
Cathelicidin Attenuates Hyperoxia-Induced Lung Injury by Inhibiting Ferroptosis in Newborn Rats

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Experimental Groups
2.2. Measurement of Ferroptosis Indicators
2.3. Determination of Iron Deposition
2.4. Western Blots of GPX4 and TOMM20
2.5. Immunohistochemistry of TOMM20 and Cathelicidin
2.6. Lung Histology
2.7. Lung Cytokine Assay
2.8. Statistical Analysis
3. Results
3.1. Survival Rate
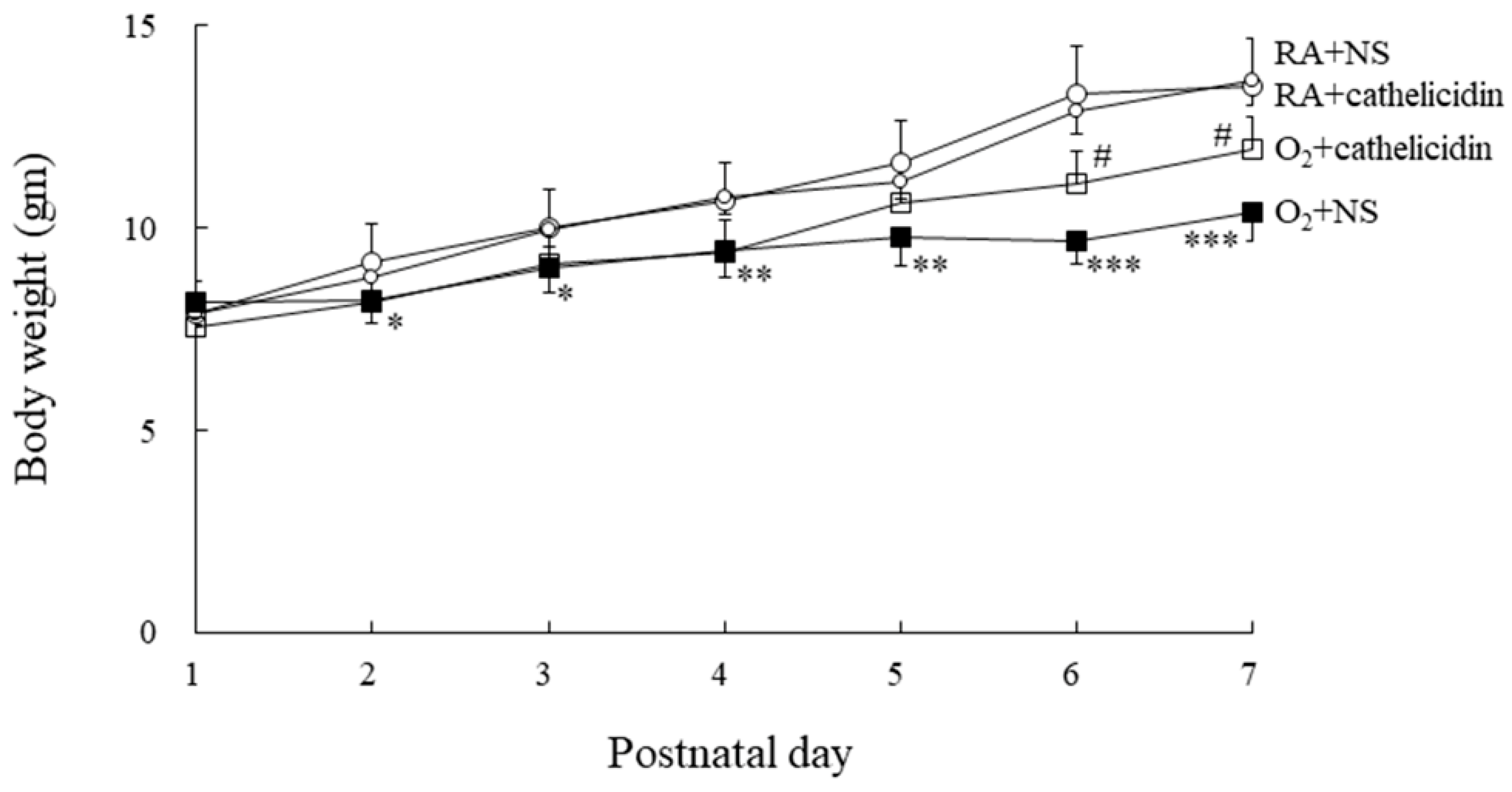
3.2. Cathelicidin Improved Hyperoxia-Induced Decrease in Body Weight
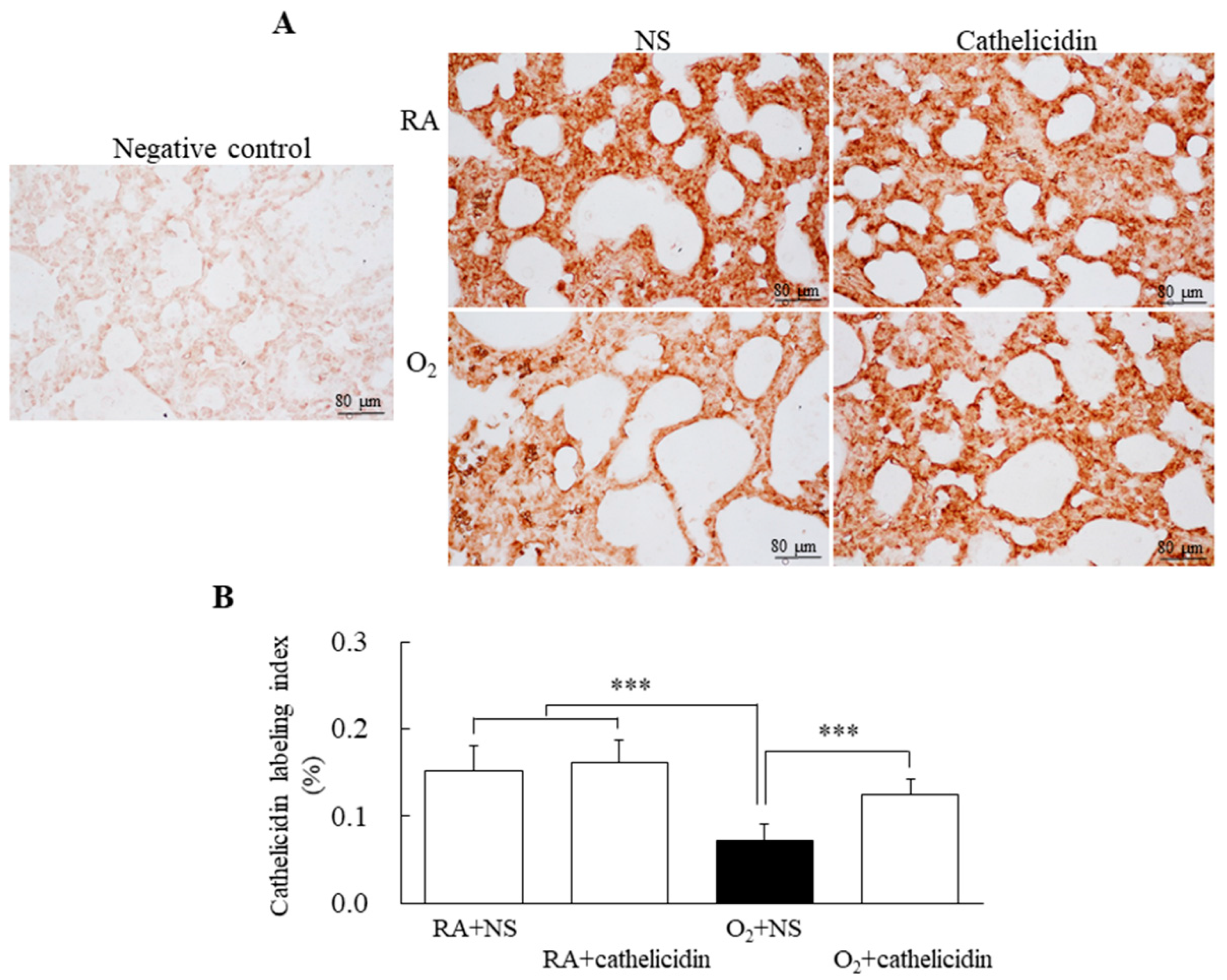
3.3. Hyperoxia Decreased Lung Cathelicidin Expression
3.4. Cathelicidin Improved Hyperoxia-Induced Ferroptosis
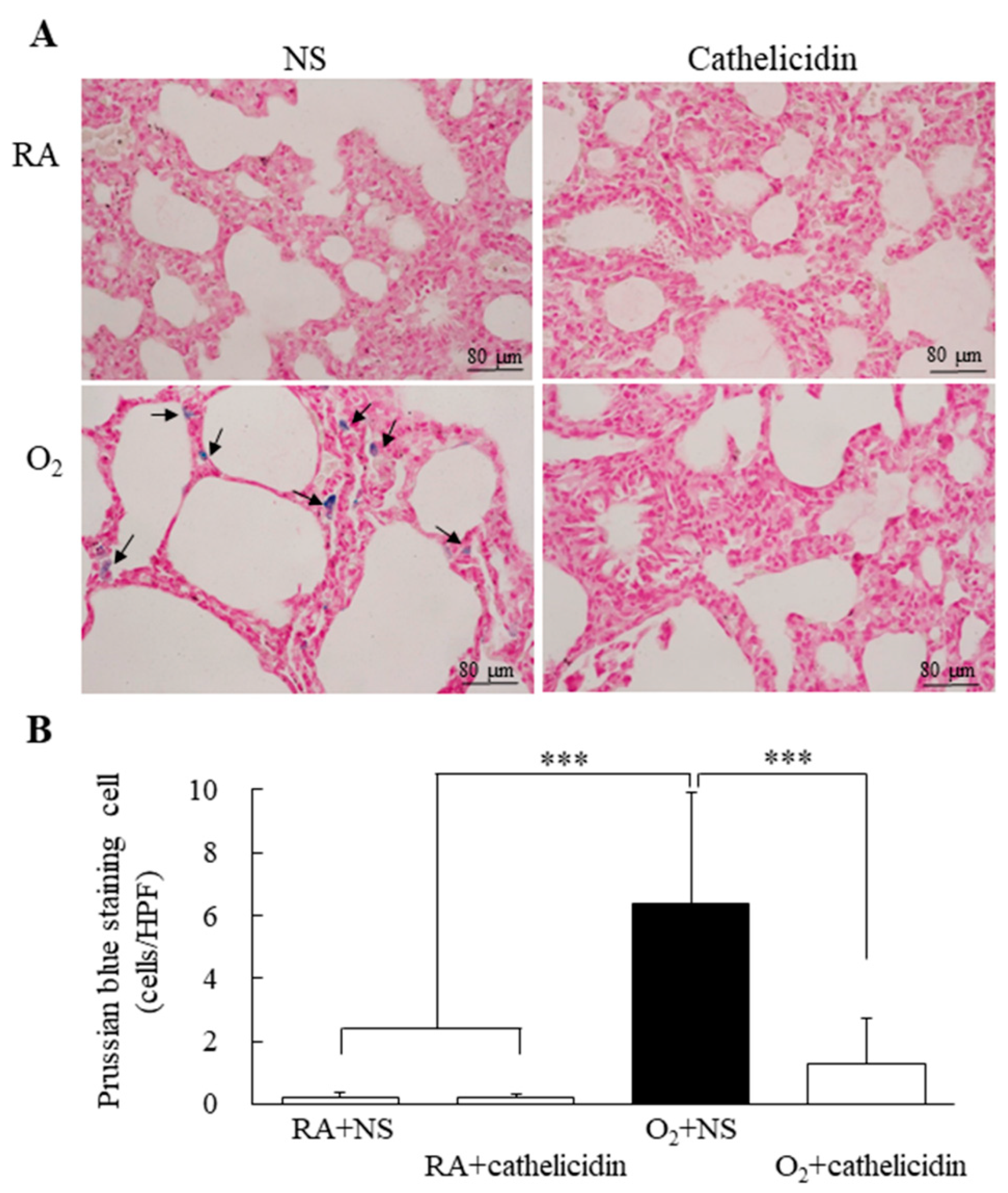
3.5. Cathelicidin Decreased Hyperoxia-Induced Increase in Iron Deposition
3.6. Cathelicidin Decreased Hyperoxia-Induced Increase in TOMM20
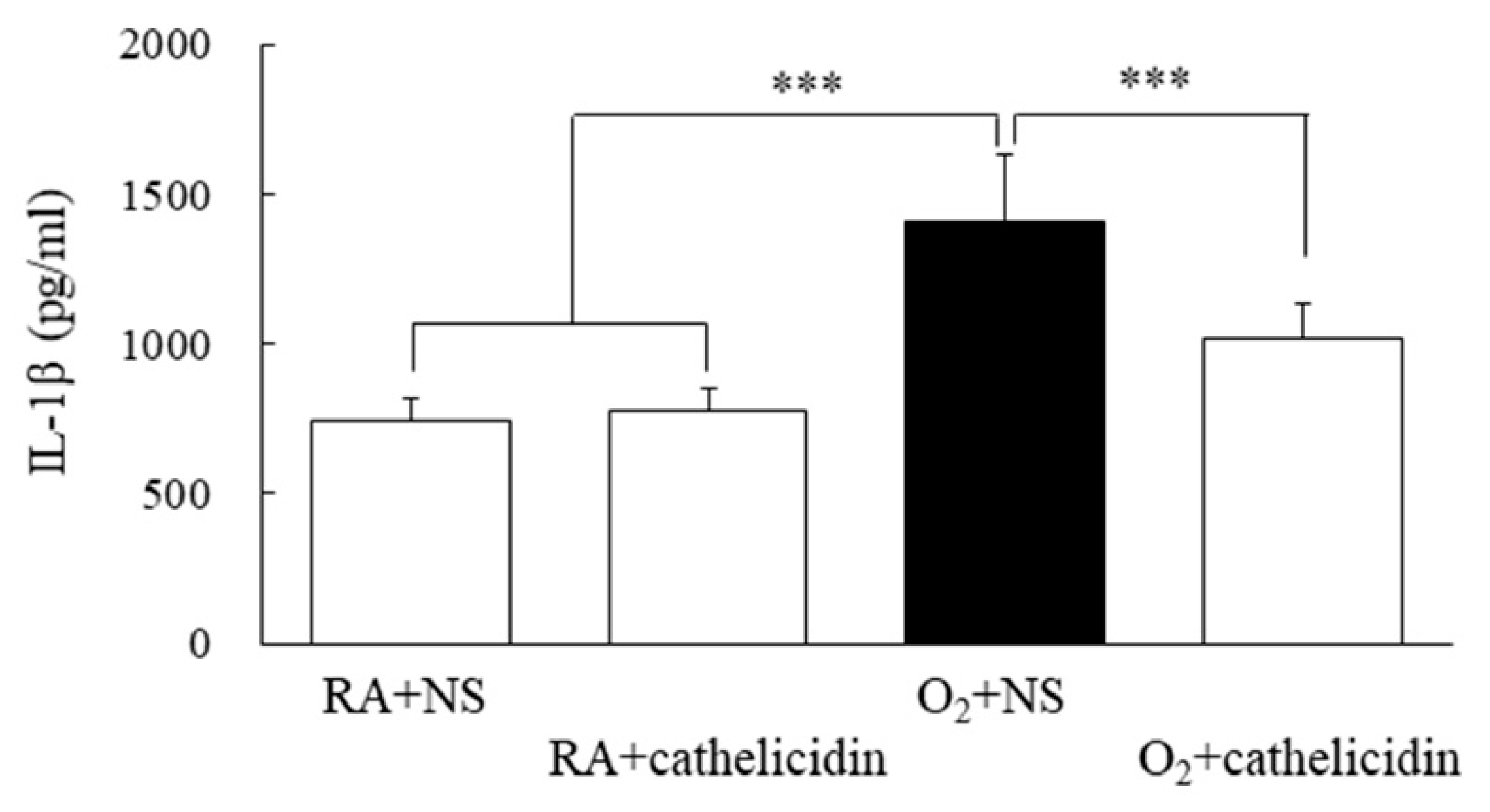
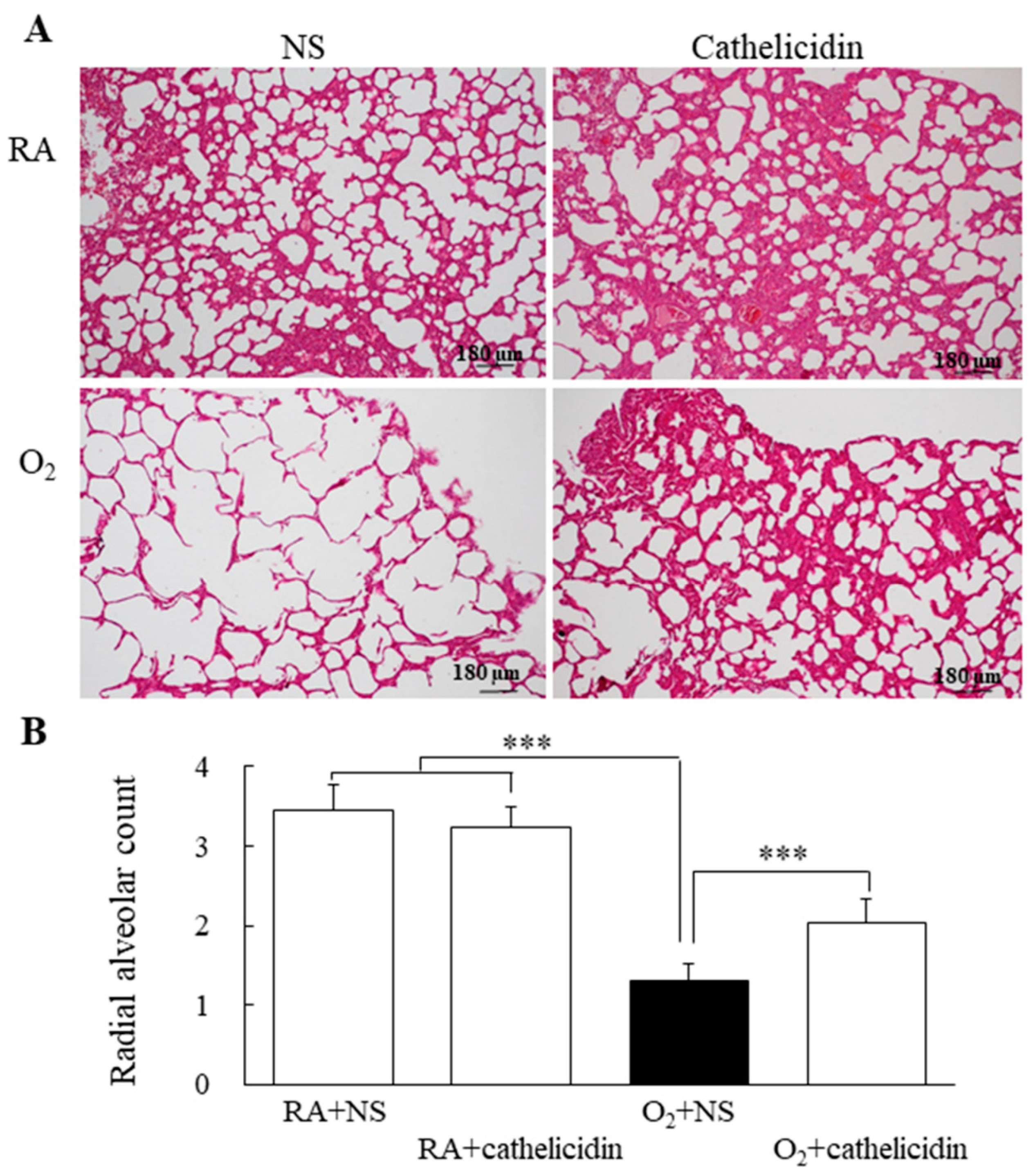
