
Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site, Conditions, and Plant Materials
2.2. Chemicals
2.3. Ash, Fiber, Moisture, Fat, Gross Energy, Carbohydrate, and Protein Estimation
2.4. Elements Estimation
2.5. Beta-Carotene
2.6. Ascorbic Acid (AsA) Estimation
2.7. Samples Extraction and Determination of Total Polyphenols (TP), Total Flavonoids (TF), and Antiradical Potential (ARP)
2.8. Samples Extraction and Determination of Phenolic Acids (PAs) by HPLC
2.9. Statistical Analysis
3. Results and Discussion
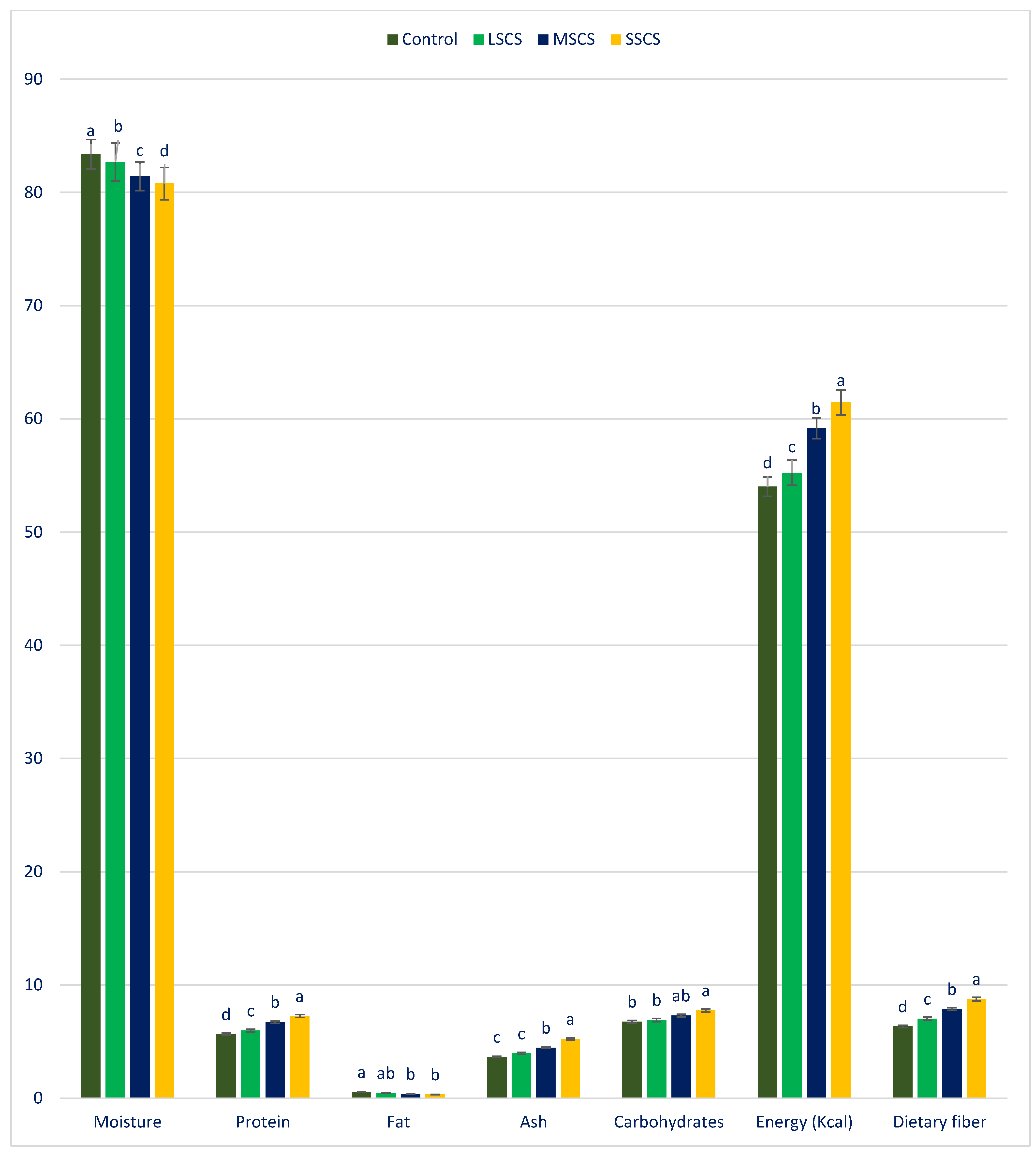
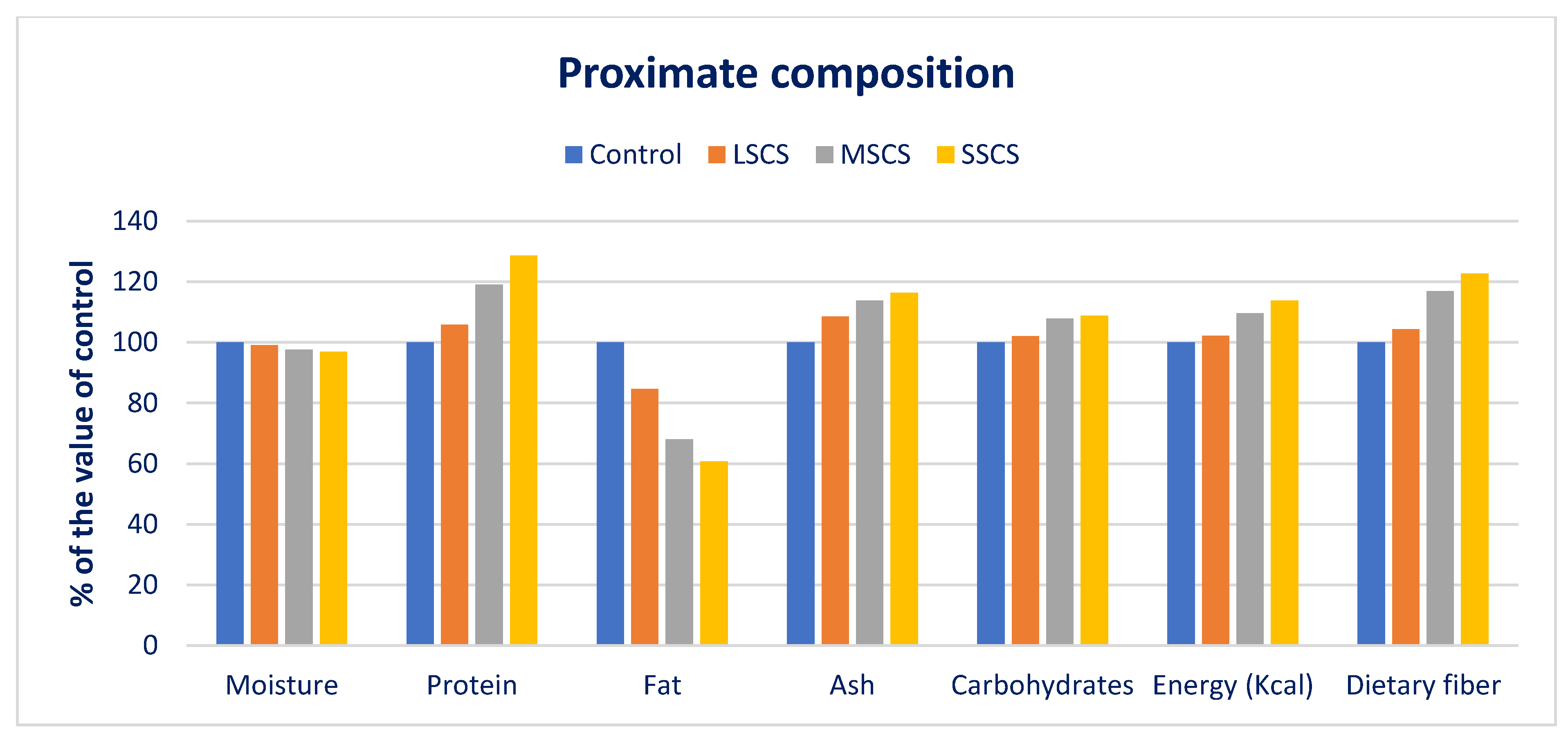
3.1. The Response of Proximate Compositions to Sodium Chloride Stress
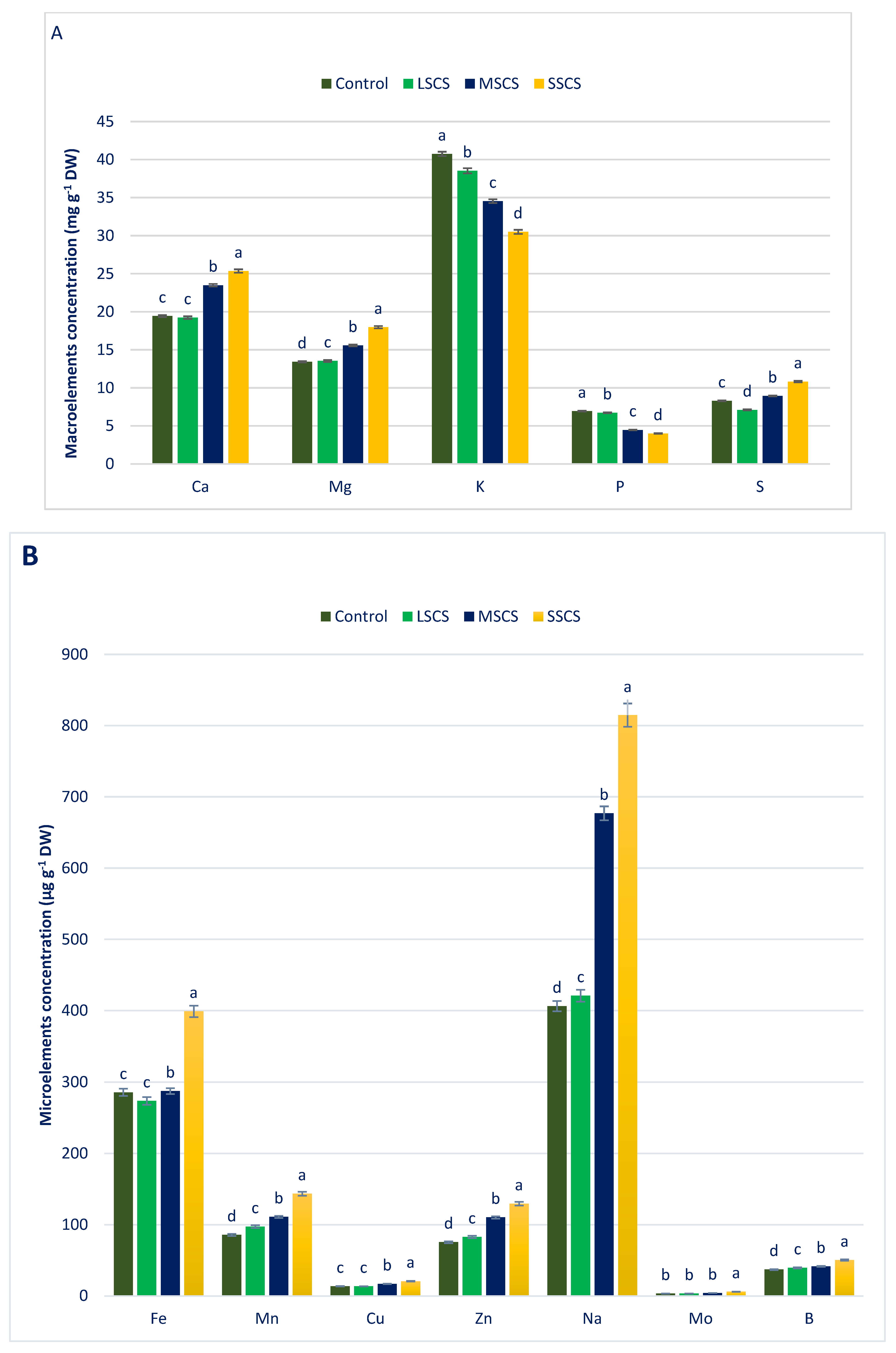
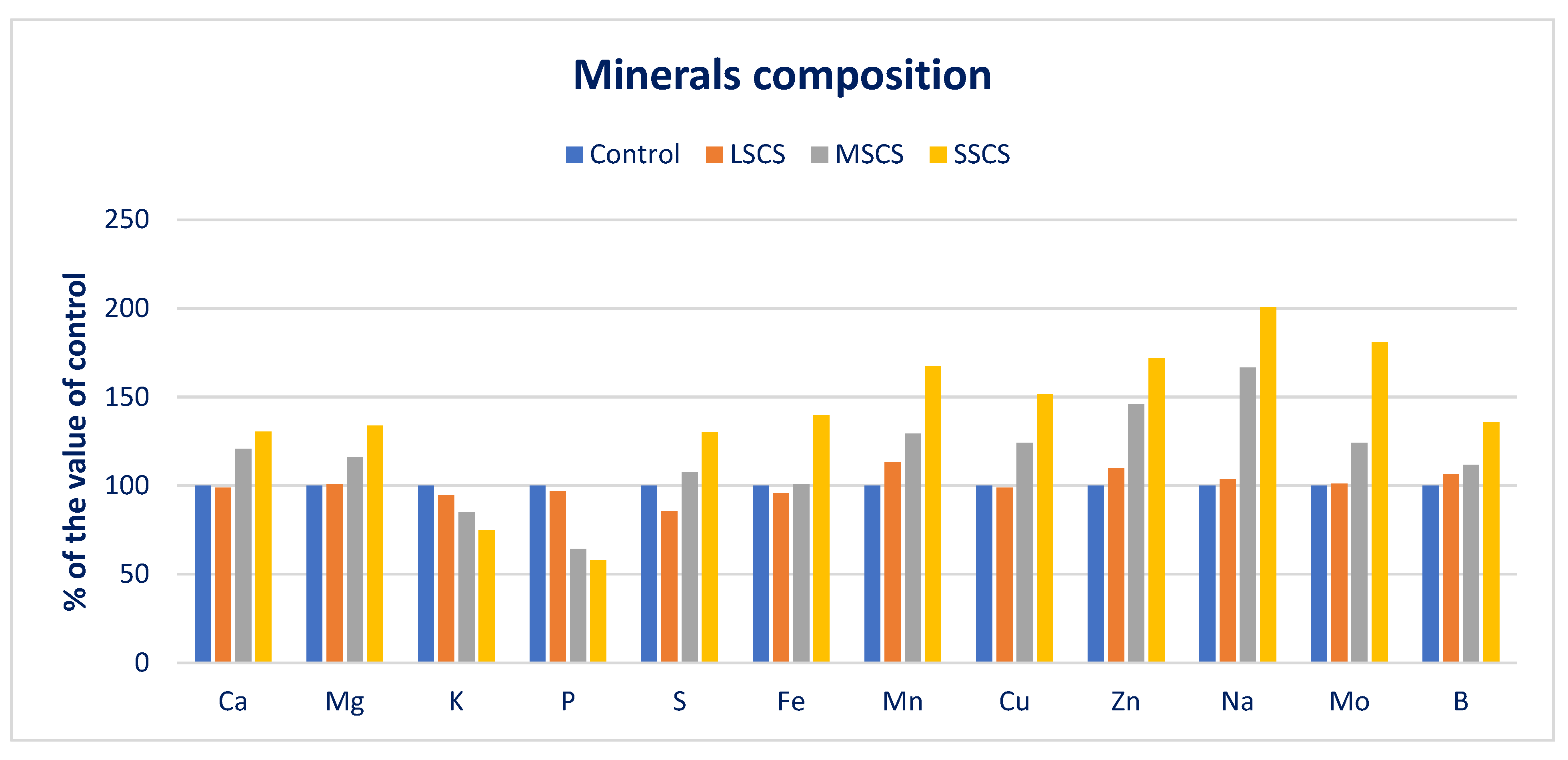
3.2. Sodium Chloride Impact on Minerals (Macroelements and Microelements) Composition
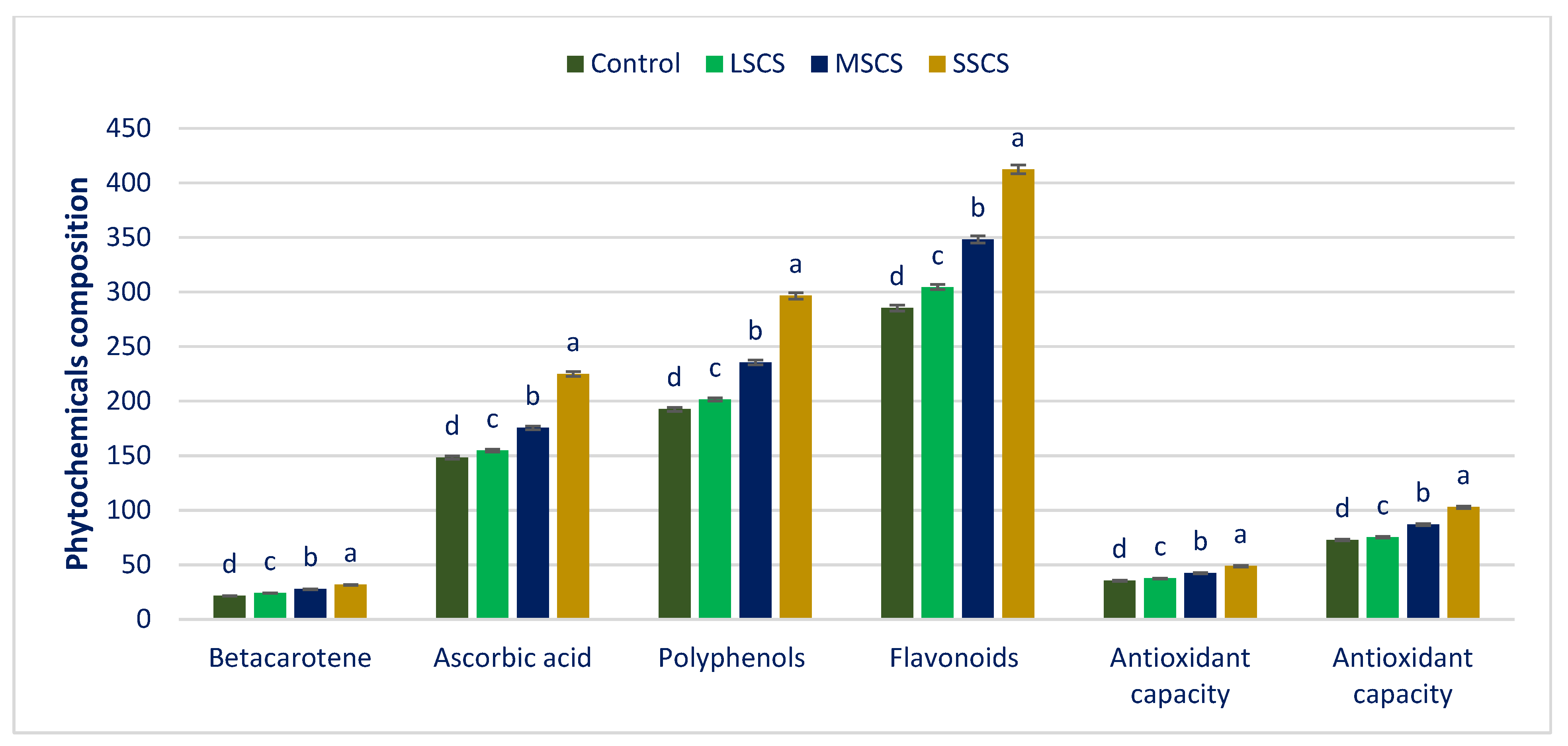
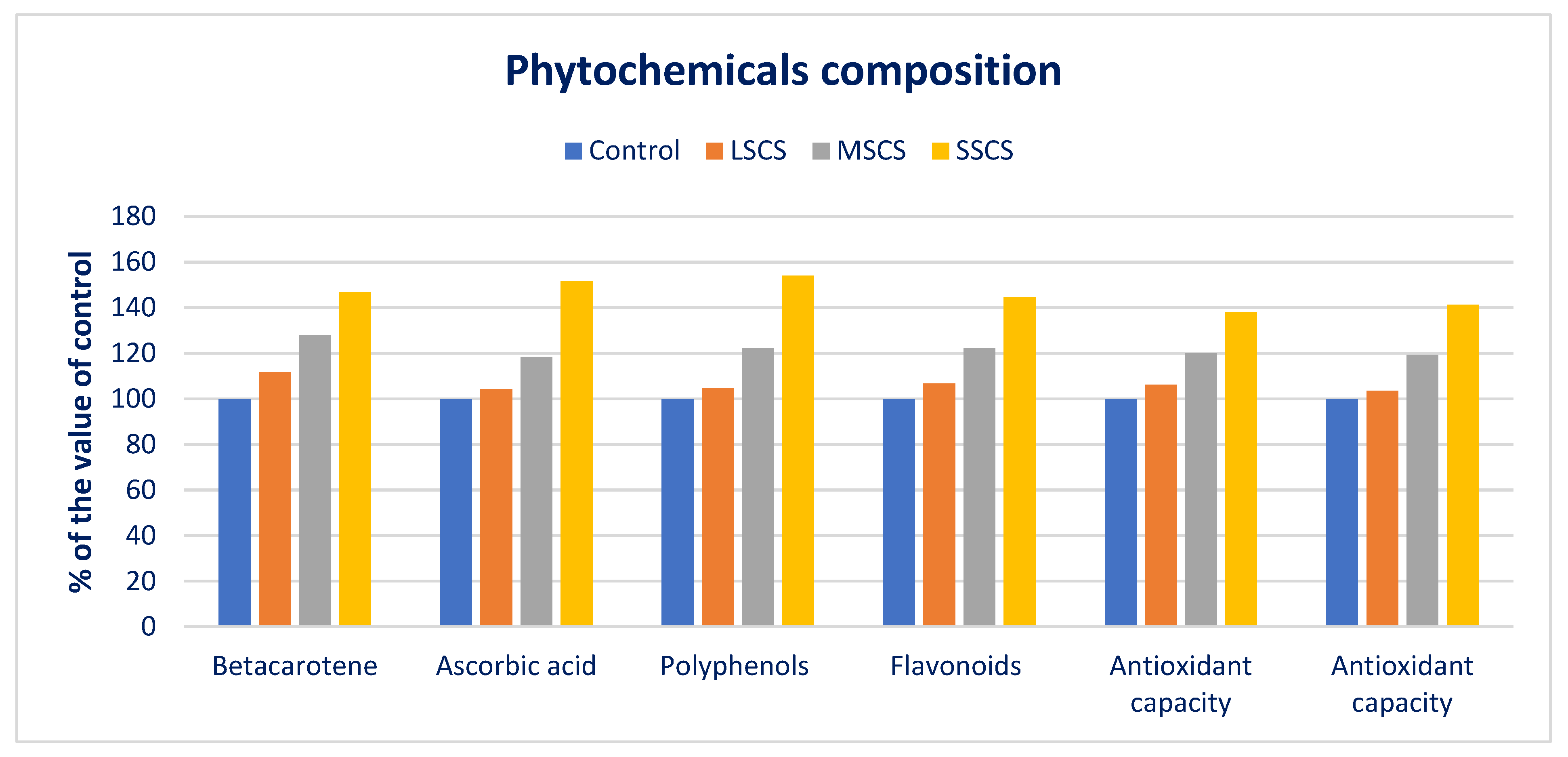
3.3. Impact of Salinity on Phytochemicals and ARP
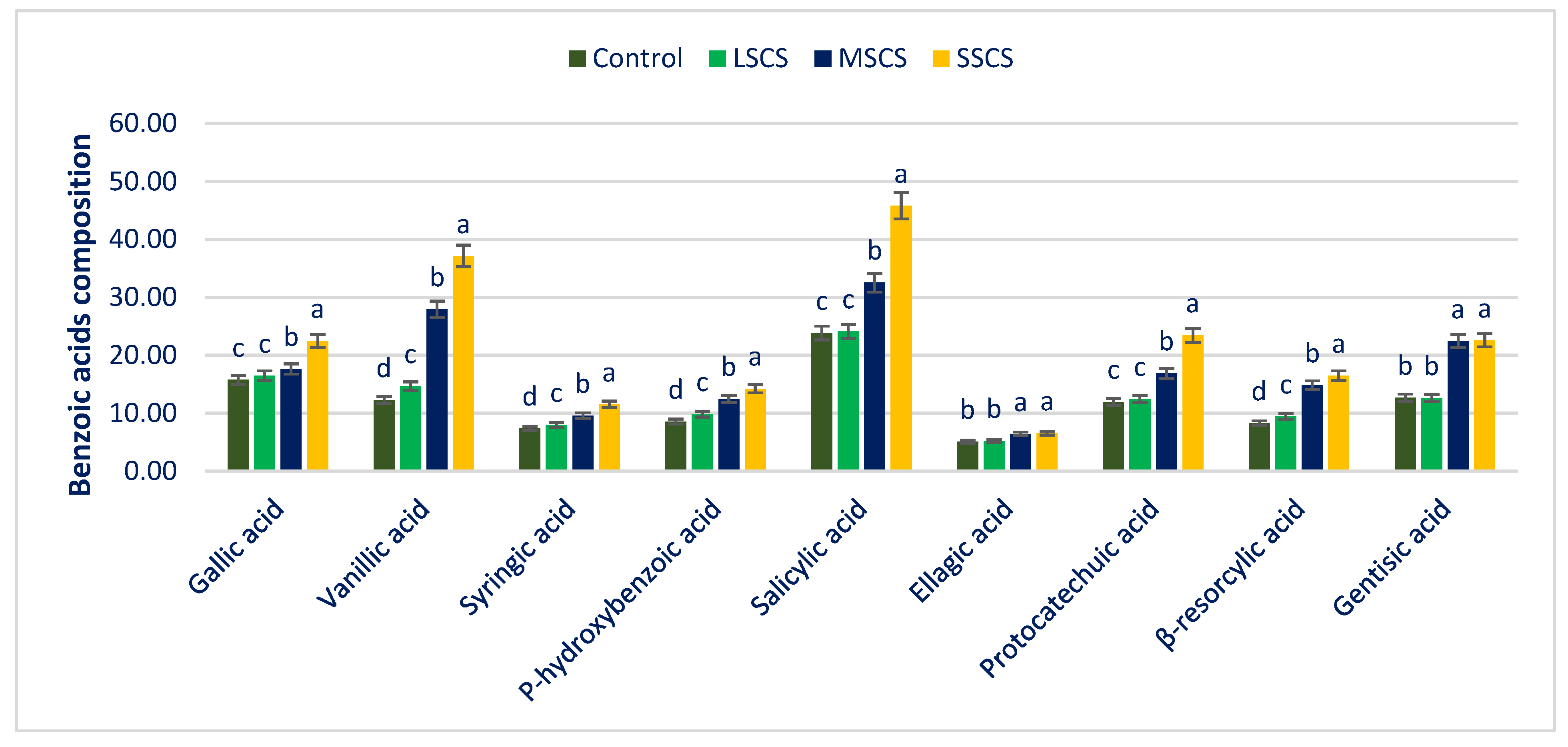
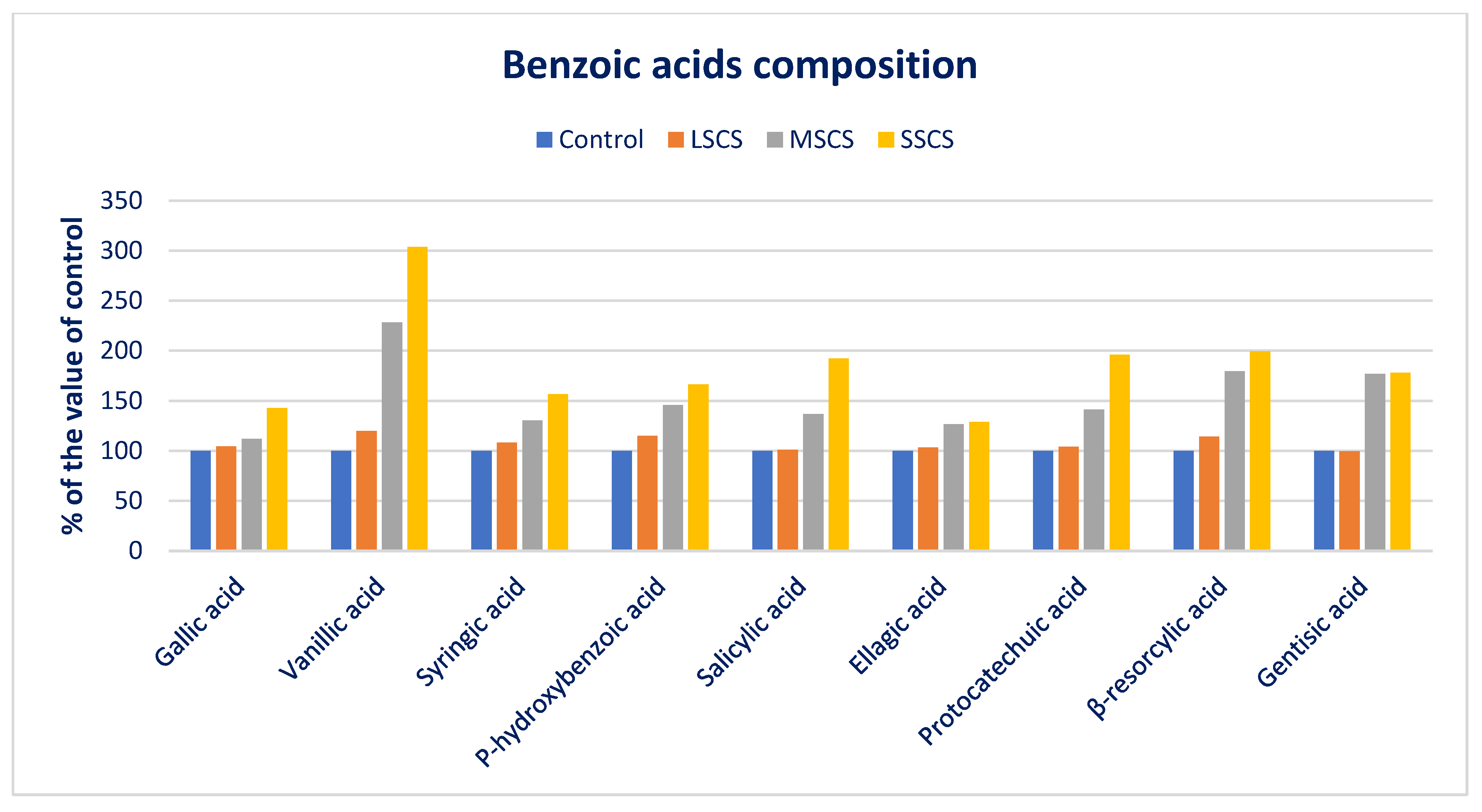
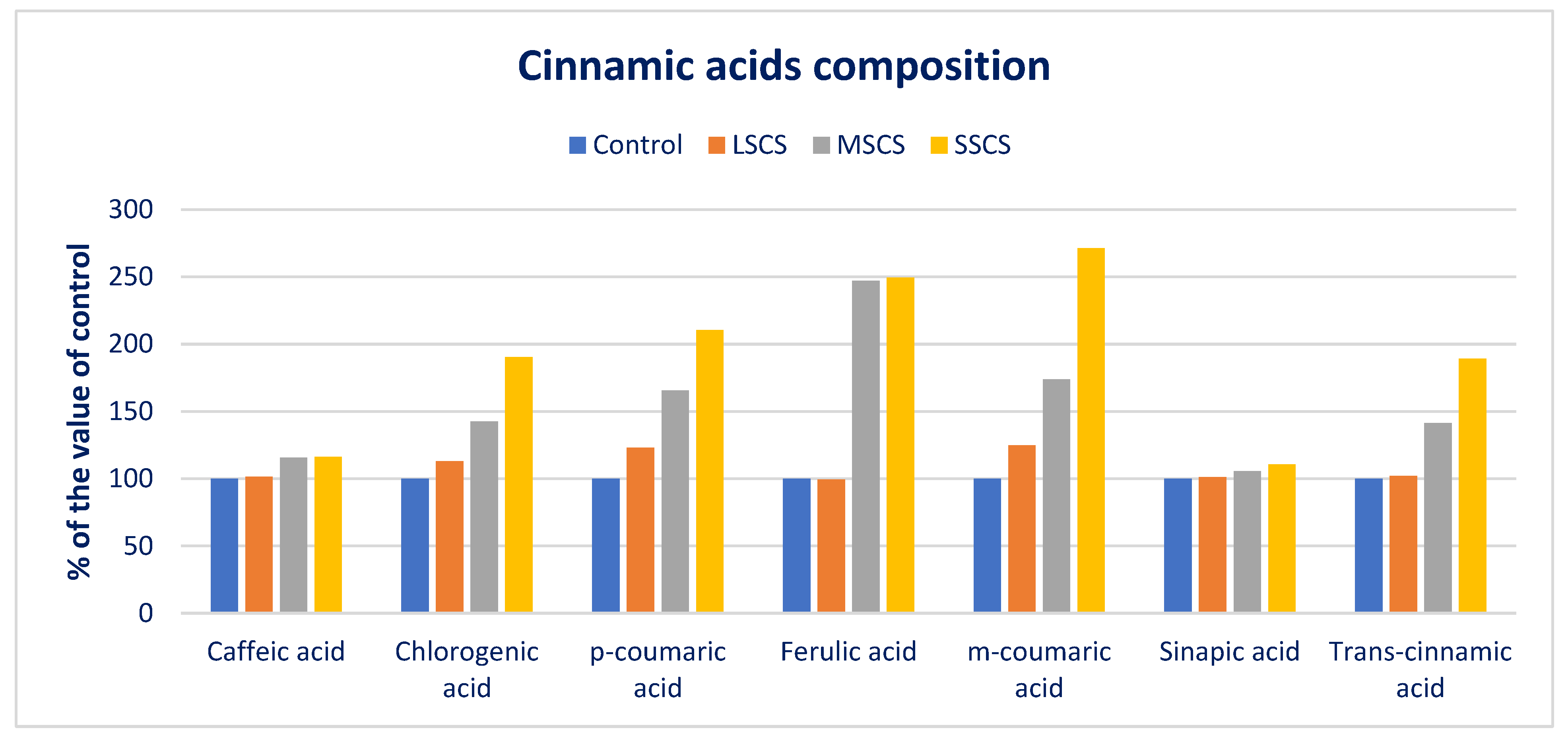
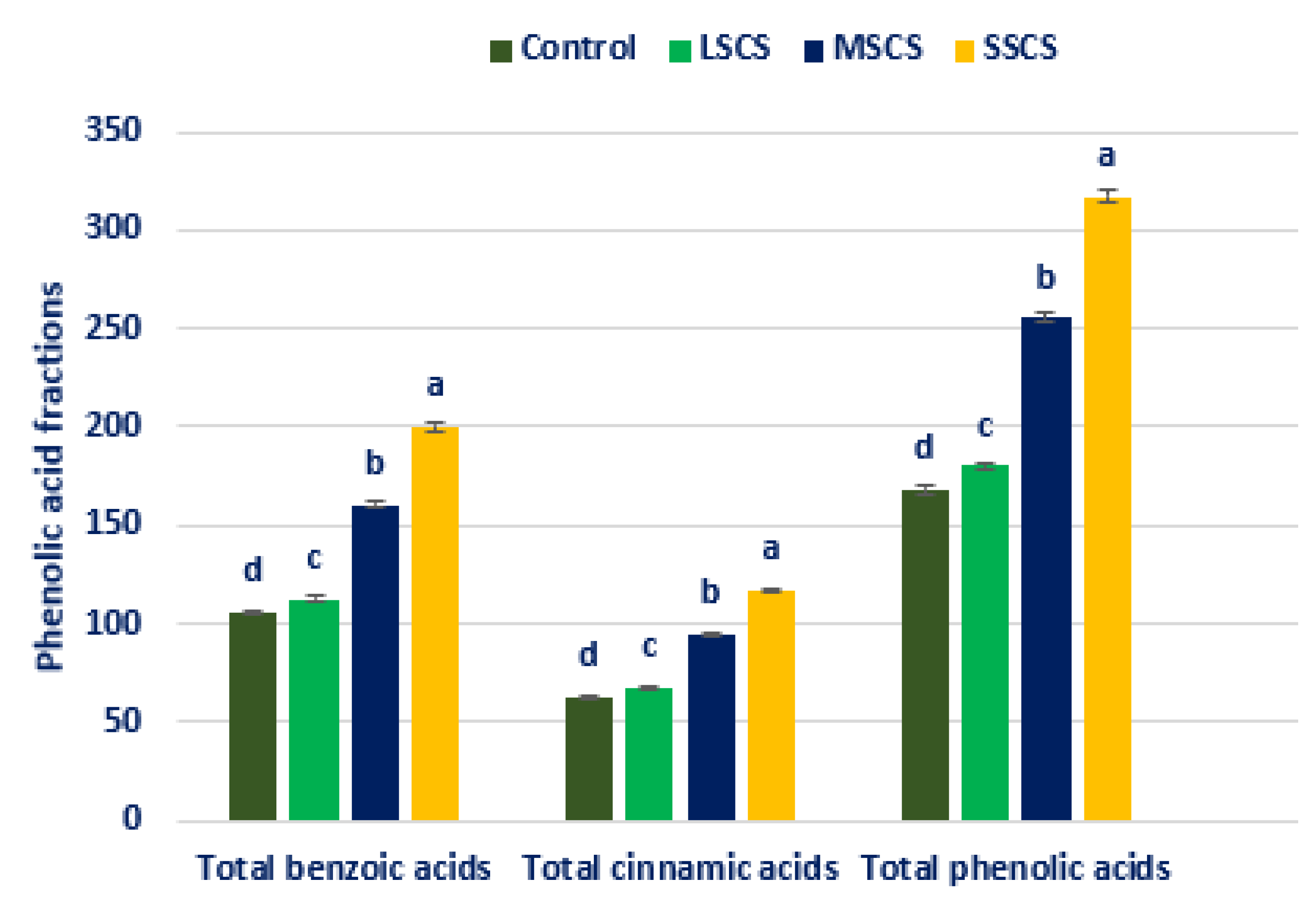
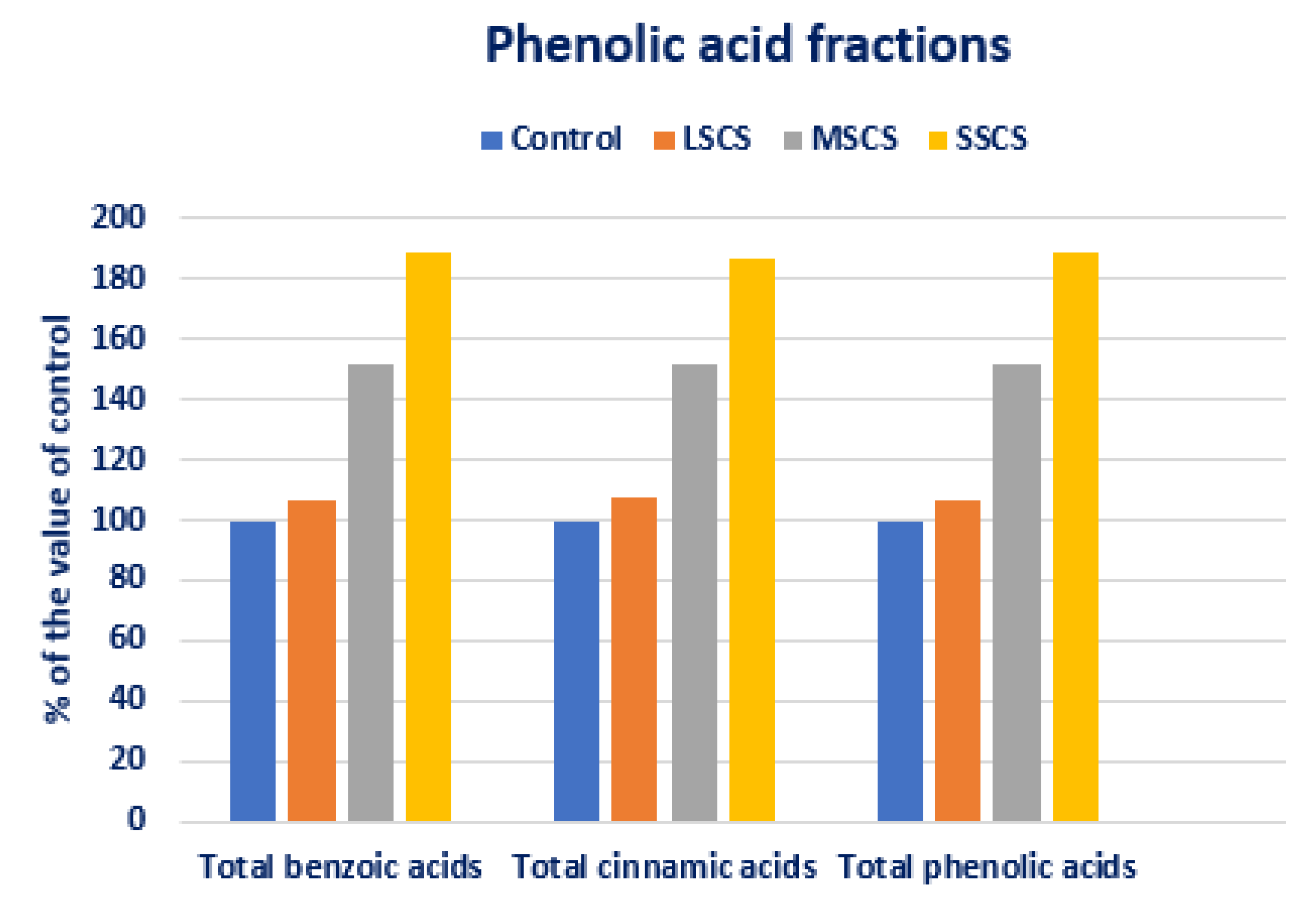
3.4. Response of Salinity on PA Profiles
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rastogi, A.; Shukla, S. Amaranth: A New Millennium Crop of Nutraceutical Values. Crit. Rev. Food Sci. Nutr. 2013, 53, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Promising Das, S. Amaranths: The Crop of Great Prospect. In Amaranthus: A Promising Crop of Future; Springer: Singapore, 2016; pp. 13–48. [Google Scholar]
- Sreelathakumary, I.; Peter, K.V. Amaranth: Amaranthus spp. In Genetic Improvement of Vegetable Crops; Elsevier: Amsterdam, The Netherlands, 1993; pp. 315–323. [Google Scholar]
- Sauer, J.D. The Grain Amaranths and Their Relatives: A Revised Taxonomic and Geographic Survey. Ann. Mo. Bot. Gard. 1967, 54, 103. [Google Scholar] [CrossRef]
- Anu, R.; Mishra, B.K.; Mrinalini, S.; Ameena, S.; Rawli, P.; Nidhi, V.; Sudhir, S. Identification of Heterotic Crosses Based on Combining Ability in Vegetable Amaranthus (Amaranthus tricolor L.). Asian J. Agric. Res. 2015, 9, 84–94. [Google Scholar]
- Nguyen, D.C.; Tran, D.S.; Tran, T.T.H.; Ohsawa, R.; Yoshioka, Y. Genetic diversity of leafy amaranth (Amaranthus tricolor L.) resources in Vietnam. Breed. Sci. 2019, 69, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Srivastava, A.; Singh, S.P. Genotypic variability in vegetable amaranth (Amaranthus tricolor L for foliage yield and its contributing traits over successive cuttings and years. Euphytica 2006, 151, 103–110. [Google Scholar] [CrossRef]
- Alvarez-Jubete, L.; Arendt, E.K.; Gallagher, E. Nutritive Value of Pseudocereals and Their Increasing Use as Functional Gluten-Free Ingredients. Trends Food Sci. Technol. 2010, 21, 106–113. [Google Scholar] [CrossRef]
- Achigan-Dako, E.G.; Sogbohossou, O.E.D.; Maundu, P. Current knowledge on Amaranthus spp.: Research avenues for improved nutritional value and yield in leafy amaranths in sub-Saharan Africa. Euphytica 2014, 197, 303–317. [Google Scholar] [CrossRef]
- Akin-Idowu, P.E.; Odunola, O.A.; Gbadegesin, M.A.; Ademoyegun, O.T.; Aduloju, A.O.; Olagunju, Y.O. Nutritional evaluation of Five Species of Grain Amaranth—An Underutilized Crop. Int. J. Sci. 2017, 3, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Alegbejo, J.O. Nutritional Value and Utilization of Amaranthus (Amaranthus spp.)—A Review. Bayero J. Pure Appl. Sci. 2014, 6, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Pandey, A.C.; Mishra, B.K. Diversity in Phenotypic and Nutritional Traits in Vegetable Amaranth (Amaranthus tricolor), A Nutritionally Underutilised Crop. J. Sci. Food Agric. 2010, 90, 139–144. [Google Scholar] [CrossRef]
