
Ishophloroglucin A Ameliorates VEGF-Induced Epithelial-Mesenchymal Transition via VEGFR2 Pathway Inhibition in Microgravity-Stimulated Human Retinal Pigment Epithelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
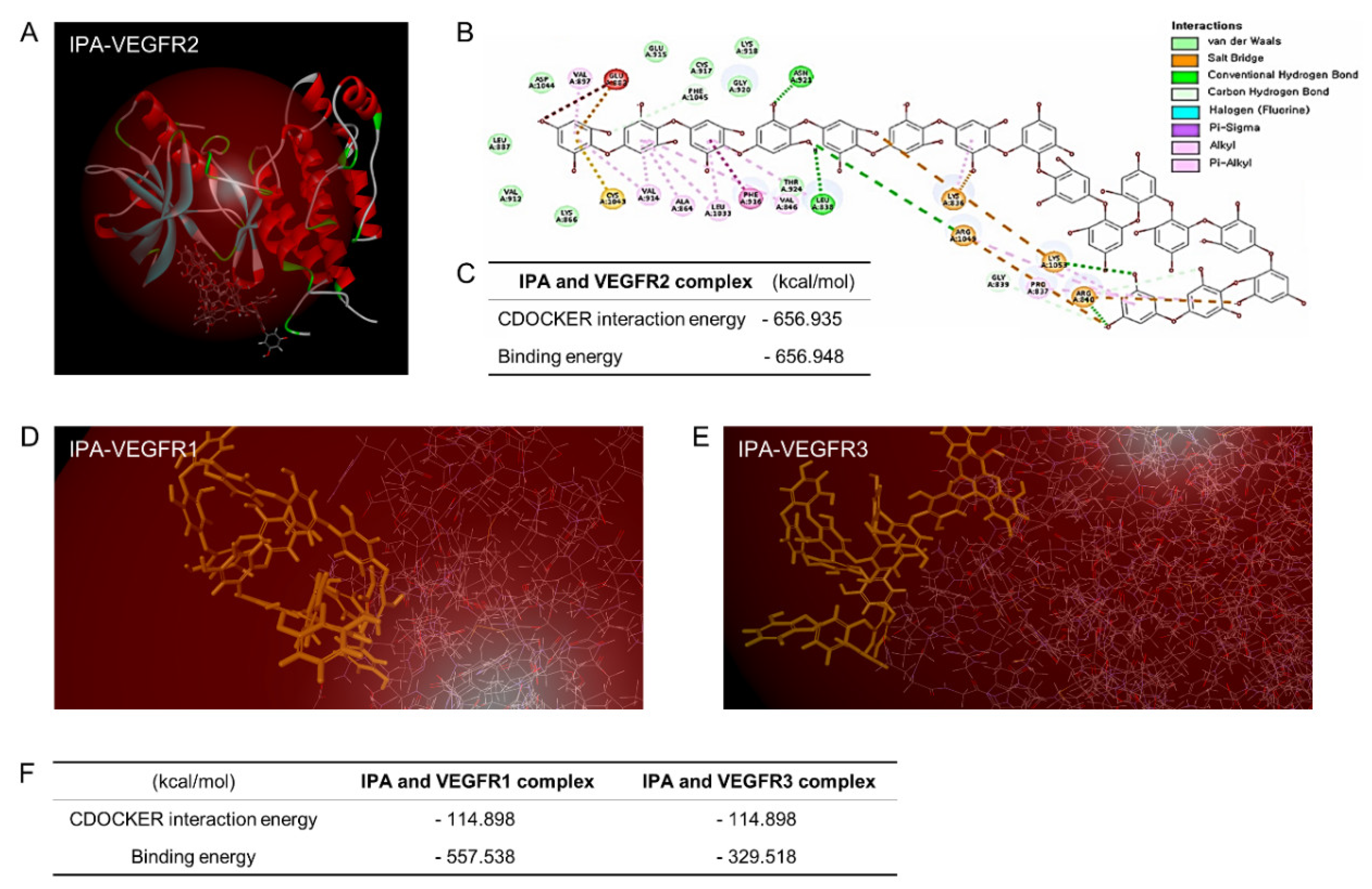
2.1. IPA Preparation and In Silico Binding Analysis of IPA-VEGFRs
