

Evaluation of the Chemical Composition, Antioxidant and Antidiabetic Activity of Rhaponticoides iconiensis Flowers: Effects on Key Enzymes Linked to Type 2 Diabetes In Vitro, In Silico and on Alloxan-Induced Diabetic Rats In Vivo
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Reagents
2.2. Extraction of the Plant Material
2.3. In Vitro Enzyme Inhibition Assays
2.4. In Vivo Antidiabetic Activity
2.4.1. Animals and Experimental Design
2.4.2. Alloxan-Induced Diabetes Model in the Rats
2.4.3. Oral Glucose Tolerance Test (OGTT)
2.5. The Total Phenolic (TPC) and Total Flavonoid (TFC) Contents
2.6. In Vitro Antioxidant Activity Assays
2.6.1. Assay for the DPPH Radical Scavenging Capacity
2.6.2. Assay for the ABTS Radical Scavenging Capacity
2.6.3. Ferric Reducing Ability Power (FRAP)
2.7. Phytochemical Analyses by LC-MS/MS
2.8. In Silico Studies
2.8.1. Molecular Docking
2.8.2. Molecular Dynamics Simulations
2.9. Statistical Analysis
3. Results and Discussion
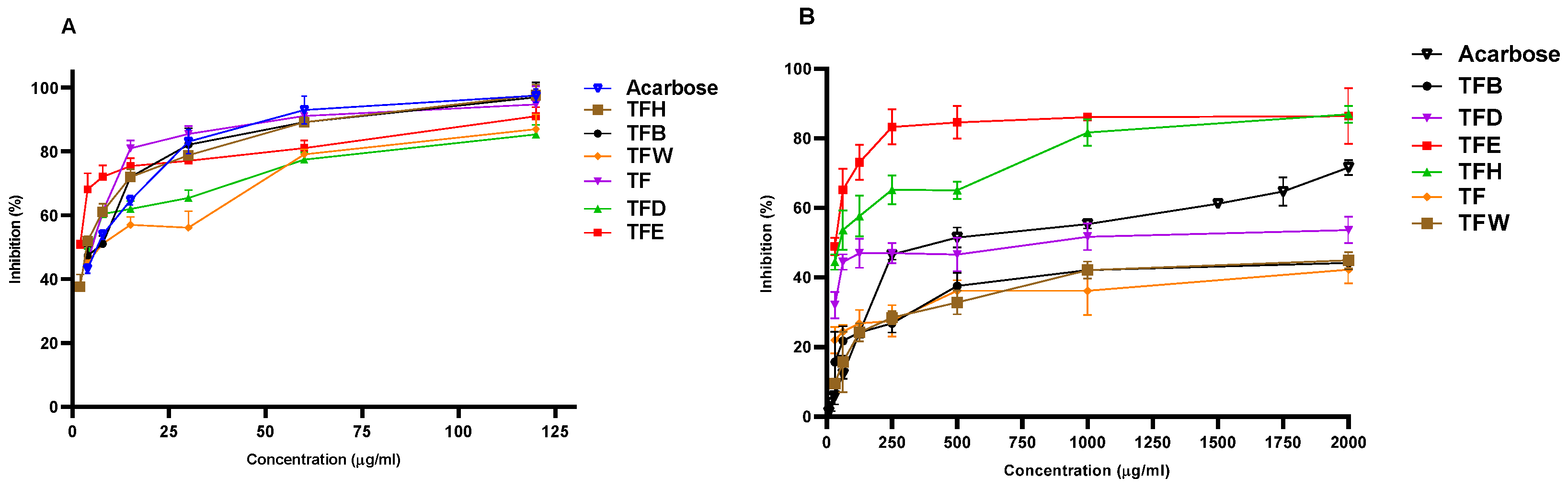
3.1. In Vitro Antidiabetic Activity
3.2. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
3.3. In Vitro Antioxidant Activity Results
3.4. In Vivo Antidiabetic Activity
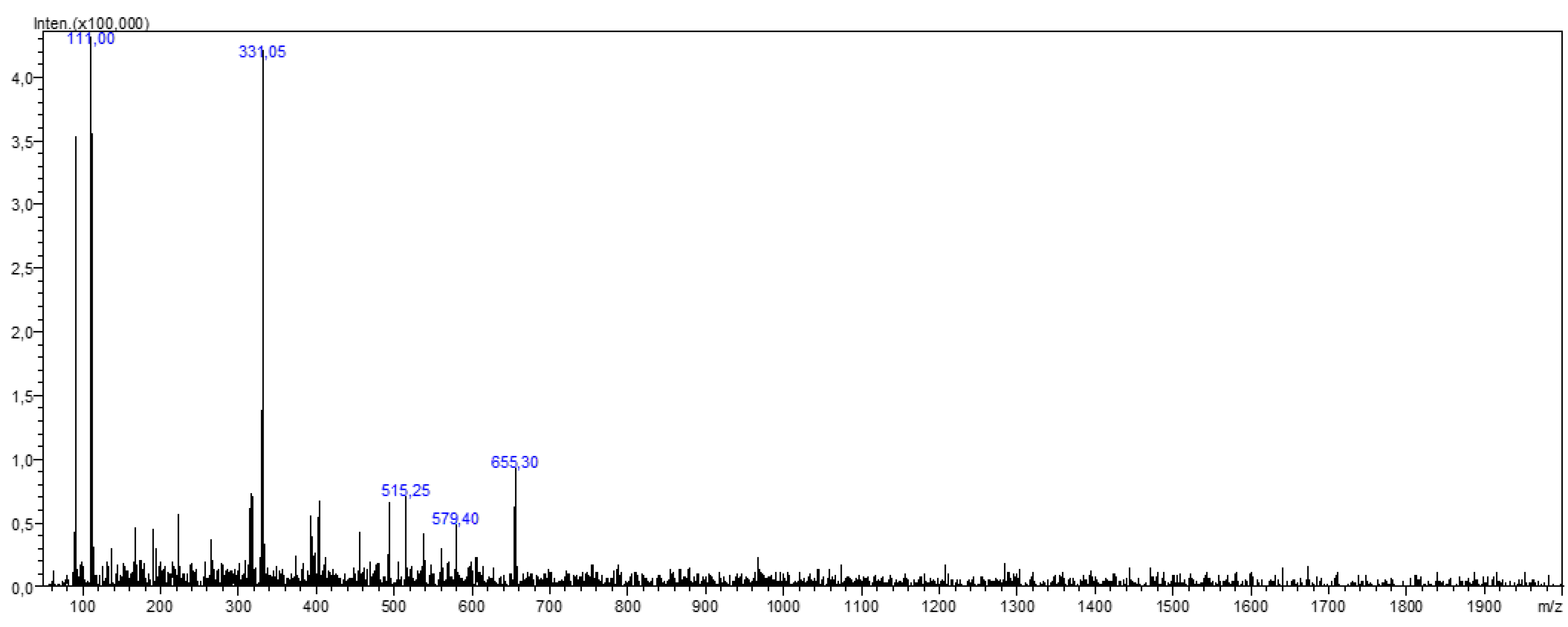
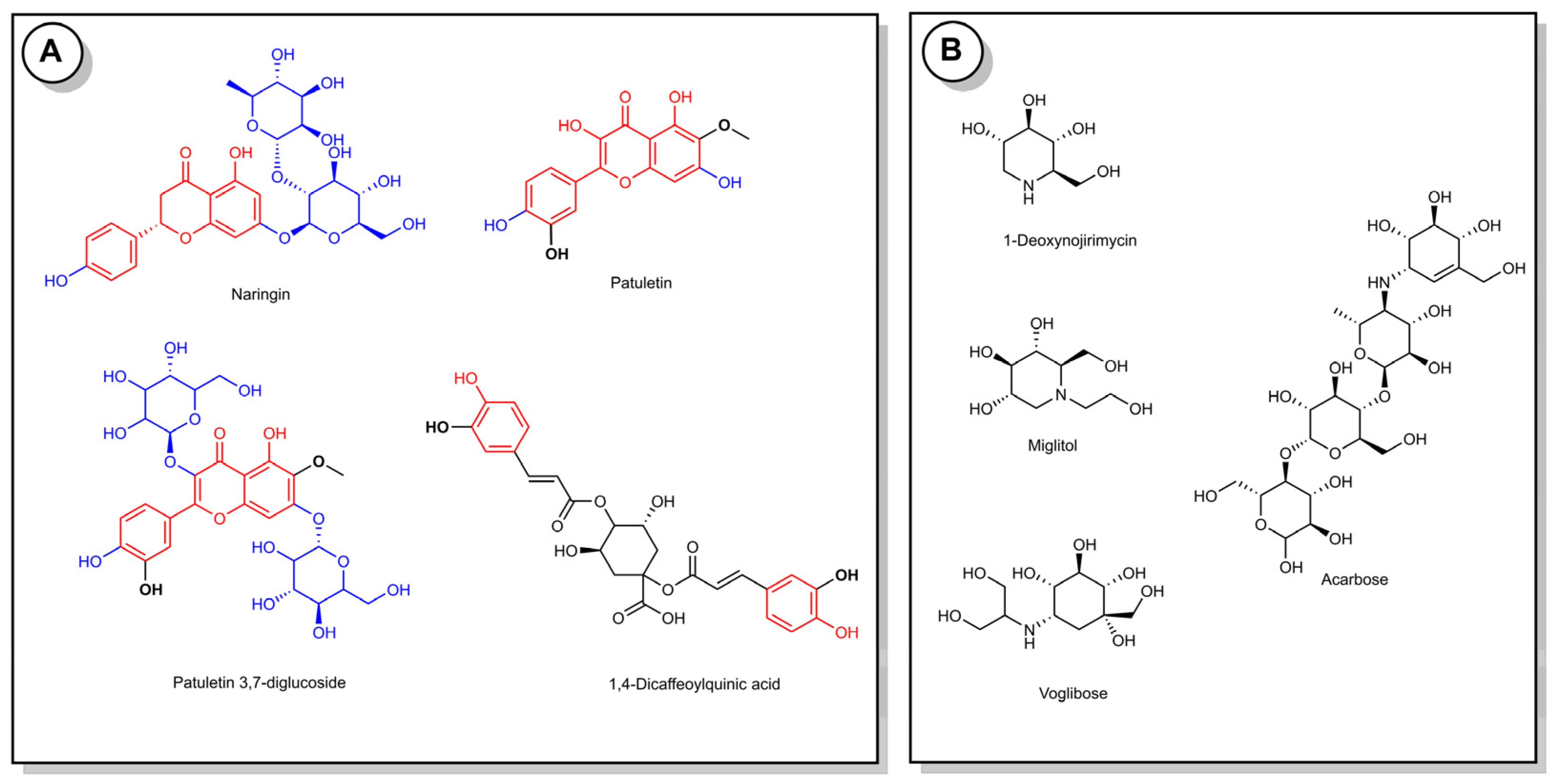
3.5. LC-MS/MS Results
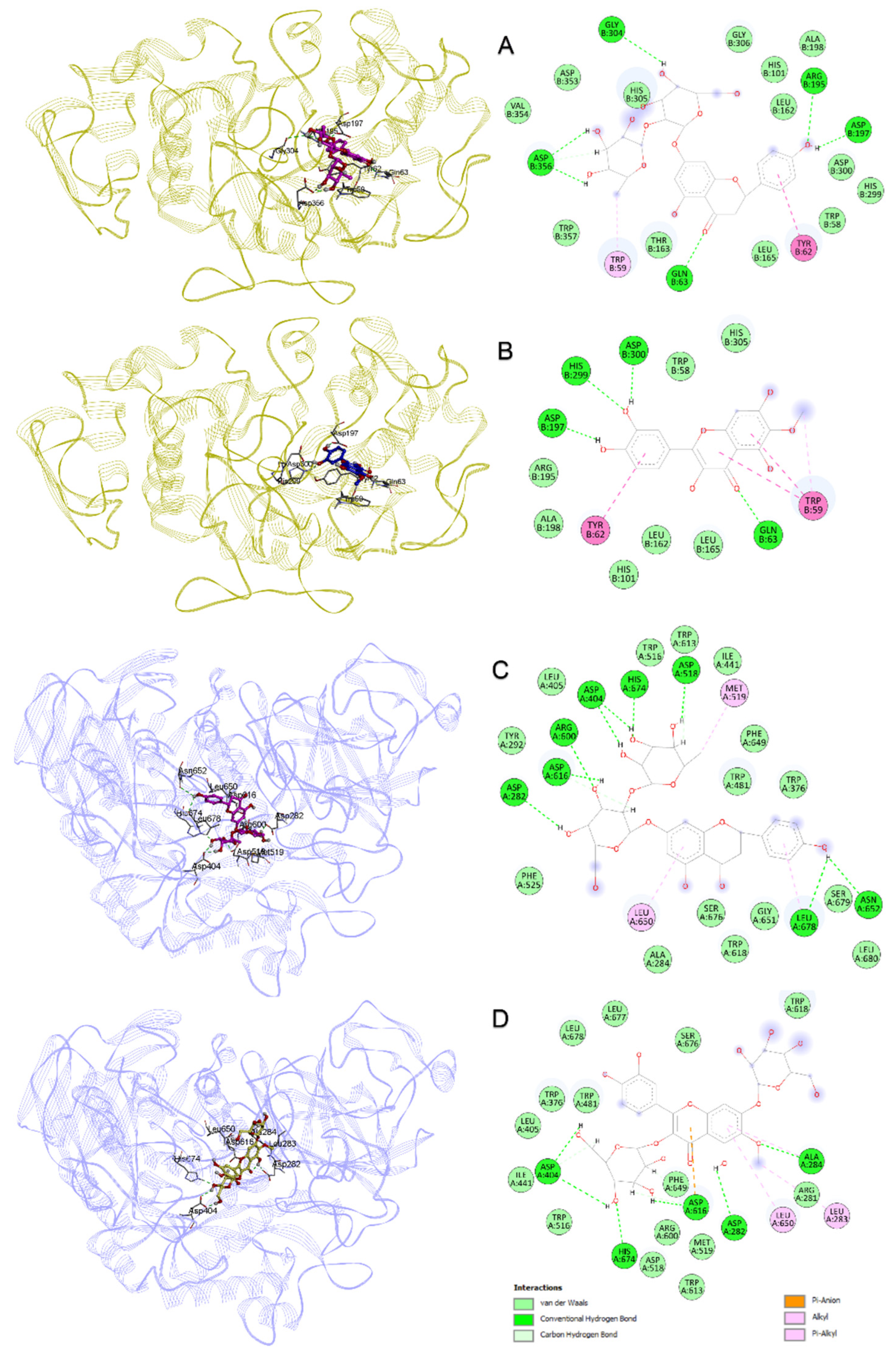