- Soriano-García, M.; Arias-Olguín, I.I.; Montes, J.P.C.; Ramírez, D.G.R.; Figueroa, J.S.M.; Flores-Valverde, E.; Valladares-Rodríguez, M.R. Nutritional functional value and therapeutic utilization of Amaranth. J. Anal. Pharm. Res. 2018, 7, 596–600. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Protein, dietary fiber, minerals, antioxidant pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE 2019, 14, e0222517. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, T.; Rabbani, G.; Oba, S. Variability, heritability and genetic association in vegetable amaranth (Amaranthus tricolor L.). Span. J. Agric. Res. 2015, 13, e0702. [Google Scholar] [CrossRef] [Green Version]
- Andini, R.; Yoshida, S.; Ohsawa, R. Variation in Protein Content and Amino Acids in the Leaves of Grain, Vegetable and Weedy Types of Amaranths. Agronomy 2013, 3, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Manólio Soares, R.A.; Mendonça, S.; Andrade de Castro, L.I.; Cardoso Corrêa Carlos Menezes, A.C.; Gomes Arêas, J.A. Major Peptides from Amaranth (Amaranthus cruentus) Protein Inhibit HMG-CoA Reductase Activity. Int. J. Mol. Sci. 2015, 16, 4150. [Google Scholar] [CrossRef] [Green Version]
- Písaríková, B.; Krácmar, S.; Herzig, I. Amino acid Contents and Biological Value of Protein Amaranth. Czech J. Anim. Sci. 2005, 50, 169–174. [Google Scholar] [CrossRef] [Green Version]
- López, D.N.; Galante, M.; Raimundo, G.; Spelzini, D.; Boeris, V. Functional properties of amaranth, quinoa and chia proteins and the biological activities of their hydrolyzates. Food Res. Int. 2019, 116, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutraceuticals, phytochemicals, and radical quenching ability of selected drought-tolerant advance lines of vegetable amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Hossain, N.; Iqbal, A.; Oba, S. Bioactive Components and Radical Scavenging Activity in Selected Advance Lines of Salt-Tolerant Vegetable Amaranth. Front. Nutr. 2020, 7, 587257. [Google Scholar] [CrossRef]
- Cai, Y.; Corke, H. Amaranthus Betacyanin Pigments Applied in Model Food Systems. J. Food Sci. 1999, 64, 869–873. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. Identification and Distribution of Simple and Acylated Betacyanins in the Amaranthaceae. J. Agric. Food Chem. 2001, 49, 1971–1978. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. HPLC Characterization of Betalains from Plants in the Amaranthaceae. J. Chromatogr. Sci. 2005, 43, 454–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Sun, M.; Wu, H.; Huang, R.; Corke, H. Characterization and Quantification of Betacyanin Pigments from Diverse Amaranthus Species. J. Agric. Food Chem. 1998, 46, 2063–2070. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. Characterization and Application of Betalain Pigments from Plants of the Amaranthaceae. Trends Food Sci. Technol. 2005, 16, 370–376. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Abderrabba, M. Chemical and Antioxidant Properties of Betalains. J. Agric. Food Chem. 2017, 65, 675–689. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Motafakkerazad, R.; Nakajima, Y.; Matsugo, S.; Rimbach, G. Free radical scavenging and antioxidant activity of betanin: Electron spin resonance spectroscopy studies and studies in cultured cells. Food Chem. Toxicol. 2014, 73, 119–126. [Google Scholar] [CrossRef]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. The Role of Phenolic Hydroxy Groups in the Free Radical Scavenging Activity of Betalains. J. Nat. Prod. 2009, 72, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Structural Implications on Color, Fluorescence, and Antiradical Activity in Betalains. Planta 2010, 232, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Purification and Antiradical Properties of the Structural Unit of Betalains. J. Nat. Prod. 2012, 75, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Biological Activities of Plant Pigments Betalains. Crit. Rev. Food Sci. Nutr. 2016, 56, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I. Plant Betalains: Safety, Antioxidant Activity, Clinical Efficacy, and Bioavailability. Compr. Rev. Food Sci. Food Saf. 2016, 15, 316–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.; Giridhar, P. Plant betalains: Chemistry and biochemistry. Phytochemistry 2015, 117, 267–295. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Functional properties of anthocyanins and betalains in plants, food, and in human nutrition. Trends Food Sci. Technol. 2004, 15, 19–38. [Google Scholar] [CrossRef]
