
The Role of Antioxidants in the Interplay between Oxidative Stress and Senescence

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Enzymatic Antioxidants
2.2. Mitochondria-Targeted Antioxidants
2.3. Vitamins
2.4. Carotenoids
2.5. Organosulphur Compounds
2.6. Nitrogen Non-Protein Compounds
2.7. Flavonoids
2.8. Non-Flavonoids
2.9. Minerals
2.10. Others—Melatonin
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Calcinotto, A.; Kohli, J.; Zagato, E.; Pellegrini, L.; Demaria, M.; Alimonti, A. Cellular Senescence: Aging, Cancer, and Injury. Physiol. Rev. 2019, 99, 1047–1078. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- di Micco, R.; Krizhanovsky, V.; Baker, D.; d’Adda di Fagagna, F. Cellular Senescence in Ageing: From Mechanisms to Therapeutic Opportunities. Nat. Rev. Mol. Cell Biol. 2021, 22, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Krizhanovsky, V.; Yon, M.; Dickins, R.A.; Hearn, S.; Simon, J.; Miething, C.; Yee, H.; Zender, L.; Lowe, S.W. Senescence of Activated Stellate Cells Limits Liver Fibrosis. Cell 2008, 134, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Sagiv, A.; Krizhanovsky, V. Immunosurveillance of Senescent Cells: The Bright Side of the Senescence Program. Biogerontology 2013, 14, 617–628. [Google Scholar] [CrossRef]
- Lozano-Torres, B.; Estepa-Fernández, A.; Rovira, M.; Orzáez, M.; Serrano, M.; Martínez-Máñez, R.; Sancenón, F. The Chemistry of Senescence. Nat. Rev. Chem. 2019, 3, 426–441. [Google Scholar] [CrossRef]
- Gao, X.; Yu, X.; Zhang, C.; Wang, Y.; Sun, Y.; Sun, H.; Zhang, H.; Shi, Y.; He, X. Telomeres and Mitochondrial Metabolism: Implications for Cellular Senescence and Age-Related Diseases. Stem Cell Rev. Rep. 2022, 1–13. [Google Scholar] [CrossRef]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Roger, L.; Tomas, F.; Gire, V. Mechanisms and Regulation of Cellular Senescence. Int. J. Mol. Sci. 2021, 22, 13173. [Google Scholar] [CrossRef]
- Casella, G.; Tsitsipatis, D.; Abdelmohsen, K.; Gorospe, M. MRNA Methylation in Cell Senescence. WIREs RNA 2019, 10, e1547. [Google Scholar] [CrossRef] [PubMed]
- Crouch, J.; Shvedova, M.; Thanapaul, R.J.R.S.; Botchkarev, V.; Roh, D. Epigenetic Regulation of Cellular Senescence. Cells 2022, 11, 672. [Google Scholar] [CrossRef]
- Lettieri-Barbato, D.; Aquilano, K.; Punziano, C.; Minopoli, G.; Faraonio, R. MicroRNAs, Long Non-Coding RNAs, and Circular RNAs in the Redox Control of Cell Senescence. Antioxidants 2022, 11, 480. [Google Scholar] [CrossRef]
- Sun, J.; Cheng, B.; Su, Y.; Li, M.; Ma, S.; Zhang, Y.; Zhang, A.; Cai, S.; Bao, Q.; Wang, S.; et al. The Potential Role of M6A RNA Methylation in the Aging Process and Aging-Associated Diseases. Front. Genet. 2022, 13, 869950. [Google Scholar] [CrossRef] [PubMed]
- Harries, L.W. Dysregulated RNA Processing and Metabolism: A New Hallmark of Ageing and Provocation for Cellular Senescence. FEBS J. 2022. [Google Scholar] [CrossRef]
- Gheitasi, I.; Azizi, A.; Omidifar, N.; Doustimotlagh, A.H. Renoprotective Effects of Origanum Majorana Methanolic L and Carvacrol on Ischemia/Reperfusion-Induced Kidney Injury in Male Rats. Evid.-Based Complement. Altern. Med. 2020, 2020, 9785932. [Google Scholar] [CrossRef]
- Gholami, A.; Emadi, F.; Amini, A.; Shokripour, M.; Chashmpoosh, M.; Omidifar, N. Functionalization of Graphene Oxide Nanosheets Can Reduce Their Cytotoxicity to Dental Pulp Stem Cells. J. Nanomater. 2020, 2020, 6942707. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative Stress, Aging, and Diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Omidifar, N.; moghadami, M.; Mousavi, S.M.; Hashemi, S.A.; Gholami, A.; Shokripour, M.; Sohrabi, Z. Trends in Natural Nutrients for Oxidative Stress and Cell Senescence. Oxidative Med. Cell. Longev. 2021, 2021, 7501424. [Google Scholar] [CrossRef]
- Rangel-Zúñiga, O.A.; Corina, A.; Lucena-Porras, B.; Cruz-Teno, C.; Gómez-Delgado, F.; Jiménez-Lucena, R.; Alcalá-Díaz, J.F.; Haro-Mariscal, C.; Yubero-Serrano, E.M.; Delgado-Lista, J.; et al. TNFA Gene Variants Related to the Inflammatory Status and Its Association with Cellular Aging: From the CORDIOPREV Study. Exp. Gerontol. 2016, 83, 56–62. [Google Scholar] [CrossRef]
- Sriram, S.; Yuan, C.; Chakraborty, S.; Tay, W.; Park, M.; Shabbir, A.; Toh, S.-A.; Han, W.; Sugii, S. Oxidative Stress Mediates Depot-Specific Functional Differences of Human Adipose-Derived Stem Cells. Stem Cell Res. Ther. 2019, 10, 141. [Google Scholar] [CrossRef]
- Yu, C.; Xiao, J.-H. The Keap1-Nrf2 System: A Mediator between Oxidative Stress and Aging. Oxidative Med. Cell. Longev. 2021, 2021, 6635460. [Google Scholar] [CrossRef]
- Lopes-Paciencia, S.; Saint-Germain, E.; Rowell, M.-C.; Ruiz, A.F.; Kalegari, P.; Ferbeyre, G. The Senescence-Associated Secretory Phenotype and Its Regulation. Cytokine 2019, 117, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.; Serrano, M. Senescence in Tumours: Evidence from Mice and Humans. Nat. Rev. Cancer 2010, 10, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, M.; Nuñez, S.; Heard, E.; Narita, M.; Lin, A.W.; Hearn, S.A.; Spector, D.L.; Hannon, G.J.; Lowe, S.W. Rb-Mediated Heterochromatin Formation and Silencing of E2F Target Genes during Cellular Senescence. Cell 2003, 113, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Serrano, M.; Lin, A.W.; McCurrach, M.E.; Beach, D.; Lowe, S.W. Oncogenic Ras Provokes Premature Cell Senescence Associated with Accumulation of P53 and P16INK4a. Cell 1997, 88, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.; Fielder, E.; Passos, J.F. Mitochondrial Dysfunction and Cell Senescence: Deciphering a Complex Relationship. FEBS Lett. 2019, 593, 1566–1579. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Meng, P.; Ling, X.; Zhou, L. Advancements in Therapeutic Drugs Targeting of Senescence. Ther. Adv. Chronic Dis. 2020, 11, 204062232096412. [Google Scholar] [CrossRef]
- Elsallabi, O.; Patruno, A.; Pesce, M.; Cataldi, A.; Carradori, S.; Gallorini, M. Fisetin as a Senotherapeutic Agent: Biopharmaceutical Properties and Crosstalk between Cell Senescence and Neuroprotection. Molecules 2022, 27, 738. [Google Scholar] [CrossRef]
- Coling, D.; Chen, S.; Chi, L.-H.; Jamesdaniel, S.; Henderson, D. Age-Related Changes in Antioxidant Enzymes Related to Hydrogen Peroxide Metabolism in Rat Inner Ear. Neurosci. Lett. 2009, 464, 22–25. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.-F. Superoxide Dismutases: Ancient Enzymes and New Insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frye, K.A.; Sendra, K.M.; Waldron, K.J.; Kehl-Fie, T.E. Old Dogs, New Tricks: New Insights into the Iron/Manganese Superoxide Dismutase Family. J. Inorg. Biochem. 2022, 230, 111748. [Google Scholar] [CrossRef] [PubMed]
- Treiber, N.; Maity, P.; Singh, K.; Ferchiu, F.; Wlaschek, M.; Scharffetter-Kochanek, K. The Role of Manganese Superoxide Dismutase in Skin Aging. Derm.-Endocrinol. 2012, 4, 232–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, G.J.; Varani, J.; Voorhees, J.J. Looking Older. Arch. Dermatol. 2008, 144, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced Expression of Connective Tissue Growth Factor (CTGF/CCN2) Mediates Collagen Loss in Chronologically Aged Human Skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, R.G.; Tresini, M.; Keogh, B.P.; Doggett, D.L.; Cristofalo, V.J. Differences in Electron Transport Potential, Antioxidant Defenses, and Oxidant Generation in Young and Senescent Fetal Lung Fibroblasts (WI-38). J. Cell Physiol 1999, 180, 114–122. [Google Scholar] [CrossRef]
- Borlon, C.; Debacq-Chainiaux, F.; Hinrichs, C.; Scharffetter-Kochanek, K.; Toussaint, O.; Wlaschek, M. The Gene Expression Profile of Psoralen plus UVA-Induced Premature Senescence in Skin Fibroblasts Resembles a Combined DNA-Damage and Stress-Induced Cellular Senescence Response Phenotype. Exp. Gerontol. 2007, 42, 911–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.-Y.; Lee, H.-C.; Fahn, H.-J.; Wei, Y.-H. Oxidative Damage Elicited by Imbalance of Free Radical Scavenging Enzymes Is Associated with Large-Scale MtDNA Deletions in Aging Human Skin. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 1999, 423, 11–21. [Google Scholar] [CrossRef]
- Meewes, C.; Brenneisen, P.; Wenk, J.; Kuhr, L.; Ma, W.; Alikoski, J.; Poswig, A.; Krieg, T.; Scharffetter-Kochanek, K. Adaptive Antioxidant Response Protects Dermal Fibroblasts from UVA-Induced Phototoxicity. Free Radic. Biol. Med. 2001, 30, 238–247. [Google Scholar] [CrossRef]
- Naderi-Hachtroudi, L.; Peters, T.; Brenneisen, P.; Meewes, C.; Hommel, C.; Razi-Wolf, Z.; Schneider, L.A.; Schüller, J.; Wlaschek, M.; Scharffetter-Kochanek, K. Induction of Manganese Superoxide Dismutase in Human Dermal Fibroblasts. Arch. Dermatol. 2002, 138, 1473–1479. [Google Scholar] [CrossRef]
- Mao, C.; Yuan, J.-Q.; Lv, Y.-B.; Gao, X.; Yin, Z.-X.; Kraus, V.B.; Luo, J.-S.; Chei, C.-L.; Matchar, D.B.; Zeng, Y.; et al. Associations between Superoxide Dismutase, Malondialdehyde and All-Cause Mortality in Older Adults: A Community-Based Cohort Study. BMC Geriatr. 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Mohammedi, K.; Bellili-Muñoz, N.; Marklund, S.L.; Driss, F.; le Nagard, H.; Patente, T.A.; Fumeron, F.; Roussel, R.; Hadjadj, S.; Marre, M.; et al. Plasma Extracellular Superoxide Dismutase Concentration, Allelic Variations in the SOD3 Gene and Risk of Myocardial Infarction and All-Cause Mortality in People with Type 1 and Type 2 Diabetes. Cardiovasc. Diabetol. 2015, 14, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, H.; Passos, J.F. Cellular Senescence: All Roads Lead to Mitochondria. FEBS J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Treiber, N.; Maity, P.; Singh, K.; Kohn, M.; Keist, A.F.; Ferchiu, F.; Sante, L.; Frese, S.; Bloch, W.; Kreppel, F.; et al. Accelerated Aging Phenotype in Mice with Conditional Deficiency for Mitochondrial Superoxide Dismutase in the Connective Tissue. Aging Cell 2011, 10, 239–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Unnikrishnan, A.; Deepa, S.S.; Liu, Y.; Li, Y.; Ikeno, Y.; Sosnowska, D.; van Remmen, H.; Richardson, A. A New Role for Oxidative Stress in Aging: The Accelerated Aging Phenotype in Sod1− Mice Is Correlated to Increased Cellular Senescence. Redox Biol. 2017, 11, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef]
- Ganini, D.; Santos, J.H.; Bonini, M.G.; Mason, R.P. Switch of Mitochondrial Superoxide Dismutase into a Prooxidant Peroxidase in Manganese-Deficient Cells and Mice. Cell Chem. Biol. 2018, 25, 413–425.e6. [Google Scholar] [CrossRef] [Green Version]
- Naranuntarat, A.; Jensen, L.T.; Pazicni, S.; Penner-Hahn, J.E.; Culotta, V.C. The Interaction of Mitochondrial Iron with Manganese Superoxide Dismutase. J. Biol. Chem. 2009, 284, 22633–22640. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Cobine, P.A.; Molik, S.; Naranuntarat, A.; Lill, R.; Winge, D.R.; Culotta, V.C. The Effects of Mitochondrial Iron Homeostasis on Cofactor Specificity of Superoxide Dismutase 2. EMBO J. 2006, 25, 1775–1783. [Google Scholar] [CrossRef] [Green Version]
- Killilea, D.W.; Atamna, H.; Liao, C.; Ames, B.N. Iron Accumulation During Cellular Senescence in Human Fibroblasts In Vitro. Antioxid. Redox Signal. 2003, 5, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Killilea, D.W.; Maier, J.A.M. A Connection between Magnesium Deficiency and Aging: New Insights from Cellular Studies. Magnes Res. 2008, 21, 77–82. [Google Scholar] [PubMed]
- Cutler, R.G. Oxidative Stress and Aging: Catalase Is a Longevity Determinant Enzyme. Rejuvenation Res. 2005, 8, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Koepke, J.I.; Wood, C.S.; Terlecky, L.J.; Walton, P.A.; Terlecky, S.R. Progeric Effects of Catalase Inactivation in Human Cells. Toxicol. Appl. Pharmacol. 2008, 232, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Koepke, J.I.; Nakrieko, K.-A.; Wood, C.S.; Boucher, K.K.; Terlecky, L.J.; Walton, P.A.; Terlecky, S.R. Restoration of Peroxisomal Catalase Import in a Model of Human Cellular Aging. Traffic 2007, 8, 1590–1600. [Google Scholar] [CrossRef]
- Wood, C.S.; Koepke, J.I.; Teng, H.; Boucher, K.K.; Katz, S.; Chang, P.; Terlecky, L.J.; Papanayotou, I.; Walton, P.A.; Terlecky, S.R. Hypocatalasemic Fibroblasts Accumulate Hydrogen Peroxide and Display Age-Associated Pathologies. Traffic 2006, 7, 97–107. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial Dysfunction and Oxidative Stress in Neurodegenerative Diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef]
- Ahmed, E.; Donovan, T.; Yujiao, L.; Zhang, Q. Mitochondrial Targeted Antioxidant in Cerebral Ischemia. J. Neurol. Neurosci. 2015, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Tarry-Adkins, J.L.; Blackmore, H.L.; Martin-Gronert, M.S.; Fernandez-Twinn, D.S.; McConnell, J.M.; Hargreaves, I.P.; Giussani, D.A.; Ozanne, S.E. Coenzyme Q10 Prevents Accelerated Cardiac Aging in a Rat Model of Poor Maternal Nutrition and Accelerated Postnatal Growth. Mol. Metab. 2013, 2, 480–490. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Hebert, V.Y.; Stadler, K.; Xue, S.Y.; Slaybaugh, K.; Luttrell-Williams, E.; Glover, M.C.; Krzywanski, D.M.; Dugas, T.R. Coenzyme Q10 Alleviates Chronic Nucleoside Reverse Transcriptase Inhibitor-Induced Premature Endothelial Senescence. Cardiovasc. Toxicol. 2019, 19, 500–509. [Google Scholar] [CrossRef]
- Ma, D.; Stokes, K.; Mahngar, K.; Domazet-Damjanov, D.; Sikorska, M.; Pandey, S. Inhibition of Stress Induced Premature Senescence in Presenilin-1 Mutated Cells with Water Soluble Coenzyme Q10. Mitochondrion 2014, 17, 106–115. [Google Scholar] [CrossRef]
- Xue, R.; Yang, J.; Wu, J.; Meng, Q.; Hao, J. Coenzyme Q10 Inhibits the Activation of Pancreatic Stellate Cells through PI3K/AKT/MTOR Signaling Pathway. Oncotarget 2017, 8, 92300–92311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Liang, S.; Zhu, X.; Wu, X.; Dong, Z. CoQ10 Suppression of Oxidative Stress and Cell Senescence Increases Bone Mass in Orchiectomized Mice. Am. J. Transl. Res. 2020, 12, 4314–4325. [Google Scholar] [PubMed]
- Mine, Y.; Takahashi, T.; Okamoto, T. Protective Effects of Coenzyme Q10 on Cell Damage Induced by Hydrogen Peroxides in Cultured Skin Fibroblasts. J. Clin. Biochem. Nutr. 2021, 69, 20–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yan, B.; Yu, S.; Zhang, C.; Wang, B.; Wang, Y.; Wang, J.; Yuan, Z.; Zhang, L.; Pan, J. Coenzyme Q10 Inhibits the Aging of Mesenchymal Stem Cells Induced by D-Galactose through Akt/MTOR Signaling. Oxidative Med. Cell. Longev. 2015, 2015, 867293. [Google Scholar] [CrossRef] [PubMed]
- Velichkovska, M.; Surnar, B.; Nair, M.; Dhar, S.; Toborek, M. Targeted Mitochondrial COQ10 Delivery Attenuates Antiretroviral-Drug-Induced Senescence of Neural Progenitor Cells. Mol. Pharm. 2019, 16, 724–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcheggiani, F.; Kordes, S.; Cirilli, I.; Orlando, P.; Silvestri, S.; Vogelsang, A.; Möller, N.; Blatt, T.; Weise, J.M.; Damiani, E.; et al. Anti-Ageing Effects of Ubiquinone and Ubiquinol in a Senescence Model of Human Dermal Fibroblasts. Free Radic. Biol. Med. 2021, 165, 282–288. [Google Scholar] [CrossRef]