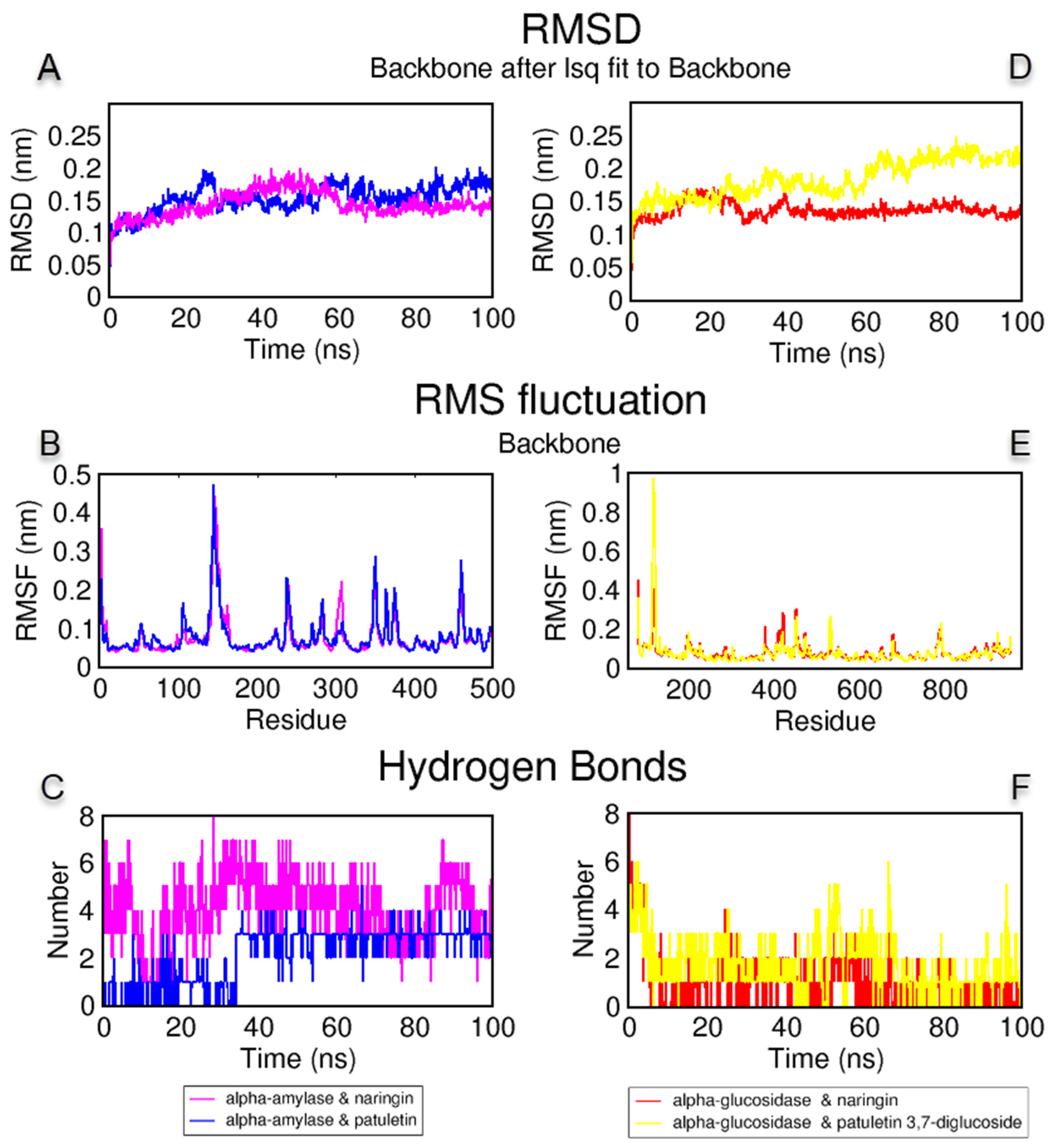
3.6. Molecular Docking and Dynamics Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, G.R.; Ignacimuthu, S.; Paulraj, M.G.; Sasikumar, P. Antihyperglycemic activity and antidiabetic effect of methyl caffeate isolated from Solanum torvum Swartz. fruit in streptozotocin induced diabetic rats. Eur. J. Pharmacol. 2011, 670, 623–631. [Google Scholar] [CrossRef]
- Chipiti, T. In Vitro Anti-Oxidative and Carbohydrates Digesting Enzymes Inhibitory Effects of Some Medicinal Plants Used for the Management of Diabetes in the Mrewa District, Zimbabwe. Ph.D. Thesis, University of KwaZulu-Natal (Westville Campus), Durban, South Africa, 2013. [Google Scholar]
- Kose, Y.B.; Iscan, G.; Goger, F.; Akalin, G.; Demirci, B.; Baser, K.H. Chemical Composition and Biological Activity of Centaurea baseri: New Species from Turkey. Chem. Biodivers. 2016, 13, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Radan, M.; Carev, I.; Tesevic, V.; Politeo, O.; Culic, V.C. Qualitative HPLC-DAD/ESI-TOF-MS Analysis, Cytotoxic, and Apoptotic Effects of Croatian Endemic Centaurea ragusina L. Aqueous Extracts. Chem Biodivers 2017, 14, e1700099. [Google Scholar] [CrossRef] [Green Version]