- Taira, J.; Tsuchida, E.; Katoh, M.C.; Uehara, M.; Ogi, T. Antioxidant capacity of betacyanins as radical scavengers for peroxyl radical and nitric oxide. Food Chem. 2015, 166, 531–536. [Google Scholar] [CrossRef]
- Sarker, U.; Lin, Y.-P.; Oba, S.; Yoshioka, Y.; Hoshikawa, K. Prospects and potentials of underutilized leafy Amaranths as vegetable use for health-promotion. Plant Physiol. Biochem. 2022, 182, 104–123. [Google Scholar] [CrossRef]
- Miguel, M.G. Betalains in Some Species of the Amaranthaceae Family: A Review. Antioxidants 2018, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Polyphenol and flavonoid profiles and radical scavenging activity in leafy vegetable Amaranthus gangeticus. BMC Plant Biol. 2020, 20, 499. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Leaf pigmentation, its profiles and radical scavenging activity in selected Amaranthus tricolor leafy vegetables. Sci. Rep. 2020, 10, 18617. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef]
- Pasko, P.; Bartón, H.; Zagrodzki, P.; Gorinstein, S.; Fołta, M.; Zachwieja, Z. Anthocyanins, Total Polyphenols and Antioxidant Activity in Amaranth and Quinoa Seeds and Sprouts During Their Growth. Food Chem. 2009, 115, 994–998. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Liu, R.; Zhu, H.; Draves, J.; Marcone, M.; Sun, Y.; Tsao, R. Characterization of phenolics, betacyanins and antioxidant activities of the seed, leaf, sprout, flower and stalk extracts of three Amaranthus species. J. Food Compos. Anal. 2015, 37, 75–81. [Google Scholar] [CrossRef]
- Barba de la Rosa, A.P.; Fomsgaard, I.S.; Laursen, B.; Mortensen, A.G.; Olvera-Martínez, L.; Silva-Sánchez, C.; Mendoza-Herrera, A.; González-Castañeda, J.; De León-Rodrígueza, A. Amaranth (Amaranthus hypochondriacus) as An Alternative Crop for Sustainable Food Production: Phenolic Acids and Flavonoids with Potential Impact on Its Nutraceutical Quality. J. Cereal Sci. 2009, 49, 117–121. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Meineri, G.; Gai, F.; Longato, E.; Amarowicz, R. Antioxidative activities and phenolic compounds of pumpkin (Cucurbita pepo) seeds and amaranth (Amaranthus caudatus) grain extracts. Nat. Prod. Res. 2017, 31, 2178–2182. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Kammerer, D.; Schieber, A.; Adama, H.; Nacoulma, O.G.; Carle, R. Betacyanins and Phenolic Com-pounds from Amaranthus spinosus L. and Boerhavia erecta L. Z. Naturforsch. C 2004, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kalinova, J.; Dadakova, E. Rutin and Total Quercetin Content in Amaranth (Amaranthus spp.). Plant Foods Hum. Nutr. 2009, 64, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Asao, M.; Watanabe, K. Functional and Bioactive Properties of Quinoa and Amaranth. Food Sci. Technol. Res. 2010, 16, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Xiao, Y.; Tang, Z.; Jin, W.; Wang, Y.; Chen, H.; Yao, H.; Shan, Z.; Bu, T.; Wang, X. Extraction of Polysaccha-rides from Amaranthus hybridus L. by Hot Water and Analysis of Their Antioxidant Activity. Peer J. 2019, 7, e7149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozsoy, N.; Yilmaz, T.; Kurt, O.; Can, A.; Yanardag, R. In vitro antioxidant activity of Amaranthus lividus L. Food Chem. 2009, 116, 867–872. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Sahinler, S.S.; Tepe, B. Astragalus gymnolobus, A. leporinus var. hirsutus, and A. onobrychis: Phyto-chemical Analysis and Biological Activity. Ind. Crop. Prod. 2020, 150, 112366. [Google Scholar] [CrossRef]
- Tatiya, A.U.; Surana, S.J.; Khope, S.D.; Gokhale, S.B.; Sutar, M.P. Phytochemical Investigation and Immunomodulatory Activity of Amaranthus spinosus L. Indian J. Pharm. Educ. Res. 2007, 444, 337–341. [Google Scholar]
