
Alpha-Lipoic Acid as an Antioxidant Strategy for Managing Neuropathic Pain
 , , and
, , and 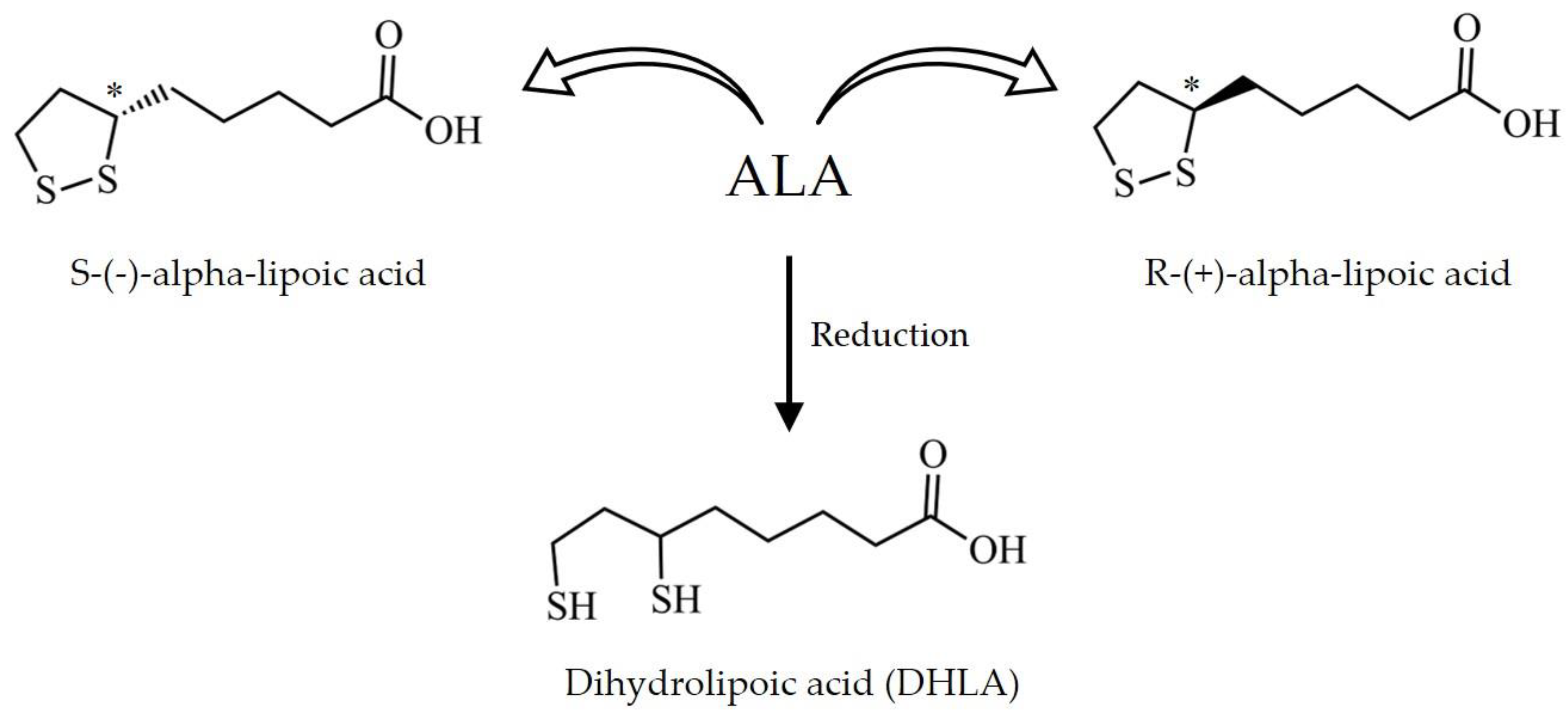{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Neuropathic Pain
3. Oxidative Stress and Neuropathic Pain
3.1. Mitochondrial Dysfunction and Pain Processing
3.2. NADPH Oxidases and Pain Processing
4. Antioxidant Potential of Alpha-Lipoic Acid in Neuropathic Pain
4.1. Chemical Properties and Pharmacokinetics
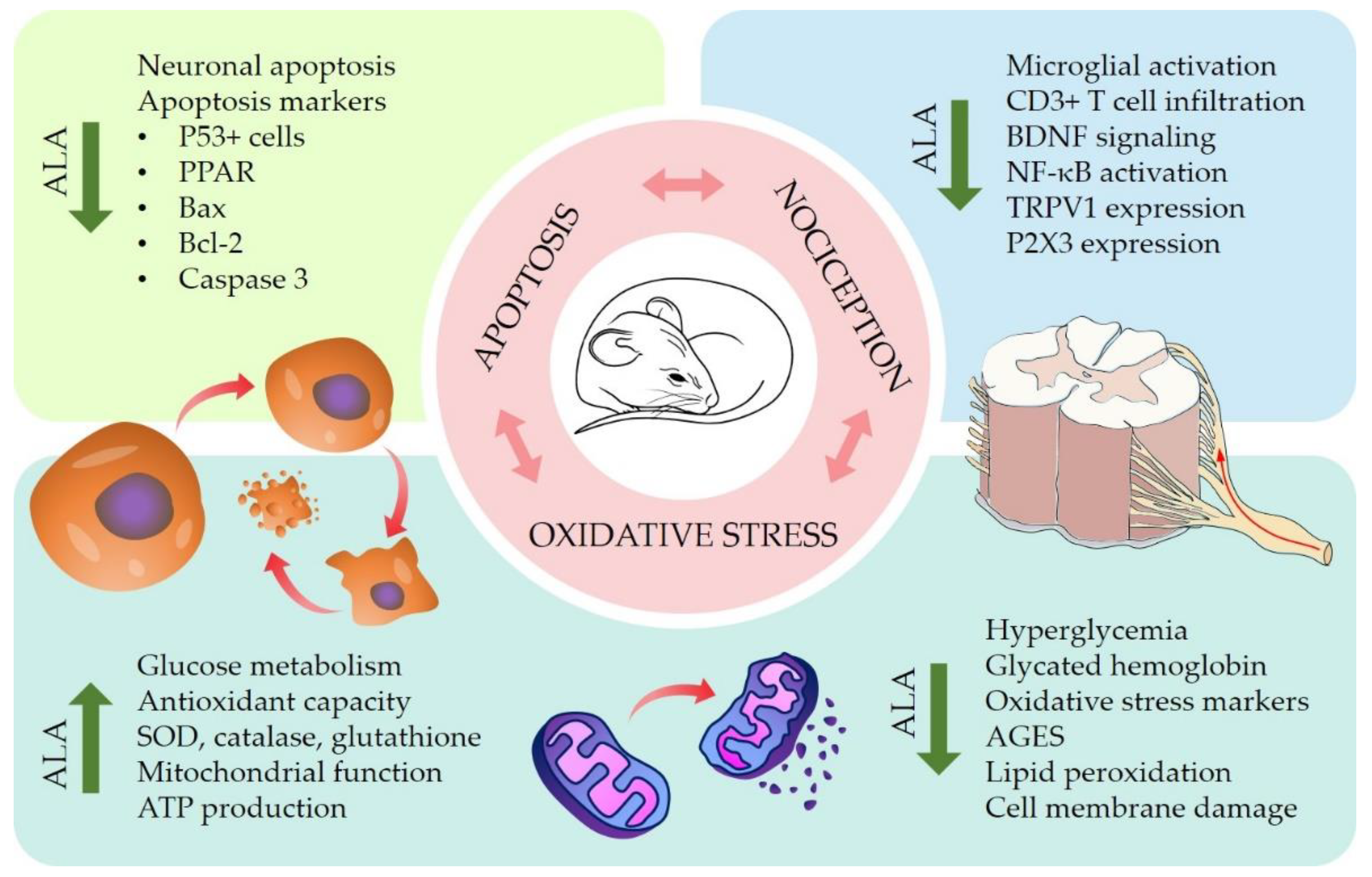
4.2. Pre-Clinical In Vitro and In Vivo Studies
4.3. Clinical Trials
5. Innovation and Alpha-Lipoic Acid-Based Products
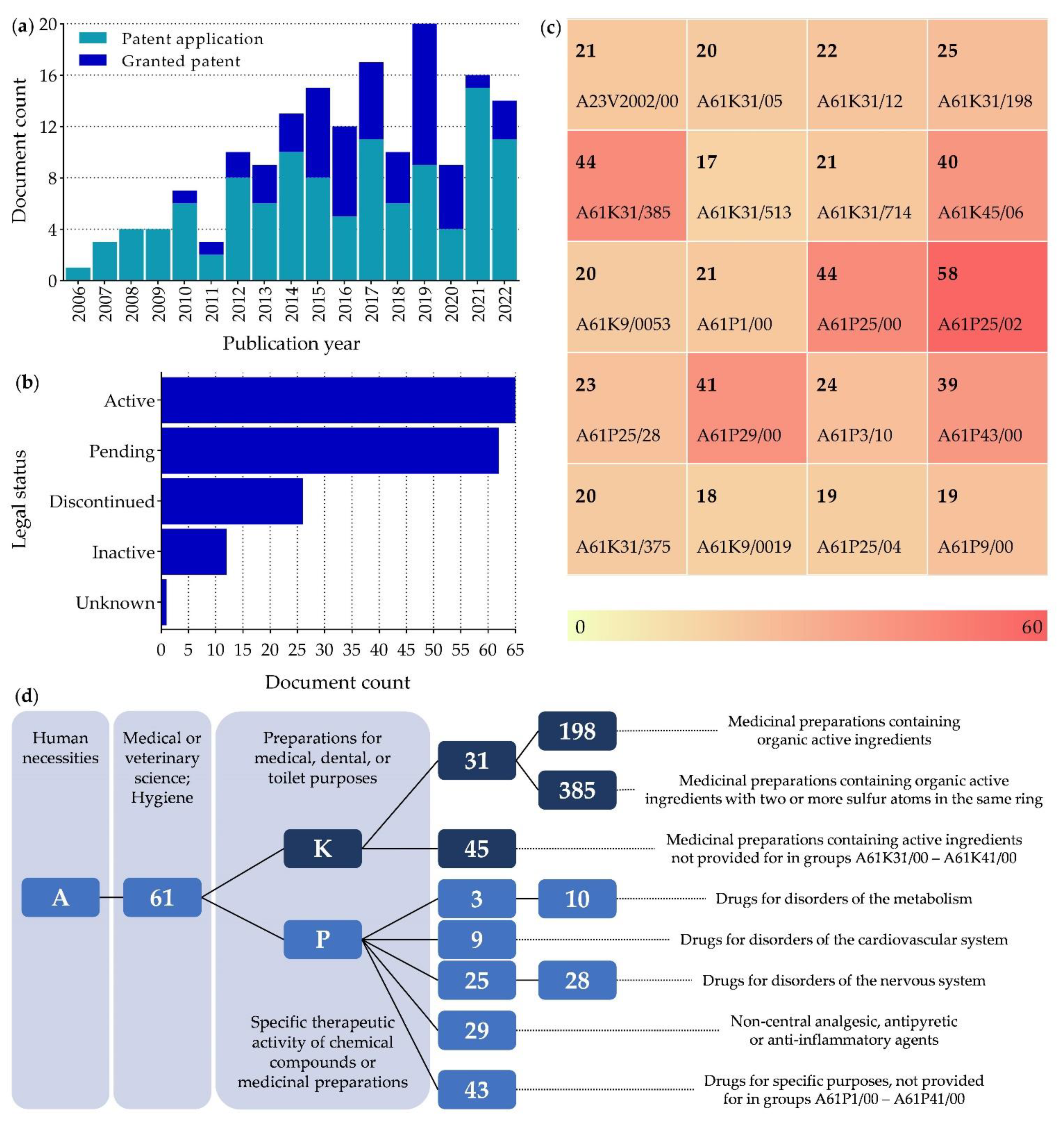
5.1. Patents
5.2. Market
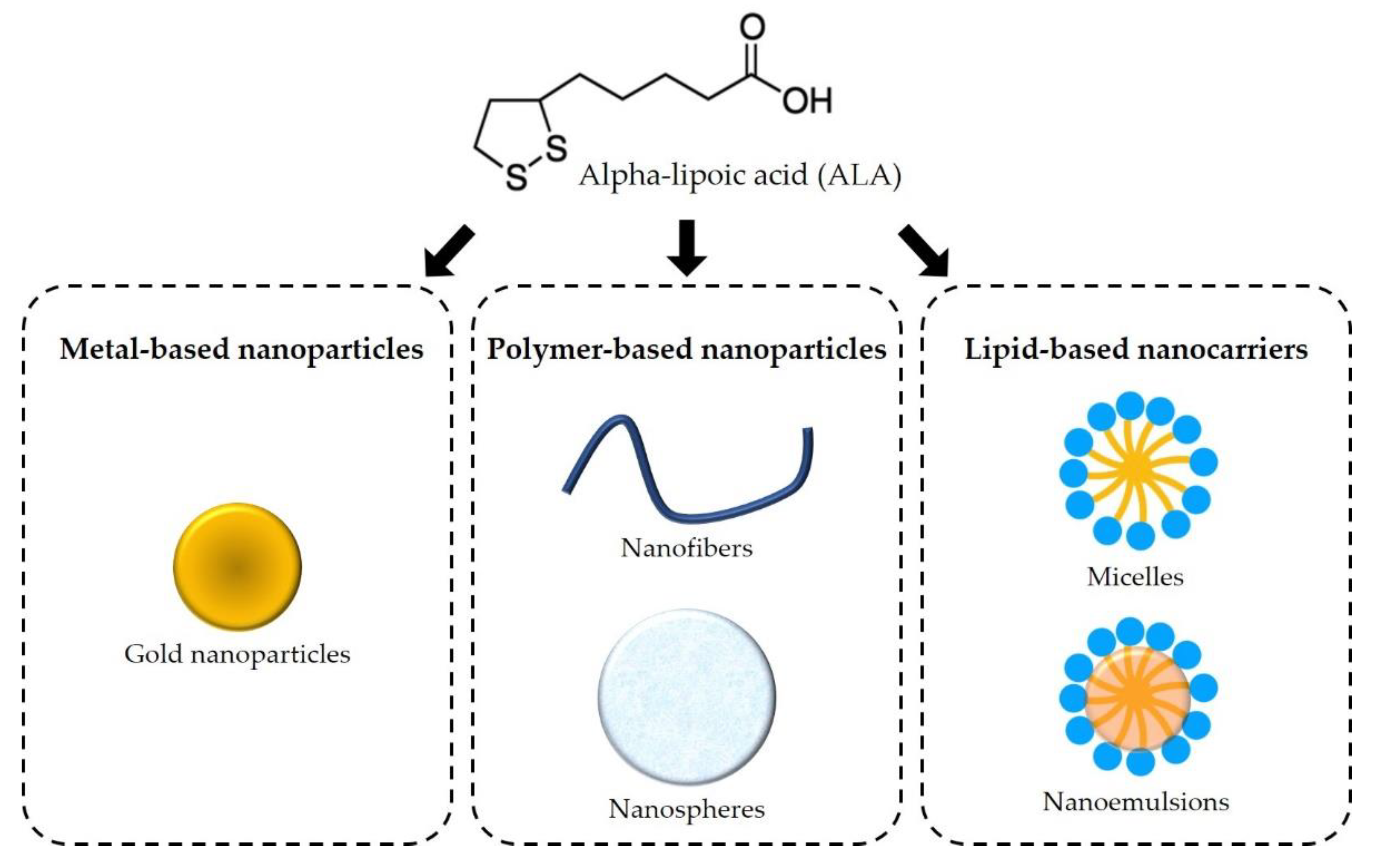
6. Novel Therapeutic Strategies
6.1. Metal-Based Nanoparticles
6.2. Polymer-Based Nanoparticles
6.3. Lipid-Based Nanocarriers
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| AGES | Advanced glycation end products |
| ALA | Alpha-lipoic acid |
| ASIC | Acid-sensing ion channels |
| ATP | Adenosine triphosphate |
| BDNF | Brain-derived neurotrophic factor |
| CPC | Cooperative patent classification |
| DHLA | Dihydrolipoic acid |
| DRG | Dorsal root ganglia |
| DSPN | Diabetic sensorimotor polyneuropathy |
| GNP | Gold nanoparticles |
| IL-1β | Interleukin 1 beta |
| IPC | International patent classification |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Nox | Nicotinamide adenine dinucleotide phosphate oxidases |
| NP | Neuropathic pain |
| PPAR | Poly (ADP-ribose) polymerase |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| STZ | Streptozotocin |
| TNF-α | Tumor necrosis factor alpha |
| TrkB | Tropomyosin receptor kinase B |
| TRPA1 | Transient receptor potential ankyrin subtype 1 |
| TRPV1 | Transient receptor potential vanilloid subtype 1 |
References
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Haroutounian, S.; Kamerman, P.; Baron, R.; Bennett, D.L.H.; Bouhassira, D.; Cruccu, G.; Freeman, R.; Hansson, P.; Nurmikko, T.; et al. Neuropathic pain: An updated grading system for research and clinical practice. Pain 2016, 157, 1599–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dworkin, R.H.; O’Connor, A.B.; Audette, J.; Baron, R.; Gourlay, G.K.; Haanpaa, M.L.; Kent, J.L.; Krane, E.J.; Lebel, A.A.; Levy, R.M.; et al. Recommendations for the pharmacological management of neuropathic pain: An overview and literature update. Mayo Clin. Proc. 2010, 85, S3–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, C.; Naziroglu, M.; Rodriguez, A.B.; Pariente, J.A. Neuropathic pain: Delving into the oxidative origin and the possible implication of transient receptor potential channels. Front. Physiol. 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wu, S.; Wang, J.; Wang, J.; Yan, Y.; Zhu, M.; Zhang, D.; Jiang, C.; Liu, T. Oxidative stress induced by NOX2 contributes to neuropathic pain via plasma membrane translocation of PKCepsilon in rat dorsal root ganglion neurons. J. Neuroinflamm. 2021, 18, 106. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Cassanego, G.; Rodrigues, P.; De Freitas Bauermann, L.; Trevisan, G. Evaluation of the analgesic effect of a-lipoic acid in treating pain disorders: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2022, 177, 106075. [Google Scholar] [CrossRef]