- Çinbilgel, İ.; Eren, Ö.; Duman, H. Rhaponticoides gokceoglui (Asteraceae), a striking new species from Turkey. Phytotaxa 2014, 170, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Güner, A.; Aslan, S. Türkiye Bitkileri Listesi: (Damarlı Bitkiler); Nezahat Gökyiǧit Botanik Bahçesi Yayınları: Istanbul, Türkiye, 2012. [Google Scholar]
- Tugay, O.; Paşayeva, L.; Demirpolat, E.; Şahin, M. Comparative evaluation of cytotoxicity and phytochemical composition of Centaurea iconiensis (Rhaponticoides iconiensis). Iran. J. Sci. Technol. Trans. A Sci. 2021, 45, 65–75. [Google Scholar] [CrossRef]
- Zheleva-Dimitrova, D.; Zengin, G.; Sinan, K.I.; Yıldıztugay, E.; Mahomoodally, M.F.; Ak, G.; Picot-Allain, M.C.N.; Gevrenova, R. Identification of bioactive compounds from Rhaponticoides iconiensis extracts and their bioactivities: An endemic plant to Turkey flora. J. Pharm. Biomed. Anal. 2020, 190, 113537. [Google Scholar] [CrossRef]
- Paşayeva, L.; İnce, U.; Fatullayev, H.; Ceylan, C.; Tugay, O. Hypoglycemic, antioxidant, antimicrobial activities and phytochemical analyses of Rhaponticoides iconiensis leafy stem extracts. Vegetos 2021, 34, 592–599. [Google Scholar] [CrossRef]
- Paşayeva, L.; Özalp, B.; Fatullayev, H. Potential enzyme inhibitory properties of extracts and fractions from fruit latex of Ficus carica-based on inhibition of α-amylase and α-glucosidase. J. Food Meas. Charact. 2020, 14, 2819–2827. [Google Scholar] [CrossRef]