- Pamela, E.A.I.; Olufemi, T.A.; Yemisi, O.O.; Aduloju, O.A.; Usifo, G.A. Phytochemical Content and Antioxidant Activity of Five Grain Amaranth Species. Am. J. Food Sci. Technol. 2017, 5, 249–255. [Google Scholar]
- Pasko, P.; Barton, H.; Fołta, M.; Gwizdz, J. Evaluation of Antioxidant Activity of Amaranth Amaranthus cruentus Grain and by-Products Flour, Popping, Cereal. Rocz. Pánstwowego Zakładu Hig. 2007, 581, 35–40. [Google Scholar]
- Nsimba, R.Y.; Kikuzaki, H.; Konishi, Y. Antioxidant Activity of Various Extracts and Fractions of Chenopodium quinoa and Amaranthus spp. Seeds. Food Chem. 2008, 106, 760–766. [Google Scholar] [CrossRef]
- Tang, Y.; Tsao, R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory, and potential health beneficial effects: A review. Mol. Nutr. Food Res. 2017, 61, 1600767. [Google Scholar] [CrossRef] [PubMed]
- Barku, V.Y.A.; Opoku-Boahen, Y.; Owusu-Ansah, E.; Mensah, E.F.; Barku, V.Y.A.; Opoku-Boahen, Y.; Owusu-Ansah, E.; Mensah, E.F. Antioxidant Activity and The Estimation of Total Phenolic and Flavonoid Contents of The Root Extract of Amaranthus spinosus. Asian J. Plant Sci. Res. 2013, 3, 69–74. [Google Scholar]
- Bulbul, I.J.; Nahar, L.; Ripa, F.A.; Haque, O. Antibacterial, Cytotoxic and Antioxidant Activity of Chloroform, N-Hexane and Ethyl Acetate Extract of Plant Amaranthus spinosus. Int. J. PharmTech Res. 2011, 33, 1675–1680. [Google Scholar]
- Kumar, B.S.A.; Lakshman, K.; Jayaveera, K.; Shekar, D.S.; Kumar, A.A.; Manoj, B. Antioxidant and antipyretic properties of methanolic extract of Amaranthus spinosus leaves. Asian Pac. J. Trop. Med. 2010, 3, 702–706. [Google Scholar] [CrossRef] [Green Version]
- Ishtiaq, S.; Ahmad, M.; Hanif, U.; Akbar, S.; Kamran, S.H. Phytochemical and in-vitro Antioxidant Evaluation of Dif-ferent Fractions of Amaranthus graecizan subsp. Silvestris Vill. Brenan. Asian Pac. J. Trop. Biomed. 2014, 412, 965–971. [Google Scholar]
- Karamać, M.; Gai, F.; Longato, E.; Meineri, G.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Antioxidant Activity and Phenolic Composition of Amaranth (Amaranthus caudatus) during Plant Growth. Antioxidants 2019, 8, 173. [Google Scholar] [CrossRef]
- Kraujalis, P.; Venskutonis, P.R.; Kraujaliene, V.; Pukalskas, A. Antioxidant properties and preliminary evaluation of phytochemical composition of different anatomical parts of amaranth. Plant Foods Hum. Nutr. 2013, 68, 322–328. [Google Scholar] [CrossRef]
- Kumari, S.; Elancheran, R.; Devi, R. Phytochemical Screening, Antioxidant, Antityrosinase, And Antigenotoxic Potential of Amaranthus viridis Extract. Indian J. Pharmacol. 2018, 50, 130–138. [Google Scholar]
- Lopez-Mejía, O.A.; Lopez-Malo, A.; Palou, E. Antioxidant Capacity of Extracts from Amaranth Amaranthus hypochondriacus L. Seeds or Leaves. Ind. Crop. Prod. 2014, 53, 55–59. [Google Scholar] [CrossRef]
- Lucero-Lopez, V.R.; Razzeto, G.S.; Gimenez, M.S.; Escudero, N.L. Antioxidant Properties of Amaranthus hypochondri-acus Seeds and Their Effect on The Liver of Alcohol-Treated Rats. Plant Foods Hum. Nutr. 2011, 66, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Salvamani, S.; Gunasekaran, B.; Shukor, M.Y.; Shaharuddin, N.A.; Sabullah, M.K.; Ahmad, S.A. Anti-HMG-CoA Re-ductase, Antioxidant, and Anti-Inflammatory Activities of Amaranthus viridis Leaf Extract as A Potential Treatment for Hypercholesterolemia. Evid. Based Complement. Altern. Med. 2016, 2016, 8090841. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Sicairos, E.S.; Milán-Noris, A.K.; Luna-Vital, D.A.; Milán-Carrillo, J.; Montoya-Rodríguez, A. Anti-Inflammatory and Antioxidant Effects of Peptides Released from Germinated Amaranth During In Vitro Simulated Gastrointestinal Digestion. Food Chem. 2021, 1, 128394. [Google Scholar] [CrossRef] [PubMed]
- Medoua, G.N.; Oldewage-Theron, W.H. Effect of Drying and Cooking on Nutritional Value and Antioxidant Capac-ity of Morogo (Amaranthus hybridus) A Traditional Leafy Vegetable Grown in South Africa. J. Food Sci. Technol. 2014, 51, 736–742. [Google Scholar] [CrossRef] [Green Version]
