
Integrative Comparative Assessment of Cold Acclimation in Evergreen and Deciduous Iris Species

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Evaluation of Leaf Freezing Tolerance
2.3. Morphological Measurements and Growth Indices
2.4. Observation of Leaf Microstructures after Exposure to Low Temperatures
2.5. Determination of Carbohydrate Concentrations
2.6. Measurement of Leaf Phytohormone Levels
2.7. Measurements of MDA, Proline, SP Contents and SOD Activities in Leaf Tissues
2.8. Expression Analysis of Genes Involved in Crucial Pathways during Natural Cold Acclimation and Deacclimation
2.9. Statistical Analyses
3. Results
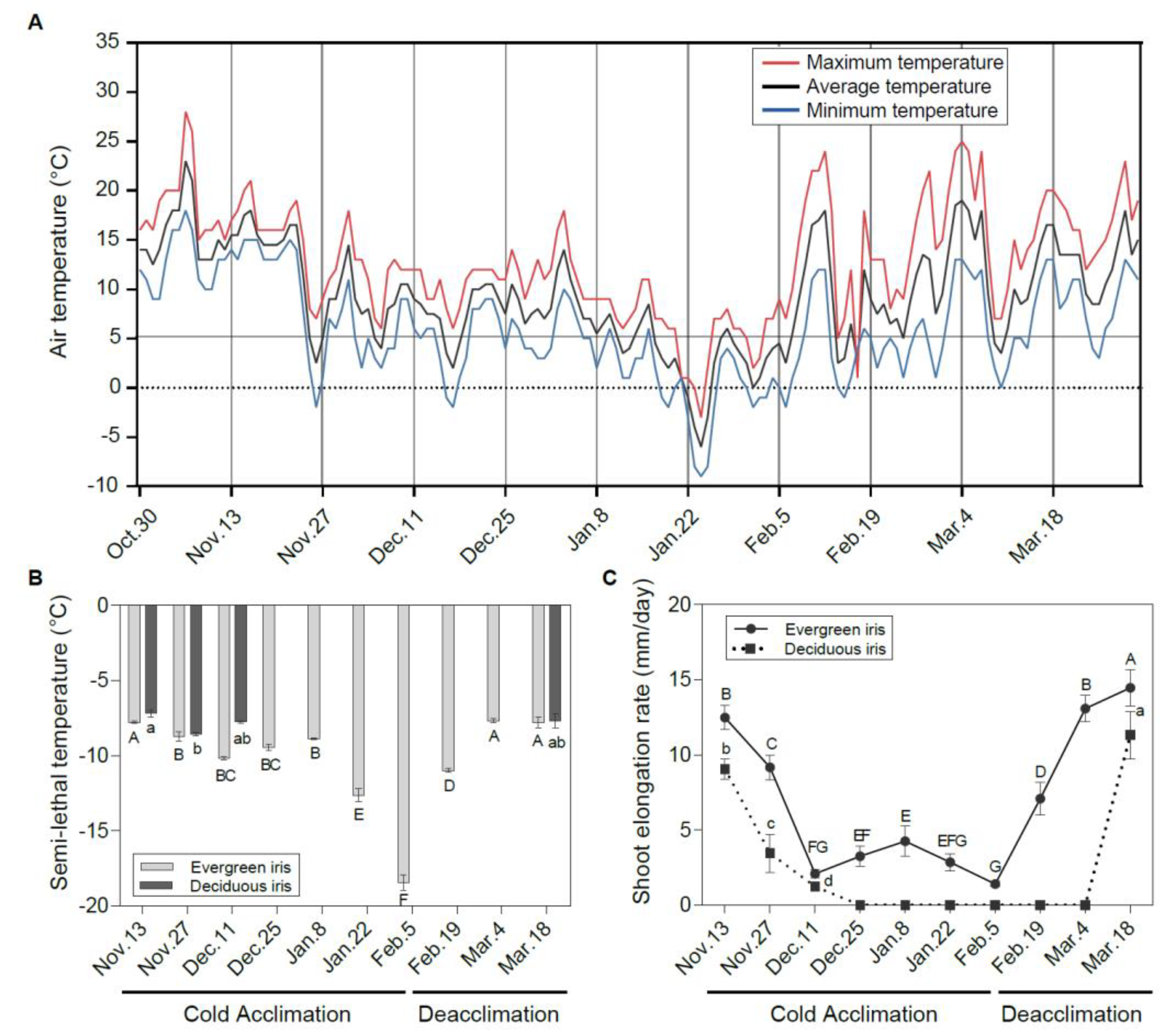
3.1. Changes in Leaf Freezing Tolerance of Two Iris Species under Natural Cold Acclimation and Deacclimation
3.2. Changes in Vegetative Growth Status of Two Iris Species during Overwintering
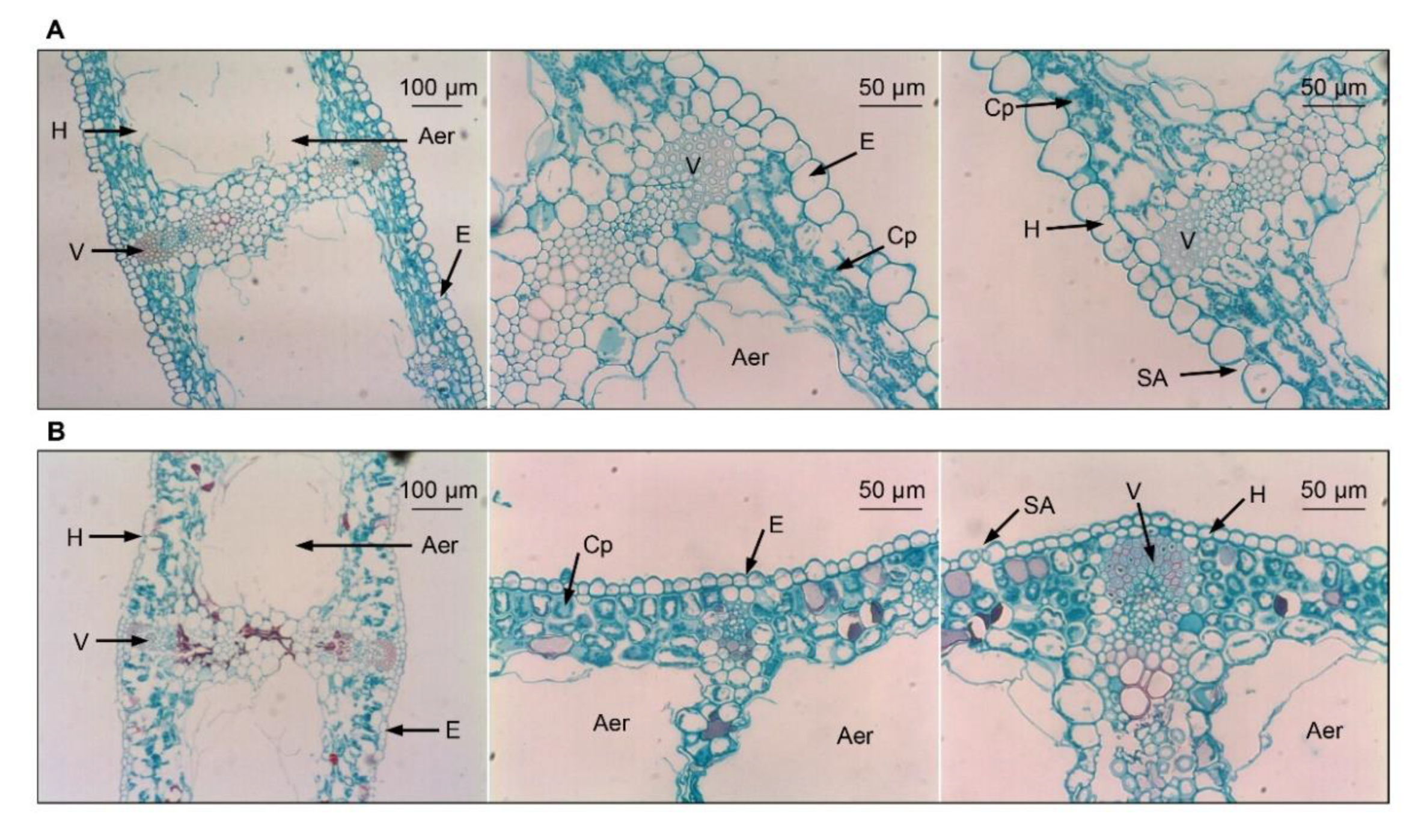
3.3. Leaf Microstructure Comparisons between Two Iris Species after Exposure to Low Temperatures
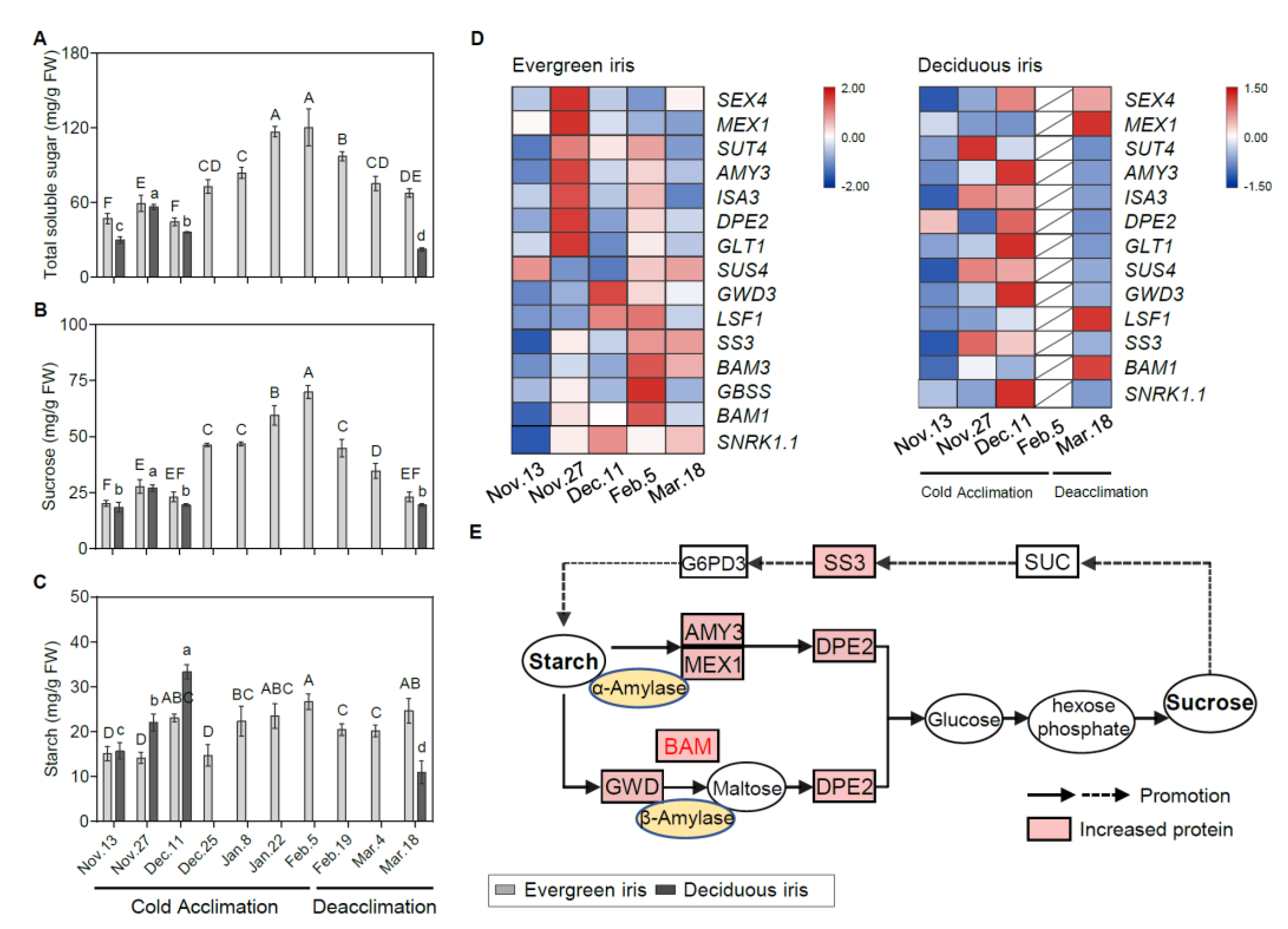
3.4. Alterations in Carbohydrate Concentrations and the Expression of Related Genes in Leaf Tissues
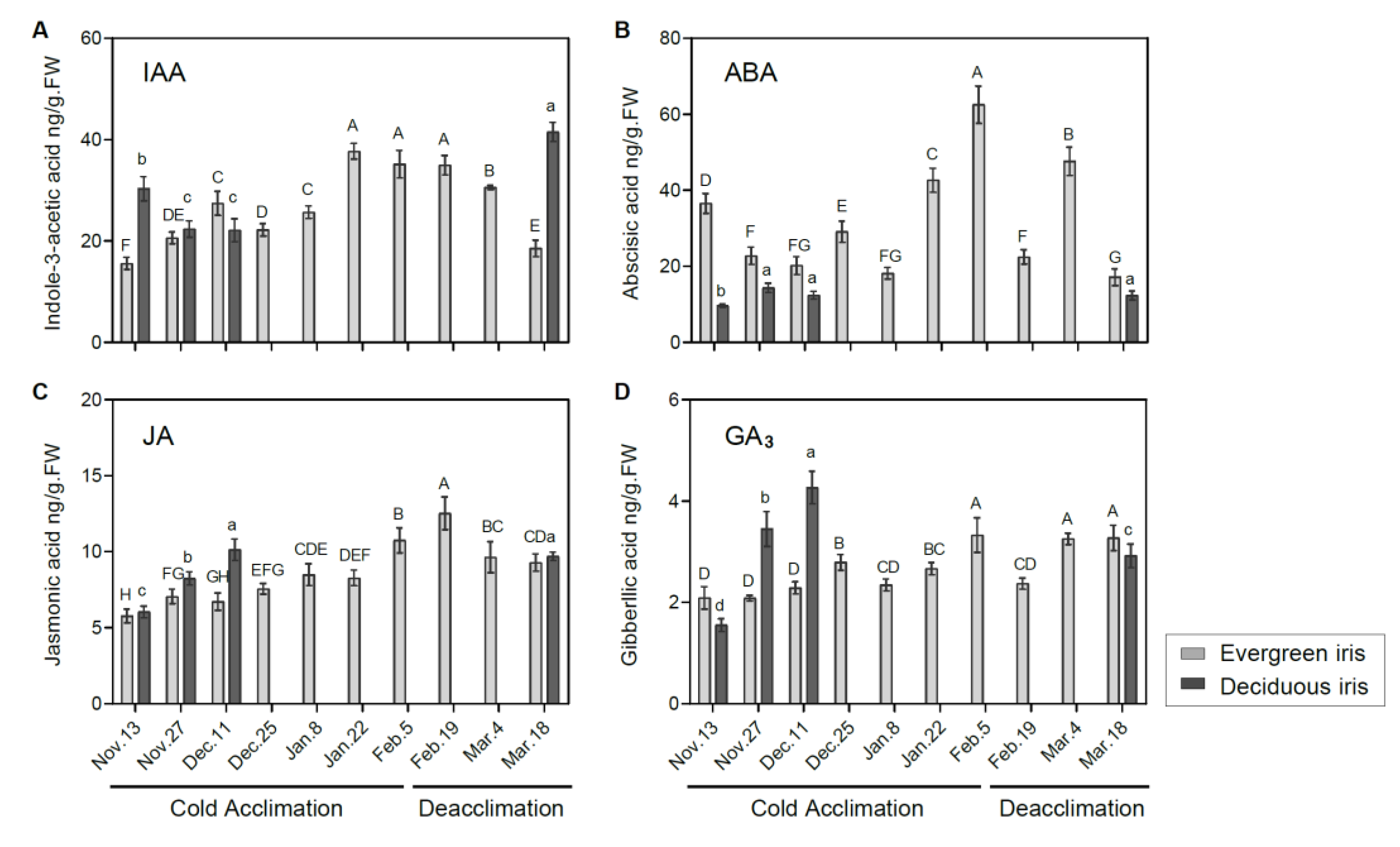
3.5. Effects of Cold Acclimation and Deacclimation on Phytohormone Levels in Leaf Tissues
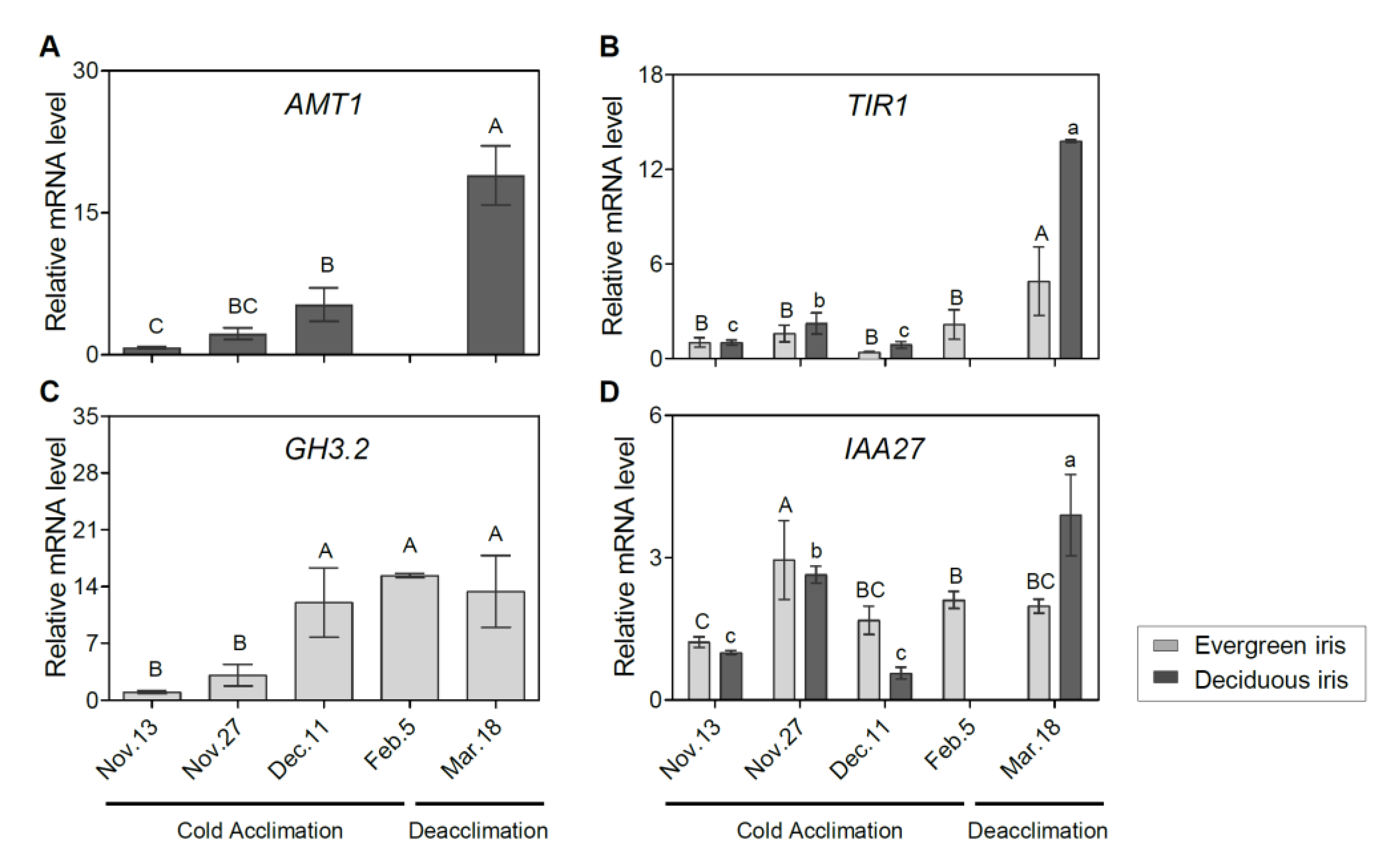
3.6. Expression Changes in Genes Associated with Phytohormone Biosynthesis and Signal Transduction in Leaf Tissues
3.7. Changes in MDA, Proline and Soluble Protein Contents and SOD Activity in Leaf Tissues and the Expression of Related Genes
4. Discussion
4.1. Different Cold Acclimation Characteristics in Evergreen and Deciduous Irises
4.2. Cold Acclimation Activates Carbohydrate Accumulation in Response to Cold for Both Evergreen and Deciduous Irises
4.3. Cold Acclimation Integrates Phytohormone Signaling to Withstand Low Temperature
4.4. Genes Belonging to the Antioxidant System Were Mainly Induced during Deacclimation
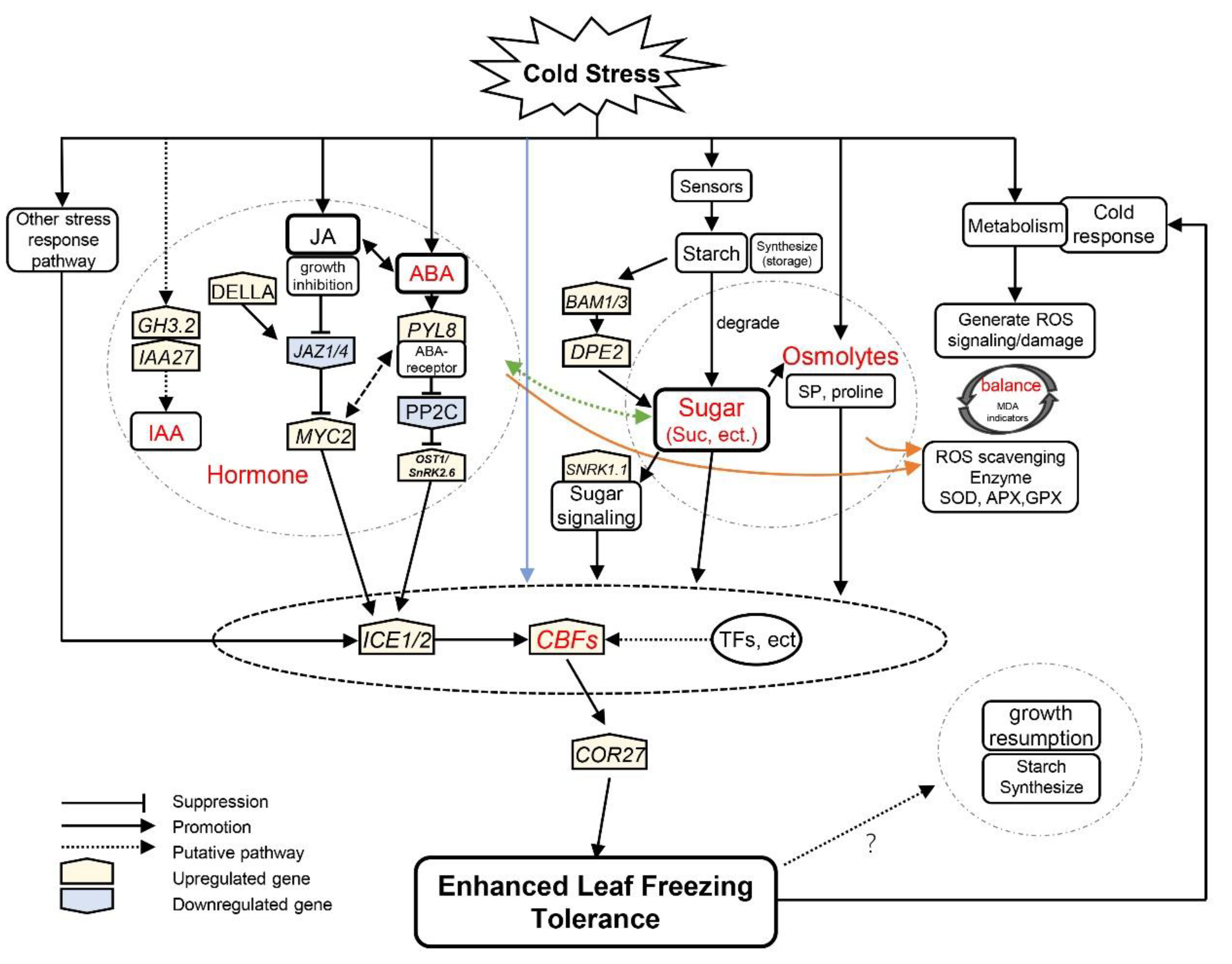
4.5. The ICE1–CBF–COR Transcriptional Cascade Integrates Multiple Stress Response Pathways during Cold Acclimation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, J.A.; Taliaferro, C.M. Freeze Tolerance of Seed-producing Turf Bermudagrasses. Crop Sci. 2002, 42, 190–192. [Google Scholar] [PubMed]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.; Clement, C.; Barka, E.A. Physiological and Molecular Changes in Plants Grown at Low Temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Gronvold, L.; Sandve, S.R.; Hvidsten, T.R.; Fjellheim, S. Evolution of Cold Acclimation and Its Role in Niche Transition in the Temperate Grass Subfamily Pooideae. Plant Physiol. 2019, 180, 404–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagter, M.; Arora, R. Winter Survival and Deacclimation of Perennials under Warming Climate: Physiological perspectives. Physiol. Plant. 2013, 147, 75–87. [Google Scholar] [CrossRef]