- Huo, J.; Xu, Z.; Hosoe, K.; Kubo, H.; Miyahara, H.; Dai, J.; Mori, M.; Sawashita, J.; Higuchi, K. Coenzyme Q10 Prevents Senescence and Dysfunction Caused by Oxidative Stress in Vascular Endothelial Cells. Oxidative Med. Cell. Longev. 2018, 2018, 3181759. [Google Scholar] [CrossRef]
- Yan, J.; Fujii, K.; Yao, J.; Kishida, H.; Hosoe, K.; Sawashita, J.; Takeda, T.; Mori, M.; Higuchi, K. Reduced Coenzyme Q10 Supplementation Decelerates Senescence in SAMP1 Mice. Exp. Gerontol. 2006, 41, 130–140. [Google Scholar] [CrossRef]
- Olivieri, F.; Lazzarini, R.; Babini, L.; Prattichizzo, F.; Rippo, M.R.; Tiano, L.; di Nuzzo, S.; Graciotti, L.; Festa, R.; Brugè, F.; et al. Anti-Inflammatory Effect of Ubiquinol-10 on Young and Senescent Endothelial Cells via MiR-146a Modulation. Free Radic. Biol. Med. 2013, 63, 410–420. [Google Scholar] [CrossRef]
- Maruoka, H.; Fujii, K.; Inoue, K.; Kido, S. Long-Term Effect of Ubiquinol on Exercise Capacity and the Oxidative Stress Regulation System in SAMP1 Mice. J. Phys. Ther. Sci. 2014, 26, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Schmelzer, C.; Kubo, H.; Mori, M.; Sawashita, J.; Kitano, M.; Hosoe, K.; Boomgaarden, I.; Döring, F.; Higuchi, K. Supplementation with the Reduced Form of Coenzyme Q10 Decelerates Phenotypic Characteristics of Senescence and Induces a Peroxisome Proliferator-Activated Receptor-α Gene Expression Signature in SAMP1 Mice. Mol. Nutr. Food Res. 2010, 54, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, I.; Orlando, P.; Marcheggiani, F.; Dludla, P.V.; Silvestri, S.; Damiani, E.; Tiano, L. The Protective Role of Bioactive Quinones in Stress-Induced Senescence Phenotype of Endothelial Cells Exposed to Cigarette Smoke Extract. Antioxidants 2020, 9, 1008. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Sawashita, J.; Kubo, H.; Nishio, S.; Hashimoto, S.; Suzuki, N.; Yoshimura, H.; Tsuruoka, M.; Wang, Y.; Liu, Y.; et al. Ubiquinol-10 Supplementation Activates Mitochondria Functions to Decelerate Senescence in Senescence-Accelerated Mice. Antioxid. Redox Signal. 2014, 20, 2606–2620. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Bakeeva, L.E.; Egormin, P.A.; Filenko, O.F.; Isakova, E.F.; Manskikh, V.N.; Mikhelson, V.M.; Panteleeva, A.A.; Pasyukova, E.G.; Pilipenko, D.I.; et al. Mitochondria-Targeted Plastoquinone Derivatives as Tools to Interrupt Execution of the Aging Program. 5. SkQ1 Prolongs Lifespan and Prevents Development of Traits of Senescence. Biochemistry 2008, 73, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, M.; Antonenko, Y.; Anisimov, V.; Chernyak, B.; Cherepanov, D.; Chistyakov, V.; Egorov, M.; Kolosova, N.; Korshunova, G.; Lyamzaev, K.; et al. Mitochondrial-Targeted Plastoquinone Derivatives. Effect on Senescence and Acute Age-Related Pathologies. Curr. Drug Targets 2011, 12, 800–826. [Google Scholar] [CrossRef]
- Kolosova, N.G.; Stefanova, N.A.; Muraleva, N.A.; Skulachev, V.P. The Mitochondria-Targeted Antioxidant SkQ1 but Not N-Acetylcysteine Reverses Aging-Related Biomarkers in Rats. Aging 2012, 4, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Manskikh, V.N.; Gancharova, O.S.; Nikiforova, A.I.; Krasilshchikova, M.S.; Shabalina, I.G.; Egorov, M.V.; Karger, E.M.; Milanovsky, G.E.; Galkin, I.I.; Skulachev, V.P.; et al. Age-Associated Murine Cardiac Lesions Are Attenuated by the Mitochondria-Targeted Antioxidant SkQ1. Histol. Histopathol. 2015, 30, 353–360. [Google Scholar] [CrossRef]
- Stefanova, N.A.; Muraleva, N.A.; Skulachev, V.P.; Kolosova, N.G. Alzheimer’s Disease-Like Pathology in Senescence-Accelerated OXYS Rats Can Be Partially Retarded with Mitochondria-Targeted Antioxidant SkQ1. J. Alzheimer’s Dis. 2013, 38, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Atamna, H.; Nguyen, A.; Schultz, C.; Boyle, K.; Newberry, J.; Kato, H.; Ames, B.N. Methylene Blue Delays Cellular Senescence and Enhances Key Mitochondrial Biochemical Pathways. FASEB J. 2008, 22, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.-M.; O’Donovan, M.; Sun, L.; Choi, J.Y.; Ren, M.; Cao, K. Anti-Aging Potentials of Methylene Blue for Human Skin Longevity. Sci. Rep. 2017, 7, 2475. [Google Scholar] [CrossRef]
- Bertolo, A.; Capossela, S.; Fränkl, G.; Baur, M.; Pötzel, T.; Stoyanov, J. Oxidative Status Predicts Quality in Human Mesenchymal Stem Cells. Stem Cell Res. Ther. 2017, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daudt, D.R.; Mueller, B.; Park, Y.H.; Wen, Y.; Yorio, T. Methylene Blue Protects Primary Rat Retinal Ganglion Cells from Cellular Senescence. Investig. Opthalmol. Vis. Sci. 2012, 53, 4657. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, I.; Heaton, R.A.; Mantle, D. Disorders of Human Coenzyme Q10 Metabolism: An Overview. Int. J. Mol. Sci. 2020, 21, 6695. [Google Scholar] [CrossRef]
- Yubero, D.; Montero, R.; Martín, M.A.; Montoya, J.; Ribes, A.; Grazina, M.; Trevisson, E.; Rodriguez-Aguilera, J.C.; Hargreaves, I.P.; Salviati, L.; et al. Secondary Coenzyme Q 10 Deficiencies in Oxidative Phosphorylation (OXPHOS) and Non-OXPHOS Disorders. Mitochondrion 2016, 30, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Mantle, D. Coenzyme Q10 Supplementation for Diabetes and Its Complications: An Overview. Br. J. Diabetes 2017, 17, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, I.; Mantle, D.; Milford, D. Chronic Kidney Disease and Coenzyme Q10 Supplementation. J. Kidney Care 2019, 4, 82–90. [Google Scholar] [CrossRef]
- Mantle, D.; Hargreaves, I. Coenzyme Q10 and Degenerative Disorders Affecting Longevity: An Overview. Antioxidants 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmanuele, V.; López, L.C.; Berardo, A.; Naini, A.; Tadesse, S.; Wen, B.; D’Agostino, E.; Solomon, M.; DiMauro, S.; Quinzii, C.; et al. Heterogeneity of Coenzyme Q10 Deficiency: Patient study and literature review. Arch. Neurol. 2012, 69, 978–983. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, T.; Li, S.; Zhang, X.; Ding, Q.; Que, H.; Yan, X.; Wei, K.; Liu, S. Comparative Proteomic Analysis of Brains of Naturally Aging Mice. Neuroscience 2008, 154, 1107–1120. [Google Scholar] [CrossRef]
- Marcheggiani, F.; Cirilli, I.; Orlando, P.; Silvestri, S.; Vogelsang, A.; Knott, A.; Blatt, T.; Weise, J.M.; Tiano, L. Modulation of Coenzyme Q10 Content and Oxidative Status in Human Dermal Fibroblasts Using HMG-CoA Reductase Inhibitor over a Broad Range of Concentrations. From Mitohormesis to Mitochondrial Dysfunction and Accelerated Aging. Aging 2019, 11, 2565–2582. [Google Scholar] [CrossRef]
- Bliznakov, E.G. Immunological Senescence in Mice and Its Reversal by Coenzyme Q10. Mech. Ageing Dev. 1978, 7, 189–197. [Google Scholar] [CrossRef]
- Sohal, R.S.; Kamzalov, S.; Sumien, N.; Ferguson, M.; Rebrin, I.; Heinrich, K.R.; Forster, M.J. Effect of Coenzyme Q10 Intake on Endogenous Coenzyme Q Content, Mitochondrial Electron Transport Chain, Antioxidative Defenses, and Life Span of Mice. Free Radic. Biol. Med. 2006, 40, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumien, N.; Heinrich, K.R.; Shetty, R.A.; Sohal, R.S.; Forster, M.J. Prolonged Intake of Coenzyme Q10 Impairs Cognitive Functions in Mice. J. Nutr. 2009, 139, 1926–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcdonald, S.R.; Sohal, R.S.; Forster, M.J. Concurrent Administration of Coenzyme Q10 and α-Tocopherol Improves Learning in Aged Mice. Free Radic. Biol. Med. 2005, 38, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Shetty, R.A.; Forster, M.J.; Sumien, N. Coenzyme Q10 Supplementation Reverses Age-Related Impairments in Spatial Learning and Lowers Protein Oxidation. Age 2013, 35, 1821–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreani, C.; Bartolacci, C.; Guescini, M.; Battistelli, M.; Stocchi, V.; Orlando, F.; Provinciali, M.; Amici, A.; Marchini, C.; Tiano, L.; et al. Combination of Coenzyme Q10 Intake and Moderate Physical Activity Counteracts Mitochondrial Dysfunctions in a SAMP8 Mouse Model. Oxidative Med. Cell. Longev. 2018, 2018, 8936251. [Google Scholar] [CrossRef] [Green Version]
- Skulachev, V.P.; Anisimov, V.N.; Antonenko, Y.N.; Bakeeva, L.E.; Chernyak, B.V.; Erichev, V.P.; Filenko, O.F.; Kalinina, N.I.; Kapelko, V.I.; Kolosova, N.G.; et al. An Attempt to Prevent Senescence: A Mitochondrial Approach. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 437–461. [Google Scholar] [CrossRef] [Green Version]
- Obukhova, L.A.; Skulachev, V.P.; Kolosova, N.G. Mitochondria-Targeted Antioxidant SkQ1 Inhibits Age-Dependent Involution of the Thymus in Normal and Senescence-Prone Rats. Aging 2009, 1, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Stefanova, N.A.; Ershov, N.I.; Kolosova, N.G. Suppression of Alzheimer’s Disease-Like Pathology Progression by Mitochondria-Targeted Antioxidant SkQ1: A Transcriptome Profiling Study. Oxidative Med. Cell. Longev. 2019, 2019, 3984906. [Google Scholar] [CrossRef] [Green Version]
- Muraleva, N.A.; Stefanova, N.A.; Kolosova, N.G. SkQ1 Suppresses the P38 MAPK Signaling Pathway Involved in Alzheimer’s Disease-Like Pathology in OXYS Rats. Antioxidants 2020, 9, 676. [Google Scholar] [CrossRef] [PubMed]
- Loshchenova, P.S.; Sinitsyna, O.I.; Fedoseeva, L.A.; Stefanova, N.A.; Kolosova, N.G. Influence of Antioxidant SkQ1 on Accumulation of Mitochondrial DNA Deletions in the Hippocampus of Senescence-Accelerated OXYS Rats. Biochemistry 2015, 80, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Ježek, J.; Engstová, H.; Ježek, P. Antioxidant Mechanism of Mitochondria-Targeted Plastoquinone SkQ1 Is Suppressed in Aglycemic HepG2 Cells Dependent on Oxidative Phosphorylation. Biochim. Biophys. Acta (BBA)-Bioenerg. 2017, 1858, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Thaivalappil, A.; Cao, K. The Potentials of Methylene Blue as an Anti-Aging Drug. Cells 2021, 10, 3379. [Google Scholar] [CrossRef] [PubMed]
- Atamna, H.; Atamna, W.; Al-Eyd, G.; Shanower, G.; Dhahbi, J.M. Combined Activation of the Energy and Cellular-Defense Pathways May Explain the Potent Anti-Senescence Activity of Methylene Blue. Redox Biol. 2015, 6, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of Antioxidants on the Fibroblast Replicative Lifespan In Vitro. Oxidative Med. Cell. Longev. 2020, 2020, 6423783. [Google Scholar] [CrossRef]
- Dawson, M.I. The Importance of Vitamin A in Nutrition. Curr Pharm Des. 2000, 6, 311–325. [Google Scholar] [CrossRef]
- Bar-El Dadon, S.; Reifen, R. Vitamin A and the Epigenome. Crit. Rev. Food Sci. Nutr. 2017, 57, 2404–2411. [Google Scholar] [CrossRef]
- Shudo, K.; Fukasawa, H.; Nakagomi, M.; Yamagata, N. Towards Retinoid Therapy for Alzheimers Disease. Curr. Alzheimer Res. 2009, 6, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Kitaoka, K.; Shimizu, N.; Ono, K.; Chikahisa, S.; Nakagomi, M.; Shudo, K.; Ishimura, K.; Séi, H.; Yoshizaki, K. The Retinoic Acid Receptor Agonist Am80 Increases Hippocampal ADAM10 in Aged SAMP8 Mice. Neuropharmacology 2013, 72, 58–65. [Google Scholar] [CrossRef]
- Fukasawa, H.; Nakagomi, M.; Yamagata, N.; Katsuki, H.; Kawahara, K.; Kitaoka, K.; Miki, T.; Shudo, K. Tamibarotene: A Candidate Retinoid Drug for Alzheimer’s Disease. Biol. Pharm. Bull. 2012, 35, 1206–1212. [Google Scholar] [CrossRef] [Green Version]
- Malaspina, A.; Michael-Titus, A.T. Is the Modulation of Retinoid and Retinoid-Associated Signaling a Future Therapeutic Strategy in Neurological Trauma and Neurodegeneration? J. Neurochem. 2008, 104, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Liang, W. ASCs-Derived Exosomes Loaded with Vitamin A and Quercetin Inhibit Rapid Senescence-like Response after Acute Liver Injury. Biochem. Biophys. Res. Commun. 2021, 572, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.A. Vitamin C in Human Health and Disease Is Still a Mystery? An Overview. Nutr. J. 2003, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Padayatty, S.J.; Katz, A.; Wang, Y.; Eck, P.; Kwon, O.; Lee, J.-H.; Chen, S.; Corpe, C.; Dutta, A.; Dutta, S.K.; et al. Vitamin C as an Antioxidant: Evaluation of Its Role in Disease Prevention. J. Am. Coll. Nutr. 2003, 22, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Monacelli, F.; Acquarone, E.; Giannotti, C.; Borghi, R.; Nencioni, A. Vitamin C, Aging and Alzheimer’s Disease. Nutrients 2017, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Ammar, M.A.; Ammar, A.A.; Condeni, M.S.; Bell, C.M. Vitamin C for Sepsis and Septic Shock. Am. J. 2021, 28, e649–e679. [Google Scholar] [CrossRef]
- Kashiouris, M.G.; L’Heureux, M.; Cable, C.A.; Fisher, B.J.; Leichtle, S.W.; Fowler, A.A. The Emerging Role of Vitamin C as a Treatment for Sepsis. Nutrients 2020, 12, 292. [Google Scholar] [CrossRef] [Green Version]
- Teng, J.; Pourmand, A.; Mazer-Amirshahi, M. Vitamin C: The next Step in Sepsis Management? J. Crit. Care 2018, 43, 230–234. [Google Scholar] [CrossRef]
- Fritz, H.; Flower, G.; Weeks, L.; Cooley, K.; Callachan, M.; McGowan, J.; Skidmore, B.; Kirchner, L.; Seely, D. Intravenous Vitamin C and Cancer. Integr. Cancer Ther. 2014, 13, 280–300. [Google Scholar] [CrossRef]
- van Gorkom, G.N.Y.; Lookermans, E.L.; van Elssen, C.H.M.J.; Bos, G.M.J. The Effect of Vitamin C (Ascorbic Acid) in the Treatment of Patients with Cancer: A Systematic Review. Nutrients 2019, 11, 977. [Google Scholar] [CrossRef] [Green Version]
- Uchio, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Ishigami, A. High Dietary Intake of Vitamin C Suppresses Age-Related Thymic Atrophy and Contributes to the Maintenance of Immune Cells in Vitamin C-Deficient Senescence Marker Protein-30 Knockout Mice. Br. J. Nutr. 2015, 113, 603–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.-S.; Park, S.-H.; Kim, H.-S.; Kang, H.-J.; Kim, M.-J.; Oh, S.-J.; Park, J.-B.; Kim, J.; Kim, S.C.; Lee, J.-Y. Ascorbic Acid Extends Replicative Life Span of Human Embryonic Fibroblast by Reducing DNA and Mitochondrial Damages. Nutr. Res. Pract. 2007, 1, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furumoto, K.; Inoue, E.; Nagao, N.; Hiyama, E.; Miwa, N. Age-Dependent Telomere Shortening Is Slowed down by Enrichment of Intracellular Vitamin C via Suppression of Oxidative Stress. Life Sci. 1998, 63, 935–948. [Google Scholar] [CrossRef]
- Chang, Z.; Huo, L.; Li, P.; Wu, Y.; Zhang, P. Ascorbic Acid Provides Protection for Human Chondrocytes against Oxidative Stress. Mol. Med. Rep. 2015, 12, 7086–7092. [Google Scholar] [CrossRef] [Green Version]
- Burger, M.; Steinitz, A.; Geurts, J.; Pippenger, B.; Schaefer, D.; Martin, I.; Barbero, A.; Pelttari, K. Ascorbic Acid Attenuates Senescence of Human Osteoarthritic Osteoblasts. Int. J. Mol. Sci. 2017, 18, 2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-M.; Lim, S.-M.; Yoo, J.-A.; Woo, M.-J.; Cho, K.-H. Consumption of High-Dose Vitamin C (1250 Mg per Day) Enhances Functional and Structural Properties of Serum Lipoprotein to Improve Anti-Oxidant, Anti-Atherosclerotic, and Anti-Aging Effects via Regulation of Anti-Inflammatory MicroRNA. Food Funct. 2015, 6, 3604–3612. [Google Scholar] [CrossRef]
- Cho, K.-H. Biomedicinal Implications of High-Density Lipoprotein: Its Composition, Structure, Functions, and Clinical Applications. BMB Rep. 2009, 42, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, G.; Bacchetti, T.; Nègre-Salvayre, A.; Salvayre, R.; Dousset, N.; Curatola, G. Structural Modifications of HDL and Functional Consequences. Atherosclerosis 2006, 184, 1–7. [Google Scholar] [CrossRef]