- Tesoriere, L.; Allegra, M.; Gentile, C.; Livrea, M.A. Betacyanins as phenol antioxidants. Chemistry and mechanistic aspects of the lipoperoxyl radical-scavenging activity in solution and liposomes. Free Radic. Res. 2009, 43, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco-Valencia, R.; Peña, J.; Kallio, H.; Salminen, S. Dietary fiber and other functional components in two varieties of crude and extruded kiwicha (Amaranthus caudatus). J. Cereal Sci. 2009, 49, 219–224. [Google Scholar] [CrossRef]
- Jo, H.-J.; Chung, K.-H.; Yoon, J.A.; Lee, K.-J.; Song, B.C.; An, J.H. Radical Scavenging Activities of Tannin Extracted from Amaranth (Amaranthus caudatus L.). J. Microbiol. Biotechnol. 2015, 25, 795–802. [Google Scholar] [CrossRef]
- Subhasree, B.; Baskar, R.; Keerthana, R.L.; Susan, R.L.; Rajasekaran, P. Evaluation of antioxidant potential in selected green leafy vegetables. Food Chem. 2009, 115, 1213–1220. [Google Scholar] [CrossRef]
- Lacatusu, I.; Arsenie, K.L.V.; Badea, G.; Popa, O.; Oprea, O.; Badea, N. New Cosmetic Formulations with Broad Pho-toprotective and Antioxidative Activities Designed by Amaranth and Pumpkin Seed Oils Nanocarriers. Ind. Crops Prod. 2018, 123, 424–433. [Google Scholar] [CrossRef]
- Steffensen, S.K.; Pedersen, H.A.; Labouriau, R.; Mortensen, A.G.; Laursen, B.; de Troiani, R.M.; Noellemeyer, E.J.; Janovska, D.; Stavelikova, H.; Taberner, A.; et al. Variation of Polyphenols and Betaines in Aerial Parts of Young, Field-Grown Amaranthus Genotypes. J. Agric. Food Chem. 2011, 59, 12073–12082. [Google Scholar] [CrossRef]
- Niveyro, S.L.; Mortensen, A.G.; Fomsgaard, I.S.; Salvo, A. Differences among five amaranth varieties (Amaranthus spp.) regarding secondary metabolites and foliar herbivory by chewing insects in the field. Arthropod-Plant Interact. 2013, 7, 235–245. [Google Scholar] [CrossRef]
- Bao, X.; Han, X.; Du, G.; Wei, C.; Zhu, X.; Ren, W.; Zeng, L.; Zhang, Y. Antioxidant Activities and Immunomodulatory Effects in Mice of Betalain in vivo. Food Sci. 2019, 40, 196–201. [Google Scholar]
- Venskutonis, P.R.; Kraujalis, P. Nutritional Components of Amaranth Seeds and Vegetables: A Review on Composition, Properties, and Uses. Comp. Rev. Food Sci. Food Saf. 2013, 12, 381–412. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco-Valencia, R.; Hellstrom, J.K.; Philava, J.M.; Mattila, P.H. Flavonoids and Other Phenolic Compounds in Andean Indigenous Grains: Quinoa (Chenopodium quinoa), Kaniwa (Chenopodium pallidicaule) and Kiwicha (Amaranthus caudatus). Food Chem. 2010, 120, 128–133. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Schiniatbeyo, V.B.; Rimbach, G. Betanin-A food colorant with biological activity. Mol. Nutr. Food Res. 2015, 59, 36–47. [Google Scholar] [CrossRef]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.-P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common fruits in Singapore. Food Chem. 2010, 123, 77–84. [Google Scholar] [CrossRef]
- Wahid, A.; Ghazanfar, A. Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J. Plant Physiol. 2006, 163, 723–730. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Effects on Growth, ROS Markers, Compatible Solutes, Phenolics, Flavonoids, and Antioxidant Activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. Antioxidant Activity of Betalains from Plants of the Amaranthaceae. J. Agric. Food Chem. 2003, 51, 2288–2294. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Betalains—Emerging prospects for food scientists. Trends Food Sci. Technol. 2007, 18, 514–525. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, T.; Oba, S. Salinity stress accelerates nutrients, dietary fiber, minerals, phytochemicals and antioxidant activity in Amaranthus tricolor leaves. PLoS ONE 2018, 13, e0206388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crop. Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I.C. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.H.; Park, K.-J.; Kim, B.-K.; Jeong, J.-W.; Kim, H.-J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef]