- Salehi, B.; Berkay Yilmaz, Y.; Antika, G.; Boyunegmez Tumer, T.; Fawzi Mahomoodally, M.; Lobine, D.; Akram, M.; Riaz, M.; Capanoglu, E.; Sharopov, F.; et al. Insights on the use of alpha-lipoic acid for therapeutic purposes. Biomolecules 2019, 9, 356. [Google Scholar] [CrossRef] [Green Version]
- Rochette, L.; Ghibu, S.; Richard, C.; Zeller, M.; Cottin, Y.; Vergely, C. Direct and indirect antioxidant properties of alpha-lipoic acid and therapeutic potential. Mol. Nutr. Food Res. 2013, 57, 114–125. [Google Scholar] [CrossRef]
- Bouhassira, D. Neuropathic pain: Definition, assessment and epidemiology. Rev. Neurol. 2019, 175, 16–25. [Google Scholar] [CrossRef]
- Abbott, C.A.; Malik, R.A.; van Ross, E.R.; Kulkarni, J.; Boulton, A.J. Prevalence and characteristics of painful diabetic neuropathy in a large community-based diabetic population in the UK. Diabetes Care 2011, 34, 2220–2224. [Google Scholar] [CrossRef] [Green Version]
- Baron, R.; Maier, C.; Attal, N.; Binder, A.; Bouhassira, D.; Cruccu, G.; Finnerup, N.B.; Haanpaa, M.; Hansson, P.; Hullemann, P.; et al. Peripheral neuropathic pain: A mechanism-related organizing principle based on sensory profiles. Pain 2017, 158, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet. Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, I.; Choucair-Jaafar, N.; Benbouzid, M.; Tessier, L.H.; Muller, A.; Hein, L.; Freund-Mercier, M.J.; Barrot, M. beta(2)-adrenoceptors are critical for antidepressant treatment of neuropathic pain. Ann. Neurol. 2009, 65, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Baron, R. Mechanisms of disease: Neuropathic pain, a clinical perspective. Nat. Clin. Pr. Neurol. 2006, 2, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Attal, N.; Lanteri-Minet, M.; Laurent, B.; Fermanian, J.; Bouhassira, D. The specific disease burden of neuropathic pain: Results of a French nationwide survey. Pain 2011, 152, 2836–2843. [Google Scholar] [CrossRef] [PubMed]
- Doth, A.H.; Hansson, P.T.; Jensen, M.P.; Taylor, R.S. The burden of neuropathic pain: A systematic review and meta-analysis of health utilities. Pain 2010, 149, 338–344. [Google Scholar] [CrossRef]
- Langley, P.C.; Van Litsenburg, C.; Cappelleri, J.C.; Carroll, D. The burden associated with neuropathic pain in Western Europe. J. Med. Econ. 2013, 16, 85–95. [Google Scholar] [CrossRef]
- Park, E.S.; Gao, X.; Chung, J.M.; Chung, K. Levels of mitochondrial reactive oxygen species increase in rat neuropathic spinal dorsal horn neurons. Neurosci. Lett. 2006, 391, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Fairchild, T.J.; Vo, L.; Drummond, P.D. Painful diabetic peripheral neuropathy: Role of oxidative stress and central sensitisation. Diabet. Med. A J. Br. Diabet. Assoc. 2022, 39, e14729. [Google Scholar] [CrossRef] [PubMed]
- Negi, G.; Kumar, A.; Joshi, R.P.; Sharma, S.S. Oxidative stress and Nrf2 in the pathophysiology of diabetic neuropathy: Old perspective with a new angle. Biochem. Biophys. Res. Commun. 2011, 408, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Makkar, T.K.; Goel, L.; Pahuja, M. Role of inflammation and oxidative stress in chemotherapy-induced neurotoxicity. Immunol. Res. 2022, 70, 725–741. [Google Scholar] [CrossRef]
- Ilari, S.; Giancotti, L.A.; Lauro, F.; Gliozzi, M.; Malafoglia, V.; Palma, E.; Tafani, M.; Russo, M.A.; Tomino, C.; Fini, M.; et al. Natural antioxidant control of neuropathic pain-exploring the role of mitochondrial SIRT3 pathway. Antioxidants 2020, 9, 1103. [Google Scholar] [CrossRef]