- Taniguchi, M.; Arai, N.; Kohno, K.; Ushio, S.; Fukuda, S. Anti-Oxidative and Anti-Aging Activities of 2-O-α-Glucopyranosyl-L-Ascorbic Acid on Human Dermal Fibroblasts. Eur. J. Pharm. 2012, 674, 126–131. [Google Scholar] [CrossRef]
- Burton, G.W.; Traber, M.G. Vitamin E: Antioxidant Activity, Biokinetics, and Bioavailability. Annu. Rev. Nutr. 1990, 10, 357–382. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Gammon, M.D.; Terry, M.B.; Wang, Q.; Bradshaw, P.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M. Telomere Length, Oxidative Damage, Antioxidants and Breast Cancer Risk. Int. J. Cancer 2009, 124, 1637–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corina, A.; Rangel-Zúñiga, O.A.; Jiménez-Lucena, R.; Alcalá-Díaz, J.F.; Quintana-Navarro, G.; Yubero-Serrano, E.M.; López-Moreno, J.; Delgado-Lista, J.; Tinahones, F.; Ordovás, J.M.; et al. Low Intake of Vitamin E Accelerates Cellular Aging in Patients With Established Cardiovascular Disease: The CORDIOPREV Study. J. Gerontol. Ser. A 2019, 74, 770–777. [Google Scholar] [CrossRef]
- Shearer, M.J.; Newman, P. Recent Trends in the Metabolism and Cell Biology of Vitamin K with Special Reference to Vitamin K Cycling and MK-4 Biosynthesis. J. Lipid Res. 2014, 55, 345–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaisar, M.A.; Prasad, S.; Cucullo, L. Protecting the BBB Endothelium against Cigarette Smoke-Induced Oxidative Stress Using Popular Antioxidants: Are They Really Beneficial? Brain Res. 2015, 1627, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, A.; Santoro, A.; Pini, E.; Cevenini, E.; Ostan, R.; Pietruszka, B.; Rolf, K.; Cano, N.; Caille, A.; Lyon-Belgy, N.; et al. A Parallel Randomized Trial on the Effect of a Healthful Diet on Inflammageing and Its Consequences in European Elderly People: Design of the NU-AGE Dietary Intervention Study. Mech. Ageing Dev. 2013, 134, 523–530. [Google Scholar] [CrossRef]
- Santoro, A.; Pini, E.; Scurti, M.; Palmas, G.; Berendsen, A.; Brzozowska, A.; Pietruszka, B.; Szczecinska, A.; Cano, N.; Meunier, N.; et al. Combating Inflammaging through a Mediterranean Whole Diet Approach: The NU-AGE Project’s Conceptual Framework and Design. Mech. Ageing Dev. 2014, 136–137, 3–13. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Shirakawa, H.; Miura, A.; Giriwono, P.E.; Sato, S.; Ohashi, A.; Iribe, M.; Goto, T.; Komai, M. Vitamin K Suppresses the Lipopolysaccharide-Induced Expression of Inflammatory Cytokines in Cultured Macrophage-like Cells via the Inhibition of the Activation of Nuclear Factor ΚB through the Repression of IKKα/β Phosphorylation. J. Nutr. Biochem. 2010, 21, 1120–1126. [Google Scholar] [CrossRef]
- Wei, N.; Lu, L.; Zhang, H.; Gao, M.; Ghosh, S.; Liu, Z.; Qi, J.; Wang, J.; Chen, J.; Huang, H. Warfarin Accelerates Aortic Calcification by Upregulating Senescence-Associated Secretory Phenotype Maker Expression. Oxidative Med. Cell. Longev. 2020, 2020, 2043762. [Google Scholar] [CrossRef]
- Nakano-Kurimoto, R.; Ikeda, K.; Uraoka, M.; Nakagawa, Y.; Yutaka, K.; Koide, M.; Takahashi, T.; Matoba, S.; Yamada, H.; Okigaki, M.; et al. Replicative Senescence of Vascular Smooth Muscle Cells Enhances the Calcification through Initiating the Osteoblastic Transition. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1673-84. [Google Scholar] [CrossRef]
- Young, A.; Lowe, G. Carotenoids—Antioxidant Properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Ross, A.C.; Caballero, B.H.; Cousins, R.J.; Tucker, K.L.; Ziegler, T.R. Modern Nutrition in Health and Disease, 11th ed.; Wolters Kluwer Health Adis (ESP): Alphen aan den Rijn, The Netherlands, 2012. [Google Scholar]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016, 2016, 3164312. [Google Scholar] [CrossRef] [Green Version]
- Rafi, M.M.; Kanakasabai, S.; Gokarn, S.V.; Krueger, E.G.; Bright, J.J. Dietary Lutein Modulates Growth and Survival Genes in Prostate Cancer Cells. J. Med. Food 2015, 18, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Zeng, F.; Li, B.; Lin, J.; Liang, Y.; Chen, Y. Higher Dietary Carotenoid Intake Associated with Lower Risk of Hip Fracture in Middle-Aged and Elderly Chinese: A Matched Case-Control Study. Bone 2018, 111, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Akbaraly, T.N.; Fontbonne, A.; Favier, A.; Berr, C. Plasma Carotenoids and Onset of Dysglycemia in an Elderly Population. Diabetes Care 2008, 31, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leermakers, E.T.; Darweesh, S.K.; Baena, C.P.; Moreira, E.M.; Melo van Lent, D.; Tielemans, M.J.; Muka, T.; Vitezova, A.; Chowdhury, R.; Bramer, W.M.; et al. The Effects of Lutein on Cardiometabolic Health across the Life Course: A Systematic Review and Meta-Analysis1,2. Am. J. Clin. Nutr. 2016, 103, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Sandmann, G. Carotenoids of Biotechnological Importance. Adv. Biochem. Eng. Biotechnol. 2014, 148, 449–467. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, Pharmacology and Treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Ames, B.N. Prolonging Healthy Aging: Longevity Vitamins and Proteins. Proc. Natl. Acad. Sci. USA 2018, 115, 10836–10844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mária, J.; Ingrid, Ž. Effects of Bioactive Compounds on Senescence and Components of Senescence Associated Secretory Phenotypes in Vitro. Food Funct. 2017, 8, 2394–2418. [Google Scholar] [CrossRef]
- Chisté, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids Inhibit Lipid Peroxidation and Hemoglobin Oxidation, but Not the Depletion of Glutathione Induced by ROS in Human Erythrocytes. Life Sci. 2014, 99, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Flori, E.; Mastrofrancesco, A.; Kovacs, D.; Ramot, Y.; Briganti, S.; Bellei, B.; Paus, R.; Picardo, M. 2,4,6-Octatrienoic Acid Is a Novel Promoter of Melanogenesis and Antioxidant Defence in Normal Human Melanocytes via PPAR-γ Activation. Pigment Cell Melanoma Res. 2011, 24, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Wertz, K.; Seifert, N.; Hunziker, P.B.; Riss, G.; Wyss, A.; Lankin, C.; Goralczyk, R. β-Carotene Inhibits UVA-Induced Matrix Metalloprotease 1 and 10 Expression in Keratinocytes by a Singlet Oxygen-Dependent Mechanism. Free Radic. Biol. Med. 2004, 37, 654–670. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and Pro-Oxidant Activities of Carotenoids and Their Oxidation Products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Feeney, J.; O’Leary, N.; Moran, R.; O’Halloran, A.M.; Nolan, J.M.; Beatty, S.; Young, I.S.; Kenny, R.A. Plasma Lutein and Zeaxanthin Are Associated With Better Cognitive Function Across Multiple Domains in a Large Population-Based Sample of Older Adults: Findings from The Irish Longitudinal Study on Aging. J. Gerontol. Ser. A 2017, 72, 1431–1436. [Google Scholar] [CrossRef] [Green Version]
- Li, L.H.; Lee, J.C.-Y.; Leung, H.H.; Lam, W.C.; Fu, Z.; Lo, A.C.Y. Lutein Supplementation for Eye Diseases. Nutrients 2020, 12, 1721. [Google Scholar] [CrossRef]
- Jia, Y.-P.; Sun, L.; Yu, H.-S.; Liang, L.-P.; Li, W.; Ding, H.; Song, X.-B.; Zhang, L.-J. The Pharmacological Effects of Lutein and Zeaxanthin on Visual Disorders and Cognition Diseases. Molecules 2017, 22, 610. [Google Scholar] [CrossRef]
- Stringham, J.M.; Johnson, E.J.; Hammond, B.R. Lutein across the Lifespan: From Childhood Cognitive Performance to the Aging Eye and Brain. Curr. Dev. Nutr. 2019, 3, nzz066. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.; Kochlik, B.; Demuth, I.; Steinhagen-Thiessen, E.; Grune, T.; Norman, K. Plasma Carotenoids, Tocopherols and Retinol-Association with Age in the Berlin Aging Study II. Redox Biol. 2020, 32, 101461. [Google Scholar] [CrossRef]
- Lindbergh, C.A.; Renzi-Hammond, L.M.; Hammond, B.R.; Terry, D.P.; Mewborn, C.M.; Puente, A.N.; Miller, L.S. Lutein and Zeaxanthin Influence Brain Function in Older Adults: A Randomized Controlled Trial. J. Int. Neuropsychol. Soc. 2018, 24, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mewborn, C.M.; Lindbergh, C.A.; Hammond, B.R.; Renzi-Hammond, L.M.; Miller, L.S. The Effects of Lutein and Zeaxanthin Supplementation on Brain Morphology in Older Adults: A Randomized, Controlled Trial. J. Aging Res. 2019, 2019, 3709402. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.; Park, S.; Park, G. Lutein Protects Human Retinal Pigment Epithelial Cells from Oxidative Stress-induced Cellular Senescence. Mol. Med. Rep. 2018, 18, 5182–5190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommerburg, O.; Keunen, J.E.E.; Bird, A.C.; van Kuijk, F.J.G.M. Fruits and Vegetables That Are Sources for Lutein and Zeaxanthin: The Macular Pigment in Human Eyes. Br. J. Ophthalmol. 1998, 82, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, S.; Knaus, M.; Suh, M. Associations between Lutein, Zeaxanthin, and Age-Related Macular Degeneration: An Overview. Crit. Rev. Food Sci. Nutr. 2009, 49, 313–326. [Google Scholar] [CrossRef]
- Paiva, S.A.R.; Russell, R.M. β-Carotene and Other Carotenoids as Antioxidants. J. Am. Coll. Nutr. 1999, 18, 426–433. [Google Scholar] [CrossRef]
- Leh, H.E.; Lee, L.K. Lycopene: A Potent Antioxidant for the Amelioration of Type II Diabetes Mellitus. Molecules 2022, 27, 2335. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, W.; Wang, X.; Zhao, K.; Negi, D.S.; Zhuo, L.; Qi, M.; Wang, X.; Zhang, X. Lycopene and Risk of Prostate Cancer. Medicine 2015, 94, e1260. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zeng, X.; Cheng, Y.; Tang, L.; Hong, D.; Yang, X. Lycopene Ameliorates Insulin Resistance and Increases Muscle Capillary Density in Aging via Activation of SIRT1. J. Nutr. Biochem. 2022, 99, 108862. [Google Scholar] [CrossRef]
- Chen, D.; Huang, C.; Chen, Z. A Review for the Pharmacological Effect of Lycopene in Central Nervous System Disorders. Biomed. Pharmacother. 2019, 111, 791–801. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.-S.; Han, Y.-S.; Lee, J.H.; Bae, I.; Yoon, Y.M.; Kwon, S.M.; Lee, S.H. Pretreatment with Lycopene Attenuates Oxidative Stress-Induced Apoptosis in Human Mesenchymal Stem Cells. Biomol. Ther. 2015, 23, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.-L.; Hu, M.-L.; Huang, C.-S. Lycopene Enhances UVA–Induced DNA Damage and Expression of Heme Oxygenase–1 in Cultured Mouse Embryo Fibroblasts. Eur. J. Nutr. 2005, 44, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, E. Photoprotection of Human Dermal Fibroblasts against Ultraviolet Light by Antioxidant Combinations Present in Tomato. Food Funct. 2014, 5, 285–290. [Google Scholar] [CrossRef]
- Liu, X.; DILXAT, T.; Shi, Q.; Qiu, T.; Lin, J. The Combination of Nicotinamide Mononucleotide and Lycopene Prevents Cognitive Impairment and Attenuates Oxidative Damage in D-Galactose Induced Aging Models via Keap1-Nrf2 Signaling. Gene 2022, 822, 146348. [Google Scholar] [CrossRef]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, H.; Zhen, C.; Liu, J.; Yang, P.; Hu, L.; Shang, P. Unraveling the Potential Role of Glutathione in Multiple Forms of Cell Death in Cancer Therapy. Oxidative Med. Cell. Longev. 2019, 2019, 3150145. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Fang, Y.-Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione Metabolism and Its Implications for Health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of Glutathione in Cancer Progression and Chemoresistance. Oxidative Med. Cell. Longev. 2013, 2013, 972913. [Google Scholar] [CrossRef] [Green Version]
- Staal, F.J.T.; Ela, S.W.; Roederer, M.; Anderson, M.T.; Herzenberg, L.A.; Herzenberg, L.A. Glutathione Deficiency and Human Immunodeficiency Virus Infection. Lancet 1992, 339, 909–912. [Google Scholar] [CrossRef]
- Waris, S.; Patel, A.; Ali, A.; Mahmood, R. Acetaldehyde-Induced Oxidative Modifications and Morphological Changes in Isolated Human Erythrocytes: An in Vitro Study. Environ. Sci. Pollut. Res. 2020, 27, 16268–16281. [Google Scholar] [CrossRef]
- Rusu, M.E.; Georgiu, C.; Pop, A.; Mocan, A.; Kiss, B.; Vostinaru, O.; Fizesan, I.; Stefan, M.-G.; Gheldiu, A.-M.; Mates, L.; et al. Antioxidant Effects of Walnut (Juglans regia L.) Kernel and Walnut Septum Extract in a D-Galactose-Induced Aging Model and in Naturally Aged Rats. Antioxidants 2020, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Morin, D.; Long, R.; Panel, M.; Laure, L.; Taranu, A.; Gueguen, C.; Pons, S.; Leoni, V.; Caccia, C.; Vatner, S.F.; et al. Hsp22 Overexpression Induces Myocardial Hypertrophy, Senescence and Reduced Life Span through Enhanced Oxidative Stress. Free Radic. Biol. Med. 2019, 137, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Armeni, T.; Ercolani, L.; Urbanelli, L.; Magini, A.; Magherini, F.; Pugnaloni, A.; Piva, F.; Modesti, A.; Emiliani, C.; Principato, G. Cellular Redox Imbalance and Changes of Protein S-Glutathionylation Patterns Are Associated with Senescence Induced by Oncogenic H-Ras. PLoS ONE 2012, 7, e52151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redondo, J.; Sarkar, P.; Kemp, K.; Heesom, K.J.; Wilkins, A.; Scolding, N.J.; Rice, C.M. Dysregulation of Mesenchymal Stromal Cell Antioxidant Responses in Progressive Multiple Sclerosis. Stem Cells Transl. Med. 2018, 7, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Gao, J.; Wei, T.; Shen, W. Angiotensin II-Induced Erythrocyte Senescence Contributes to Oxidative Stress. Rejuvenation Res. 2022, 25, 30–38. [Google Scholar] [CrossRef]
- Kurz, D.J.; Decary, S.; Hong, Y.; Trivier, E.; Akhmedov, A.; Erusalimsky, J.D. Chronic Oxidative Stress Compromises Telomere Integrity and Accelerates the Onset of Senescence in Human Endothelial Cells. J. Cell Sci. 2004, 117, 2417–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Johansson, E.; Fan, Y.; Shertzer, H.G.; Vasiliou, V.; Nebert, D.W.; Dalton, T.P. Early Onset Senescence Occurs When Fibroblasts Lack the Glutamate–Cysteine Ligase Modifier Subunit. Free Radic. Biol. Med. 2009, 47, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Probin, V.; Wang, Y.; Zhou, D. Busulfan-Induced Senescence Is Dependent on ROS Production Upstream of the MAPK Pathway. Free Radic. Biol. Med. 2007, 42, 1858–1865. [Google Scholar] [CrossRef] [Green Version]
- Fafián-Labora, J.A.; Rodríguez-Navarro, J.A.; O’Loghlen, A. Small Extracellular Vesicles Have GST Activity and Ameliorate Senescence-Related Tissue Damage. Cell Metab. 2020, 32, 71–86.e5. [Google Scholar] [CrossRef]
- Tong, J.; Fitzmaurice, P.S.; Moszczynska, A.; Mattina, K.; Ang, L.-C.; Boileau, I.; Furukawa, Y.; Sailasuta, N.; Kish, S.J. Do Glutathione Levels Decline in Aging Human Brain? Free Radic. Biol. Med. 2016, 93, 110–117. [Google Scholar] [CrossRef]
- Barilani, M.; Lovejoy, C.; Piras, R.; Abramov, A.Y.; Lazzari, L.; Angelova, P.R. Age-related Changes in the Energy of Human Mesenchymal Stem Cells. J. Cell. Physiol. 2022, 237, 1753–1767. [Google Scholar] [CrossRef] [PubMed]
- Martin-de-Pablos, A.; Córdoba-Fernández, A.; Fernández-Espejo, E. Analysis of Neurotrophic and Antioxidant Factors Related to Midbrain Dopamine Neuronal Loss and Brain Inflammation in the Cerebrospinal Fluid of the Elderly. Exp. Gerontol. 2018, 110, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mori, A. Age-Associated Changes in Superoxide Dismutase Activity, Thiobarbituric Acid Reactivity and Reduced Glutathione Level in the Brain and Liver in Senescence Accelerated Mice (SAM): A Comparison with DdY Mice. Mech. Ageing Dev. 1993, 71, 23–30. [Google Scholar] [CrossRef]