2.2. Experimental Cell Model
2.2.1. ARPE19 Cell Culture
2.2.2. Clinostat Setup
2.3. RNA Extraction and cDNA Synthesis
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. Protein Extraction
2.6. Western Blotting
2.7. Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Scratch-Wound Migration Assay and Transwell Migration Assay
2.9. Statistical Analysis
3. Results
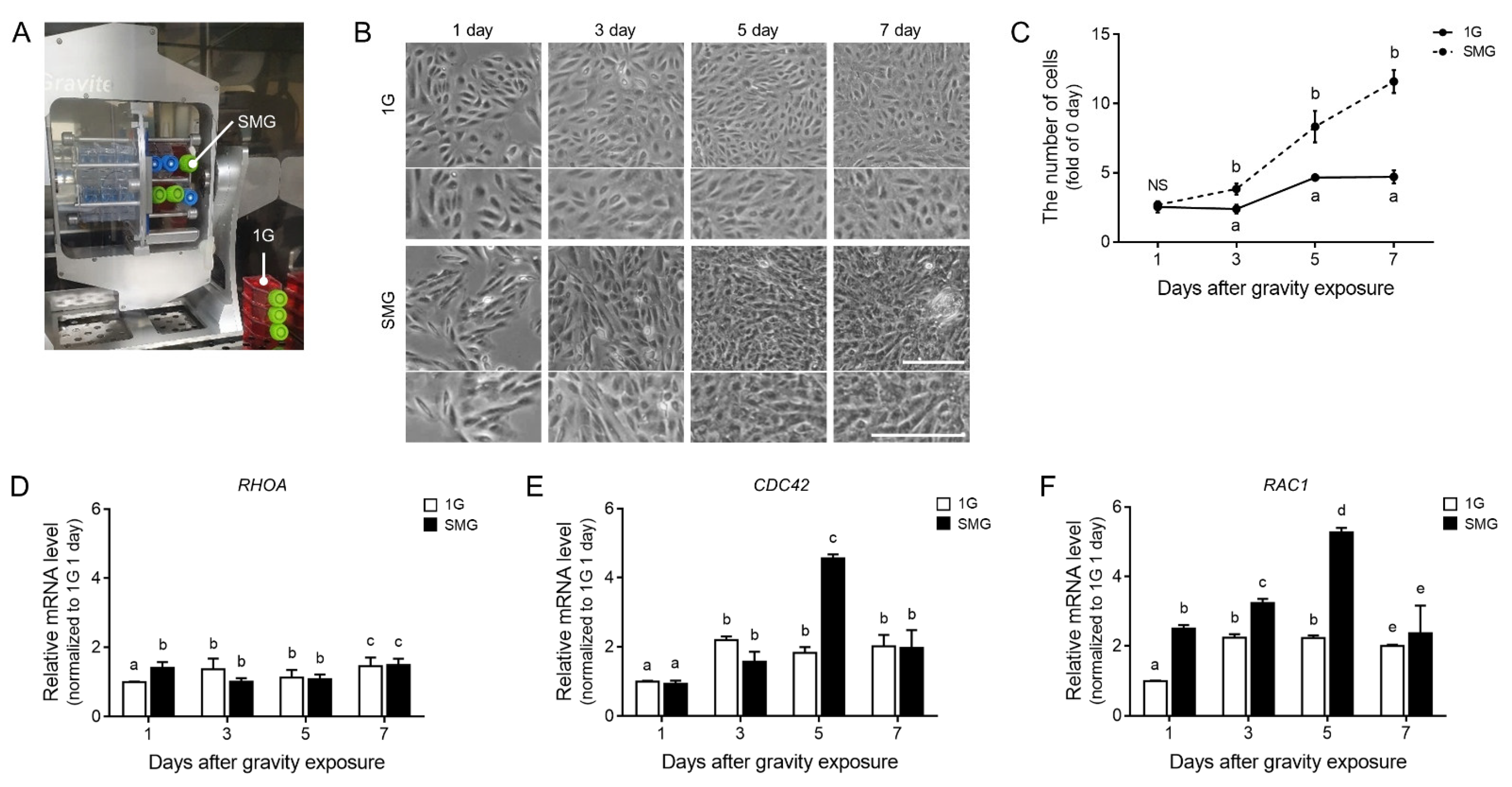
3.1. Actin Cytoskeleton and Cell Population Changed during Microgravity-Stimulated Development in ARPE19 Cells
3.2. VEGF Expression and EMT-Related Markers Increased during Microgravity-Stimulated Development in ARPE19 Cells