3.7. Cathelicidin Decreased Lung Cytokine and Improved Alveolarization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramanathan, R. Surfactant therapy in preterm infants with respiratory distress syndrome and in near-term or term newborns with acute RDS. J. Perinatol. 2006, 26, S51–S56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayton, A.; Timoney, P.; Vargo, L.; Perez, J.A. A review of oxygen physiology and appropriate management of oxygen levels in premature neonates. Adv. Neonatal Care 2018, 18, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Manji, J.S.; O’Kelly, C.J.; Leung, W.I.; Olson, D.M. Timing of hyperoxic exposure during alveolarization influences damage mediated by leukotrienes. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 281, L799–L806. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.M.; Wang, L.F.; Chou, H.C.; Lan, Y.D.; Lai, Y.P. Up-regulation of connective tissue growth factor in hyperoxia-induced lung fibrosis. Pediatr. Res. 2007, 62, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gien, J.; Kinsella, J.P. Pathogenesis and treatment of bronchopulmonary dysplasia. Curr. Opin. Pediatr. 2011, 23, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Mouradian, G.C., Jr.; Alvarez-Argote, S.; Gorzek, R.; Thuku, G.; Michkalkiewicz, T.; Wong-Riley, M.T.T.; Konduri, G.G.; Hodges, M.R. Acute and chronic changes in the control of breathing in a rat model of bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L506–L518. [Google Scholar] [CrossRef]
- Chen, C.M.; Hwang, J.; Chou, H.C. Maternal Tn immunization attenuates hyperoxia-induced lung injury in neonatal rats through suppression of oxidative stress and inflammation. Front. Immunol. 2019, 10, 681. [Google Scholar] [CrossRef] [Green Version]
- Saugstad, O.D. Update on oxygen radical disease in neonatology. Curr. Opin. Obstet. Gynecol. 2001, 13, 147–153. [Google Scholar] [CrossRef]
- Dietz, R.M.; Wright, C.J. Oxidative stress diseases unique to the perinatal period: A window into the developing innate immune response. Am. J. Reprod. Immunol. 2018, 79, e12787. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, W.; Yu, J.; Li, S.; Lin, L.; Chen, X. Induction of ferroptosis and mitochondrial dysfunction by oxidative stress in PC12 cells. Sci. Rep. 2018, 8, 574. [Google Scholar] [CrossRef]