- Iskusnykh, I.Y.; Zakharova, A.A.; Pathak, D. Glutathione in Brain Disorders and Aging. Molecules 2022, 27, 324. [Google Scholar] [CrossRef]
- Barardo, D.; Thornton, D.; Thoppil, H.; Walsh, M.; Sharifi, S.; Ferreira, S.; Anžič, A.; Fernandes, M.; Monteiro, P.; Grum, T.; et al. The DrugAge Database of Aging-Related Drugs. Aging Cell 2017, 16, 594–597. [Google Scholar] [CrossRef] [Green Version]
- Rebrin, I.; Zicker, S.; Wedekind, K.J.; Paetau-Robinson, I.; Packer, L.; Sohal, R.S. Effect of Antioxidant-Enriched Diets on Glutathione Redox Status in Tissue Homogenates and Mitochondria of the Senescence-Accelerated Mouse. Free Radic. Biol. Med. 2005, 39, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Homma, T.; Fujii, J. Application of Glutathione as Anti-Oxidative and Anti-Aging Drugs. Curr. Drug Metab. 2015, 16, 560–571. [Google Scholar] [CrossRef]
- Kumar, P.; Osahon, O.W.; Sekhar, R.V. GlyNAC (Glycine and N-Acetylcysteine) Supplementation in Mice Increases Length of Life by Correcting Glutathione Deficiency, Oxidative Stress, Mitochondrial Dysfunction, Abnormalities in Mitophagy and Nutrient Sensing, and Genomic Damage. Nutrients 2022, 14, 1114. [Google Scholar] [CrossRef]
- Sekhar, R.V.; Patel, S.G.; Guthikonda, A.P.; Reid, M.; Balasubramanyam, A.; Taffet, G.E.; Jahoor, F. Deficient Synthesis of Glutathione Underlies Oxidative Stress in Aging and Can Be Corrected by Dietary Cysteine and Glycine Supplementation. Am. J. Clin. Nutr. 2011, 94, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Sekhar, R.V. GlyNAC Supplementation Improves Glutathione Deficiency, Oxidative Stress, Mitochondrial Dysfunction, Inflammation, Aging Hallmarks, Metabolic Defects, Muscle Strength, Cognitive Decline, and Body Composition: Implications for Healthy Aging. J. Nutr. 2021, 151, 3606–3616. [Google Scholar] [CrossRef]
- Ansary, J.; Forbes-Hernández, T.Y.; Gil, E.; Cianciosi, D.; Zhang, J.; Elexpuru-Zabaleta, M.; Simal-Gandara, J.; Giampieri, F.; Battino, M. Potential Health Benefit of Garlic Based on Human Intervention Studies: A Brief Overview. Antioxidants 2020, 9, 619. [Google Scholar] [CrossRef] [PubMed]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; G. Wasef, L.; Elewa, Y.H.A.; A. Al-Sagan, A.; Abd El-Hack, M.E.; Taha, A.E.; M. Abd-Elhakim, Y.; Prasad Devkota, H. Chemical Constituents and Pharmacological Activities of Garlic (Allium Sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elosta, A.; Slevin, M.; Rahman, K.; Ahmed, N. Aged Garlic Has More Potent Antiglycation and Antioxidant Properties Compared to Fresh Garlic Extract in Vitro. Sci. Rep. 2017, 7, 39613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, T.; Saito, H.; Nishiyama, N. Aged Garlic Extract Prolongs Longevity and Improves Spatial Memory Deficit in Senescence-Accelerated Mouse. Biol. Pharm. Bull. 1996, 19, 305–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, T.; Takashina, K.; Chu, P.; Saito, H.; Nishiyama, N. Prolongation of Life Span and Improved Learning in the Senescence Accelerated Mouse Produced by Aged Garlic Extract. Biol. Pharm. Bull. 1994, 17, 1589–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, H.; Higuchi, O.; Tateshita, K.; Tomobe, K.; Okuma, Y.; Nomura, Y. Antioxidative Activity and Ameliorative Effects of Memory Impairment of Sulfur-Containing Compounds in Allium Species. BioFactors 2006, 26, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Nakai, T.; Masutani, T.; Unno, K.; Akao, Y. Improvement of Learning and Memory in Senescence-Accelerated Mice by S-Allylcysteine in Mature Garlic Extract. Nutrients 2020, 12, 1834. [Google Scholar] [CrossRef]
- Chen, P.; Chang, C.; Lin, W.; Nagabhushanam, K.; Ho, C.; Pan, M. S-Allylcysteine Ameliorates Aging Features via Regulating Mitochondrial Dynamics in Naturally Aged C57BL/6J Mice. Mol. Nutr. Food Res. 2022, 66, 2101077. [Google Scholar] [CrossRef]
- Borek, C. Antioxidant Health Effects of Aged Garlic Extract. J. Nutr. 2001, 131, 1010S–1015S. [Google Scholar] [CrossRef] [Green Version]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA Maturation by Trans-Encoded Small RNA and Host Factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Kim, H. Protective Effect of Garlic on Cellular Senescence in UVB-Exposed HaCaT Human Keratinocytes. Nutrients 2016, 8, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, S.; Suzuki, J.; Takashima, M.; Ishida, M.; Kokubo, H.; Yoshizumi, M. S-1-Propenylcysteine Promotes IL-10-Induced M2c Macrophage Polarization through Prolonged Activation of IL-10R/STAT3 Signaling. Sci. Rep. 2021, 11, 22469. [Google Scholar] [CrossRef] [PubMed]
- Roumeliotis, S.; Roumeliotis, A.; Dounousi, E.; Eleftheriadis, T.; Liakopoulos, V. Dietary Antioxidant Supplements and Uric Acid in Chronic Kidney Disease: A Review. Nutrients 2019, 11, 1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Lozada, L.G. The Pathophysiology of Uric Acid on Renal Diseases. Contrib. Neprhol. 2018, 192, 17–24. [Google Scholar] [CrossRef]
- Kang, D.-H.; Ha, S.-K. Uric Acid Puzzle: Dual Role as Anti-Oxidantand Pro-Oxidant. Electrolytes Blood Press. 2014, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Vazirpanah, N.; Radstake, T.; Broen, J. Inflamm-Ageing and Senescence in Gout: The Tale of an Old King’s Disease. Curr. Aging Sci. 2015, 8, 186–201. [Google Scholar] [CrossRef]
- Goldberg, E.L.; Dixit, V.D. Drivers of Age-Related Inflammation and Strategies for Healthspan Extension. Immunol. Rev. 2015, 265, 63–74. [Google Scholar] [CrossRef]
- Fan, Y.; Xia, J.; Jia, D.; Zhang, M.; Zhang, Y.; Huang, G.; Wang, Y. Mechanism of Ginsenoside Rg1 Renal Protection in a Mouse Model of D-Galactose-Induced Subacute Damage. Pharm. Biol. 2016, 54, 1815–1821. [Google Scholar] [CrossRef]
- Cheong, K.; Lee, A. Guanine Deaminase Stimulates Ultraviolet-Induced Keratinocyte Senescence in Seborrhoeic Keratosis via Guanine Metabolites. Acta Derm. Venereol. 2020, 100, 1–10. [Google Scholar] [CrossRef]
- Yu, M.-A.; Sánchez-Lozada, L.G.; Johnson, R.J.; Kang, D.-H. Oxidative Stress with an Activation of the Renin-Angiotensin System in Human Vascular Endothelial Cells as a Novel Mechanism of Uric Acid-Induced Endothelial Dysfunction. J. Hypertens 2010, 28, 1234–1242. [Google Scholar] [CrossRef]
- Battelli, M.G.; Bortolotti, M.; Bolognesi, A.; Polito, L. Pro-Aging Effects of Xanthine Oxidoreductase Products. Antioxidants 2020, 9, 839. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-C.; Kuo, C.-S.; Wu, C.-C.; Tsai, H.-Y.; Lin, C.-P.; Li, S.-Y.; Chou, R.-H.; Huang, P.-H.; Chen, J.-W.; Lin, S.-J. Chronic Hyperuricemia Impairs Blood Flow Recovery in the Ischemic Hindlimb through Suppression of Endothelial Progenitor Cells. Oncotarget 2018, 9, 9285–9298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mladenov, M.; Gokik, M.; Hadzi-Petrushev, N.; Gjorgoski, I.; Jankulovski, N. The Relationship Between Antioxidant Enzymes and Lipid Peroxidation in Senescent Rat Erythrocytes. Physiol. Res. 2015, 64, 891. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Shin, D.G.; Kim, J.R.; Cho, K.H. Senescence-Related Truncation and Multimerization of Apolipoprotein A-I in High-Density Lipoprotein With an Elevated Level of Advanced Glycated End Products and Cholesteryl Ester Transfer Activity. J. Gerontol. Ser. A: Biomed. Sci. Med. Sci. 2010, 65, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.; Belwal, T.; Efferth, T.; Farooqi, A.A.; Sanches-Silva, A.; Vacca, R.A.; Nabavi, S.F.; Khan, F.; Prasad Devkota, H.; Barreca, D.; et al. Targeting Epigenetics in Cancer: Therapeutic Potential of Flavonoids. Crit. Rev. Food Sci. Nutr. 2021, 61, 1616–1639. [Google Scholar] [CrossRef]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as Antiobesity Agents: A Review. Med. Res. Rev. 2021, 41, 556–585. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, E.K.; Kim, D.H.; Yu, B.P.; Chung, H.Y. Kaempferol Modulates Pro-Inflammatory NF-ΚB Activation by Suppressing Advanced Glycation Endproducts-Induced NADPH Oxidase. Age 2010, 32, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Hua, Y.Q.; Zeng, Y.; Xu, J.; Xu, X. le Naringenin Alleviates Nonalcoholic Steatohepatitis in Middle-Aged Apoe−/−mice: Role of SIRT1. Phytomedicine 2021, 81, 153412. [Google Scholar] [CrossRef]
- Chattopadhyay, D.; Sen, S.; Chatterjee, R.; Roy, D.; James, J.; Thirumurugan, K. Context- and Dose-Dependent Modulatory Effects of Naringenin on Survival and Development of Drosophila Melanogaster. Biogerontology 2016, 17, 383–393. [Google Scholar] [CrossRef]
- Zhu, Y.; Tchkonia, T.; Pirtskhalava, T.; Gower, A.C.; Ding, H.; Giorgadze, N.; Palmer, A.K.; Ikeno, Y.; Hubbard, G.B.; Lenburg, M.; et al. The Achilles’ Heel of Senescent Cells: From Transcriptome to Senolytic Drugs. Aging Cell 2015, 14, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M.; Evans, S.A.; Fielder, E.; Victorelli, S.; Kruger, P.; Salmonowicz, H.; Weigand, B.M.; Patel, A.D.; Pirtskhalava, T.; Inman, C.L.; et al. Whole-body Senescent Cell Clearance Alleviates Age-related Brain Inflammation and Cognitive Impairment in Mice. Aging Cell 2021, 20, e13296. [Google Scholar] [CrossRef] [PubMed]
- Dookun, E.; Passos, J.F.; Arthur, H.M.; Richardson, G.D. Therapeutic Potential of Senolytics in Cardiovascular Disease. Cardiovasc. Drugs Ther. 2022, 36, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, M.J.; Zhu, Y.; McGowan, S.J.; Angelini, L.; Fuhrmann-Stroissnigg, H.; Xu, M.; Ling, Y.Y.; Melos, K.I.; Pirtskhalava, T.; Inman, C.L.; et al. Fisetin Is a Senotherapeutic That Extends Health and Lifespan. EBioMedicine 2018, 36, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Shen, L.; Yu, J.; Wan, H.; Guo, A.; Chen, J.; Long, Y.; Zhao, J.; Pei, G. Rapamycin and Other Longevity-Promoting Compounds Enhance the Generation of Mouse Induced Pluripotent Stem Cells. Aging Cell 2011, 10, 908–911. [Google Scholar] [CrossRef]
- Perrott, K.M.; Wiley, C.D.; Desprez, P.-Y.; Campisi, J. Apigenin Suppresses the Senescence-Associated Secretory Phenotype and Paracrine Effects on Breast Cancer Cells. Geroscience 2017, 39, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Ji, S.; Li, M.; Zheng, S.; Zhou, X.; Guo, H.; Deng, S.; Zhu, J.; Li, D.; Xie, Z. Theaflavin-Regulated Imd Condensates Control Drosophila Intestinal Homeostasis and Aging. iScience 2021, 24, 102150. [Google Scholar] [CrossRef]
- Xiao, Y.-Z.; Yang, M.; Xiao, Y.; Guo, Q.; Huang, Y.; Li, C.-J.; Cai, D.; Luo, X.-H. Reducing Hypothalamic Stem Cell Senescence Protects against Aging-Associated Physiological Decline. Cell Metab. 2020, 31, 534–548.e5. [Google Scholar] [CrossRef]
- Büchter, C.; Ackermann, D.; Havermann, S.; Honnen, S.; Chovolou, Y.; Fritz, G.; Kampkötter, A.; Wätjen, W. Myricetin-Mediated Lifespan Extension in Caenorhabditis Elegans Is Modulated by DAF-16. Int. J. Mol. Sci. 2013, 14, 11895–11914. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.-Y.; Lee, D.; Ryu, H.G.; Choi, B.-H.; Go, Y.; Lee, N.; Lee, D.; Son, H.G.; Jeon, J.; Kim, S.-H.; et al. Myricetin Improves Endurance Capacity and Mitochondrial Density by Activating SIRT1 and PGC-1α. Sci. Rep. 2017, 7, 6237. [Google Scholar] [CrossRef]
- Chattopadhyay, D.; Thirumurugan, K. Longevity-Promoting Efficacies of Rutin in High Fat Diet Fed Drosophila Melanogaster. Biogerontology 2020, 21, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, S.; Feng, T.; Dong, J.; Li, Y.; Li, H. Rutin Protects against Aging-Related Metabolic Dysfunction. Food Funct. 2016, 7, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-L.; Li, Y.-N.; Zhang, H.; Su, Y.-J.; Zhou, W.-W.; Zhang, Z.-P.; Wang, S.-W.; Xu, P.-X.; Wang, Y.-J.; Liu, R.-T. Rutin Inhibits Amylin-Induced Neurocytotoxicity and Oxidative Stress. Food Funct. 2015, 6, 3296–3306. [Google Scholar] [CrossRef] [PubMed]
- Burton, M.D.; Rytych, J.L.; Amin, R.; Johnson, R.W. Dietary Luteolin Reduces Proinflammatory Microglia in the Brain of Senescent Mice. Rejuvenation Res. 2016, 19, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Xiang, L.; Ishihara, S.; Matsuura, A.; Sakagami, Y.; Qi, J. Anti-Aging Effects of Hesperidin on Saccharomyces Cerevisiae via Inhibition of Reactive Oxygen Species and UTH1 Gene Expression. Biosci. Biotechnol. Biochem. 2012, 76, 640–645. [Google Scholar] [CrossRef] [Green Version]
- Elavarasan, J.; Velusamy, P.; Ganesan, T.; Ramakrishnan, S.K.; Rajasekaran, D.; Periandavan, K. Hesperidin-Mediated Expression of Nrf2 and Upregulation of Antioxidant Status in Senescent Rat Heart. J. Pharm. Pharmacol. 2012, 64, 1472–1482. [Google Scholar] [CrossRef]
- Fan, X.; Zeng, Y.; Fan, Z.; Cui, L.; Song, W.; Wu, Q.; Gao, Y.; Yang, D.; Mao, X.; Zeng, B.; et al. Dihydromyricetin Promotes Longevity and Activates the Transcription Factors FOXO and AOP in Drosophila. Aging 2021, 13, 460–476. [Google Scholar] [CrossRef]
- Qian, J.; Wang, X.; Cao, J.; Zhang, W.; Lu, C.; Chen, X. Dihydromyricetin Attenuates D-Galactose-Induced Brain Aging of Mice via Inhibiting Oxidative Stress and Neuroinflammation. Neurosci. Lett. 2021, 756, 135963. [Google Scholar] [CrossRef]
- Martínez-Coria, H.; Mendoza-Rojas, M.X.; Arrieta-Cruz, I.; López-Valdés, H.E. Preclinical Research of Dihydromyricetin for Brain Aging and Neurodegenerative Diseases. Front. Pharmacol. 2019, 10, 1334. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Sanchez, I.; Mansour, C.; Navarrete-Yañez, V.; Ayala-Hernandez, M.; Guevara, G.; Castillo, C.; Loredo, M.; Bustamante, M.; Ceballos, G.; Villarreal, F.J. (−)-Epicatechin Induced Reversal of Endothelial Cell Aging and Improved Vascular Function: Underlying Mechanisms. Food Funct. 2018, 9, 4802–4813. [Google Scholar] [CrossRef]
- Si, H.; Wang, X.; Zhang, L.; Parnell, L.D.; Ahmed, B.; LeRoith, T.; Ansah, T.-A.; Zhang, L.; Li, J.; Ordovás, J.M.; et al. Dietary Epicatechin Improves Survival and Delays Skeletal Muscle Degeneration in Aged Mice. FASEB J. 2019, 33, 965–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete-Yañez, V.; Garate-Carrillo, A.; Rodriguez, A.; Mendoza-Lorenzo, P.; Ceballos, G.; Calzada-Mendoza, C.; Hogan, M.C.; Villarreal, F.; Ramirez-Sanchez, I. Effects of (−)-Epicatechin on Neuroinflammation and Hyperphosphorylation of Tau in the Hippocampus of Aged Mice. Food Funct. 2020, 11, 10351–10361. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Kim, J.-R.; Choi, H.C. Genistein-Induced LKB1–AMPK Activation Inhibits Senescence of VSMC through Autophagy Induction. Vasc. Pharmacol. 2016, 81, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Uehara, Y.; Choi, Y.J.; Ha, Y.M.; Ye, B.H.; Yu, B.P.; Chung, H.Y. Mechanism of Attenuation of Pro-Inflammatory Ang II-Induced NF-ΚB Activation by Genistein in the Kidneys of Male Rats during Aging. Biogerontology 2011, 12, 537–550. [Google Scholar] [CrossRef]
- Bonet-Costa, V.; Herranz-Pérez, V.; Blanco-Gandía, M.; Mas-Bargues, C.; Inglés, M.; Garcia-Tarraga, P.; Rodriguez-Arias, M.; Miñarro, J.; Borras, C.; Garcia-Verdugo, J.M.; et al. Clearing Amyloid-β through PPARγ/ApoE Activation by Genistein Is a Treatment of Experimental Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 51, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Fan, Z.; Yang, Z.; Huang, T.; Tong, Y.; Yang, D.; Mao, X.; Yang, M. Flavonoids—Natural Gifts to Promote Health and Longevity. Int. J. Mol. Sci. 2022, 23, 2176. [Google Scholar] [CrossRef]