- Alam, A.; Juraimi, A.S.; Rafii, M.Y.; Hamid, A.A.; Aslani, F.; Alam, M.Z. Effects of salinity and salinity-induced augmented bioactive compounds in purslane (Portulaca oleracea L.) for possible economical use. Food Chem. 2015, 169, 439–447. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Cao, F.; Han, Y.; Nadira, U.A.; Zhang, G.; Wu, F. Differential changes in grain ultrastructure, amylase, protein and amino acid profiles between Tibetan wild and cultivated barleys under drought and salinity alone and combined stress. Food Chem. 2013, 141, 2743–2750. [Google Scholar] [CrossRef]
- Hossain, N.; Sarker, U.; Raihan, S.; Al-Huqail, A.A.; Siddiqui, M.H.; Oba, S. Influence of Salinity Stress on Color Parameters, Leaf Pigmentation, Polyphenol and Flavonoid Contents, and Antioxidant Activity of Amaranthus lividus Leafy Vegetables. Molecules 2022, 27, 1821. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Rabbani, G.; Oba, S.; Eldehna, W.M.; Al-Rashood, S.T.; Mostafa, N.M.; Eldahshan, O.A. Phytonutrients, Colorant Pigments, Phytochemicals, and Antioxidant Potential of Orphan Leafy Amaranthus Species. Molecules 2022, 27, 2899. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Dola, D.B.; Mannan, A.; Sarker, U.; Al Mamun, A.; Islam, T.; Ercisli, S.; Saleem, M.H.; Ali, B.; Pop, O.L.; Marc, R.A. Nano-iron oxide accelerates growth, yield, and quality of Glycine max seed in water deficits. Front. Plant Sci. 2022, 13, 992535. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, A.; Mannan, M.A.; Sarker, U.; Dola, D.B.; Higuchi, H.; Ercisli, S.; Ali, B.; Saleem, M.H.; Babalola, O.O. Foliar application of seaweed extracts enhances yield and drought tolerance of soybean. Front. Plant Sci. 2022, 13, 992880. [Google Scholar] [CrossRef]
- Sarker, U.; Azam, M.; Alam Talukder, Z. Genetic variation in mineral profiles, yield contributing agronomic traits, and foliage yield of stem amaranth. Genetika 2022, 54, 91–108. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Ercisli, S.; Assouguem, A.; Alotaibi, A.; Ullah, R. Bioactive Phytochemicals and Quenching Activity of Radicals in Selected Drought-Resistant Amaranthus tricolor Vegetable Amaranth. Antioxidants 2022, 11, 578. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Alsanie, W.F.; Gaber, A. Characterization of Phytochemicals, Nutrients, and Antiradical Potential in Slim Amaranth. Antioxidants 2022, 11, 1089. [Google Scholar] [CrossRef]
- Sarker, U.; Iqbal, A.; Hossain, N.; Oba, S.; Ercisli, S.; Muresan, C.C.; Marc, R.A. Colorant Pigments, Nutrients, Bioactive Components, and Antiradical Potential of Danta Leaves (Amaranthus lividus). Antioxidants 2022, 11, 1206. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Faysal, A.S.M.; Ali, L.; Azam, G.; Sarker, U.; Ercisli, S.; Golokhvast, K.S.; Marc, R.A. Genetic Variability, Character Association, and Path Coefficient Analysis in Transplant Aman Rice Genotypes. Plants 2022, 11, 2952. [Google Scholar] [CrossRef] [PubMed]
- Hassan, J.; Jahan, F.; Rajib, M.R.; Sarker, U.; Miyajima, I.; Ozaki, Y.; Ercisli, S.; Golokhvast, K.S.; Marc, R.A. Color and physiochemical attributes of pointed gourd (Trichosanthes dioica Roxb.) influenced by modified atmosphere packaging and postharvest treatment during storage. Front. Plant Sci. 2022, 13, 1016324. [Google Scholar] [CrossRef] [PubMed]
- Kulsum, U.; Sarker, U.; Rasul, G. Genetic variability, heritability and interrelationship in salt-tolerant lines of T. Aman rice. Genetika 2022, 54, 761–776. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Sarker, U.; Matin, M.Q.I.; Shahin, N.H.; Kabir, M.S.; Ercisli, S.; Marc, R.A. Assessment of GGE, AMMI, Regression, and Its Deviation Model to Identify Stable Rice Hybrids in Bangladesh. Plants 2022, 11, 2336. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Sarker, U.; Swapan, A.H.; Raihan, M.S.; Oba, S.; Alamri, S.; Siddiqui, M.H. Combining Ability Analysis and Marker-Based Prediction of Heterosis in Yield Reveal Prominent Heterotic Combinations from Diallel Population of Rice. Agronomy 2022, 12, 1797. [Google Scholar] [CrossRef]