- Doss, M.; White, M.R.; Tecle, T.; Hartshorn, K.L. Human defensins and LL-37 in mucosal immunity. J. Leukoc. Biol. 2010, 87, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.P.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, H.C.; Chen, C.M. Hyperoxia induces ferroptosis and impairs lung development in neonatal mice. Antioxidants 2022, 11, 641. [Google Scholar] [CrossRef]
- Jiang, J.S.; Chou, H.C.; Chen, C.M. Cathelicidin attenuates hyperoxia-induced lung injury by inhibiting oxidative stress in newborn rats. Free Radic. Biol. Med. 2020, 150, 23–29. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Pan, Q.; Zhang, W.; Xiang, J.; Jiang, X. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Martín-Gari, M.; Ayala, V.; Pamplona, R. The advanced lipoxidation end-product malondialdehyde-lysine in aging and longevity. Antioxidants 2020, 9, 1132. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Margison, K.; Pratt, D.A. The potency of diarylamine radical-trapping antioxidants as inhibitors of ferroptosis underscores the role of autoxidation in the mechanism of cell death. ACS Chem. Biol. 2017, 12, 2538–2545. [Google Scholar] [CrossRef]
- Cooney, T.P.; Thurlbeck, W.M. The radial alveolar count method of Emery and Mithal: A reappraisal 2—Intrauterine and early postnatal lung growth. Thorax 1982, 37, 580–583. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, M.; Thébaud, B. Animal models of bronchopulmonary dysplasia. The term rat models. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L948–L958. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Murakami, T.; Kuwahara-Arai, K.; Tamura, H.; Hiramatsu, K.; Nagaoka, I. Human anti-microbial cathelicidin peptide LL-37 suppresses the LPS-induced apoptosis of endothelial cells. Int. Immunol. 2011, 23, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alva, R.; Mirza, M.; Baiton, A.; Lazuran, L.; Samokysh, L.; Bobinski, A.; Cowan, C.; Jaimon, A.; Obioru, D.; Al Makhoul, T.; et al. Oxygen toxicity: Cellular mechanisms in normobaric hyperoxia. Cell Biol. Toxicol. 2022, 16, 1–33. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, P.; Zhai, B.; Zhang, M.; Xiang, Y.; Fang, J.; Xu, S.; Gao, Y.; Chen, X.; Sui, X.; et al. The emerging role of ferroptosis in inflammation. Biomed. Pharmacother. 2020, 127, 110108. [Google Scholar] [CrossRef]