- Zhang, J.; Hong, Y.; Liuyang, Z.; Li, H.; Jiang, Z.; Tao, J.; Liu, H.; Xie, A.; Feng, Y.; Dong, X.; et al. Quercetin Prevents Radiation-Induced Oral Mucositis by Upregulating BMI-1. Oxidative Med. Cell. Longev. 2021, 2021, 2231680. [Google Scholar] [CrossRef]
- Smith, M.J.; Fowler, M.; Naftalin, R.J.; Siow, R.C.M. UVA Irradiation Increases Ferrous Iron Release from Human Skin Fibroblast and Endothelial Cell Ferritin: Consequences for Cell Senescence and Aging. Free Radic. Biol. Med. 2020, 155, 49–57. [Google Scholar] [CrossRef]
- Li, S.; Cao, H.; Shen, D.; Jia, Q.; Chen, C.; Xing, S. Quercetin Protects against Ox-LDL-induced Injury via Regulation of ABCAl, LXR-α and PCSK9 in RAW264.7 Macrophages. Mol. Med. Rep. 2018, 18, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Sohn, E.-J.; Kim, J.M.; Kang, S.-H.; Kwon, J.; An, H.J.; Sung, J.-S.; Cho, K.A.; Jang, I.-S.; Choi, J.-S. Restoring Effects of Natural Anti-Oxidant Quercetin on Cellular Senescent Human Dermal Fibroblasts. Am. J. Chin. Med. 2018, 46, 853–873. [Google Scholar] [CrossRef]
- Zoico, E.; Nori, N.; Darra, E.; Tebon, M.; Rizzatti, V.; Policastro, G.; de Caro, A.; Rossi, A.P.; Fantin, F.; Zamboni, M. Senolytic Effects of Quercetin in an in Vitro Model of Pre-Adipocytes and Adipocytes Induced Senescence. Sci. Rep. 2021, 11, 23237. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Fu, J.; Wu, W.; Ma, P.; Ren, L.; Yi, Z.; Wu, J. Quercetin Prevents Oxidative Stress-Induced Injury of Periodontal Ligament Cells and Alveolar Bone Loss in Periodontitis. Drug Des. Dev. Ther. 2021, 15, 3509–3522. [Google Scholar] [CrossRef] [PubMed]
- Zefzoufi, M.; Fdil, R.; Bouamama, H.; Gadhi, C.; Katakura, Y.; Mouzdahir, A.; Sraidi, K. Effect of Extracts and Isolated Compounds Derived from Retama Monosperma (L.) Boiss. on Anti-Aging Gene Expression in Human Keratinocytes and Antioxidant Activity. J. Ethnopharmacol. 2021, 280, 114451. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Kapeta, S.; Chinou, I.; Vassilatou, K.; Papassideri, I.; Gonos, E.S. Anti-Ageing and Rejuvenating Effects of Quercetin. Exp. Gerontol. 2010, 45, 763–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickson, L.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Corrigendum to ‘Senolytics Decrease Senescent Cells in Humans: Preliminary Report from a Clinical Trial of Dasatinib plus Quercetin in Individuals with Diabetic Kidney Disease’ EBioMedicine 47 (2019) 446–456. EBioMedicine 2020, 52, 102595. [Google Scholar] [CrossRef] [PubMed]
- Abharzanjani, F.; Hemmati, M. Protective Effects of Quercetin and Resveratrol on Aging Markers in Kidney under High Glucose Condition: In Vivo and in Vitro Analysis. Mol. Biol. Rep. 2021, 48, 5435–5442. [Google Scholar] [CrossRef]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics Improve Physical Function and Increase Lifespan in Old Age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef]
- Saccon, T.D.; Nagpal, R.; Yadav, H.; Cavalcante, M.B.; de Nunes, A.D.C.; Schneider, A.; Gesing, A.; Hughes, B.; Yousefzadeh, M.; Tchkonia, T.; et al. Senolytic Combination of Dasatinib and Quercetin Alleviates Intestinal Senescence and Inflammation and Modulates the Gut Microbiome in Aged Mice. J. Gerontol. Ser. A 2021, 76, 1895–1905. [Google Scholar] [CrossRef]
- Molagoda, I.M.N.; Kavinda, M.H.D.; Choi, Y.H.; Lee, H.; Kang, C.-H.; Lee, M.-H.; Lee, C.-M.; Kim, G.-Y. Fisetin Protects HaCaT Human Keratinocytes from Fine Particulate Matter (PM2.5)-Induced Oxidative Stress and Apoptosis by Inhibiting the Endoplasmic Reticulum Stress Response. Antioxidants 2021, 10, 1492. [Google Scholar] [CrossRef]
- Giri, S.; Takada, A.; Paudel, D.; Yoshida, K.; Furukawa, M.; Kuramitsu, Y.; Matsushita, K.; Abiko, Y.; Furuichi, Y. An in Vitro Senescence Model of Gingival Epithelial Cell Induced by Hydrogen Peroxide Treatment. Odontology 2022, 110, 44–53. [Google Scholar] [CrossRef]
- Singh, S.; Garg, G.; Singh, A.K.; Bissoyi, A.; Rizvi, S.I. Fisetin, a Potential Caloric Restriction Mimetic, Attenuates Senescence Biomarkers in Rat Erythrocytes. Biochem. Cell Biol. 2019, 97, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.J.; Bernier, M.; Mattison, J.A.; Aon, M.A.; Kaiser, T.A.; Anson, R.M.; Ikeno, Y.; Anderson, R.M.; Ingram, D.K.; de Cabo, R. Daily Fasting Improves Health and Survival in Male Mice Independent of Diet Composition and Calories. Cell Metab. 2019, 29, 221–228.e3. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Garg, G.; Singh, A.K.; Tripathi, S.S.; Rizvi, S.I. Fisetin, a Potential Caloric Restriction Mimetic, Modulates Ionic Homeostasis in Senescence Induced and Naturally Aged Rats. Arch. Physiol. Biochem. 2022, 128, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Boesten, D.M.P.H.J.; de Vos-Houben, J.M.J.; Timmermans, L.; den Hartog, G.J.M.; Bast, A.; Hageman, G.J. Accelerated Aging during Chronic Oxidative Stress: A Role for PARP-1. Oxidative Med. Cell. Longev. 2013, 2013, 680414. [Google Scholar] [CrossRef] [PubMed]
- Mas-Bargues, C.; Borrás, C.; Viña, J. Genistein, a Tool for Geroscience. Mech. Ageing Dev. 2022, 204, 111665. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Wu, W.; Chen, H.C.; Fang, H. Genistein Protects against UVB-Induced Senescence-like Characteristics in Human Dermal Fibroblast by P66Shc down-Regulation. J. Dermatol. Sci. 2010, 58, 19–27. [Google Scholar] [CrossRef]
- Zhang, H.; Pang, X.; Yu, H.; Zhou, H. Genistein Suppresses Ox-LDL-elicited Oxidative Stress and Senescence in HUVECs through the SIRT1-p66shc-Foxo3a Pathways. J. Biochem. Mol. Toxicol. 2022, 36, e22939. [Google Scholar] [CrossRef]
- Prasanth, M.; Sivamaruthi, B.; Chaiyasut, C.; Tencomnao, T. A Review of the Role of Green Tea (Camellia Sinensis) in Antiphotoaging, Stress Resistance, Neuroprotection, and Autophagy. Nutrients 2019, 11, 474. [Google Scholar] [CrossRef] [Green Version]
- Kochman, J.; Jakubczyk, K.; Antoniewicz, J.; Mruk, H.; Janda, K. Health Benefits and Chemical Composition of Matcha Green Tea: A Review. Molecules 2020, 26, 85. [Google Scholar] [CrossRef]
- Cabrera, C.; Artacho, R.; Giménez, R. Beneficial Effects of Green Tea—A Review. J. Am. Coll Nutr 2006, 25, 79–99. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wang, Y.; Li, D.; Chen, Y.; Qiao, X.; Fardous, R.; Lewandowski, A.; Liu, J.; Chan, T.-H.; Dou, Q.P. Perspectives on the Recent Developments with Green Tea Polyphenols in Drug Discovery. Expert Opin. Drug. Discov. 2018, 13, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Tea Polyphenols in Promotion of Human Health. Nutrients 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unno, K.; Takabayashi, F.; Yoshida, H.; Choba, D.; Fukutomi, R.; Kikunaga, N.; Kishido, T.; Oku, N.; Hoshino, M. Daily Consumption of Green Tea Catechin Delays Memory Regression in Aged Mice. Biogerontology 2007, 8, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Unno, K.; Ishikawa, Y.; Takabayashi, F.; Sasaki, T.; Takamori, N.; Iguchi, K.; Hoshino, M. Daily Ingestion of Green Tea Catechins from Adulthood Suppressed Brain Dysfunction in Aged Mice. Biofactors 2008, 34, 263–271. [Google Scholar] [CrossRef]
- Unno, K.; Takabayashi, F.; Kishido, T.; Oku, N. Suppressive Effect of Green Tea Catechins on Morphologic and Functional Regression of the Brain in Aged Mice with Accelerated Senescence (SAMP10). Exp. Gerontol. 2004, 39, 1027–1034. [Google Scholar] [CrossRef]
- Hsu, Y.-W.; Chen, W.-K.; Tsai, C.-F. Senescence-Mediated Redox Imbalance in Liver and Kidney: Antioxidant Rejuvenating Potential of Green Tea Extract. Int. J. Environ. Res. Public Health 2021, 19, 260. [Google Scholar] [CrossRef] [PubMed]
- Kishido, T.; Unno, K.; Yoshida, H.; Choba, D.; Fukutomi, R.; Asahina, S.; Iguchi, K.; Oku, N.; Hoshino, M. Decline in Glutathione Peroxidase Activity Is a Reason for Brain Senescence: Consumption of Green Tea Catechin Prevents the Decline in Its Activity and Protein Oxidative Damage in Ageing Mouse Brain. Biogerontology 2007, 8, 423–430. [Google Scholar] [CrossRef]
- Srividhya, R.; Jyothilakshmi, V.; Arulmathi, K.; Senthilkumaran, V.; Kalaiselvi, P. Attenuation of Senescence-induced Oxidative Exacerbations in Aged Rat Brain by (−)-epigallocatechin-3-gallate. Int. J. Dev. Neurosci. 2008, 26, 217–223. [Google Scholar] [CrossRef]
- Marrazzo, P.; Angeloni, C.; Freschi, M.; Lorenzini, A.; Prata, C.; Maraldi, T.; Hrelia, S. Combination of Epigallocatechin Gallate and Sulforaphane Counteracts In Vitro Oxidative Stress and Delays Stemness Loss of Amniotic Fluid Stem Cells. Oxidative Med. Cell. Longev. 2018, 2018, 5263985. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Jeon, H.-J.; Park, J.; Chang, M.-S. Epigallocatechin-3-Gallate Prevents Oxidative Stress-Induced Cellular Senescence in Human Mesenchymal Stem Cells via Nrf2. Int. J. Mol. Med. 2016, 38, 1075–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-C.; Liu, H.-W.; Chan, Y.-C.; Hu, S.-H.; Liu, M.-Y.; Chang, S.-J. The Green Tea Polyphenol Epigallocatechin-3-Gallate Attenuates Age-Associated Muscle Loss via Regulation of MiR-486-5p and Myostatin. Arch. Biochem. Biophys. 2020, 692, 108511. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Gu, C.; Chen, D.; Yu, B.; He, J. Oxidative Stress-Induced Diseases and Tea Polyphenols. Oncotarget 2017, 8, 81649–81661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanet, A.; Milenkovic, D.; Manach, C.; Mazur, A.; Morand, C. Citrus Flavanones: What Is Their Role in Cardiovascular Protection? J. Agric. Food Chem. 2012, 60, 8809–8822. [Google Scholar] [CrossRef]
- Chularojmontri, L.; Gerdprasert, O.; Wattanapitayakul, S.K. Pummelo Protects Doxorubicin-Induced Cardiac Cell Death by Reducing Oxidative Stress, Modifying Glutathione Transferase Expression, and Preventing Cellular Senescence. Evid.-Based Complement. Altern. Med. 2013, 2013, 254835. [Google Scholar] [CrossRef]
- Mulero, J.; Bernabé, J.; Cerdá, B.; García-Viguera, C.; Moreno, D.A.; Albaladejo, M.D.; Avilés, F.; Parra, S.; Abellán, J.; Zafrilla, P. Variations on Cardiovascular Risk Factors in Metabolic Syndrome after Consume of a Citrus-Based Juice. Clin. Nutr. 2012, 31, 372–377. [Google Scholar] [CrossRef]
- Buachan, P.; Chularojmontri, L.; Wattanapitayakul, S. Selected Activities of Citrus Maxima Merr. Fruits on Human Endothelial Cells: Enhancing Cell Migration and Delaying Cellular Aging. Nutrients 2014, 6, 1618–1634. [Google Scholar] [CrossRef] [Green Version]
- Testai, L.; Piragine, E.; Piano, I.; Flori, L.; da Pozzo, E.; Miragliotta, V.; Pirone, A.; Citi, V.; di Cesare Mannelli, L.; Brogi, S.; et al. The Citrus Flavonoid Naringenin Protects the Myocardium from Ageing-Dependent Dysfunction: Potential Role of SIRT1. Oxidative Med. Cell. Longev. 2020, 2020, 4650207. [Google Scholar] [CrossRef] [Green Version]
- da Pozzo, E.; Costa, B.; Cavallini, C.; Testai, L.; Martelli, A.; Calderone, V.; Martini, C. The Citrus Flavanone Naringenin Protects Myocardial Cells against Age-Associated Damage. Oxidative Med. Cell. Longev. 2017, 2017, 9536148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, B.; Chen, X.; Zhang, N.; Li, G.; Zhang, L.-H.; Tan, L.-Y. Naringenin Ameliorates Behavioral Dysfunction and Neurological Deficits in a D-Galactose-Induced Aging Mouse Model Through Activation of PI3K/Akt/Nrf2 Pathway. Rejuvenation Res. 2017, 20, 462–472. [Google Scholar] [CrossRef]
- Nakajima, A.; Aoyama, Y.; Nguyen, T.-T.L.; Shin, E.-J.; Kim, H.-C.; Yamada, S.; Nakai, T.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; et al. Nobiletin, a Citrus Flavonoid, Ameliorates Cognitive Impairment, Oxidative Burden, and Hyperphosphorylation of Tau in Senescence-Accelerated Mouse. Behav. Brain Res. 2013, 250, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Aoyama, Y.; Shin, E.-J.; Nam, Y.; Kim, H.-C.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; Yokoi, T.; Ohizumi, Y.; et al. Nobiletin, a Citrus Flavonoid, Improves Cognitive Impairment and Reduces Soluble Aβ Levels in a Triple Transgenic Mouse Model of Alzheimer’s Disease (3XTg-AD). Behav. Brain Res. 2015, 289, 69–77. [Google Scholar] [CrossRef] [PubMed]
- da Pozzo, E.; de Leo, M.; Faraone, I.; Milella, L.; Cavallini, C.; Piragine, E.; Testai, L.; Calderone, V.; Pistelli, L.; Braca, A.; et al. Antioxidant and Antisenescence Effects of Bergamot Juice. Oxidative Med. Cell. Longev. 2018, 2018, 9395804. [Google Scholar] [CrossRef]
- Kashyap, P.; Shikha, D.; Thakur, M.; Aneja, A. Functionality of Apigenin as a Potent Antioxidant with Emphasis on Bioavailability, Metabolism, Action Mechanism and in Vitro and in Vivo Studies: A Review. J. Food Biochem. 2022, 46, e13950. [Google Scholar] [CrossRef]
- Choi, S.; Youn, J.; Kim, K.; Joo, D.H.; Shin, S.; Lee, J.; Lee, H.K.; An, I.-S.; Kwon, S.; Youn, H.J.; et al. Apigenin Inhibits UVA-Induced Cytotoxicity in Vitro and Prevents Signs of Skin Aging in Vivo. Int. J. Mol. Med. 2016, 38, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.S.; Zhu, R.Z.; Lim, S.-H.; Seo, J.H.; Choi, B.-M. Apigenin Alleviates Oxidative Stress-Induced Cellular Senescence via Modulation of the SIRT1-NAD+-CD38 Axis. Am. J. Chin. Med. 2021, 49, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Zhang, F.; Wang, H.; Yao, J.; Chen, R.; Zhou, Z.; Yang, K.; Xie, Y.; Wan, T.; Ding, H. Apigenin Exhibits Protective Effects in a Mouse Model of D-Galactose-Induced Aging via Activating the Nrf2 Pathway. Food Funct. 2017, 8, 2331–2340. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Hunter, K.D.; Fonseca, F.P.; Radhakrishnan, R. Emerging Role of Cellular Senescence in the Pathogenesis of Oral Submucous Fibrosis and Its Malignant Transformation. Head Neck 2021, 43, 3153–3164. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, R.; Zhao, H.; Chen, G.; Jiang, Y.; Lyu, X.; Wu, T. Reduction of Aging-Induced Oxidative Stress and Activation of Autophagy by Bilberry Anthocyanin Supplementation via the AMPK–MTOR Signaling Pathway in Aged Female Rats. J. Agric. Food Chem. 2019, 67, 7832–7843. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wu, Y.; He, D.; Zhu, X.; Li, H.; Liu, H.; Liu, H. Anti-Aging Effects of Ribes Meyeri Anthocyanins on Neural Stem Cells and Aging Mice. Aging 2020, 12, 17738–17753. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-H.; Chan, Y.-C.; Liao, J.-W.; Wang, M.-F.; Yen, G.-C. Antioxidant and Cognitive Promotion Effects of Anthocyanin-Rich Mulberry (Morus Atropurpurea L.) on Senescence-Accelerated Mice and Prevention of Alzheimer’s Disease. J. Nutr. Biochem. 2010, 21, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Hoang, T.; Jung, E.; Jung, S.; Han, S.; Chung, M.; Chae, S.; Chae, H. Anthocyanins Attenuate Endothelial Dysfunction through Regulation of Uncoupling of Nitric Oxide Synthase in Aged Rats. Aging Cell 2020, 19, e13279. [Google Scholar] [CrossRef]