- Sornalakshmi, V.; Tresina Soris, P.; Paulpriya, K.; Packia Lincy, M.; Mohan, V. Oral glucose tolerance test (OGTT) in normal control and glucose induced hyperglycemic rats with Hedyotis leschenaultiana DC. Group 2016, 1, 59–62. [Google Scholar]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolics and total flavonoids in Bulgarian fruits and vegetables. J. Univ. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Gyamfi, M.A.; Aniya, Y. Antioxidant properties of Thonningianin A, isolated from the African medicinal herb, Thonningia sanguinea. Biochem. Pharmacol. 2002, 63, 1725–1737. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Guo, C.J.; Yang, J.J.; Wei, J.Y.; Li, Y.F.; Xu, J.; Jiang, Y.G. Antioxidant activities of peel, pulp and seed fractions of common fruits as determined by FRAP assay. Nutr. Res. 2003, 23, 1719–1726. [Google Scholar] [CrossRef]
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of human lysosomal acid alpha-glucosidase-a guide for the treatment of Pompe disease. Nat. Commun. 2017, 8, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilles, C.; Astier, J.P.; Marchis-Mouren, G.; Cambillau, C.; Payan, F. Crystal structure of pig pancreatic alpha-amylase isoenzyme II, in complex with the carbohydrate inhibitor acarbose. Eur. J. Biochem. 1996, 238, 561–569. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Wu, C.; Ghoreishi, D.; Chen, W.; Wang, L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; Von Bargen, C.D.; et al. OPLS4: Improving Force Field Accuracy on Challenging Regimes of Chemical Space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Celik, I.; Sarialtin, S.Y.; Coban, T.; Kilcigil, G. Design, Synthesis, in Vitro and in Silico Studies of Benzimidazole-Linked Oxadiazole Derivatives as Anti-inflammatory Agents. Chemistryselect 2022, 7, e202201548. [Google Scholar] [CrossRef]
- Işık, A.; Acar Çevik, U.; Karayel, A.; Celik, I.; Erçetin, T.; Koçak, A.; Özkay, Y.; Kaplancıklı, Z. Synthesis and molecular modelling of thiadizole based hydrazone derivatives as acetylcholinesterase and butyrylcholinesterase inhibitory activities. SAR QSAR Environ. Res. 2022, 33, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmuller, H.; MacKerell, A.D., Jr. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paşayeva, L.; Eruygur, N.; Ayaz, F.; Tugay, O.; Fatullayev, H. Assessment of the antioxidant and enzyme inhibition activities of Cousinia iconica with focus on phytochemical investigation by LC-MS/MS. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 1178–1188. [Google Scholar] [CrossRef]
- WRoMS, D. Wiley Registry of Mass Spectral Data 11/NIST 17 Mass Spectrometer Database; Scientific Instrument Services Inc.: Ringoes, NJ, USA.