- Li, J.Y.; Yao, Y.M.; Tian, Y.P. Ferroptosis: A trigger of proinflammatory state progression to immunogenicity in necroinflammatory disease. Front. Immunol. 2021, 12, 701163. [Google Scholar] [CrossRef]
- Linkermann, A.; Stockwell, B.R.; Krautwald, S.; Anders, H.J. Regulated cell death and inflammation: An auto-amplification loop causes organ failure. Nat. Rev. Immunol. 2014, 14, 759–767. [Google Scholar] [CrossRef]
- Beyer, A.M.; Norwood Toro, L.E.; Hughes, W.E.; Young, M.; Clough, A.V.; Gao, F.; Medhora, M.; Audi, S.H.; Jacobs, E.R. Autophagy, TERT, and mitochondrial dysfunction in hyperoxia. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H985–H1003. [Google Scholar] [CrossRef]
- Ratner, V.; Starkov, A.; Matsiukevich, D.; Polin, R.A.; Ten, V.S. Mitochondrial dysfunction contributes to alveolar developmental arrest in hyperoxia-exposed mice. Am. J. Respir. Cell Mol. Biol. 2009, 40, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, J.; Rezonzew, G.; Jilling, T.; Ballinger, S.; Ambalavanan, N. Mitochondrial DNA variation modulates alveolar development in newborn mice exposed to hyperoxia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 317, L740–L747. [Google Scholar] [CrossRef]
- Piedboeuf, B.; Horowitz, S.; Johnston, C.J.; Gamache, M.; Bélanger, S.; Poubelle, P.E.; Welty, S.E.; Watkins, R.H. Interleukin-1 expression during hyperoxic lung injury in the mouse. Free Radic. Biol. Med. 1998, 24, 1446–1454. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, H.-C.; Chen, C.-M. Cathelicidin Attenuates Hyperoxia-Induced Lung Injury by Inhibiting Ferroptosis in Newborn Rats. Antioxidants 2022, 11, 2405. https://doi.org/10.3390/antiox11122405
Chou H-C, Chen C-M. Cathelicidin Attenuates Hyperoxia-Induced Lung Injury by Inhibiting Ferroptosis in Newborn Rats. Antioxidants. 2022; 11(12):2405. https://doi.org/10.3390/antiox11122405
Chicago/Turabian StyleChou, Hsiu-Chu, and Chung-Ming Chen. 2022. "Cathelicidin Attenuates Hyperoxia-Induced Lung Injury by Inhibiting Ferroptosis in Newborn Rats" Antioxidants 11, no. 12: 2405. https://doi.org/10.3390/antiox11122405