K.P.; Asgharpour, A.; Vincent, R.; Mirshahi, F.; Le, P.; Contos, M.J.; Tonascia, J.; Chalasani, N.P.; et al. Haptoglobin 2 Allele is Associated with Histologic Response to Vitamin E in Subjects with Nonalcoholic Steatohepatitis. J. Clin. Gastroenterol. 2019, 53, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Boettger, L.M.; Salem, R.M.; Handsaker, R.E.; Peloso, G.M.; Kathiresan, S.; Hirschhorn, J.N.; McCarroll, S.A. Recurring exon deletions in the HP (haptoglobin) gene contribute to lower blood cholesterol levels. Nat. Genet. 2016, 48, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; the Chinese NAFLD Clinical Research Network (CNAFLD CRN); Liu, J.; Sheng, H.; You, N.; Chen, J.; Mi, X.; Yang, W.; Zang, S.; Shi, J. Haptoglobin 2-2 Genotype is Associated with More Advanced Disease in Subjects with Non-Alcoholic Steatohepatitis: A Retrospective Study. Adv. Ther. 2019, 36, 880–895. [Google Scholar] [CrossRef] [PubMed]
- Zang, S.; On behalf of the Chinese NAFLD Clinical Research Network (CNAFLD CRN); Chen, J.; Song, Y.; Bai, L.; Chen, J.; Chi, X.; He, F.; Sheng, H.; Wang, J.; et al. Haptoglobin Genotype and Vitamin E Versus Placebo for the Treatment of Nondiabetic Patients with Nonalcoholic Steatohepatitis in China: A Multicenter, Randomized, Placebo-Controlled Trial Design. Adv. Ther. 2018, 35, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Zöhrer, E.; Alisi, A.; Jahnel, J.; Mosca, A.; Della Corte, C.; Crudele, A.; Fauler, G.; Nobili, V. Efficacy of docosahexaenoic acid–choline–vitamin E in paediatric NASH: A randomized controlled clinical trial. Appl. Physiol. Nutr. Metab. 2017, 42, 948–954. [Google Scholar] [CrossRef]

| % Patients: Placebo vs. Vitamin E | ||||||
|---|---|---|---|---|---|---|
| Improvement of Primary Outcome | Steatosis | Inflammation | Ballooning | NASH Resolution | Reference | |
| PIVENS | 19 vs. 43 * | 31 vs. 54 * | 35 vs. 54 * | 29 vs. 50 * | 21 vs. 36 | [22] |
| TONIC | 27 vs. 31 | 20 vs. 22 | 10 vs. 22 * | 28 vs. 58 * | [23] | |
| Bril et al. | 46 vs. 68 | 43 vs. 36 | 35 vs. 50 | 12 vs. 33 * | [16] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Civelek, M.; Podszun, M.C. Genetic Factors Associated with Response to Vitamin E Treatment in NAFLD. Antioxidants 2022, 11, 1284. https://doi.org/10.3390/antiox11071284
Civelek M, Podszun MC. Genetic Factors Associated with Response to Vitamin E Treatment in NAFLD. Antioxidants. 2022; 11(7):1284. https://doi.org/10.3390/antiox11071284
Chicago/Turabian StyleCivelek, Mehtap, and Maren C. Podszun. 2022. "Genetic Factors Associated with Response to Vitamin E Treatment in NAFLD" Antioxidants 11, no. 7: 1284. https://doi.org/10.3390/antiox11071284