- Parzonko, A.; Oświt, A.; Bazylko, A.; Naruszewicz, M. Anthocyans-Rich Aronia Melanocarpa Extract Possesses Ability to Protect Endothelial Progenitor Cells against Angiotensin II Induced Dysfunction. Phytomedicine 2015, 22, 1238–1246. [Google Scholar] [CrossRef]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health Benefits of the Mediterranean Diet: Metabolic and Molecular Mechanisms. J. Gerontol. Ser. A 2018, 73, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Gantenbein, K.V.; Kanaka-Gantenbein, C. Mediterranean Diet as an Antioxidant: The Impact on Metabolic Health and Overall Wellbeing. Nutrients 2021, 13, 1951. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Li, J. Resveratrol Protects against Oxidized Low-Density Lipoprotein-Induced Human Umbilical Vein Endothelial Cell Apoptosis via Inhibition of Mitochondrial-Derived Oxidative Stress. Mol. Med. Rep. 2017, 15, 2457–2464. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.-S.; Zhang, Z.-S.; Yang, B.; He, W. Resveratrol Attenuates Oxidative Damage and Ameliorates Cognitive Impairment in the Brain of Senescence-Accelerated Mice. Life Sci. 2012, 91, 872–877. [Google Scholar] [CrossRef]
- Tung, B.T.; Rodríguez-Bies, E.; Talero, E.; Gamero-Estévez, E.; Motilva, V.; Navas, P.; López-Lluch, G. Anti-Inflammatory Effect of Resveratrol in Old Mice Liver. Exp. Gerontol. 2015, 64, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rahimifard, M.; Baeeri, M.; Bahadar, H.; Moini-Nodeh, S.; Khalid, M.; Haghi-Aminjan, H.; Mohammadian, H.; Abdollahi, M. Therapeutic Effects of Gallic Acid in Regulating Senescence and Diabetes; an In Vitro Study. Molecules 2020, 25, 5875. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Park, S.-Y.; Lee, H.J.; Lee, T.Y.; Sun, Z.; Yi, T.H. Gallic Acid Regulates Skin Photoaging in UVB-Exposed Fibroblast and Hairless Mice. Phytother. Res. 2014, 28, 1778–1788. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Zhou, B. Antioxidative, Anti-Inflammatory and Anti-Apoptotic Effects of Ellagic Acid in Liver and Brain of Rats Treated by D-Galactose. Sci. Rep. 2018, 8, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, H.J.; Kim, K.B.; Bae, S.; Choi, B.G.; An, S.; Ahn, K.J.; Kim, S.Y. Pretreatment of Ferulic Acid Protects Human Dermal Fibroblasts against Ultraviolet A Irradiation. Ann. Dermatol. 2016, 28, 740. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; You, Y.; Xi, Y.; Ni, B.; Chu, X.; Zhang, R.; You, H. P-Coumaric Acid Attenuates IL-1β-Induced Inflammatory Responses and Cellular Senescence in Rat Chondrocytes. Inflammation 2020, 43, 619–628. [Google Scholar] [CrossRef]
- Shen, Y.; Song, X.; Li, L.; Sun, J.; Jaiswal, Y.; Huang, J.; Liu, C.; Yang, W.; Williams, L.; Zhang, H.; et al. Protective Effects of P-Coumaric Acid against Oxidant and Hyperlipidemia-an in Vitro and in Vivo Evaluation. Biomed. Pharmacother. 2019, 111, 579–587. [Google Scholar] [CrossRef]
- Sun, C.Y.; Qi, S.S.; Zhou, P.; Cui, H.R.; Chen, S.X.; Dai, K.Y.; Tang, M.L. Neurobiological and Pharmacological Validity of Curcumin in Ameliorating Memory Performance of Senescence-Accelerated Mice. Pharmacol. Biochem. Behav. 2013, 105, 76–82. [Google Scholar] [CrossRef]
- Takano, K.; Tatebe, J.; Washizawa, N.; Morita, T. Curcumin Inhibits Age-Related Vascular Changes in Aged Mice Fed a High-Fat Diet. Nutrients 2018, 10, 1476. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Lin, S.; Yu, Y.; Chien, M.; Su, K.; Lin, C.; Way, T.; Yiang, G.; Lin, C.; Chan, D.; et al. Isochaihulactone Protects PC12 Cell against H2O2 Induced Oxidative Stress and Exerts the Potent Anti-Aging Effects in D-Galactose Aging Mouse Model. Acta Pharmacol. Sin. 2010, 31, 1532–1540. [Google Scholar] [CrossRef]
- Jang, H.-J.; Yang, K.E.; Oh, W.K.; Lee, S.-I.; Hwang, I.-H.; Ban, K.-T.; Yoo, H.-S.; Choi, J.-S.; Yeo, E.-J.; Jang, I.-S. Nectandrin B-Mediated Activation of the AMPK Pathway Prevents Cellular Senescence in Human Diploid Fibroblasts by Reducing Intracellular ROS Levels. Aging 2019, 11, 3731–3749. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and Roles of Stilbenes in Plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Flamini, R.; de Rosso, M. High-Resolution Mass Spectrometry and Biological Properties of Grapevine and Wine Stilbenoids. Stud. Nat. Prod. Chem. 2019, 61, 175–210. [Google Scholar]
- Griñán-Ferré, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillón, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C.; et al. The Pleiotropic Neuroprotective Effects of Resveratrol in Cognitive Decline and Alzheimer’s Disease Pathology: From Antioxidant to Epigenetic Therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef]
- Li, B.; Hou, D.; Guo, H.; Zhou, H.; Zhang, S.; Xu, X.; Liu, Q.; Zhang, X.; Zou, Y.; Gong, Y.; et al. Resveratrol Sequentially Induces Replication and Oxidative Stresses to Drive P53-CXCR2 Mediated Cellular Senescence in Cancer Cells. Sci. Rep. 2017, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential of Resveratrol: The in Vivo Evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Lu, X.; Ma, L.; Ruan, L.; Kong, Y.; Mou, H.; Zhang, Z.; Wang, Z.; Wang, J.M.; Le, Y. Resveratrol Differentially Modulates Inflammatory Responses of Microglia and Astrocytes. J. Neuroinflamm. 2010, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Carrizzo, A.; Forte, M.; Damato, A.; Trimarco, V.; Salzano, F.; Bartolo, M.; Maciag, A.; Puca, A.A.; Vecchione, C. Antioxidant Effects of Resveratrol in Cardiovascular, Cerebral and Metabolic Diseases. Food Chem. Toxicol. 2013, 61, 215–226. [Google Scholar] [CrossRef]
- Simão, F.; Matté, A.; Pagnussat, A.S.; Netto, C.A.; Salbego, C.G. Resveratrol Prevents CA1 Neurons against Ischemic Injury by Parallel Modulation of Both GSK-3β and CREB through PI3-K/Akt Pathways. Eur. J. Neurosci. 2012, 36, 2899–2905. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Jang, J.-H.; Li, M.-H.; Surh, Y.-J. Resveratrol Upregulates Heme Oxygenase-1 Expression via Activation of NF-E2-Related Factor 2 in PC12 Cells. Biochem. Biophys. Res. Commun. 2005, 331, 993–1000. [Google Scholar] [CrossRef]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health Benefits of Resveratrol: Evidence from Clinical Studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Dou, Y.; Li, A. Resveratrol Boosts Cognitive Function by Targeting SIRT1. Neurochem. Res. 2018, 43, 1705–1713. [Google Scholar] [CrossRef]
- Ohtsu, A.; Shibutani, Y.; Seno, K.; Iwata, H.; Kuwayama, T.; Shirasuna, K. Advanced Glycation End Products and Lipopolysaccharides Stimulate Interleukin-6 Secretion via the RAGE/TLR4-NF-κB-ROS Pathways and Resveratrol Attenuates These Inflammatory Responses in Mouse Macrophages. Exp. Ther. Med. 2017, 14, 4363–4370. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Li, Z.; Xu, K.; Wang, Y.; Wang, Z. Resveratrol Protects against High-Fat Diet Induced Renal Pathological Damage and Cell Senescence by Activating SIRT1. Biol. Pharm. Bull. 2016, 39, 1448–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-R.; Li, S.; Lin, C.-C. Effect of Resveratrol and Pterostilbene on Aging and Longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Chen, L.; Kowal, J.M.; Okla, M.; Manikandan, M.; AlShehri, M.; AlMana, Y.; AlObaidan, R.; AlOtaibi, N.; Hamam, R.; et al. Resveratrol Inhibits Adipocyte Differentiation and Cellular Senescence of Human Bone Marrow Stromal Stem Cells. Bone 2020, 133, 115252. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.-H.; Kim, S.Y. Resveratrol-Enriched Rice Attenuates UVB-ROS-Induced Skin Aging via Downregulation of Inflammatory Cascades. Oxidative Med. Cell. Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef] [PubMed]
- Kilic Eren, M.; Kilincli, A.; Eren, Ö. Resveratrol Induced Premature Senescence Is Associated with DNA Damage Mediated SIRT1 and SIRT2 Down-Regulation. PLoS ONE 2015, 10, e0124837. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.A.M.; Coelho, B.P.; Behr, G.; Pettenuzzo, L.F.; Souza, I.C.C.; Moreira, J.C.F.; Borojevic, R.; Gottfried, C.; Guma, F.C.R. Resveratrol Induces Pro-Oxidant Effects and Time-Dependent Resistance to Cytotoxicity in Activated Hepatic Stellate Cells. Cell Biochem. Biophys. 2014, 68, 247–257. [Google Scholar] [CrossRef]
- Heiss, E.H.; Schilder, Y.D.C.; Dirsch, V.M. Chronic Treatment with Resveratrol Induces Redox Stress- and Ataxia Telangiectasia-Mutated (ATM)-Dependent Senescence in P53-Positive Cancer Cells. J. Biol. Chem. 2007, 282, 26759–26766. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekara, A. Phenolic Acids. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 535–545. [Google Scholar]
- Guo, L.; Cao, J.; Wei, T.; Li, J.; Feng, Y.; Wang, L.; Sun, Y.; Chai, Y. Gallic Acid Attenuates Thymic Involution in the D-Galactose Induced Accelerated Aging Mice. Immunobiology 2020, 225, 151870. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-C.; Chen, S.-C.; Huang, H.-P.; Wang, C.-J. Gallic Acid Inhibits Bladder Cancer Cell Proliferation and Migration via Regulating Fatty Acid Synthase (FAS). J. Food Drug Anal. 2018, 26, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Punithavathi, V.R.; Stanely Mainzen Prince, P.; Kumar, M.R.; Selvakumari, C.J. Protective Effects of Gallic Acid on Hepatic Lipid Peroxide Metabolism, Glycoprotein Components and Lipids in Streptozotocin-Induced Type II Diabetic Wistar Rats. J. Biochem. Mol. Toxicol. 2011, 25, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Moghtaderi, H.; Sepehri, H.; Delphi, L.; Attari, F. Gallic Acid and Curcumin Induce Cytotoxicity and Apoptosis in Human Breast Cancer Cell MDA-MB-231. BioImpacts 2018, 8, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A Role of Gallic Acid in Oxidative Damage Diseases: A Comprehensive Review. Nat. Prod. Commun. 2019, 14, 1934578X1987417. [Google Scholar] [CrossRef] [Green Version]
- Dludla, P.; Nkambule, B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Orlando, P.; Tiano, L.; Louw, J.; Mazibuko-Mbeje, S. Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients 2018, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Varma, S.R.; Sivaprakasam, T.O.; Mishra, A.; Kumar, L.M.S.; Prakash, N.S.; Prabhu, S.; Ramakrishnan, S. Protective Effects of Triphala on Dermal Fibroblasts and Human Keratinocytes. PLoS ONE 2016, 11, e0145921. [Google Scholar] [CrossRef]
- Baek, B.; Lee, S.H.; Kim, K.; Lim, H.-W.; Lim, C.-J. Ellagic Acid Plays a Protective Role against UV-B-Induced Oxidative Stress by up-Regulating Antioxidant Components in Human Dermal Fibroblasts. Korean J. Physiol. Pharmacol. 2016, 20, 269. [Google Scholar] [CrossRef] [Green Version]
- Vattem, D.A.; Shetty, K. Biological Functionality of Ellagic Acid: A Review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]
- Hwang, J.M.; Cho, J.S.; Kim, T.H.; Lee, Y.I. Ellagic Acid Protects Hepatocytes from Damage by Inhibiting Mitochondrial Production of Reactive Oxygen Species. Biomed. Pharmacother. 2010, 64, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Uzar, E.; Alp, H.; Cevik, M.U.; Fırat, U.; Evliyaoglu, O.; Tufek, A.; Altun, Y. Ellagic Acid Attenuates Oxidative Stress on Brain and Sciatic Nerve and Improves Histopathology of Brain in Streptozotocin-Induced Diabetic Rats. Neurol. Sci. 2012, 33, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Baeeri, M.; Momtaz, S.; Navaei-Nigjeh, M.; Niaz, K.; Rahimifard, M.; Ghasemi-Niri, S.F.; Sanadgol, N.; Hodjat, M.; Sharifzadeh, M.; Abdollahi, M. Molecular Evidence on the Protective Effect of Ellagic Acid on Phosalone-Induced Senescence in Rat Embryonic Fibroblast Cells. Food Chem. Toxicol. 2017, 100, 8–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthaiyah, B.; Essa, M.M.; Lee, M.; Chauhan, V.; Kaur, K.; Chauhan, A. Dietary Supplementation of Walnuts Improves Memory Deficits and Learning Skills in Transgenic Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 42, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Ou, L.; Kong, L.-Y.; Zhang, X.-M.; Niwa, M. Oxidation of Ferulic Acid by Momordica Charantia Peroxidase and Related Anti-Inflammation Activity Changes. Biol. Pharm. Bull. 2003, 26, 1511–1516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Lin, D.; Jiang, R.; Li, H.; Wan, J.; Li, H. Ferulic Acid Exerts Antitumor Activity and Inhibits Metastasis in Breast Cancer Cells by Regulating Epithelial to Mesenchymal Transition. Oncol. Rep. 2016, 36, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Q.; Ma, Z.-C.; Huang, H.; Wang, Y.-G.; Tan, H.-L.; Xiao, C.-R.; Liang, Q.-D.; Zhang, H.-T.; Gao, Y. Antithrombotic Activities of Ferulic Acid via Intracellular Cyclic Nucleotide Signaling. Eur. J. Pharmacol. 2016, 777, 1–8. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial Activity and Mode of Action of Ferulic and Gallic Acids Against Pathogenic Bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Wagle, S.; Sim, H.-J.; Bhattarai, G.; Choi, K.-C.; Kook, S.-H.; Lee, J.-C.; Jeon, Y.-M. Supplemental Ferulic Acid Inhibits Total Body Irradiation-Mediated Bone Marrow Damage, Bone Mass Loss, Stem Cell Senescence, and Hematopoietic Defect in Mice by Enhancing Antioxidant Defense Systems. Antioxidants 2021, 10, 1209. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Balasubashini, M.S.; Rukkumani, R.; Viswanathan, P.; Menon, V.P. Ferulic Acid Alleviates Lipid Peroxidation in Diabetic Rats. Phytother. Res. 2004, 18, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Zhang, R.; Li, Y.; Li, Y.; Yang, Z.; Yang, H. Ferulic Acid Exerts Neuroprotective Effects against Cerebral Ischemia/Reperfusion-Induced Injury via Antioxidant and Anti-Apoptotic Mechanisms In Vitro and In Vivo. Int. J. Mol. Med. 2017, 40, 1444–1456. [Google Scholar] [CrossRef] [Green Version]
- Meng, G.; Meng, X.; Ma, X.; Zhang, G.; Hu, X.; Jin, A.; Zhao, Y.; Liu, X. Application of Ferulic Acid for Alzheimer’s Disease: Combination of Text Mining and Experimental Validation. Front. Neuroinform. 2018, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic Acid Exerts Its Antidiabetic Effect by Modulating Insulin-Signalling Molecules in the Liver of High-Fat Diet and Fructose-Induced Type-2 Diabetic Adult Male Rat. Appl. Physiol. Nutr. Metab. 2015, 40, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Stompor-Gorący, M.; Machaczka, M. Recent Advances in Biological Activity, New Formulations and Prodrugs of Ferulic Acid. Int. J. Mol. Sci. 2021, 22, 12889. [Google Scholar] [CrossRef]
- Kim, S.; Kim, J.; Lee, Y.I.; Jang, S.; Song, S.Y.; Lee, W.J.; Lee, J.H. Particulate Matter-induced Atmospheric Skin Aging Is Aggravated by UVA and Inhibited by a Topical L-Ascorbic Acid Compound. Photodermatol. Photoimmunol. Photomed. 2022, 38, 123–131. [Google Scholar] [CrossRef]
- Igarashi, K.; Kurata, D. Effect of High-Oleic Peanut Intake on Aging and Its Hippocampal Markers in Senescence-Accelerated Mice (SAMP8). Nutrients 2020, 12, 3461. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. P-Coumaric Acid and Its Conjugates: Dietary Sources, Pharmacokinetic Properties and Biological Activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. P-Coumaric Acid Kills Bacteria through Dual Damage Mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Amalan, V.; Vijayakumar, N.; Indumathi, D.; Ramakrishnan, A. Antidiabetic and Antihyperlipidemic Activity of P-Coumaric Acid in Diabetic Rats, Role of Pancreatic GLUT 2: In Vivo Approach. Biomed. Pharmacother. 2016, 84, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Nasr Bouzaiene, N.; Kilani Jaziri, S.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The Effects of Caffeic, Coumaric and Ferulic Acids on Proliferation, Superoxide Production, Adhesion and Migration of Human Tumor Cells in Vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.-H.; Cheon, S.-R.; Kim, J.-H.; Choi, K.-C.; Kim, M.-K.; Lee, J.-C. Dietary Hydroxycinnamates Prevent Oxidative Damages to Liver, Spleen, and Bone Marrow Cells in Irradiation-Exposed Mice. Food Sci. Biotechnol. 2017, 26, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Widowati, W.; Fauziah, N.; Herdiman, H.; Afni, M.; Afifah, E.; Kusuma, H.S.W.; Nufus, H.; Arumwardana, S.; Rihibiha, D.D. Antioxidant and Anti Aging Assays of Oryza Sativa Extracts, Vanillin and Coumaric Acid. J. Nat. Remedies 2016, 16, 88. [Google Scholar] [CrossRef] [Green Version]