- Hincha, D.K.; Zuther, E. Introduction: Plant Cold Acclimation and Winter Survival. Methods Mol. Biol. 2020, 2156, 1–7. [Google Scholar]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Novak, A.; Boldizsar, A.; Adam, E.; Kozma-Bognar, L.; Majlath, I.; Baga, M.; Toth, B.; Chibbar, R.; Galiba, G. Light-quality and Temperature-dependent CBF14 Gene Expression Modulates Freezing Tolerance in Cereals. J. Exp. Bot. 2016, 67, 1285–1295. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Ding, Y.; Yang, S. Molecular Regulation of CBF Signaling in Cold Acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef]
- Wingler, A.; Tijero, V.; Muller, M.; Yuan, B.; Munne-Bosch, S. Interactions between Sucrose and Jasmonate Signalling in the Response to Cold Stress. BMC Plant Biol. 2020, 20, 176. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Zhu, X.; Duan, N.; Liu, J.H. PtrBAM1, a Beta-amylase-coding Gene of Poncirus trifoliata, is a CBF Regulon Member with Function in Cold Tolerance by Modulating Soluble Sugar Levels. Plant Cell Environ. 2014, 37, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.L.; Wang, L.; Zhou, Y.H.; Huang, Y.T.; Hao, X.Y.; Wang, Y.C.; Wang, B.; Yang, Y.J.; Wang, X.C. Effects of Cold Acclimation on Sugar Metabolism and Sugar-related Gene Expression in Tea Plant during the Winter Season. Plant Mol Biol. 2015, 88, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Ahres, M.; Palmai, T.; Gierczik, K.; Dobrev, P.; Vankova, R.; Galiba, G. The Impact of Far-Red Light Supplementation on Hormonal Responses to Cold Acclimation in Barley. Biomolecules 2021, 11, 450. [Google Scholar] [CrossRef]
- Lv, X.; Li, H.; Chen, X.; Xiang, X.; Guo, Z.; Yu, J.; Zhou, Y. The Role of Calcium-dependent Protein Kinase in Hydrogen Peroxide, Nitric Oxide and ABA-dependent Cold Acclimation. J. Exp. Bot. 2018, 69, 4127–4139. [Google Scholar] [CrossRef]
- Wang, H.; Blakeslee, J.J.; Jones, M.L.; Chapin, L.J.; Dami, I.E. Exogenous Abscisic Acid Enhances Physiological, Metabolic, and Transcriptional Cold Acclimation Responses in Greenhouse-grown Grapevines. Plant Sci. 2020, 293, 110437. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 Family Member, OsGH3-2, Modulates Auxin and Abscisic Acid Levels and Differentially Affects Drought and Cold Tolerance in Rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate Regulates the Inducer of CBF Expression-C-repeat Binding Factor/DRE Binding Factor1 Cascade and Freezing Tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Guo, Z.; Li, H.; Wang, M.; Onac, E.; Zhou, J.; Xia, X.; Shi, K.; Yu, J.; Zhou, Y. Phytochrome A and B Function Antagonistically to Regulate Cold Tolerance via Abscisic Acid-Dependent Jasmonate Signaling. Plant Physiol. 2016, 170, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Han, X.; Wang, Q.; Wang, X.; Chen, X.; Li, L.; Fu, X.; Gao, D. EARLY BUD BREAK 1 Triggers Bud Break in Peach Trees by Regulating Hormone Metabolism, the Cell Cycle, and Cell Wall Modifications. J. Exp. Bot. 2020, 71, 3512–3523. [Google Scholar] [CrossRef]
- Li, N.N.; Yue, C.; Cao, H.L.; Qian, W.J.; Hao, X.Y.; Wang, Y.C.; Wang, L.; Ding, C.Q.; Wang, X.C.; Yang, Y.J. Transcriptome Sequencing Dissection of the Mechanisms Underlying Differential Cold Sensitivity in Young and Mature Leaves of the Tea Plant (Camellia sinensis). J Plant Physiol. 2018, 224, 144–155. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar Signalling and Antioxidant Network Connections in Plant Cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef] [PubMed]