- Saifi, G.M.; Szigeti, K.; Snipes, G.J.; Garcia, C.A.; Lupski, J.R. Molecular mechanisms, diagnosis, and rational approaches to management of and therapy for Charcot-Marie-Tooth disease and related peripheral neuropathies. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2003, 51, 261–283. [Google Scholar] [CrossRef]
- Siotto, M.; Aprile, I.; Simonelli, I.; Pazzaglia, C.; Ventriglia, M.; Santoro, M.; Imbimbo, I.; Squitti, R.; Padua, L. An exploratory study of BDNF and oxidative stress marker alterations in subacute and chronic stroke patients affected by neuropathic pain. J. Neural Transm. 2017, 124, 1557–1566. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Salvemini, D.; Little, J.W.; Doyle, T.; Neumann, W.L. Roles of reactive oxygen and nitrogen species in pain. Free Radic. Biol. Med. 2011, 51, 951–966. [Google Scholar] [CrossRef] [Green Version]
- Janes, K.; Doyle, T.; Bryant, L.; Esposito, E.; Cuzzocrea, S.; Ryerse, J.; Bennett, G.J.; Salvemini, D. Bioenergetic deficits in peripheral nerve sensory axons during chemotherapy-induced neuropathic pain resulting from peroxynitrite-mediated post-translational nitration of mitochondrial superoxide dismutase. Pain 2013, 154, 2432–2440. [Google Scholar] [CrossRef]
- Munoz, F.M.; Gao, R.; Tian, Y.; Henstenburg, B.A.; Barrett, J.E.; Hu, H. Neuronal P2X7 receptor-induced reactive oxygen species production contributes to nociceptive behavior in mice. Sci. Rep. 2017, 7, 3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallenborn-Gerhardt, W.; Schröder, K.; Schmidtko, A. NADPH oxidases in pain processing. Antioxidants 2022, 11, 1162. [Google Scholar] [CrossRef] [PubMed]
- Doyle, T.M.; Salvemini, D. Mini-Review: Mitochondrial dysfunction and chemotherapy-induced neuropathic pain. Neurosci. Lett. 2021, 760, 136087. [Google Scholar] [CrossRef] [PubMed]
- Román-Pintos, L.M.; Villegas-Rivera, G.; Rodríguez-Carrizalez, A.D.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. Diabetic polyneuropathy in type 2 diabetes mellitus: Inflammation, oxidative stress, and mitochondrial function. J. Diabetes Res. 2016, 2016, 3425617. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.L.L.; Santos, G.G.L.; Espirito-Santo, R.F.; Silva, G.S.A.; Evangelista, A.F.; Silva, D.N.; Soares, M.B.P.; Villarreal, C.F. Reestablishment of redox homeostasis in the nociceptive primary afferent as a mechanism of antinociception promoted by mesenchymal stem/stromal cells in oxaliplatin-induced chronic peripheral neuropathy. Stem Cells Int. 2021, 2021, 8815206. [Google Scholar] [CrossRef]
- Leo, M.; Schmitt, L.I.; Küsterarent, P.; Kutritz, A.; Rassaf, T.; Kleinschnitz, C.; Hendgen-Cotta, U.B.; Hagenacker, T. Platinum-based drugs cause mitochondrial dysfunction in cultured dorsal root ganglion neurons. Int. J. Mol. Sci. 2020, 21, 8636. [Google Scholar] [CrossRef]
- Flatters, S.J.L.; Bennett, G.J. Studies of peripheral sensory nerves in paclitaxel-induced painful peripheral neuropathy: Evidence for mitochondrial dysfunction. Pain 2006, 122, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.C.; Bartol, T.M.; Phan, S.; Bushong, E.A.; Perkins, G.; Sejnowski, T.J.; Ellisman, M.H.; Skupin, A. Mitochondrial morphology provides a mechanism for energy buffering at synapses. Sci. Rep. 2019, 9, 18306. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.H.; Bennett, G.J. Effects of mitochondrial poisons on the neuropathic pain produced by the chemotherapeutic agents, paclitaxel and oxaliplatin. Pain 2012, 153, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Ludman, T.; Melemedjian, O.K. Bortezomib-induced aerobic glycolysis contributes to chemotherapy-induced painful peripheral neuropathy. Mol. Pain 2019, 15, 1744806919837429. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Fabisiak, T.; Patel, M. Crosstalk between neuroinflammation and oxidative stress in epilepsy. Front. Cell Dev. Biol. 2022, 10, 976953. [Google Scholar] [CrossRef] [PubMed]
- Bettiol, A.; Galora, S.; Argento, F.R.; Fini, E.; Emmi, G.; Mattioli, I.; Bagni, G.; Fiorillo, C.; Becatti, M. Erythrocyte oxidative stress and thrombosis. Expert Rev. Mol. Med. 2022, 24, e31. [Google Scholar] [CrossRef]
- De Logu, F.; Nassini, R.; Materazzi, S.; Carvalho Gonçalves, M.; Nosi, D.; Rossi Degl’Innocenti, D.; Marone, I.M.; Ferreira, J.; Li Puma, S.; Benemei, S.; et al. Schwann cell TRPA1 mediates neuroinflammation that sustains macrophage-dependent neuropathic pain in mice. Nat. Commun. 2017, 8, 1887. [Google Scholar] [CrossRef] [Green Version]
- Kallenborn-Gerhardt, W.; Hohmann, S.W.; Syhr, K.M.; Schröder, K.; Sisignano, M.; Weigert, A.; Lorenz, J.E.; Lu, R.; Brüne, B.; Brandes, R.P.; et al. Nox2-dependent signaling between macrophages and sensory neurons contributes to neuropathic pain hypersensitivity. Pain 2014, 155, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Geis, C.; Geuss, E.; Sommer, C.; Schmidt, H.H.; Kleinschnitz, C. NOX4 is an early initiator of neuropathic pain. Exp. Neurol. 2017, 288, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Sun, B.; Liu, Z.; Yao, X.; Wang, H.; Shen, X.; Jiang, H.; Chen, J. Advanced oxidative protein products cause pain hypersensitivity in rats by inducing dorsal root ganglion neurons apoptosis via NADPH oxidase 4/c-Jun N-terminal kinase pathways. Front. Mol. Neurosci. 2017, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Miao, F.; Wang, R.; Cui, G.; Li, X.; Wang, T.; Li, X. Engagement of microRNA-155 in exaggerated oxidative stress signal and TRPA1 in the dorsal horn of the spinal cord and neuropathic pain during chemotherapeutic oxaliplatin. Neurotox. Res. 2019, 36, 712–723. [Google Scholar] [CrossRef]
- Kallenborn-Gerhardt, W.; Schröder, K.; Del Turco, D.; Lu, R.; Kynast, K.; Kosowski, J.; Niederberger, E.; Shah, A.M.; Brandes, R.P.; Geisslinger, G.; et al. NADPH oxidase-4 maintains neuropathic pain after peripheral nerve injury. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 10136–10145. [Google Scholar] [CrossRef]
- Eid, S.A.; El Massry, M.; Hichor, M.; Haddad, M.; Grenier, J.; Dia, B.; Barakat, R.; Boutary, S.; Chanal, J.; Aractingi, S.; et al. Targeting the NADPH oxidase-4 and liver X receptor pathway preserves Schwann cell integrity in diabetic mice. Diabetes 2020, 69, 448–464. [Google Scholar] [CrossRef]
- Zhao, W.C.; Zhang, B.; Liao, M.J.; Zhang, W.X.; He, W.Y.; Wang, H.B.; Yang, C.X. Curcumin ameliorated diabetic neuropathy partially by inhibition of NADPH oxidase mediating oxidative stress in the spinal cord. Neurosci. Lett. 2014, 560, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Lu, J.H.; Xie, C.S.; Shen, Y.J.; Wang, J.W.; Ye, X.Y.; Zhang, M.B.; Jia, G.L.; Tao, Y.X.; Li, J.; et al. Caveolin-1 in spinal cord modulates type-2 diabetic neuropathic pain through the Rac1/NOX2/NR2B signaling pathway. Am. J. Transl. Res. 2020, 12, 1714–1727. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Sharma, S.; Chopra, K. Lycopene attenuates thermal hyperalgesia in a diabetic mouse model of neuropathic pain. Eur. J. Pain 2008, 12, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Goel, R.; Tyagi, N. Potential contribution of antioxidant mechanism in the defensive effect of lycopene against partial sciatic nerve ligation induced behavioral, biochemical and histopathological modification in Wistar rats. Drug Res. 2016, 66, 633–638. [Google Scholar] [CrossRef]
- Recalde, M.D.; Miguel, C.A.; Noya-Riobó, M.V.; González, S.L.; Villar, M.J.; Coronel, M.F. Resveratrol exerts anti-oxidant and anti-inflammatory actions and prevents oxaliplatin-induced mechanical and thermal allodynia. Brain Res. 2020, 1748, 147079. [Google Scholar] [CrossRef]
- Joseph, E.K.; Chen, X.; Bogen, O.; Levine, J.D. Oxaliplatin acts on IB4-positive nociceptors to induce an oxidative stress-dependent acute painful peripheral neuropathy. J. Pain 2008, 9, 463–472. [Google Scholar] [CrossRef]
- Moura, F.A.; de Andrade, K.Q.; dos Santos, J.C.; Goulart, M.O. Lipoic acid: Its antioxidant and anti-inflammatory role and clinical applications. Curr. Top. Med. Chem. 2015, 15, 458–483. [Google Scholar] [CrossRef]
- Kishimoto-Urata, M.; Urata, S.; Fujimoto, C.; Yamasoba, T. Role of oxidative stress and antioxidants in acquired inner ear disorders. Antioxidants 2022, 11, 1469. [Google Scholar] [CrossRef]
- Brookes, M.H.; Golding, B.T.; Howes, D.A.; Hudson, A.T. Proof that the absolute configuration of natural alpha-lipoic acid is R by the synthesis of its enantiomer [(S)-(–)-alpha-lipoic acid] from (S)-malic acid. J. Chem. Soc. Chem. Commun. 1983, 19, 1051–1053. [Google Scholar] [CrossRef]
- Ghibu, S.; Richard, C.; Vergely, C.; Zeller, M.; Cottin, Y.; Rochette, L. Antioxidant properties of an endogenous thiol: Alpha-lipoic acid, useful in the prevention of cardiovascular diseases. J. Cardiovasc. Pharmacol. 2009, 54, 391–398. [Google Scholar] [CrossRef]
- Szeląg, M.; Mikulski, D.; Molski, M. Quantum-chemical investigation of the structure and the antioxidant properties of α-lipoic acid and its metabolites. J. Mol. Model. 2012, 18, 2907–2916. [Google Scholar] [CrossRef] [Green Version]
- Akiba, S.; Matsugo, S.; Packer, L.; Konishi, T. Assay of protein-bound lipoic acid in tissues by a new enzymatic method. Anal. Biochem. 1998, 258, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Hermann, R.; Niebch, G.D.; Borbe, H.O.; Fieger-Büschges, H.; Ruus, P.; Nowak, H.; Riethmüller-Winzen, H.; Peukert, M.; Blume, H.H. Enantioselective pharmacokinetics and bioavailability of different racemic α-lipoic acid formulations in healthy volunteers. Eur. J. Pharm. Sci. 1996, 4, 167–174. [Google Scholar] [CrossRef]
- Golbidi, S.; Badran, M.; Laher, I. Diabetes and alpha lipoic acid. Front. Pharmacol. 2011, 2, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochette, L.; Ghibu, S.; Muresan, A.; Vergely, C. Alpha-lipoic acid: Molecular mechanisms and therapeutic potential in diabetes. Can. J. Physiol. Pharmacol. 2015, 93, 1021–1027. [Google Scholar] [CrossRef]
- Mignini, F.; Nasuti, C.; Gioventu, G.; Napolioni, V.; Di Martino, P. Human bioavailability and pharmacokinetic profile of different formulations delivering alpha lipoic acid. J. Clin. Cell. Immunol. 2012, 1, 418. [Google Scholar] [CrossRef]