- Pieńkowska, N.; Bartosz, G.; Pichla, M.; Grzesik-Pietrasiewicz, M.; Gruchala, M.; Sadowska-Bartosz, I. Effect of Antioxidants on the H2O2-Induced Premature Senescence of Human Fibroblasts. Aging 2020, 12, 1910–1927. [Google Scholar] [CrossRef]
- Morita, H.; Abe, I.; Noguchi, H. Plant Type III PKS. In Comprehensive Natural Products II; Elsevier: Amsterdam, The Netherlands, 2010; pp. 171–225. [Google Scholar]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological Activities of Curcuminoids, Other Biomolecules from Turmeric and Their Derivatives—A Review. J. Tradit. Complement. Med. 2017, 7, 205–233. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, T. Curcumin as a Functional Food-Derived Factor: Degradation Products, Metabolites, Bioactivity, and Future Perspectives. Food Funct. 2018, 9, 705–714. [Google Scholar] [CrossRef]
- Tomeh, M.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [Green Version]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Curcumin, a Natural Antimicrobial Agent with Strain-Specific Activity. Pharmaceuticals 2020, 13, 153. [Google Scholar] [CrossRef]
- Nabavi, S.; Thiagarajan, R.; Rastrelli, L.; Daglia, M.; Sobarzo-Sanchez, E.; Alinezhad, H.; Nabavi, S. Curcumin: A Natural Product for Diabetes and Its Complications. Curr. Top. Med. Chem. 2015, 15, 2445–2455. [Google Scholar] [CrossRef]
- Donatus, I.A.; Sardjoko; Vermeulen, N.P.E. Cytotoxic and Cytoprotective Activities of Curcumin. Biochem. Pharmacol. 1990, 39, 1869–1875. [Google Scholar] [CrossRef]
- Zia, A.; Farkhondeh, T.; Pourbagher-Shahri, A.M.; Samarghandian, S. The Role of Curcumin in Aging and Senescence: Molecular Mechanisms. Biomed. Pharmacother. 2021, 134, 111119. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.; Alsahli, M.; Aly, S.; Khan, M.; Aldebasi, Y. Role of Curcumin in Disease Prevention and Treatment. Adv. Biomed. Res. 2018, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, Y.S.; Kim, E.; Kim, Y.; Kim, Y. Curcumin and Hesperetin Attenuate D-Galactose-Induced Brain Senescence In Vitro and In Vivo. Nutr. Res. Pract. 2020, 14, 438. [Google Scholar] [CrossRef]
- Lee, S.-J.; Chandrasekran, P.; Mazucanti, C.H.; O’Connell, J.F.; Egan, J.M.; Kim, Y. Dietary Curcumin Restores Insulin Homeostasis in Diet-Induced Obese Aged Mice. Aging 2022, 14, 225–239. [Google Scholar] [CrossRef]
- Santos-Parker, J.R.; Lubieniecki, K.L.; Rossman, M.J.; van Ark, H.J.; Bassett, C.J.; Strahler, T.R.; Chonchol, M.B.; Justice, J.N.; Seals, D.R. Curcumin Supplementation and Motor-Cognitive Function in Healthy Middle-Aged and Older Adults. Nutr. Healthy Aging 2018, 4, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Tavakol, S.; Zare, S.; Hoveizi, E.; Tavakol, B.; Rezayat, S.M. The Impact of the Particle Size of Curcumin Nanocarriers and the Ethanol on Beta_1-Integrin Overexpression in Fibroblasts: A Regenerative Pharmaceutical Approach in Skin Repair and Anti-Aging Formulations. DARU J. Pharm. Sci. 2019, 27, 159–168. [Google Scholar] [CrossRef]
- Schiborr, C.; P. Eckert, G.; Weissenberger, J.; E. Muller, W.; Schwamm, D.; Grune, T.; Rimbach, G.; Frank, J. Cardiac Oxidative Stress and Inflammation Are Similar in SAMP8 and SAMR1 Mice and Unaltered by Curcumin and Ginkgo Biloba Extract Intake. Curr. Pharm. Biotechnol. 2010, 11, 861–867. [Google Scholar] [CrossRef]
- Yoder, S.C.; Lancaster, S.M.; Hullar, M.A.J.; Lampe, J.W. Gut Microbial Metabolism of Plant Lignans. In Diet-Microbe Interactions in the Gut; Elsevier: Amsterdam, The Netherlands, 2015; pp. 103–117. [Google Scholar]
- Adolphe, J.L.; Whiting, S.J.; Juurlink, B.H.J.; Thorpe, L.U.; Alcorn, J. Health Effects with Consumption of the Flax Lignan Secoisolariciresinol Diglucoside. Br. J. Nutr. 2010, 103, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Le, T.D.; Nakahara, Y.; Ueda, M.; Okumura, K.; Hirai, J.; Sato, Y.; Takemoto, D.; Tomimori, N.; Ono, Y.; Nakai, M.; et al. Sesamin Suppresses Aging Phenotypes in Adult Muscular and Nervous Systems and Intestines in a Drosophila Senescence-Accelerated Model. Eur. Rev. Med. Pharm. Sci. 2019, 23, 1826–1839. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. An Overview of Neuroprotective and Cognitive Enhancement Properties of Lignans from Schisandra Chinensis. Biomed. Pharmacother. 2018, 97, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Velalopoulou, A.; Chatterjee, S.; Pietrofesa, R.; Koziol-White, C.; Panettieri, R.; Lin, L.; Tuttle, S.; Berman, A.; Koumenis, C.; Christofidou-Solomidou, M. Synthetic Secoisolariciresinol Diglucoside (LGM2605) Protects Human Lung in an Ex Vivo Model of Proton Radiation Damage. Int. J. Mol. Sci. 2017, 18, 2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenkin, A. Micronutrients in Health and Disease. Postgrad. Med. J. 2006, 82, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehri, A. Trace Elements in Human Nutrition (II)—An Update. Int. J. Prev. Med. 2020, 11, 2. [Google Scholar] [CrossRef]
- Rudolf, E.; Rudolf, K. Low Zinc Environment Induces Stress Signaling, Senescence and Mixed Cell Death Modalities in Colon Cancer Cells. Apoptosis 2015, 20, 1651–1665. [Google Scholar] [CrossRef]
- Rudolf, E.; Cervinka, M. Stress Responses of Human Dermal Fibroblasts Exposed to Zinc Pyrithione. Toxicol. Lett. 2011, 204, 164–173. [Google Scholar] [CrossRef]
- Malavolta, M.; Costarelli, L.; Giacconi, R.; Basso, A.; Piacenza, F.; Pierpaoli, E.; Provinciali, M.; Ogo, O.A.; Ford, D. Changes in Zn Homeostasis during Long Term Culture of Primary Endothelial Cells and Effects of Zn on Endothelial Cell Senescence. Exp. Gerontol. 2017, 99, 35–45. [Google Scholar] [CrossRef]
- Legrain, Y.; Touat-Hamici, Z.; Chavatte, L. Interplay between Selenium Levels, Selenoprotein Expression, and Replicative Senescence in WI-38 Human Fibroblasts. J. Biol. Chem. 2014, 289, 6299–6310. [Google Scholar] [CrossRef] [Green Version]
- Jobeili, L.; Rousselle, P.; Béal, D.; Blouin, E.; Roussel, A.-M.; Damour, O.; Rachidi, W. Selenium Preserves Keratinocyte Stemness and Delays Senescence by Maintaining Epidermal Adhesion. Aging 2017, 9, 2302–2315. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.T.Y.; Cao, L.; Chen, B.P.C.; Cheng, W.-H. Selenoprotein H Suppresses Cellular Senescence through Genome Maintenance and Redox Regulation. J. Biol. Chem. 2014, 289, 34378–34388. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.T.; Cao, L.; Mattson, E.; Witwer, K.W.; Cao, J.; Zeng, H.; He, X.; Combs, G.F.; Cheng, W. Opposing Impacts on Healthspan and Longevity by Limiting Dietary Selenium in Telomere Dysfunctional Mice. Aging Cell 2017, 16, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Dickens, B.F.; Weglicki, W.B.; Li, Y.-S.; Mak, I.T. Magnesium Deficiency in Vitro Enhances Free Radical-Induced Intracellular Oxidation and Cytotoxicity in Endothelial Cells. FEBS Lett. 1992, 311, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Wolf, F.I.; Trapani, V.; Simonacci, M.; Ferré, S.; Maier, J.A.M. Magnesium Deficiency and Endothelial Dysfunction: Is Oxidative Stress Involved? Magnes Res. 2008, 21, 58–64. [Google Scholar] [PubMed]
- Yang, Y.; Wu, Z.; Chen, Y.; Qiao, J.; Gao, M.; Yuan, J.; Nie, W.; Guo, Y. Magnesium Deficiency Enhances Hydrogen Peroxide Production and Oxidative Damage in Chick Embryo Hepatocyte In Vitro. BioMetals 2006, 19, 71–81. [Google Scholar] [CrossRef]
- Killilea, D.W.; Ames, B.N. Magnesium Deficiency Accelerates Cellular Senescence in Cultured Human Fibroblasts. Proc. Natl. Acad. Sci. USA 2008, 105, 5768–5773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blache, D.; Devaux, S.; Joubert, O.; Loreau, N.; Schneider, M.; Durand, P.; Prost, M.; Gaume, V.; Adrian, M.; Laurant, P.; et al. Long-Term Moderate Magnesium-Deficient Diet Shows Relationships between Blood Pressure, Inflammation and Oxidant Stress Defense in Aging Rats. Free Radic. Biol. Med. 2006, 41, 277–284. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Loutsidou, A.C.; Spiliopoulou, C.A.; Stefanidou, M.E. Zinc and Human Health: An Update. Arch. Toxicol. 2012, 86, 521–534. [Google Scholar] [CrossRef]
- Cousins, R.J.; Blanchard, R.K.; Moore, J.B.; Cui, L.; Green, C.L.; Liuzzi, J.P.; Cao, J.; Bobo, J.A. Regulation of Zinc Metabolism and Genomic Outcomes. J. Nutr. 2003, 133, 1521S–1526S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maret, W. Zinc Biochemistry: From a Single Zinc Enzyme to a Key Element of Life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Liu, X.; Pan, Z. Zinc Deficiency and Cellular Oxidative Stress: Prognostic Implications in Cardiovascular Diseases. Acta Pharmacol. Sin. 2018, 39, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.R. Critical Role of Zinc as Either an Antioxidant or a Prooxidant in Cellular Systems. Oxidative Med. Cell. Longev. 2018, 2018, 9156285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marreiro, D.; Cruz, K.; Morais, J.; Beserra, J.; Severo, J.; de Oliveira, A. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef]
- Yu, K.-N.; Yoon, T.-J.; Minai-Tehrani, A.; Kim, J.-E.; Park, S.J.; Jeong, M.S.; Ha, S.-W.; Lee, J.-K.; Kim, J.S.; Cho, M.-H. Zinc Oxide Nanoparticle Induced Autophagic Cell Death and Mitochondrial Damage via Reactive Oxygen Species Generation. Toxicol. Vitr. 2013, 27, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- O’Dell, B.L. Role of Zinc in Plasma Membrane Function. J. Nutr. 2000, 130, 1432S–1436S. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. The Oxidative Stress of Zinc Deficiency. Metallomics 2011, 3, 1124. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Kim, M.J.; Kim, K.J.; Kim, J.-R. POZ/BTB and AT-Hook-Containing Zinc Finger Protein 1 (PATZ1) Inhibits Endothelial Cell Senescence through a P53 Dependent Pathway. Cell Death Differ. 2012, 19, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Ji, Q.; Song, M.; Ren, J.; Liu, Z.; Wang, Z.; Liu, X.; Yan, K.; Hu, J.; Jing, Y.; et al. ZKSCAN3 Counteracts Cellular Senescence by Stabilizing Heterochromatin. Nucleic Acids Res. 2020, 48, 6001–6018. [Google Scholar] [CrossRef]
- Igata, T.; Tanaka, H.; Etoh, K.; Hong, S.; Tani, N.; Koga, T.; Nakao, M. Loss of the Transcription Repressor ZHX3 Induces Senescence-Associated Gene Expression and Mitochondrial-Nucleolar Activation. PLoS ONE 2022, 17, e0262488. [Google Scholar] [CrossRef]
- Liu, C.; la Rosa, S.; Hagos, E.G. Oxidative DNA Damage Causes Premature Senescence in Mouse Embryonic Fibroblasts Deficient for Krüppel-like Factor 4. Mol. Carcinog. 2015, 54, 889–899. [Google Scholar] [CrossRef]
- Gao, B.; Li, K.; Wei, Y.-Y.; Zhang, J.; Li, J.; Zhang, L.; Gao, J.-P.; Li, Y.-Y.; Huang, L.-G.; Lin, P.; et al. Zinc Finger Protein 637 Protects Cells against Oxidative Stress-Induced Premature Senescence by MTERT-Mediated Telomerase Activity and Telomere Maintenance. Cell Death Dis. 2014, 5, e1334. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lu, Y.; Yang, P.; Chen, Q.; Wang, Y.; Ding, Q.; Xu, T.; Li, X.; Li, C.; Huang, C.; et al. MicroRNA-145 Induces the Senescence of Activated Hepatic Stellate Cells through the Activation of P53 Pathway by ZEB2. J. Cell. Physiol. 2019, 234, 7587–7599. [Google Scholar] [CrossRef]
- Yi, Y.; Xie, H.; Xiao, X.; Wang, B.; Du, R.; Liu, Y.; Li, Z.; Wang, J.; Sun, L.; Deng, Z.; et al. Ultraviolet A Irradiation Induces Senescence in Human Dermal Fibroblasts by Down-Regulating DNMT1 via ZEB1. Aging 2018, 10, 212–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, A.; Gagné, A.; Laflamme, P.; Marcoux, M.; Orain, M.; Plante, S.; Joubert, D.; Joubert, P.; Laplante, M. ZNF768 Expression Associates with High Proliferative Clinicopathological Features in Lung Adenocarcinoma. Cancers 2021, 13, 4136. [Google Scholar] [CrossRef]
- Salazar, G. NADPH Oxidases and Mitochondria in Vascular Senescence. Int. J. Mol. Sci. 2018, 19, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, H.J.; Hondal, R.J. Why Nature Chose Selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Opstad, T.B.; Alexander, J.; Larsson, A.; Aaseth, J. Impact of Selenium on Biomarkers and Clinical Aspects Related to Ageing. A Review. Biomolecules 2021, 11, 1478. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and Selenoproteins: It’s Role in Regulation of Inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Selenius, M.; Rundlöf, A.-K.; Olm, E.; Fernandes, A.P.; Björnstedt, M. Selenium and the Selenoprotein Thioredoxin Reductase in the Prevention, Treatment and Diagnostics of Cancer. Antioxid. Redox Signal. 2010, 12, 867–880. [Google Scholar] [CrossRef]
- Ebert, R.; Ulmer, M.; Zeck, S.; Meissner-Weigl, J.; Schneider, D.; Stopper, H.; Schupp, N.; Kassem, M.; Jakob, F. Selenium Supplementation Restores the Antioxidative Capacity and Prevents Cell Damage in Bone Marrow Stromal Cells In Vitro. Stem cells 2006, 24, 1226–1235. [Google Scholar] [CrossRef]
- Hartwig, A. Role of Magnesium in Genomic Stability. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2001, 475, 113–121. [Google Scholar] [CrossRef]
- Maguire, D.; Neytchev, O.; Talwar, D.; McMillan, D.; Shiels, P. Telomere Homeostasis: Interplay with Magnesium. Int. J. Mol. Sci. 2018, 19, 157. [Google Scholar] [CrossRef] [Green Version]
- Mazur, A.; Maier, J.A.M.; Rock, E.; Gueux, E.; Nowacki, W.; Rayssiguier, Y. Magnesium and the Inflammatory Response: Potential Physiopathological Implications. Arch. Biochem. Biophys. 2007, 458, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, G.G.; van Noorden, S.; Balato, A.; Cimini, V. Copper/Zinc Superoxide Dismutase in Human Skin: Current Knowledge. Front. Med. 2020, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Kinnula, V.L.; Crapo, J.D. Superoxide Dismutases in the Lung and Human Lung Diseases. Am. J. Respir. Crit. Care Med. 2003, 167, 1600–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajac, D. Mineral Micronutrients in Asthma. Nutrients 2021, 13, 4001. [Google Scholar] [CrossRef]
- Majidinia, M.; Reiter, R.J.; Shakouri, S.K.; Yousefi, B. The Role of Melatonin, a Multitasking Molecule, in Retarding the Processes of Ageing. Ageing Res. Rev. 2018, 47, 198–213. [Google Scholar] [CrossRef]
- Lahiri, D.K.; Ghosh, C. Interactions between Melatonin, Reactive Oxygen Species, and Nitric Oxide. Ann. N. Y. Acad. Sci. 1999, 893, 325–330. [Google Scholar] [CrossRef]