- Trovato, M.; Mattioli, R.; Costantino, P. Multiple Roles of Proline in Plant Stress Tolerance and Development. Rend. Lincei.-Sci. Fis. Nat. 2008, 19, 325–346. [Google Scholar] [CrossRef]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Morris, P.; Ribeiro, D.; Wilson, I. Nitric Oxide, Stomatal Closure, and Abiotic Stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, L.; Leus, L.; De Keyser, E.; Van Labeke, M.C. Seasonal Changes in Cold Hardiness and Carbohydrate Metabolism in Four Garden Rose Cultivars. J. Plant Physiol. 2019, 232, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, J.; Zhang, J.; Li, K.; Xia, Y. Green Period Characteristics and Foliar Cold Tolerance in 12 Iris Species and Cultivars in the Yangtze Delta, China. HortTechnology 2017, 27, 399–407. [Google Scholar] [CrossRef]
- Cheng, L.; Feng, S.; Xiao, Y.; Yu, F.; Zhu, S.; Wang, X. Advances in Molecular Phylogeny of Iris spp. Guihaia. Front. Plant Sci. 2021, 12, 756300. [Google Scholar]
- Wilson, C. Phylogenetic Relationships Among the Recognized Series in Iris Section Limniris. Syst. Bot. 2009, 34, 277–284. [Google Scholar] [CrossRef]
- Li, D.; Xia, Y.; Lou, J.; Zhang, D.; Wang, X.; Zhang, J. A Comparative Study between Evergreen and Deciduous Daylily Species Reveals the Potential Contributions of Winter Shoot Growth and Leaf Freezing Tolerance to Foliar Habits. J. Plant Growth Regul. 2019, 39, 1030–1045. [Google Scholar] [CrossRef]
- Wang, X.; Li, D.; Zhang, D.; Shi, X.; Wu, Y.; Qi, Z.; Ding, H.; Zhu, K.; Xia, Y.; Zhang, J. Improving Crucial Details and Selecting the Optimal Model for Evaluating the Chilling Requirement of Paeonia lactiflora Pall. at Low Latitudes during Four Winters. Sci. Hortic. 2020, 265, 109175. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, F.D. The LT50 and Cold Tolerance Adaptability of Chrysanthemum during a Natural Drop in Temperature. Acta Hortic Sin. 2008, 35, 559–564. [Google Scholar]
- Liu, B.; Zhou, H.; Cao, S.; Xia, Y.; Arora, R. Comparative Physiology of Natural Deacclimation in Ten Azalea Cultivars. Hort. Sci. 2017, 52, 1451–1457. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.P. Improvement of Traditional Paraffin Section Preparation Methods. J. Biol. 2006, 4, 45–46. [Google Scholar]
- Sun, T.X.; Jiang, X.S. H2O2-CH2COOH Maceration-a Simple and Perfect Method for Leaf Epidermis. Guihaia 2009, 29, 44–47. [Google Scholar]
- Wang, X.; Zhang, R.; Huang, Q.; Shi, X.; Li, D.; Shao, L.; Xu, T.; Horvath, D.P.; Xia, Y.; Zhang, J. Comparative Study on Physiological Responses and Gene Expression of Bud Endodormancy Release Between Two Herbaceous Peony Cultivars (Paeonia lactiflora Pall.) with Contrasting Chilling Requirements. Front. Plant Sci. 2021, 12, 772285. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sun, M.; Zhang, J.; Zhang, L.; Ren, Z.; Min, R.; Wang, X.; Xia, Y. Differential Effects of Paclobutrazol on the Bulblet Growth of Oriental Lily Cultured In Vitro: Growth Behavior, Carbohydrate Metabolism, and Antioxidant Capacity. J. Plant Growth Regul. 2018, 38, 359–372. [Google Scholar] [CrossRef]