- Brufani, M.; Figliola, R. (R)-α-lipoic acid oral liquid formulation: Pharmacokinetic parameters and therapeutic efficacy. Acta Bio-Med. Atenei Parm. 2014, 85, 108–115. [Google Scholar]
- Gleiter, C.H.; Schug, B.S.; Hermann, R.; Elze, M.; Blume, H.H.; Gundert-Remy, U. Influence of food intake on the bioavailability of thioctic acid enantiomers. Eur. J. Clin. Pharmacol. 1996, 50, 513–514. [Google Scholar] [CrossRef]
- Packer, L.; Kraemer, K.; Rimbach, G. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition 2001, 17, 888–895. [Google Scholar] [CrossRef]
- May, J.M.; Qu, Z.C.; Nelson, D.J. Uptake and reduction of alpha-lipoic acid by human erythrocytes. Clin. Biochem. 2007, 40, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Schupke, H.; Hempel, R.; Peter, G.; Hermann, R.; Wessel, K.; Engel, J.; Kronbach, T. New metabolic pathways of alpha-lipoic acid. Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 855–862. [Google Scholar] [PubMed]
- Bock, E.; Schneeweiss, J. Ein beitrag zur therapie der neuropathia diabetica. Munch. Med. Wochenschr. 1959, 101, 1911–1912. [Google Scholar] [PubMed]
- Packer, L. alpha-Lipoic acid: A metabolic antioxidant which regulates NF-kappa B signal transduction and protects against oxidative injury. Drug Metab. Rev. 1998, 30, 245–275. [Google Scholar] [CrossRef] [PubMed]
- Biewenga, G.P.; Haenen, G.R.; Bast, A. The pharmacology of the antioxidant lipoic acid. Gen. Pharmacol. 1997, 29, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.J.; Tsuchiya, M.; Packer, L. Thioctic acid and dihydrolipoic acid are novel antioxidants which interact with reactive oxygen species. Free Radic. Res. Commun. 1991, 15, 255–263. [Google Scholar] [CrossRef]
- Wray, D.W.; Nishiyama, S.K.; Harris, R.A.; Zhao, J.; McDaniel, J.; Fjeldstad, A.S.; Witman, M.A.; Ives, S.J.; Barrett-O’Keefe, Z.; Richardson, R.S. Acute reversal of endothelial dysfunction in the elderly after antioxidant consumption. Hypertension 2012, 59, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Reljanovic, M.; Reichel, G.; Rett, K.; Lobisch, M.; Schuette, K.; Möller, W.; Tritschler, H.J.; Mehnert, H. Treatment of diabetic polyneuropathy with the antioxidant thioctic acid (alpha-lipoic acid): A two year multicenter randomized double-blind placebo-controlled trial (ALADIN II). Alpha Lipoic Acid in Diabetic Neuropathy. Free Radic. Res. 1999, 31, 171–179. [Google Scholar] [CrossRef]
- Akter, N. Diabetic peripheral neuropathy: Epidemiology, physiopathology, diagnosis and treatment. Delta Med. Coll. J. 2019, 7, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Pang, L.; Lian, X.; Liu, H.; Zhang, Y.; Li, Q.; Cai, Y.; Ma, H.; Yu, X. Understanding diabetic neuropathy: Focus on oxidative stress. Oxidative Med. Cell. Longev. 2020, 2020, 9524635. [Google Scholar] [CrossRef]
- Chen, J.; Li, Q. Lipoic acid decreases the expression of poly ADP-ribose polymerase and inhibits apoptosis in diabetic rats. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1725–1731. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Zhang, Y.L.; Sun, Q.; Zhang, P.A.; Wang, X.X.; Xu, G.Y.; Hu, J.; Zhang, H.H. Alpha-lipoic acid downregulates TRPV1 receptor via NF-κB and attenuates neuropathic pain in rats with diabetes. CNS Neurosci. Ther. 2020, 26, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Sadeghiyan Galeshkalami, N.; Abdollahi, M.; Najafi, R.; Baeeri, M.; Jamshidzade, A.; Falak, R.; Davoodzadeh Gholami, M.; Hassanzadeh, G.; Mokhtari, T.; Hassani, S.; et al. Alpha-lipoic acid and coenzyme Q10 combination ameliorates experimental diabetic neuropathy by modulating oxidative stress and apoptosis. Life Sci. 2019, 216, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Najafi, R.; Sharifi, A.M.; Hosseini, A. Protective effects of alpha lipoic acid on high glucose-induced neurotoxicity in PC12 cells. Metab. Brain Dis. 2015, 30, 731–738. [Google Scholar] [CrossRef]