- Verspohl, E.J. Recommended testing in diabetes research. Planta Med. 2002, 68, 581–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubacey, T.M.; Haggag, E.G.; El-Toumy, S.A.; Ahmed, A.A.; El-Ashmawy, I.M.; Youns, M.M. Biological activity and flavonoids from Centaurea alexanderina leaf extract. J. Pharm. Res 2012, 5, 3352–3361. [Google Scholar]
- Ahmed, O.M.; Mahmoud, A.M.; Abdel-Moneim, A.; Ashour, M.B. Antidiabetic effects of hesperidin and naringin in type 2 diabetic rats. Diabetol. Croat. 2012, 41, 53–67. [Google Scholar]
- Ahmed, O.M.; Hassan, M.A.; Abdel-Twab, S.M.; Azeem, M.N.A. Navel orange peel hydroethanolic extract, naringin and naringenin have anti-diabetic potentials in type 2 diabetic rats. Biomed. Pharmacother. 2017, 94, 197–205. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ahmed, O.M.; Ashour, M.B.; Abdel-Moneim, A. In vivo and in vitro antidiabetic effects of citrus flavonoids; a study on the mechanism of action. Int. J. Diabetes Dev. Ctries. 2015, 35, 250–263. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, Y.; Han, K.; Park, H.; Lee, J.; Chung, S. Antidiabetic effect and mechanisms of DKY in KKAy mice. In Proceedings of the PSK Conference, Seoul, Republic of Korea, 20 April 2001; p. 130.2. [Google Scholar]
- Sahnoun, M.; Trabelsi, S.; Bejar, S. Citrus flavonoids collectively dominate the α-amylase and α-glucosidase inhibitions. Biologia 2017, 72, 764–773. [Google Scholar] [CrossRef]
- Asraoui, F.; Kounnoun, A.; Cacciola, F.; El Mansouri, F.; Kabach, I.; Oulad El Majdoub, Y.; Alibrando, F.; Arena, K.; Trovato, E.; Mondello, L. Phytochemical profile, antioxidant capacity, α-amylase and α-glucosidase inhibitory potential of wild Moroccan inula viscosa (L.) aiton leaves. Molecules 2021, 26, 3134. [Google Scholar] [CrossRef]
- Gao, H.; Huang, Y.-N.; Gao, B.; Xu, P.-Y.; Inagaki, C.; Kawabata, J. α-Glucosidase inhibitory effect by the flower buds of Tussilago farfara L. Food Chem. 2008, 106, 1195–1201. [Google Scholar] [CrossRef]
- Jabeen, A.; Mesaik, M.A.; Simjee, S.U.; Bano, S.; Faizi, S. Anti-TNF-α and anti-arthritic effect of patuletin: A rare flavonoid from Tagetes patula. Int. Immunopharmacol. 2016, 36, 232–240. [Google Scholar] [CrossRef]
- Alvarado-Sansininea, J.J.; Sánchez-Sánchez, L.; López-Muñoz, H.; Escobar, M.L.; Flores-Guzmán, F.; Tavera-Hernández, R.; Jiménez-Estrada, M. Quercetagetin and patuletin: Antiproliferative, necrotic and apoptotic activity in tumor cell lines. Molecules 2018, 23, 2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junejo, J.A.; Zaman, K.; Rudrapal, M.; Celik, I.; Attah, E.I. Antidiabetic bioactive compounds from Tetrastigma angustifolia (Roxb.) Deb and Oxalis debilis Kunth.: Validation of ethnomedicinal claim by in vitro and in silico studies. S. Afr. J. Bot. 2021, 143, 164–175. [Google Scholar] [CrossRef]
- Karrouchi, K.; Celik, I.; Fettach, S.; Karthick, T.; Bougrin, K.; Radi, S.; Faouzi, M.E.; Ansar, M.; Renjith, R. Synthesis and investigations of reactive properties, photophysical properties and biological activities of a pyrazole-triazole hybrid molecule. J. Mol. Struct. 2022, 1265, 133363. [Google Scholar] [CrossRef]