- Wang, L.; Yao, L.; Hao, X.; Li, N.; Wang, Y.; Ding, C.; Lei, L.; Qian, W.; Zeng, J.; Yang, Y.; et al. Transcriptional and Physiological Analyses Reveal the Association of ROS Metabolism with Cold Tolerance in Tea Plant. Environ. Exp. Bot. 2019, 160, 45–58. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S.; Wang, W. Hormonal Changes in the Grains of Rice Subjected to Water Stress during Grain Filling. Plant Physiol. 2001, 127, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Fan, Z.; Qiu, L.; Che, Q.; Wang, T.; Li, Y.; Wang, Y. MdbHLH130, an Apple bHLH Transcription Factor, Confers Water Stress Resistance by Regulating Stomatal Closure and ROS Homeostasis in Transgenic Tobacco. Front. Plant Sci. 2020, 11, 543696. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantification of Microgram Quantities of Protein Utilizing the Principle of Protein Dye-binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher-plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Shao, L.; Zhang, J.; Wang, X.; Zhang, D.; Horvath, D.P.; Zhang, L.; Zhang, J.; Xia, Y. MADS-box Transcription Factors Determine the Duration of Temporary Winter Dormancy in Closely Related Evergreen and Deciduous Iris spp. J. Exp. Bot. 2022, 73, 1429–1449. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, Y.; Chang, L.; Zhang, T.; An, J.; Liu, Y.; Cao, Y.; Zhao, X.; Sha, X.; Hu, T.; et al. MsZEP, a Novel Zeaxanthin Epoxidase Gene from Alfalfa (Medicago sativa), Confers Ddrought and Salt Tolerance in Transgenic Tobacco. Plant Cell Rep. 2016, 35, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.X.; Li, J. The Effects of Exogenous ABA Applied to Maize (Zea mays L.) Roots on Plant Responses to Chilling Stress. Acta Physiol. Plant. 2018, 40, 77. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts Across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Wen, L.; Yun, X.; Zheng, X.; Xu, H.; Zhan, R.; Chen, W.; Xu, Y.; Chen, Y.; Zhang, J. Transcriptomic Comparison Reveals Candidate Genes for Triterpenoid Biosynthesis in Two Closely Related Ilex Species. Front. Plant Sci. 2017, 8, 634. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Lin, L.; Jiang, P.; Wang, D.; Xing, Y. A Comparison of RNA-Seq and High-density Exon Array for Detecting Differential Gene Expression between Closely Related Species. Nucleic Acids Res. 2011, 39, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Oh, D.H.; Duan, L.; Ramachandran, P.; Ramirez, A.; Bartlett, A.; Tran, K.N.; Wang, G.; Dassanayake, M.; Dinneny, J.R. Divergence in the ABA Gene Regulatory Network Underlies Differential Growth Control. Nat. Plants. 2022, 10, 1038. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Leus, L.; De Keyser, E.; Van Labeke, M.C. Cold Acclimation and Deacclimation of Two Garden Rose Cultivars Under Controlled Daylength and Temperature. Front. Plant Sci. 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Hong, X.; Hu, D.; Liu, C.; Yang, J.; Li, Y.; Huang, Y.; Feng, Y.; Gong, H.; et al. Functional Inactivation of UDP-N-acetylglucosamine Pyrophosphorylase 1 (UAP1) Induces Early Leaf Senescence and Defence Responses in Rice. J. Exp. Bot. 2015, 66, 973–987. [Google Scholar] [CrossRef] [Green Version]