- Hardeland, R. Aging, Melatonin, and the Pro- and Anti-Inflammatory Networks. Int. J. Mol. Sci. 2019, 20, 1223. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Liu, X.; Chen, D.; Lu, Y.; Li, J.; Du, F.; Zhang, C.; Lu, L. Melatonin Prevents Endothelial Dysfunction in SLE by Activating the Nuclear Receptor Retinoic Acid-Related Orphan Receptor-α. Int. Immunopharmacol. 2020, 83, 106365. [Google Scholar] [CrossRef]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A Pleiotropic Molecule Regulating Inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Yoon, Y.M.; Song, K.; Noh, H.; Lee, S.H. Melatonin Suppresses Senescence-derived Mitochondrial Dysfunction in Mesenchymal Stem Cells via the HSPA1L–Mitophagy Pathway. Aging Cell 2020, 19, e13111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhao, C.; Liu, P.; Huang, H.; Zhang, S.; Wang, X. Anti-Apoptosis and Autophagy Effects of Melatonin Protect Rat Chondrocytes against Oxidative Stress via Regulation of AMPK/Foxo3 Pathways. CARTILAGE 2021, 13, 1041S–1053S. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.K. Control of Mesenchymal Stromal Cell Senescence by Tryptophan Metabolites. Int. J. Mol. Sci. 2021, 22, 697. [Google Scholar] [CrossRef]
- Cai, B.; Ma, W.; Bi, C.; Yang, F.; Zhang, L.; Han, Z.; Huang, Q.; Ding, F.; Li, Y.; Yan, G.; et al. Long Noncoding RNA H19 Mediates Melatonin Inhibition of Premature Senescence of C-Kit+ Cardiac Progenitor Cells by Promoting MiR-675. J. Pineal Res. 2016, 61, 82–95. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, X.; Liu, T.; Gong, Y.; Chen, S.; Pan, G.; Cui, W.; Luo, Z.-P.; Pei, M.; Yang, H.; et al. Melatonin Reverses H2O2-Induced Premature Senescence in Mesenchymal Stem Cells via the SIRT1-Dependent Pathway. J. Pineal Res. 2015, 59, 190–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, N.; Shi, Y.; Zhang, C.; Zheng, Y.; Wang, Y.; Zhao, B.; Zeng, Y.; Liu, X.; Liu, J. Antioxidants Inhibit Cell Senescence and Preserve Stemness of Adipose Tissue-Derived Stem Cells by Reducing ROS Generation during Long-Term In Vitro Expansion. Stem Cell Res. Ther. 2019, 10, 306. [Google Scholar] [CrossRef] [Green Version]
- Shuai, Y.; Liao, L.; Su, X.; Yu, Y.; Shao, B.; Jing, H.; Zhang, X.; Deng, Z.; Jin, Y. Melatonin Treatment Improves Mesenchymal Stem Cells Therapy by Preserving Stemness during Long-Term In Vitro Expansion. Theranostics 2016, 6, 1899–1917. [Google Scholar] [CrossRef]
- Puig, Á.; Rancan, L.; Paredes, S.D.; Carrasco, A.; Escames, G.; Vara, E.; Tresguerres, J.A.F. Melatonin Decreases the Expression of Inflammation and Apoptosis Markers in the Lung of a Senescence-Accelerated Mice Model. Exp. Gerontol. 2016, 75, 1–7. [Google Scholar] [CrossRef]
- Parisotto, E.B.; Vidal, V.; García-Cerro, S.; Lantigua, S.; Wilhelm Filho, D.; Sanchez-Barceló, E.J.; Martínez-Cué, C.; Rueda, N. Chronic Melatonin Administration Reduced Oxidative Damage and Cellular Senescence in the Hippocampus of a Mouse Model of Down Syndrome. Neurochem. Res. 2016, 41, 2904–2913. [Google Scholar] [CrossRef] [Green Version]
- Okatani, Y.; Wakatsuki, A.; Reiter, R.J.; Miyahara, Y. Melatonin Reduces Oxidative Damage of Neural Lipids and Proteins in Senescence-Accelerated Mouse. Neurobiol. Aging 2002, 23, 639–644. [Google Scholar] [CrossRef]
- Sumsuzzman, D.M.; Khan, Z.A.; Choi, J.; Hong, Y. Differential Role of Melatonin in Healthy Brain Aging: A Systematic Review and Meta-Analysis of the SAMP8 Model. Aging 2021, 13, 9373–9397. [Google Scholar] [CrossRef] [PubMed]
- Bin-Jaliah, I.; Sakr, H. Melatonin Ameliorates Brain Oxidative Stress and Upregulates Senescence Marker Protein-30 and Osteopontin in a Rat Model of Vascular Dementia. Physiol. Int. 2018, 105, 38–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.P.; Demmitt, B.A.; Nath, R.D.; Brunet, A. The Genetics of Aging: A Vertebrate Perspective. Cell 2019, 177, 200–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redman, L.M.; Smith, S.R.; Burton, J.H.; Martin, C.K.; Il’yasova, D.; Ravussin, E. Metabolic Slowing and Reduced Oxidative Damage with Sustained Caloric Restriction Support the Rate of Living and Oxidative Damage Theories of Aging. Cell Metab. 2018, 27, 805–815.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzotti, D.R.; Guindalini, C.; dos Moraes, W.A.S.; Andersen, M.L.; Cendoroglo, M.S.; Ramos, L.R.; Tufik, S. Human Longevity Is Associated with Regular Sleep Patterns, Maintenance of Slow Wave Sleep, and Favorable Lipid Profile. Front. Aging Neurosci. 2014, 6, 134. [Google Scholar] [CrossRef] [Green Version]
- Epel, E.S.; Lithgow, G.J. Stress Biology and Aging Mechanisms: Toward Understanding the Deep Connection Between Adaptation to Stress and Longevity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, S10–S16. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, C.E.; Dickens, A.P.; Jones, K.; Thompson-Coon, J.; Taylor, R.S.; Rogers, M.; Bambra, C.L.; Lang, I.; Richards, S.H. Is Volunteering a Public Health Intervention? A Systematic Review and Meta-Analysis of the Health and Survival of Volunteers. BMC Public Health 2013, 13, 773. [Google Scholar] [CrossRef] [Green Version]
- Young, R.D. Validated Living Worldwide Supercentenarians, Living and Recently Deceased: February 2018. Rejuvenation Res. 2018, 21, 67–69. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Lyublinskaya, O.; Kornienko, J.; Ivanova, J.; Pugovkina, N.; Alekseenko, L.; Lyublinskaya, E.; Tyuryaeva, I.; Smirnova, I.; Grinchuk, T.; Shorokhova, M.; et al. Induction of Premature Cell Senescence Stimulated by High Doses of Antioxidants Is Mediated by Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2021, 22, 11851. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, J.S.; Smirnova, I.S.; Pugovkina, N.A.; Ivanova, J.S.; Shilina, M.A.; Grinchuk, T.M.; Shatrova, A.N.; Aksenov, N.D.; Zenin, V.V.; Nikolsky, N.N.; et al. High Doses of Synthetic Antioxidants Induce Premature Senescence in Cultivated Mesenchymal Stem Cells. Sci. Rep. 2019, 9, 1296. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.P.; Harries, L.W. Senotherapeutic Drugs: A New Avenue for Skincare? Plast. Reconstr. Surg. 2021, 148, 21S–26S. [Google Scholar] [CrossRef]
- Wissler Gerdes, E.O.; Zhu, Y.; Tchkonia, T.; Kirkland, J.L. Discovery, Development, and Future Application of Senolytics: Theories and Predictions. FEBS J. 2020, 287, 2418–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkland, J.L.; Tchkonia, T. Senolytic Drugs: From Discovery to Translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef] [PubMed]



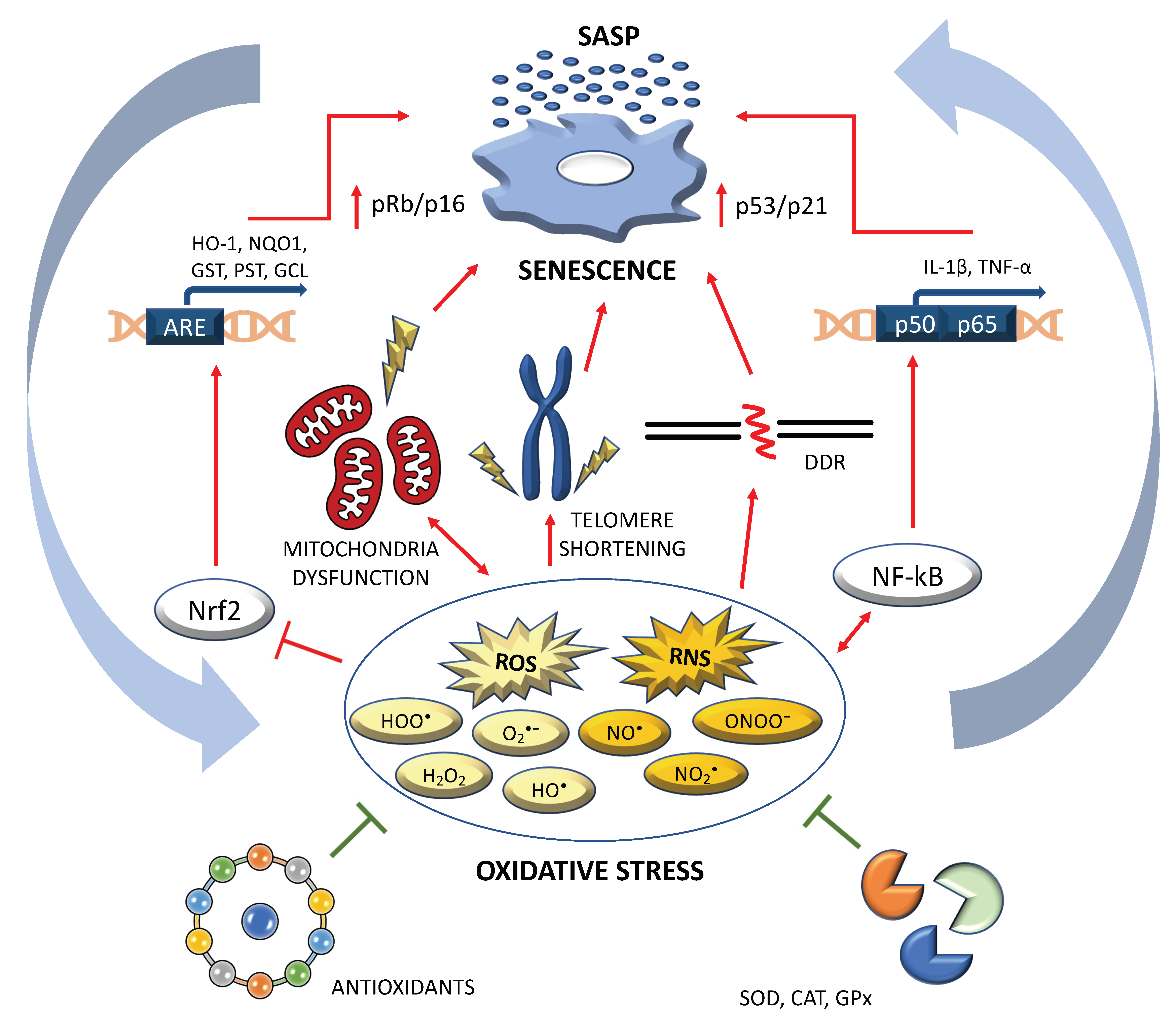{kind=link}
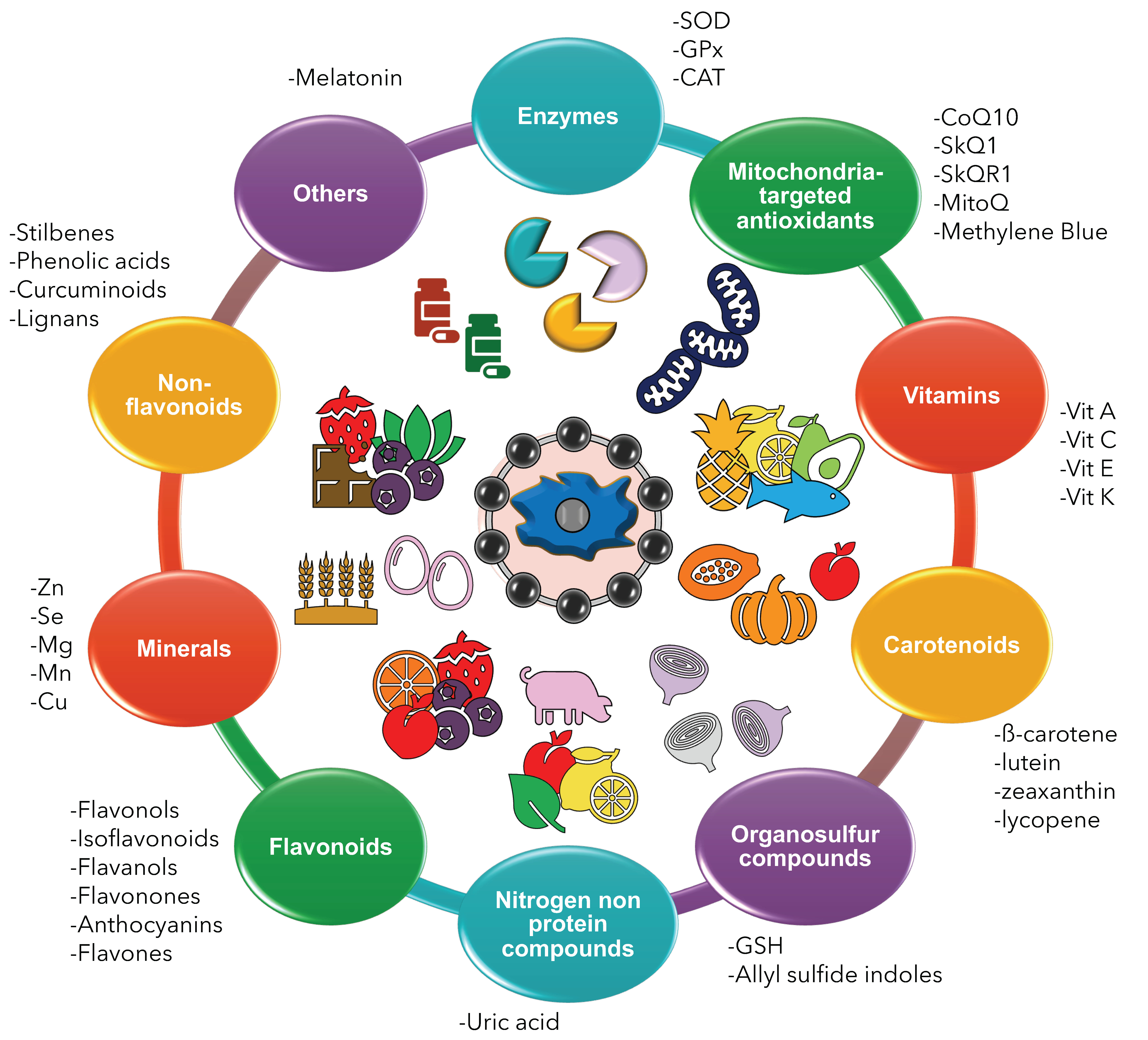{kind=link}
| Ref. | Study Design | Treatment | Form | Results | Conclusion |
|---|---|---|---|---|---|
| Chen et al., 2019 [59] | HAEC chronically treated with NRTI | 5 μM CoQ10 continuously applied across passages | CoQ10 | ↓ NRTI-induced senescence ↓ ROS ↑ Mitochondrial respiration rate | CoQ10 reduces cardiovascular side effects of NRTI treatment |
| Tarry-Adkins et al., 2013 [58] | Mouse model of low birth-weight and catch-up growth | Post-weaning dietary supplement | CoQ10 | ↓ Nitrosative and OS ↓ DNA damage ↓ Cellular senescence ↓Telomere shortening ↓ Apoptosis | CoQ10 prevents cardiac aging and cardiovascular risk |
| Ma et al., 2014 [60] | PS-1-mutated AD fibroblasts | Medium with 50 μg/mL WS-CoQ10 | WS-CoQ10 | ↓ ROS ↑ Cell doubling ↓ SIPS ↑ PCNA expression ↓ MnSOD, p21, p16Ink4A, and Rb ↑ Autophagy | WS-CoQ10 inhibits SIPS and improves autophagy |
| Xue et al., 2017 [61] | Mouse PSCs | Cell treatment with 1/10/100 μM CoQ10 for 24/48/72 h | CoQ10 | ↓ Apoptosis ↓ SA-β-Gal ↓ ROS and malondialdehyde after 72 h treatment | CoQ10 may act as a target in PSC-related pathologies |
| Wu et al., 2020 [62] | ORX mice | CoQ10 50 mg/kg/day | CoQ10 | ↓ OS ↓ Cell senescence ↑ Osteoblastic bone formation | CoQ10 is anti-osteoporosis and -senescence |
| Mine et al., 2021 [63] | H2O2-induced SIPS in human skin fibroblasts | 1 μM and 10 μM CoQ10 | CoQ10 | ↑ Cell viability ↓ OS ↓ SA-β-Gal ↓ SASP | CoQ10 can contribute to increase lifespan |
| Zhang et al., 2015 [64] | D-galactose -induced aging in MSC | 1/10/100 mmol/L CoQ10 for 48 h | CoQ10 | ↓ p-AKT and p-mTOR ↓ MSC senescence | CoQ10 inhibits MSC senescence and aging |
| Velichokovska et al., 2019 [65] | NPCs exposed to ART | NP-based delivery of CoQ10 to mitochondria | CoQ10 | ↓ ROS ↑ SIRT3 ↑ Cell proliferation ↓ SA-β-Gal ↑ Telomere length | ART-induced senescence can be reversed by NP-CoQ10 |
| Marcheggiani et al., 2021 [66] | CoQ10-deprived HDF | 5, 10 or 15 μg/mL of either CoQ10 or CoQ10H2 for 24 h | CoQ10 or CoQ10H2 | ↓ SA-β-Gal ↓ OS ↓ p21 ↑ Elastin, collagen type I | CoQ10 or CoQ10H2 prevent skin aging and support skin vitality |
| Huo et al., 2018 [67] | HUVEC treated with H2O2 | 24 h in medium with 10 μM CoQ10H2 | CoQ10H2 | ↓ SA-β-Gal positive cells ↓ SASP ↓ ROS ↓ Apoptosis ↑ NO and eNOS ↑ Mitochondrial function | CoQ10H2 delays vascular aging |
| Yan et al., 2006 [68] | SAMP1 mice | 250 mg/kg/day lifelong supplement | CoQ10H2 | ↓ Senescence grading scores ↑ Female body weight = Lifespan = Urinary 8-OHdG and acrolein-lysine adduct | CoQ10H2 decreases cellular senescence in middle-aged SAMP1 mice |
| Olivieri et al., 2013 [69] | Senescent HUVECs in presence or absence of LPS | 10 µM CoQ10H2 for 24 h or 60 days | CoQ10H2 | ↓ LPS-induced NF-kB activation ↓ SASP | CoQ10H2 may prevent aged-induced endothelial dysfunction |
| Maruoka et al., 2014 [70] | SAMP1 mice | 300 mg/kg (Group A) or 30 mg/kg CoQ10H2 (Group B) | CoQ10H2 | ↓ Senescence score at 10 months in Group A compared to B ↓ OS ↑ Antioxidant potential | CoQ10H2 reduces senescence and OS in a dose-dependent manner |
| Schmelzer et al., 2010 [71] | Middle aged SAMP1 mice | 500 mg/kg/day of CoQ10H2 for 6 or 14 months | CoQ10H2 | ↓ Senescence grading score ↑ PPAR-alpha | CoQ10H2 decelerates degeneration in SAMP1 mice |
| Cirilli et al., 2020 [72] | HUVEC treated with CSE for 24 h | 10 µM CoQ10H2 | CoQ10H2 and menaquinone 7 | ↓ OS ↓ Inflammation ↓ Apoptosis ↓ SA-β-Gal | CoQ10H2 and menaquinone-7 counteract CSE-induced damage |
| Tian et al., 2014 [73] | SAMP1 mice | Dietary CoQ10H2 (0.3% w/w) | CoQ10H2 | ↑ PGC-1α, SOD2, IDH2, SIRT1, SIRT3 ↑ Mitochondrial complex I activity ↓ OS ↑ GSH/GSSG ratio | CoQ10H2 protects against aging progression |
| Compound | Sample | Treatment | Results | Ref. |
|---|---|---|---|---|
| SkQ1 | Podospora anserina, Ceriodaphnia affinis, Drosophila melanogaster, and mouse | Nano- and subnanomolar concentrations of SkQ1 | -Prolonged lifespan -Reduced senescence | [74,75] |
| SkQ1 | Wistar and senescence-accelerated rats | 250 nmol per kg/day SkQ1 (starting from 19 months of age) | -Reduced and reversed age-related decline | [76] |
| SkQ1 | BALB/c and C57BL/6 mice | Lifelong administration of SkQ1 | -Decreased cardiomyopathy, fibrosis and cardiac hypertrophy | [77] |
| SkQ1 | Senescence-accelerated rats | 250 nmol/kg body weight, daily (from 1.5 to 23 months of age) | -Reduced Alzheimer’s disease pathology | [78] |
| MB | Human IMR90 fibroblasts | 10, 100 or 1000 nM of MB for 4 days | -Delayed senescence -Improved mitochondrial function | [79] |
| MB | Human skin fibroblasts derived from progeria patients | 100 nM MB | -Effective ROS scavenging -Improved skin fibroblast proliferation -Delayed senescence | [80] |
| MB | Human bone marrow-derived MSCs | 200 nM MB | -Improved expansion in vigorous MSCs -Improved differentiation in vigorous MSCs | [81] |
| MB | Primary rat RGCs | 1 μM and 10 μM MB | -Stimulated mitochondrial function -Enhanced neuroprotection | [82] |
| Type of Flavonoid | Effect | Reference |
|---|---|---|
| 4,4′-dimethoxychalcone |
|
|
| Naringenin |
|
|
| Nobiletin: Rutaceae family |
|
|
| Quercetin |
| |
| Fisetin |
| |
| Apigenin |
|
|
| Theaflavin |
| |
| Myrecitin |
| |
| Rutin |
| |
| Luteonil |
|
|
| Kaempferol |
|
|
| Hesperidin |
| |
| Dyhydromericetin |
| |
| Epicatechin |
| |
| Genistein |
|
| Non-Flavonoid | Model | Effects | Reference |
|---|---|---|---|
| Resveratrol | HUVEC cells |
| [320] |
| Senescence-accelerated mice |
| [321] | |
| Old male mice |
| [322] | |
| Gallic acid | Rat embryonic fibroblast cells |
| [323] |
| UVB-irradiated human fibroblast cells |
| [324] | |
| UVB-irradiated hairless mice |
| ||
| Ellagic acid | D-galactose-treated rats |
| [325] |
| Ferulic acid | UVA-irradiated nHDF |
| [326] |
| p-coumaric acid | Rat chondrocytes |
| [327] |
| Mice fed with high-fat diet (HFD) |
| [328] | |
| Curcumin | Senescence-accelerated mice |
| [329] |
| Mice fed with HFD |
| [330] | |
| Lignans | nPC12 cells |
| [331] |
| D-galactose aging mice |
| ||
| Old HDFs |
| [332] |
| Mineral | Sample | Treatment/Condition | Result | Ref. |
|---|---|---|---|---|
| Zinc | Colon cancer lines SW480 and SW620 | ↓ Zinc | ↑ Oxidative stress, cellular proliferation, stress signalling morphological changes, cell death | [409] |
| Zinc | Dermal fibroblast | ↑ Zinc | ↑ Oxidative stress and DNA damage | [410] |
| Zinc | HCAECs | ↑ Zinc | ↑ Senescence | [411] |
| Selenium | Bone marrow stromal cells | ↑ Selenium | ↓ Senescence | [412] |
| Selenium | Keratinocytes | ↑ Selenium | ↓ Senescence | [413] |
| Selenium | Human fibroblasts | ↑ Selenium | ↓ Senescence | [414] |
| Selenium | Mice | ↓ Selenium | ↑ Senescence | [415] |
| Magnesium | Endothelial cells | ↓ Magnesium | ↑ Oxidative stress and cell death | [416] |
| Magnesium | Endothelial cells | ↓ Magnesium | ↑ Pro-inflammatory molecules | [417] |
| Magnesium | Embryo-hepatocytes | ↓ Magnesium | ↑ Oxidative stress | [418] |
| Magnesium | Human fibroblasts | ↓ Magnesium | ↑ Telomere shortening | [419] |
| Magnesium | Rats | ↓ Magnesium | ↑ Age-related diseases | [420] |
| Mineral | Sample | Zinc-Finger Proteins | Ref. |
|---|---|---|---|
| Zinc | Endothelial cells | PATZ1 is downregulated in senescence | [430] |
| Zinc | Mesenchymal stem cells | ZKSCAN3 upregulation contrast senescence | [431] |
| Zinc | Human diploid fibroblast | ZHX3 is downregulated in senescence | [432] |
| Zinc | Mouse embryonic fibroblasts | KLF4 reduces cellular senescence and DNA damage | [433] |
| Zinc | NIH3T3 and C2C12 cells | ZFP637 protects from oxidative stress | [434] |
| Zinc | Hepatic stellate cells | ZEB2 protects from oxidative stress and senescence | [435] |
| Zinc | Dermal fibroblasts | ZEB1 protects from oxidative stress and senescence | [436] |
| Zinc | Cell lines (A549, NCI-H441 and NCI-H460, 293T) | ZNF768 depletion induces senescence | [437] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varesi, A.; Chirumbolo, S.; Campagnoli, L.I.M.; Pierella, E.; Piccini, G.B.; Carrara, A.; Ricevuti, G.; Scassellati, C.; Bonvicini, C.; Pascale, A. The Role of Antioxidants in the Interplay between Oxidative Stress and Senescence. Antioxidants 2022, 11, 1224. https://doi.org/10.3390/antiox11071224
Varesi A, Chirumbolo S, Campagnoli LIM, Pierella E, Piccini GB, Carrara A, Ricevuti G, Scassellati C, Bonvicini C, Pascale A. The Role of Antioxidants in the Interplay between Oxidative Stress and Senescence. Antioxidants. 2022; 11(7):1224. https://doi.org/10.3390/antiox11071224
Chicago/Turabian StyleVaresi, Angelica, Salvatore Chirumbolo, Lucrezia Irene Maria Campagnoli, Elisa Pierella, Gaia Bavestrello Piccini, Adelaide Carrara, Giovanni Ricevuti, Catia Scassellati, Cristian Bonvicini, and Alessia Pascale. 2022. "The Role of Antioxidants in the Interplay between Oxidative Stress and Senescence" Antioxidants 11, no. 7: 1224. https://doi.org/10.3390/antiox11071224