- Yeşilçayır, E.; Çelik, İ.; Şen, H.T.; Gürpınar, S.S.; Eryılmaz, M.; Kilcigil, G. Novel Benzimidazole-Based Compounds as Antimicrobials: Synthesis, Molecular Docking, Molecular Dynamics and in silico ADME Profile Studies. Acta Chim. Slov. 2022, 69, 419–429. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Names | α-glucosidase Assay IC50 (µg/mL) | α-amylase Assay IC50 (µg/mL) |
|---|---|---|
| TF | 10.582 ± 0.470 a,d | - |
| TFH | 9.438 ± 0.599 b,a,d | 248.867 ± 0.547 a |
| TFD | 20.741 ± 0.089 c,g | 93.912 ± 0.048 b |
| TFE | 11.737 ± 0.823 d | 84.247 ± 0.721 c |
| TFB | 17.691 ± 0.587 e,g | - |
| TFW | 33.732 ± 0.940 f | - |
| Acarbose | 17.707 ± 0.569 g | 217.133 ± 0.001 d |
| Samples | TPC (mg GAE/gextract) | TFC (mg CA/gextract) | FRAP (mmol Fe2+/gextract) | DPPH (IC50) (mg/mL) | ABTS (µM Trolox/gextract) |
|---|---|---|---|---|---|
| TF | 70.002 ± 0.003 a | 23.500 ± 2.816 a | 26.000 ± 1.001 a | 0.331 ± 0.001 a | 0.284 ± 0.001 a |
| TFH | 26.100 ± 1.054 b,f | 16.647 ± 0.905 b | 2741.123 ± 1.152 b | 0.756 ± 0.003 b | 0.137 ± 0.002 b,f |
| TFD | 107.333 ± 2.065 c | 58.300 ± 1.539 c | 236.589 ± 0.045 c | 0.654 ± 0.004 c | 0.482 ± 0.003 c |
| TFE | 292.333 ± 2.082 d | 204.667 ± 1.528 d | 3112.052 ± 2.023 d | 0.126 ± 0.002 d | 0.608 ± 0.005 d,g |
| TFB | 161.001 ± 1.002 e | 46.633 ± 2.801 e,f | 1807.333 ± 2.517 e | 0.241 ± 0.001 e | 0.431 ± 0.001 e |
| TFW | 26.647 ± 0.905 f | 47.333 ± 2.003 f | 328.467 ± 0.945 f | 0.540 ± 0.005 f | 0.132 ± 0.002 f |
| Trolox | - | - | 7911.014 ± 1.468 g | - | - |
| BHA | - | - | - | 0.057 ± 0.001 g | 0.620 ± 0.001 g |
| Blood Glucose Levels (mg/dL) | ||||||
|---|---|---|---|---|---|---|
| Treatment | Dose | 0 min (Fasted) | 30 min | 60 min | 90 min | 120 min |
| Saline | - | 119.3 ± 2.6 | 177.7 ± 4.3 | 108.7 ± 2.0 | 114.7 ± 2.9 | 113.3 ± 2.0 |
| Sub-extract | 300 mg/kg | 117.5 ± 6.5 | 144.0 ± 12.4 * | 148.0 ± 11.3 ** | 121.0 ± 4.5 | 118.8 ± 3.4 |
| Sub-extract | 600 mg/kg | 106.0 ± 5.0 | 131.5 ± 5.0 *** | 149.5 ± 5.5 ** | 128.0 ± 8.1 | 105.5 ± 4.0 |
| Acarbose | 5 mg/kg | 100.8 ± 9.6 | 137.0 ± 4.6 ** | 112.0 ± 14.2 | 127.8 ± 11.4 | 110.0 ± 2.9 |
| Blood Glucose Levels (mg/dL) | ||||||
|---|---|---|---|---|---|---|
| Treatment | Dose | 0 min (Fasted) | 30 min | 60 min | 90 min | 120 min |
| Alloxan | 100 mg/kg | 423.3 ± 14.5 | 529.7 ± 2.4 | 554.3 ± 14.3 | 561.3 ± 9.2 | 584.0 ± 8.3 |
| Sub-extract | 300 mg/kg | 455.3 ± 10.1 | 480.0 ± 11.6 | 516.6 ± 44.1 | 513.3 ± 27.3 | 476.7 ± 12.0 * |
| Sub-extract | 600 mg/kg | 444.3 ± 6.3 | 448.4 ± 14.1 ** | 463.7 ± 23.4 ** | 480.0 ± 30.5 ** | 420.2 ± 17.4 *** |
| Acarbose | 5 mg/kg | 466.6 ± 8.8 | 461.0 ± 4.9 * | 443.4 ± 3.5 *** | 380.6 ± 18.3 *** | 371.6 ± 4.4 *** |
| [M-H]− (m/z) | MS/MS (m/z) | Compounds | Reference |
|---|---|---|---|
| 331 | 269, 183, 159, 151, 149, 117, 107, 83, 65 | Patuletin | [7] |
| 655 | 493, 331 | Patuletin 3,7-diglucoside | [7] |
| 579 | 271, 151 | Naringin | [27] |
| 515 | 353, 203, 191, 179, 173 | 1,4-dicaffeoylquinic acid | [8,28] |
| α-Amylase (PDB ID: 1B2Y) | α-Glucosidase (PDB ID: 5NN8) | ||||||
|---|---|---|---|---|---|---|---|
| Compounds Name | PubChem ID | Docking Score | Protein–Ligand Interactions | Docking Score | Protein–Ligand Interactions | ||