- Stefania, T.; Alice, T.; Antonio, F.; Daniela, R.J.E.H. Physiological Mechanisms for Delaying the Leaf Yellowing of Potted Geranium Plants. Sci. Hortic. 2018, 242, 146–154. [Google Scholar]
- Lim, P.O.; Kim, H.J.; Nam, H.G. Leaf senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Arora, R.; Wisniewski, M.E.; Scorza, R. Cold Acclimation in Genetically Related (Sibling) Deciduous and Evergreen Peach (Prunus persica [L.] Batsch): I. Seasonal Changes in Cold Hardiness and Polypeptides of Bark and Xylem Tissues. Plant Physiol. 1992, 99, 1562–1568. [Google Scholar] [CrossRef] [Green Version]
- Yano, R.; Nakamura, M.; Yoneyama, T.; Nishida, I. Starch-related Alpha-glucan/water Dikinase is Involved in the Cold-induced Development of Freezing Tolerance in Arabidopsis. Plant Physiol. 2005, 138, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Yang, M.; Hu, D.; Yang, Z.; Ma, S.; Li, X.; Xiong, L. The OsMYB30 Transcription Factor Suppresses Cold Tolerance by Interacting with a JAZ Protein and Suppressing beta-Amylase Expression. Plant Physiol. 2017, 173, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, Y.; Liu, X.; Huang, J.; Wang, Q.; Gu, J.; Lu, Y. Transcriptome Profiling of the Cold Response and Signaling Pathways in Lilium lancifolium. BMC Genom. 2014, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Xie, S.; Nie, L.; Zheng, Y.; Wang, J.; Huang, J.; Zhao, M.; Zhu, S.; Hou, J.; Chen, G.; et al. Comparative Proteomics Reveals Cold Acclimation Machinery Through Enhanced Carbohydrate and Amino Acid Metabolism in Wucai (Brassica Campestris L.). Plants 2019, 8, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicher, R. Carbon Partitioning and the Impact of Starch Deficiency on the Initial Response of Arabidopsis to Chilling Temperatures. Plant Sci. 2011, 181, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Teng, L.; Yan, X.; Zhao, M.; Shen, S. The Cold Responsive Mechanism of the Paper Mulberry: Decreased Photosynthesis Capacity and Increased Starch Accumulation. BMC Genom. 2015, 16, 898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiaens, A.; De Keyser, E.; Lootens, P.; Pauwels, E.; Roldan-Ruiz, I.; De Riek, J.; Gobin, B.; Van Labeke, M.C. Cold Storage to Overcome Dormancy Affects the Carbohydrate Status and Photosynthetic Capacity of Rhododendron simsii. Plant Biol. 2015, 17, 97–105. [Google Scholar] [CrossRef]
- Liu, Y.S.; Geng, J.C.; Sha, X.Y.; Zhao, Y.X.; Hu, T.M.; Yang, P.Z. Effect of Rhizobium Symbiosis on Low-Temperature Tolerance and Antioxidant Response in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2019, 10, 538. [Google Scholar] [CrossRef]
- Maruyama, K.; Takeda, M.; Kidokoro, S.; Yamada, K.; Sakuma, Y.; Urano, K.; Fujita, M.; Yoshiwara, K.; Matsukura, S.; Morishita, Y.; et al. Metabolic Pathways Involved in Cold Acclimation Identified by Integrated Analysis of Metabolites and Transcripts Regulated by DREB1A and DREB2A. Plant Physiol. 2009, 150, 1972–1980. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, F.; Guy, C.L. β-Amylase Induction and the Protective Role of Maltose during Temperature Shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Tong, Z.; He, F.; Li, X. Response of Alfalfa (Medicago sativa L.) to Abrupt Chilling as Reflected by Changes in Freezing Tolerance and Soluble Sugars. Agronomy 2020, 10, 255. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, J.; Zhang, W.; Yan, S.; Wang, R.; Zhao, J.; Li, Y.; Qi, Z.; Sun, Z.; Zhu, Z. The Putative Auxin Efflux Carrier OsPIN3t is Involved in the Drought Stress Response and Drought Tolerance. Plant J. 2012, 72, 805–816. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Huang, Z.; Mi, L.; Xu, K.; Wu, J.; Fan, Y.; Ma, S.; Jiang, D. Effects of Low Temperature at Booting Stage on Sucrose Metabolism and Endogenous Hormone Contents in Winter Wheat Spikelet. Front. Plant Sci. 2019, 10, 498. [Google Scholar] [CrossRef]
- Shoji, K.; Addicott, F.T.; Swets, W.A. Auxin in Relation to Leaf Blade Abscission. Plant Physiol. 1951, 26, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Majlath, I.; Szalai, G.; Soos, V.; Sebestyen, E.; Balazs, E.; Vankova, R.; Dobrev, P.I.; Tari, I.; Tandori, J.; Janda, T. Effect of Light on the Gene Expression and Hormonal Status of Winter and Spring Wheat Plants during Cold Hardening. Physiol. Plant 2012, 145, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, Z.; Li, X.; Lv, T.; Liu, H.; Wang, L.; Niu, H.; Bu, Q. Characterization and Functional Analysis of Pyrabactin Resistance-Like Abscisic Acid Receptor Family in Rice. Rice 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Ge, H.; Wang, X.; Tang, R.; Wang, Y.; Zhao, F.; Lan, W.; Luan, S.; Yang, L. Overexpression of Pyrabactin Resistance-Like Abscisic Acid Receptors Enhances Drought, Osmotic, and Cold Tolerance in Transgenic Poplars. Front. Plant Sci. 2017, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Rubio, S.; Noriega, X.; Pérez, F.J. Abscisic acid (ABA) and Low Temperatures Synergistically Increase the Expression of CBF/DREB1 Transcription Factors and Cold-hardiness in Grapevine Dormant Buds. Ann. Bot. 2019, 123, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, Y.; Han, X.; Wang, H.; Pan, J.; Yu, D. Jasmonate Regulates Leaf Senescence and Tolerance to Cold Stress: Crosstalk with Other Phytohormones. J. Exp. Bot. 2017, 68, 1361–1369. [Google Scholar] [CrossRef]
- Kalapos, B.; Dobrev, P.; Nagy, T.; Vitamvas, P.; Gyorgyey, J.; Kocsy, G.; Marincs, F.; Galiba, G. Transcript and Hormone Analyses Reveal the Involvement of ABA-signalling, Hormone Crosstalk and Genotype-specific Biological Processes in Cold-shock Response in Wheat. Plant Sci. 2016, 253, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liu, H.; Xiong, L. Endogenous Auxin and Jasmonic Acid Levels are Differentially Modulated by Abiotic Stresses in Rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Juvany, M.; Cuthbert, C.; Munne-Bosch, S. Adaptation to Altitude Affects the Senescence Response to Chilling in the Perennial Plant Arabis alpina. J. Exp. Bot. 2015, 66, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.R.; Harberd, N.P. Integration of Plant Responses to Environmentally Activated Phytohormonal Signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.; Duan, X.; Jiang, Y.; Zhang, P. Increased Expression of Native Cytosolic Cu/Zn Superoxide Dismutase and Ascorbate Peroxidase Improves Tolerance to Oxidative and Chilling Stresses in Cassava (Manihot esculenta Crantz). BMC Plant Biol. 2014, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous Abscisic Acid Increases Antioxidant Enzymes and Related Gene Expression in Pepper (Capsicum annuum) Leaves Subjected to Chilling Stress. Genet. Mol. Res. 2012, 11, 4063–4080. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Cang, J.; Lu, Q.; Fan, B.; Xu, Q.; Li, W.; Wang, X. ABA Enhanced Cold Tolerance of Wheat dn1 via Increasing ROS Scavenging System. Plant Signal. Behav. 2020, 15, 1780403. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, J.; Chang, Q.; Gu, C.; Song, A.; Chen, S.; Dong, B.; Chen, F. Cold Acclimation Induces Freezing Tolerance via Antioxidative Enzymes, Proline Metabolism and Gene Expression Changes in Two Chrysanthemum Species. Mol. Biol. Rep. 2014, 41, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Chai, H.; Yin, H.; Yang, M.; Hu, G.; Guo, M.; Yi, R.; Zhang, P. Full-length Transcriptome Sequencing Reveals the Low-temperature-tolerance Mechanism of Medicago falcata Roots. BMC Plant Biol. 2019, 19, 575. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A Regulator of Cold-induced Transcriptome and Freezing Tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.F.; Kang, H.G.; Park, M.Y.; Jeong, H.; Sun, H.J.; Song, P.S.; Lee, H.Y. Zoysia japonica MYC type Transcription Factor ZjICE1 Regulates Cold Tolerance in Transgenic Arabidopsis. Plant Sci. 2019, 289, 110254. [Google Scholar] [CrossRef]
- Kyonoshin, M.; Daisuke, T.; Junya, M.; Takuya, Y.; Satoshi, K.; Satoko, M.; Hironori, T.; Tetsuya, S.; Yamamoto, Y.Y.; Kyouko, Y.J.D.R. Identification of Cis-Acting Promoter Elements in Cold- and Dehydration-Induced Transcriptional Pathways in Arabidopsis, Rice, and Soybean. DNA Res. 2011, 19, 37–49. [Google Scholar]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschik, P. The Cold-inducible CBF1 Factor-dependent Signaling Pathway Modulates the Accumulation of the Growth-repressing DELLA Proteins via Its Effect on Gibberellin Metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Beckles, D.M. Dynamic Changes in the Starch-sugar Interconversion within Plant Source and Sink Tissues Promote a Better Abiotic Stress Response. J. Plant Physiol. 2019, 234, 80–93. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evergreen Iris | LT50 | SER | RCC | LW | FW | NFL | TSS | SUC | STA | IAA | ABA | JA | GA3 | SOD | MDA | PRO | SP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LT50 | 1 | ||||||||||||||||
| SER | 0.68 * | 1 | |||||||||||||||
| RCC | 0.55 | 0.45 | 1 | ||||||||||||||
| LW | 0.60 | 0.90 ** | 0.61 | 1 | |||||||||||||
| FW | 0.66 * | 0.85 ** | 0.61 | 0.93 ** | 1 | ||||||||||||
| NFL | −0.14 | 0.28 | −0.55 | 0.20 | 0.19 | 1 | |||||||||||
| TSS | −0.76 * | −0.45 | −0.86 ** | −0.61 | −0.68 * | 0.44 | 1 | ||||||||||
| SUC | −0.82 ** | −0.67 * | −0.82 ** | −0.82 ** | −0.85 ** | 0.24 | 0.94 ** | 1 | |||||||||
| STA | −0.56 | −0.29 | −0.61 | −0.23 | −0.17 | 0.59 | 0.55 | 0.44 | 1 | ||||||||
| IAA | −0.68 * | −0.55 | −0.88 ** | −0.56 | −0.69 * | 0.36 | 0.81 ** | 0.76 * | 0.57 | 1 | |||||||