- Siniscalco, D.; Fuccio, C.; Giordano, C.; Ferraraccio, F.; Palazzo, E.; Luongo, L.; Rossi, F.; Roth, K.A.; Maione, S.; de Novellis, V. Role of reactive oxygen species and spinal cord apoptotic genes in the development of neuropathic pain. Pharmacol. Res. 2007, 55, 158–166. [Google Scholar] [CrossRef]
- Jain, S.K.; Lim, G. Lipoic acid decreases lipid peroxidation and protein glycosylation and increases (Na+ + K+)- and Ca++-ATPase activities in high glucose-treated human erythrocytes. Free Radic. Biol. Med. 2000, 29, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Qin, X.; Song, Z.Y.; Yang, P.P.; Feng, Y.; Sun, Q.; Xu, G.Y.; Zhang, H.H. Alpha-lipoic acid suppresses P2X receptor activities and visceral hypersensitivity to colorectal distention in diabetic rats. Sci. Rep. 2017, 7, 3928. [Google Scholar] [CrossRef]
- Fei, X.; He, X.; Tai, Z.; Wang, H.; Qu, S.; Chen, L.; Hu, Q.; Fang, J.; Jiang, Y. Electroacupuncture alleviates diabetic neuropathic pain in rats by suppressing P2X3 receptor expression in dorsal root ganglia. Purinergic Signal. 2020, 16, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.; Momeni, Z.; Theaker, M.; Jagadeeshan, S.; Yamamoto, Y.; Ianowski, J.P.; Campanucci, V.A. RAGE-dependent potentiation of TRPV1 currents in sensory neurons exposed to high glucose. PLoS ONE 2018, 13, e0193312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, M.B.; Negrato, C.A. Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Martorana, F.; Foti, M.; Virtuoso, A.; Gaglio, D.; Aprea, F.; Latronico, T.; Rossano, R.; Riccio, P.; Papa, M.; Alberghina, L.; et al. Differential Modulation of NF-κB in Neurons and Astrocytes Underlies Neuroprotection and Antigliosis Activity of Natural Antioxidant Molecules. Oxidative Med. Cell. Longev. 2019, 2019, 8056904. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.X.; Wang, H.; Song, H.L.; Wang, J.; Zhang, Z.J. Neuroinflammation Involved in Diabetes-Related Pain and Itch. Front. Pharmacol. 2022, 13, 921612. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lou, Z.; Xi, H.; Li, Z.; Li, L.; Li, Z.; Zhang, K.; Asakawa, T. Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model. Open Life Sci. 2021, 16, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Gordon, R.; Woodruff, T.M.; Smith, M.T. Antiallodynic effects of alpha lipoic acid in an optimized RR-EAE mouse model of MS-neuropathic pain are accompanied by attenuation of upregulated BDNF-TrkB-ERK signaling in the dorsal horn of the spinal cord. Pharmacol. Res. Perspect. 2015, 3, e00137. [Google Scholar] [CrossRef] [PubMed]
- Melli, G.; Taiana, M.; Camozzi, F.; Triolo, D.; Podini, P.; Quattrini, A.; Taroni, F.; Lauria, G. Alpha-lipoic acid prevents mitochondrial damage and neurotoxicity in experimental chemotherapy neuropathy. Exp. Neurol. 2008, 214, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E. Mouse Sleeping. Available online: https://www.scidraw.io/drawing/285 (accessed on 11 November 2022).