| Hydrogen Bond | Hydrophobic | Hydrogen Bond | Hydrophobic | ||||
| Naringin | 442428 | −7.635 | Gln63 (2.15 Å), Arg195 (2.98 Å), Gly304 (2.83 Å), Asp356 (2.16 Å), Asp356 (1.77 Å and 2.56 Å), Asp197 (1.65 Å) | Trp59, Tyr62, Asp356 | −6.930 | Arg600 (2.10 Å), His674 (1.65 Å), Asp616 (1.61 Å), Asp282 (2.27 Å), Asp404 (2.11 Å and 1.84 Å), Asp51 (2.04 Å), Asn652 (2.82 Å), Leu678 (2.77 Å) | Leu650, Met519, Leu678 |
| Patuletin | 5281678 | −7.360 | Gln63 (2.09 Å), His299 (2.30 Å), Asp300 (1.72 Å), Asp197 (1.58 Å) | Trp59, Tyr62 | −5.274 | His674 (1.86 Å), Asp282 (1.76 Å), Asp616 (1.89 Å), Asp404 (2.04 Å) | Asp518, Asp616, Trp376, Phe649, Trp481, Phe525 |
| Patuletin 3,7-diglucoside | 44259785 | −7.227 | Arg195 (2.28 Å), Lys200 (2.02 Å), Lys200 (2.84 Å), Asp197 (1.54 Å), Asp300 (1.80 Å), Asp197 (1.86 Å), Gly306 (2.06 Å), His305 (2.00 Å) | Glu233, Asp300, Glu233, Glu240, Tyr151, Leu162, Ile235, Leu162 | −5.511 | Ala284 (2.32 Å), His674 (2.21 Å), Asp616 (1.72 Å), Asp404 (2.47 Å and 1.88 Å) and Asp282 (2.35 Å) | Asp404, Asp616, Leu283, Ala284, Leu650 |
| 1,4-Dicaffeoylquinic acid | 12358846 | −5.763 | Trp59 (2.64 Å), Gln63 (2.15 Å), Thr163 (2.95 Å) Lys200 (1.97 Å), Asp356 (1.62 Å), Asp356 (1.93 Å) | Ile235 | −4.832 | Ala284 (2.64 Å), Trp481 (2.39 Å), Gly651 (2.72 Å), Asp282 (1.95 Å), Asp518 (1.70 Å), Ser676 (1.89 Å), Asp404 (2.59 Å) | Leu283, Ser676, Asp282, Asp616, Arg600, Asp518, Leu650, Ala284 |
| Acarbose | 445421 | −8.480 | Gln63 (1.99 and 2.79 Å), Arg195 (2.62 and 2.19 Å), Lys200 (2.71 Å and 2.06 Å), His299 (2.99 Å), Glu240 (2.44 Å), Gly306 (1.97 Å), His201 (1.93 Å), Glu233 (1.91 Å), Asp300 (1.94 and 2.52 Å), His305 (2.02 Å and 2.61 Å), Trp59 (1.83 Å) | Glu233, Asp300, Asp197, Gly306, His201, Thr163, Leu165 | −6.130 | Trp481 (3.04 Å), Arg600 (2.21 Å), His674 (2.23 Å), Asp282 (2.00 Å), Asp282 (2.04 Å), Asp616 (1.86 Å), Asp404 (1.89 Å and 2.02 Å) | Asp282, Asp518, Asp616, Asp404, Trp376, Trp481, Phe649 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paşayeva, L.; Fatullayev, H.; Celik, I.; Unal, G.; Bozkurt, N.M.; Tugay, O.; Abdellattif, M.H. Evaluation of the Chemical Composition, Antioxidant and Antidiabetic Activity of Rhaponticoides iconiensis Flowers: Effects on Key Enzymes Linked to Type 2 Diabetes In Vitro, In Silico and on Alloxan-Induced Diabetic Rats In Vivo. Antioxidants 2022, 11, 2284. https://doi.org/10.3390/antiox11112284
Paşayeva L, Fatullayev H, Celik I, Unal G, Bozkurt NM, Tugay O, Abdellattif MH. Evaluation of the Chemical Composition, Antioxidant and Antidiabetic Activity of Rhaponticoides iconiensis Flowers: Effects on Key Enzymes Linked to Type 2 Diabetes In Vitro, In Silico and on Alloxan-Induced Diabetic Rats In Vivo. Antioxidants. 2022; 11(11):2284. https://doi.org/10.3390/antiox11112284
Chicago/Turabian StylePaşayeva, Leyla, Hanifa Fatullayev, Ismail Celik, Gokhan Unal, Nuh Mehmet Bozkurt, Osman Tugay, and Magda H. Abdellattif. 2022. "Evaluation of the Chemical Composition, Antioxidant and Antidiabetic Activity of Rhaponticoides iconiensis Flowers: Effects on Key Enzymes Linked to Type 2 Diabetes In Vitro, In Silico and on Alloxan-Induced Diabetic Rats In Vivo" Antioxidants 11, no. 11: 2284. https://doi.org/10.3390/antiox11112284