| ABA | −0.65 * | −0.18 | −0.48 | −0.31 | −0.38 | 0.21 | 0.56 | 0.60 | 0.25 | 0.48 | 1 | ||||||
| JA | −0.43 | −0.05 | −0.61 | −0.08 | −0.26 | 0.75 * | 0.68 * | 0.52 | 0.51 | 0.65 * | 0.22 | 1 | |||||
| GA | −0.33 | 0.10 | −0.57 | −0.01 | 0.09 | 0.82 ** | 0.46 | 0.38 | 0.56 | 0.29 | 0.52 | 0.50 | 1 | ||||
| SOD | −0.30 | −0.49 | −0.48 | −0.43 | −0.31 | 0.18 | 0.40 | 0.42 | 0.40 | 0.38 | −0.24 | 0.33 | 0.24 | 1 | |||
| MDA | 0.50 | 0.50 | 0.21 | 0.42 | 0.56 | 0.11 | −0.27 | −0.47 | 0.23 | −0.37 | −0.44 | −0.26 | −0.08 | −0.09 | 1 | ||
| PRO | −0.65 * | −0.31 | −0.43 | −0.46 | −0.45 | 0.38 | 0.70 * | 0.75 * | 0.28 | 0.26 | 0.46 | 0.52 | 0.53 | 0.26 | −0.42 | 1 | |
| SP | −0.14 | −0.05 | −0.67 * | −0.17 | −0.08 | 0.77 ** | 0.51 | 0.42 | 0.48 | 0.39 | −0.01 | 0.67 * | 0.73 * | 0.69 * | −0.03 | 0.44 | 1 |
| Deciduous Iris | LT50 | SER | RCC | LW | FW | NFL | TSS | SUC | STA | IAA | ABA | JA | GA3 | SOD | MDA | PRO | SP |
| LT50 | 1 | ||||||||||||||||
| SER | 0.54 | 1 | |||||||||||||||
| RCC | 0.58 | 0.90 | 1 | ||||||||||||||
| LW | 0.38 | 0.23 | 0.63 | 1 | |||||||||||||
| FW | 0.23 | 0.90 | 0.65 | −0.18 | 1 | ||||||||||||
| NFL | 0.12 | −0.12 | 0.32 | 0.93 | −0.48 | 1 | |||||||||||
| TSS | −0.85 | −0.70 | −0.52 | 0.08 | −0.61 | 0.39 | 1 | ||||||||||
| SUC | −0.94 | −0.43 | −0.35 | −0.05 | −0.24 | 0.19 | 0.92 | 1 | |||||||||
| STA | −0.29 | −0.96 * | −0.89 | −0.29 | −0.89 | 0.02 | 0.46 | 0.14 | 1 | ||||||||
| IAA | 0.46 | 0.93 | 0.69 | −0.13 | 0.97 * | −0.47 | −0.79 | −0.48 | −0.85 | 1 | |||||||
| ABA | −0.96 * | −0.50 | −0.66 | −0.62 | −0.11 | −0.39 | 0.68 | 0.81 | 0.30 | −0.33 | 1 | ||||||
| JA | −0.35 | −0.32 | −0.69 | −0.99 ** | 0.07 | −0.90 | −0.07 | 0.00 | 0.39 | 0.04 | 0.59 | 1 | |||||
| GA | −0.61 | −0.75 | −0.96 * | −0.82 | −0.40 | −0.56 | 0.39 | 0.32 | 0.74 | −0.47 | 0.75 | 0.86 | 1 | ||||
| SOD | −0.29 | −0.73 | −0.36 | 0.49 | −0.91 | 0.77 | 0.75 | 0.44 | 0.62 | −0.92 | 0.08 | −0.42 | 0.10 | 1 | |||
| MDA | 0.98 * | 0.35 | 0.46 | 0.43 | 0.02 | 0.23 | −0.73 | −0.91 | −0.11 | 0.26 | −0.96 * | −0.37 | −0.54 | −0.10 | 1 | ||
| PRO | −0.54 | −0.81 | −0.98 * | −0.75 | −0.51 | −0.49 | 0.38 | 0.26 | 0.83 | −0.55 | 0.66 | 0.81 | 0.99 * | 0.19 | −0.44 | 1 | |
| SP | 0.81 | 0.61 | 0.39 | −0.19 | 0.55 | −0.47 | −0.99 ** | −0.93 | −0.35 | 0.74 | −0.63 | 0.19 | −0.26 | −0.74 | 0.71 | −0.25 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, L.; Xu, T.; Wang, X.; Zhang, R.; Wang, X.; Ren, Z.; Zhang, J.; Xia, Y.; Li, D. Integrative Comparative Assessment of Cold Acclimation in Evergreen and Deciduous Iris Species. Antioxidants 2022, 11, 977. https://doi.org/10.3390/antiox11050977
Shao L, Xu T, Wang X, Zhang R, Wang X, Ren Z, Zhang J, Xia Y, Li D. Integrative Comparative Assessment of Cold Acclimation in Evergreen and Deciduous Iris Species. Antioxidants. 2022; 11(5):977. https://doi.org/10.3390/antiox11050977
Chicago/Turabian StyleShao, Lingmei, Tong Xu, Xiaobin Wang, Runlong Zhang, Xiuyun Wang, Ziming Ren, Jiaping Zhang, Yiping Xia, and Danqing Li. 2022. "Integrative Comparative Assessment of Cold Acclimation in Evergreen and Deciduous Iris Species" Antioxidants 11, no. 5: 977. https://doi.org/10.3390/antiox11050977