- Azam, G.; Sarker, U.; Hossain, A.; Iqbal, S.; Islam, R.; Hossain, F.; Ercisli, S.; Kul, R.; Assouguem, A.; Al-Huqail, A.A.; et al. Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids. Plants 2022, 11, 1774. [Google Scholar] [CrossRef]
- Hassan, J.; Rajib, M.R.; Sarker, U.; Akter, M.; Khan, N.-E.; Khandaker, S.; Khalid, F.; Rahman, G.K.M.M.; Ercisli, S.; Muresan, C.C.; et al. Optimizing textile dyeing wastewater for tomato irrigation through physiochemical, plant nutrient uses and pollution load index of irrigated soil. Sci. Rep. 2022, 12, 10088. [Google Scholar] [CrossRef]
- Prodhan, M.; Sarker, U.; Hoque, A.; Biswas, S.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almutairi, M.H.; Mohamed, H.R.H.; Najda, A. Foliar Application of GA3 Stimulates Seed Production in Cauliflower. Agronomy 2022, 12, 1394. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R.; Sayed, A.A.; Peluso, I. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Majumder, R.R.; Sarker, U. Genotypic Variability for Grain Quality Attributes in Restorer Lines of Hybrid Rice. Genetika 2020, 52, 973–989 doi 102298/GENSR2003973H. [Google Scholar] [CrossRef]
- Hasan-Ud-Daula, M.; Sarker, U. Variability, Heritability, Character Association, and Path Coefficient Analysis in Advanced Breeding Lines of Rice (Oryza sativa L.). Genetika 2020, 52, 711–726. [Google Scholar] [CrossRef]
- Rashad, M.M.I.; Sarker, U. Genetic Variations in Yield and Yield Contributing Traits of Green Amaranth. Genetika 2020, 52, 393–407. [Google Scholar] [CrossRef]
- Sun, H.; Mu, T.; Xi, L.; Zhang, M.; Chen, J. Sweet potato (Ipomoea batatas L.) leaves as nutritional and functional foods. Food Chem. 2014, 156, 380–389. [Google Scholar] [CrossRef]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Srivastava, J.; Singh, N.; Singh, S.P. Mineral profile and variability in vegetable amaranth (Amaranthus tricolor). Plant Food. Hum. Nutri. 2006, 61, 23–28. [Google Scholar] [CrossRef]
- Madruga, M.S.; Camara, F.S. The chemical composition of “Multimistura” as a food supplement. Food Chem. 2000, 68, 41–44. [Google Scholar] [CrossRef]
- Shahidi, F.; Chavan, U.; Bal, A.; McKenzie, D. Chemical composition of beach pea (Lathyrus maritimus L.) plant parts. Food Chem. 1999, 64, 39–44. [Google Scholar] [CrossRef]
- Jiménez-Aguilar, D.M.; Grusak, M.A. Minerals, vitamin C, phenolics, flavonoids and antioxidant activity of Amaranthus leafy vegetables. J. Food Compos. Anal. 2017, 58, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Khanam, U.K.S.; Oba, S.; Yanase, E.; Murakami, Y. Phenolic acids, flavonoids and total antioxidant capacity of selected leafy vegetables. J. Funct. Foods. 2012, 4, 979–987. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic Acids in Foods: An Overview of Analytical Methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and disease: A mechanistic review. Iran J. Basic Med. Sci 2019, 22, 225–237. [Google Scholar] [CrossRef]
- Kaur, J.; Gulati, M.; Singh, S.K.; Kuppusamy, G.; Kapoor, B.; Mishra, V.; Gupta, S.; Arshad, M.F.; Porwal, O.; Jha, N.K.; et al. Discovering multifaceted role of vanillic acid beyond flavours: Nutraceutical and therapeutic potential. Trends Food Sci. Technol. 2022, 122, 187–200. [Google Scholar] [CrossRef]
- Cheemanapalli, S.; Mopuri, R.; Golla, R.; Anurudha, C.M.; Chitta, S.K. Syringic acid (SA)—A review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxidative Med. Cell. Longev. 2022, 2022, 3848084. [Google Scholar] [CrossRef] [PubMed]
- Semaming, Y.; Pannengpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Pharmacological Properties of Protocatechuic Acid and Its Potential Roles as Complementary Medicine. Evid. Based Complement. Altern. Med. 2015, 11, 593902. [Google Scholar] [CrossRef] [Green Version]