- Georgievckaya, K. Spine Cord. Available online: https://www.scidraw.io/drawing/454 (accessed on 11 November 2022).
- Mijnhout, G.S.; Kollen, B.J.; Alkhalaf, A.; Kleefstra, N.; Bilo, H.J. Alpha lipoic acid for symptomatic peripheral neuropathy in patients with diabetes: A meta-analysis of randomized controlled trials. Int. J. Endocrinol. 2012, 2012, 456279. [Google Scholar] [CrossRef]
- Kulaklı, F. The effect of alpha lipoic acid in the treatment of multiple sclerosis induced neuropathic pain: A case report. Eurasian J. Med. Oncol. 2018, 2, 179–181. [Google Scholar] [CrossRef]
- Ziegler, D.; Papanas, N.; Schnell, O.; Nguyen, B.D.T.; Nguyen, K.T.; Kulkantrakorn, K.; Deerochanawong, C. Current concepts in the management of diabetic polyneuropathy. J. Diabetes Investig. 2021, 12, 464–475. [Google Scholar] [CrossRef]
- Abdelrahman, K.A.; Ibrahim, A.S.; Osman, A.M.; Aly, M.G.; Ali, A.S.; Farrag, W.S. Alpha lipoic acid with pulsed radiofrequency in treatment of chronic lumbosacral radicular pain: A prospective, randomized study. Medicine 2021, 100, e26344. [Google Scholar] [CrossRef]
- Barreto da Silva, L.; Camargo, S.B.; Moraes, R.D.A.; Medeiros, C.F.; Jesus, A.M.; Evangelista, A.; Villarreal, C.F.; Quintans-Júnior, L.J.; Silva, D.F. Antihypertensive effect of carvacrol is improved after incorporation in β-cyclodextrin as a drug delivery system. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1798–1807. [Google Scholar] [CrossRef]
- Crescenzo, A.D.; Cacciatore, I.; Petrini, M.; D’Alessandro, M.; Petragnani, N.; Boccio, P.D.; Profio, P.D.; Boncompagni, S.; Spoto, G.; Turkez, H.; et al. Gold nanoparticles as scaffolds for poor water soluble and difficult to vehiculate antiparkinson codrugs. Nanotechnology 2017, 28, 025102. [Google Scholar] [CrossRef]
- Piersimoni, M.E.; Teng, X.; Cass, A.E.G.; Ying, L. Antioxidant lipoic acid ligand-shell gold nanoconjugates against oxidative stress caused by α-synuclein aggregates. Nanoscale Adv. 2020, 2, 5666–5681. [Google Scholar] [CrossRef] [PubMed]
- Aljaeid, B.M.; El-Moselhy, M.A. Loading of gentamicin and alpha lipoic acid on a biodegradable polymer for more effective and less nephrotoxic formula. Int. J. Pharmacol. 2018, 14, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Haidar, M.K.; Timur, S.S.; Kazanci, A.; Turkoglu, O.F.; Gürsoy, R.N.; Nemutlu, E.; Sargon, M.F.; Bodur, E.; Gök, M.; Ulubayram, K.; et al. Composite nanofibers incorporating alpha lipoic acid and atorvastatin provide neuroprotection after peripheral nerve injury in rats. Eur. J. Pharm. Biopharm. Off. J. Arb. Fur Pharm. Verfahr. 2020, 153, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, O.I.; Berezhnoy, D.S.; Stvolinsky, S.L.; Lopachev, A.V.; Orlova, V.S.; Fedorova, T.N. Neuroprotective effect of the carnosine—α-lipoic acid nanomicellar complex in a model of early-stage Parkinson’s disease. Regul. Toxicol. Pharmacol. RTP 2018, 95, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Musashi, M.; Nagasawa, T.; Shimura, N.; Igarashi, R.; Yamaguchi, Y. Novel nanocapsule of α-lipoic acid reveals pigmentation improvement: α-Lipoic acid stimulates the proliferation and differentiation of keratinocyte in murine skin by topical application. Exp. Dermatol. 2019, 28 (Suppl. 1), 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çoban, Ö.; Yıldırım, S.; Bakır, T. Alpha-lipoic acid and cyanocobalamin co-loaded nanoemulsions: Development, characterization, and evaluation of stability. J. Pharm. Innov. 2022, 17, 510–520. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viana, M.D.M.; Lauria, P.S.S.; Lima, A.A.d.; Opretzka, L.C.F.; Marcelino, H.R.; Villarreal, C.F. Alpha-Lipoic Acid as an Antioxidant Strategy for Managing Neuropathic Pain. Antioxidants 2022, 11, 2420. https://doi.org/10.3390/antiox11122420
Viana MDM, Lauria PSS, Lima AAd, Opretzka LCF, Marcelino HR, Villarreal CF. Alpha-Lipoic Acid as an Antioxidant Strategy for Managing Neuropathic Pain. Antioxidants. 2022; 11(12):2420. https://doi.org/10.3390/antiox11122420
Chicago/Turabian StyleViana, Max Denisson Maurício, Pedro Santana Sales Lauria, Alyne Almeida de Lima, Luiza Carolina França Opretzka, Henrique Rodrigues Marcelino, and Cristiane Flora Villarreal. 2022. "Alpha-Lipoic Acid as an Antioxidant Strategy for Managing Neuropathic Pain" Antioxidants 11, no. 12: 2420. https://doi.org/10.3390/antiox11122420