- Wagle, B.R.; Upadhyay, A.; Arsi, K.; Shrestha, S.; Venkitanarayanan, K.; Donoghue, A.M.; Donoghue, D.J. Application of β-Resorcylic Acid as Potential Antimicrobial Feed Additive to Reduce Campylobacter Colonization in Broiler Chickens. Front. Microbiol. 2017, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Abedi, F.; Razavi, B.M.; Hosseinzadeh, H. A review on gentisic acid as a plant derived phenolic acid and me-tabolite of aspirin: Comprehensive pharmacology, toxicology, and some pharmaceutical aspects. Phytother. Res. 2020, 34, 729–734. [Google Scholar] [CrossRef]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Rosario, A.C.R.S.; Da Silva, A.H.M.; Silva, A.G.B.; Vieira, A.P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Boz, H. p-Coumaric acid in cereals: Presence, antioxidant and antimicrobial effects. Int. J. Food Sci. Technol. 2015, 50, 2323–2328. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Nićiforović, N.; Abramovič, H. Sinapic Acid and Its Derivatives: Natural Sources and Bioactivity. Compr. Rev. Food Sci. Food Saf. 2014, 13, 34–51. [Google Scholar] [CrossRef] [PubMed]
- Pontiki, E.; Hadjipavlou-Litina, D.; Litinas, K.; Geromichalos, G. Novel Cinnamic Acid Derivatives as Antioxidant and Anticancer Agents: Design, Synthesis and Modeling Studies. Molecules 2014, 19, 9655–9674. [Google Scholar] [CrossRef] [PubMed]
- Klados, E.; Tzortzakis, N. Effects of Substrate and Salinity in Hydroponically Grown Cichorium spinosum. J. Soil Sci. Plant Nutr. 2014, 14, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Neffati, M.; Sriti, J.; Hamdaoui, G.; Kchouk, M.E.; Marzouk, B. Salinity Impact on Fruit Yield, Essential Oil Composition and Antioxidant Activities of Coriandrum sativum Fruit Extracts. Food Chem. 2011, 124, 221–225. [Google Scholar] [CrossRef]
- Omamt, E.N.; Hammes, P.S.; Robbertse, P.J. Differences in Salinity Tolerance for Growth and Water-Use Efficiency in Some Amaranth (Amaranthus spp.) genotypes. New Zealand J. Crop Hortic. Sci. 2006, 34, 11–22. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
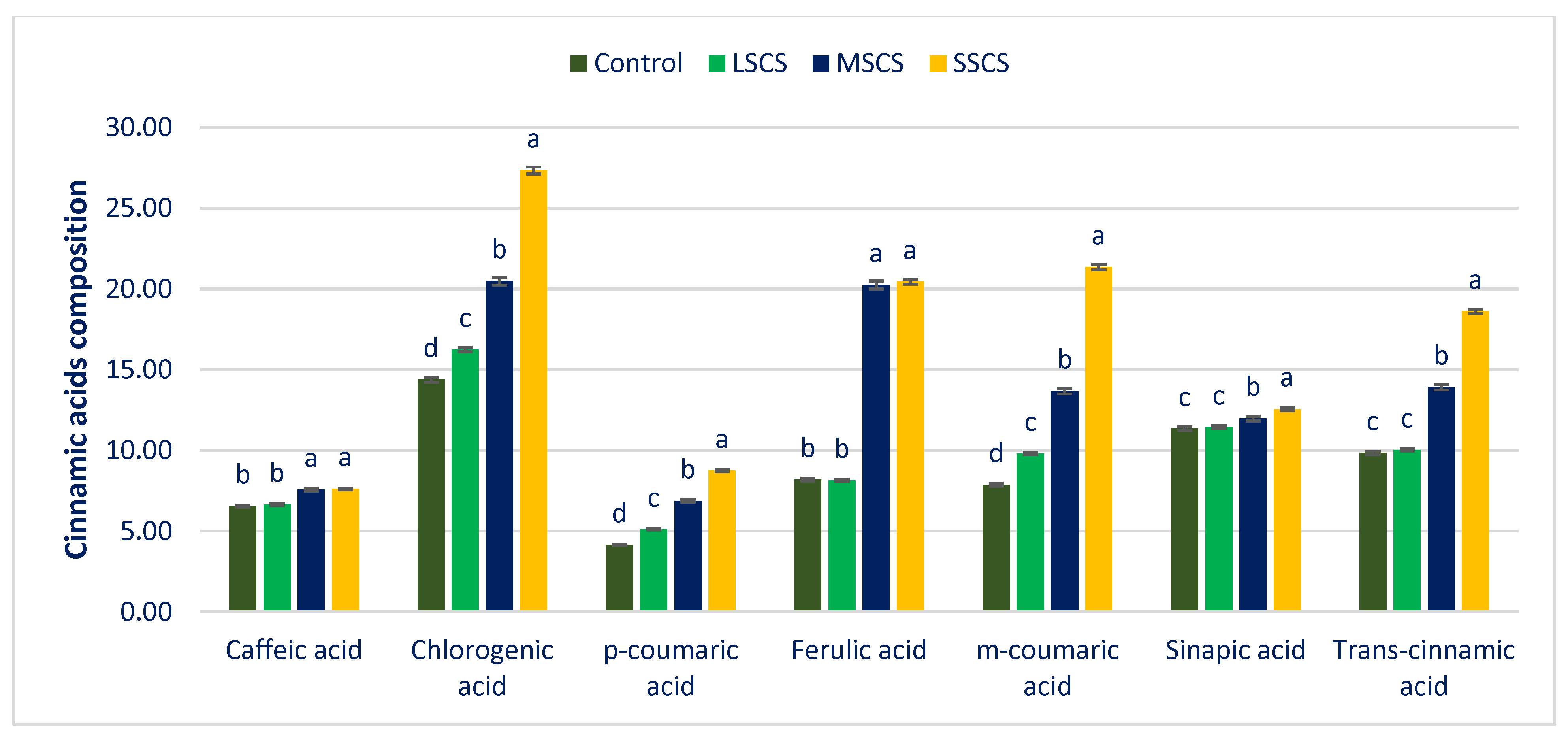{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No | Rt (min) | λmax (nm) | Molecular Ion [M-H]− (m/z) | MS2 (m/z) | Identity of Tentative Phenolic Acids |
|---|---|---|---|---|---|
| 1 | 9.1 | 254 | 169.1142 | 169.1563 | 3,4,5 Trihydroxybenzoic acid |
| 2 | 30.6 | 254 | 167.1214 | 167.1564 | 4-Hydroxy-3-methoxybenzoic acid |
| 3 | 34.8 | 254 | 197.1132 | 197.1104 | 3,5-Dimethoxy-4-hydroxybenzoic acid |
| 4 | 31.5 | 254 | 137.0213 | 137.1574 | 4-Hydroxybenzoic acid |
| 5 | 48.2 | 254 | 137.2113 | 137.1582 | 2-Hydroxybenzoic acid |
| 6 | 52.5 | 254 | 301.0423 | 301.0643 | 2,3,7,8-Tetrahydroxy-chromeno [5,4,3-cde] chromene-5,10-dione |
| 7 | 2.2 | 280 | 154.1212 | 154.1157 | 3,4-Dihydroxybenzoic acid |
| 8 | 4.0 | 280 | 154.1212 | 154.0156 | 2,4-Dihydroxybenzoic acid |
| 9 | 3.7 | 280 | 154.1212 | 154.1157 | 2,5- Dihydroxybenzoic acid |
| 10 | 32.0 | 280 | 179.0821 | 179.0687 | 3,4-Dihydroxy-trans-cinnamate |
| 11 | 31.1 | 280 | 353.1253 | 353.1542 | 3-(3,4-Dihydroxy cinnamoyl) quinic acid |
| 12 | 42.0 | 280 | 163.0658 | 163.1241 | 4-Hydroxy cinnamic acid |
| 13 | 47.9 | 280 | 193.1726 | 193.1649 | 3-Methoxy-4-hydroxy cinnamic acid |
| 14 | 49.6 | 280 | 163.2547 | 163.2872 | 3-Hydroxy cinnamic acid |
| 15 | 49.0 | 280 | 223.1568 | 223.1748 | 4-Hydroxy-3,5-dimethoxy cinnamic acid |
| 16 | 67.3 | 280 | 147.1142 | 147.1103 | 3-Phenyl acrylic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarker, U.; Ercisli, S. Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves. Antioxidants 2022, 11, 2434. https://doi.org/10.3390/antiox11122434
Sarker U, Ercisli S. Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves. Antioxidants. 2022; 11(12):2434. https://doi.org/10.3390/antiox11122434
Chicago/Turabian StyleSarker, Umakanta, and Sezai Ercisli. 2022. "Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves" Antioxidants 11, no. 12: 2434. https://doi.org/10.3390/antiox11122434