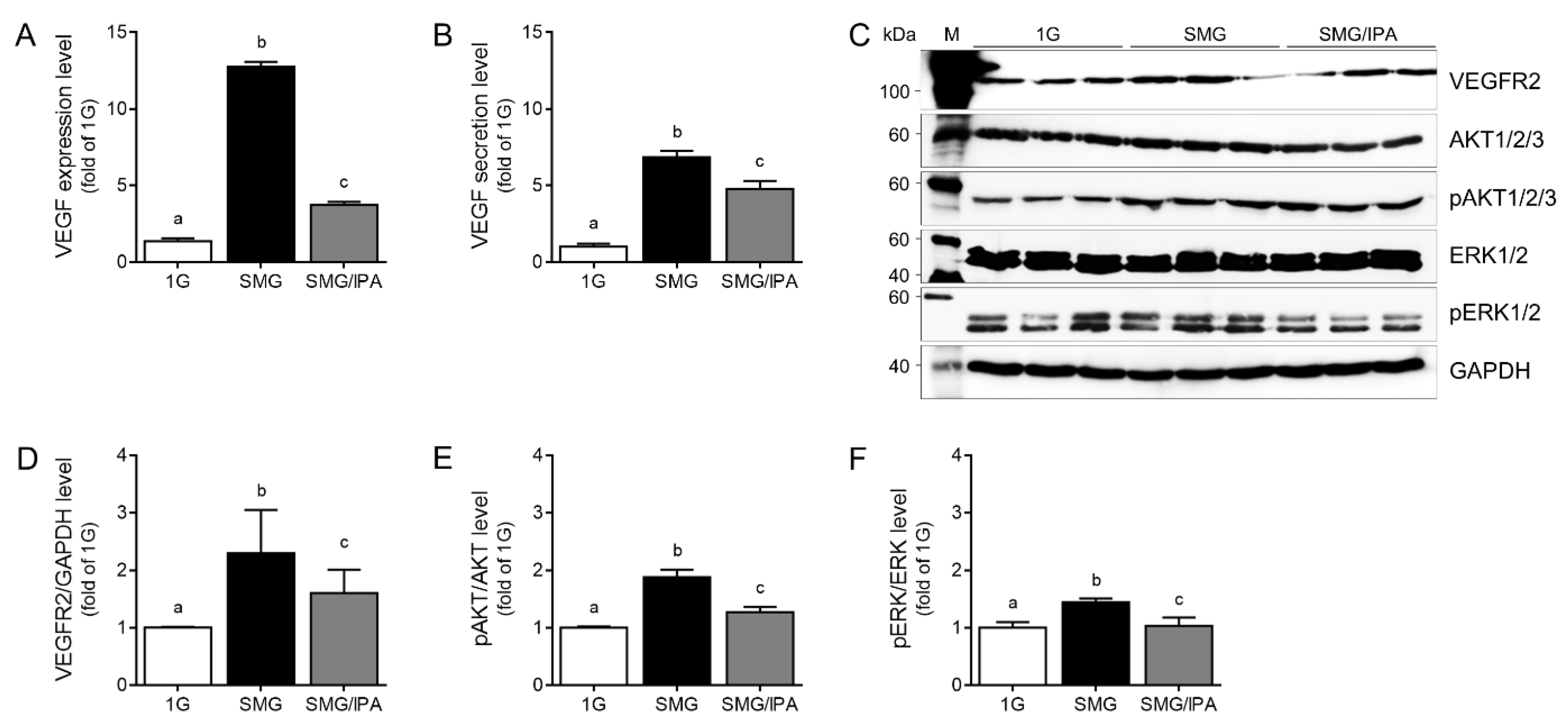
3.3. IPA Disrupted VEGFR2 Signaling Activation by Inhibiting VEGF–VEGFR2 Binding in Microgravity-Stimulated ARPE19 Cells
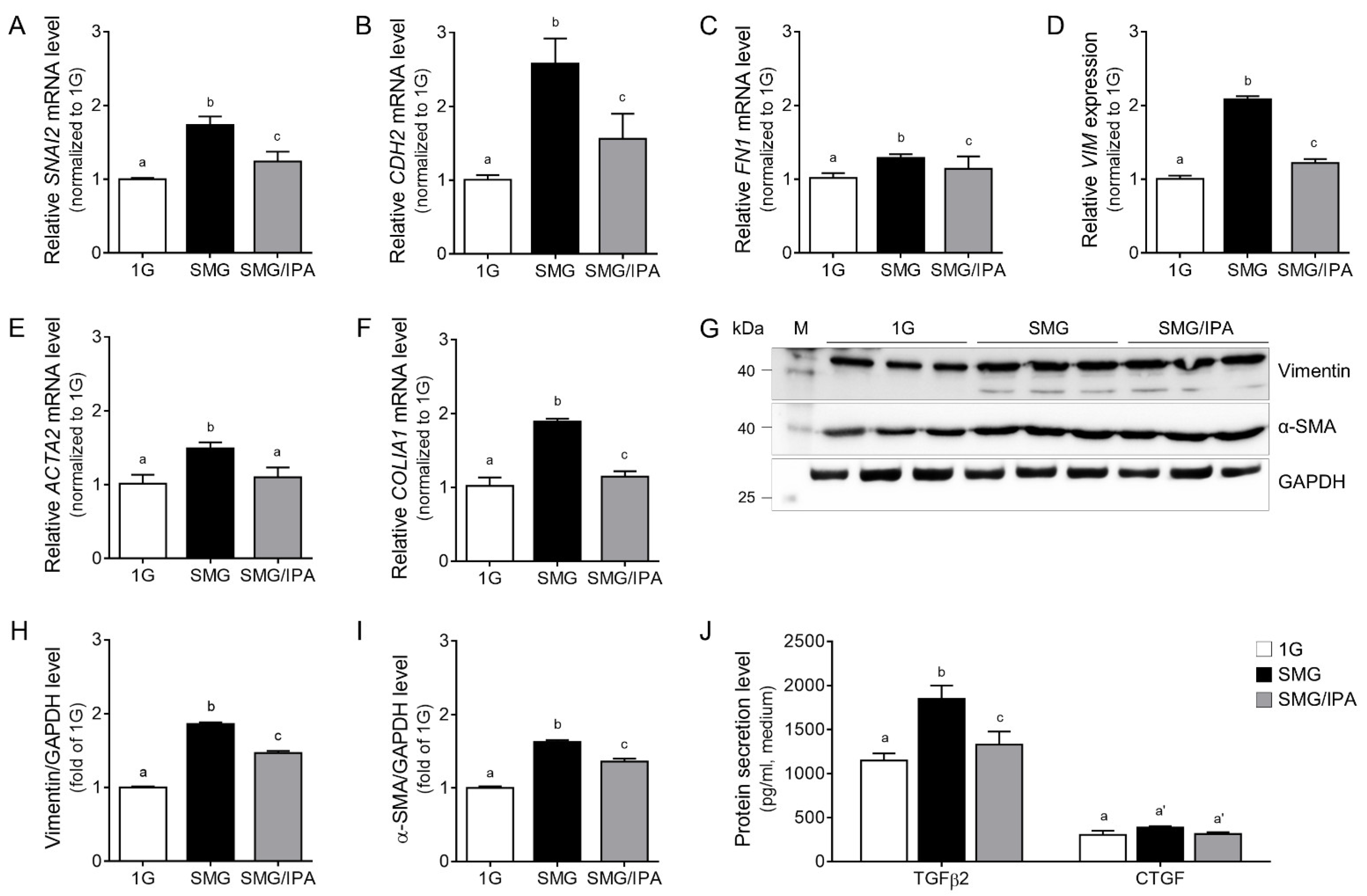
3.4. IPA Protected Microgravity-Induced EMT in ARPE19 Cells
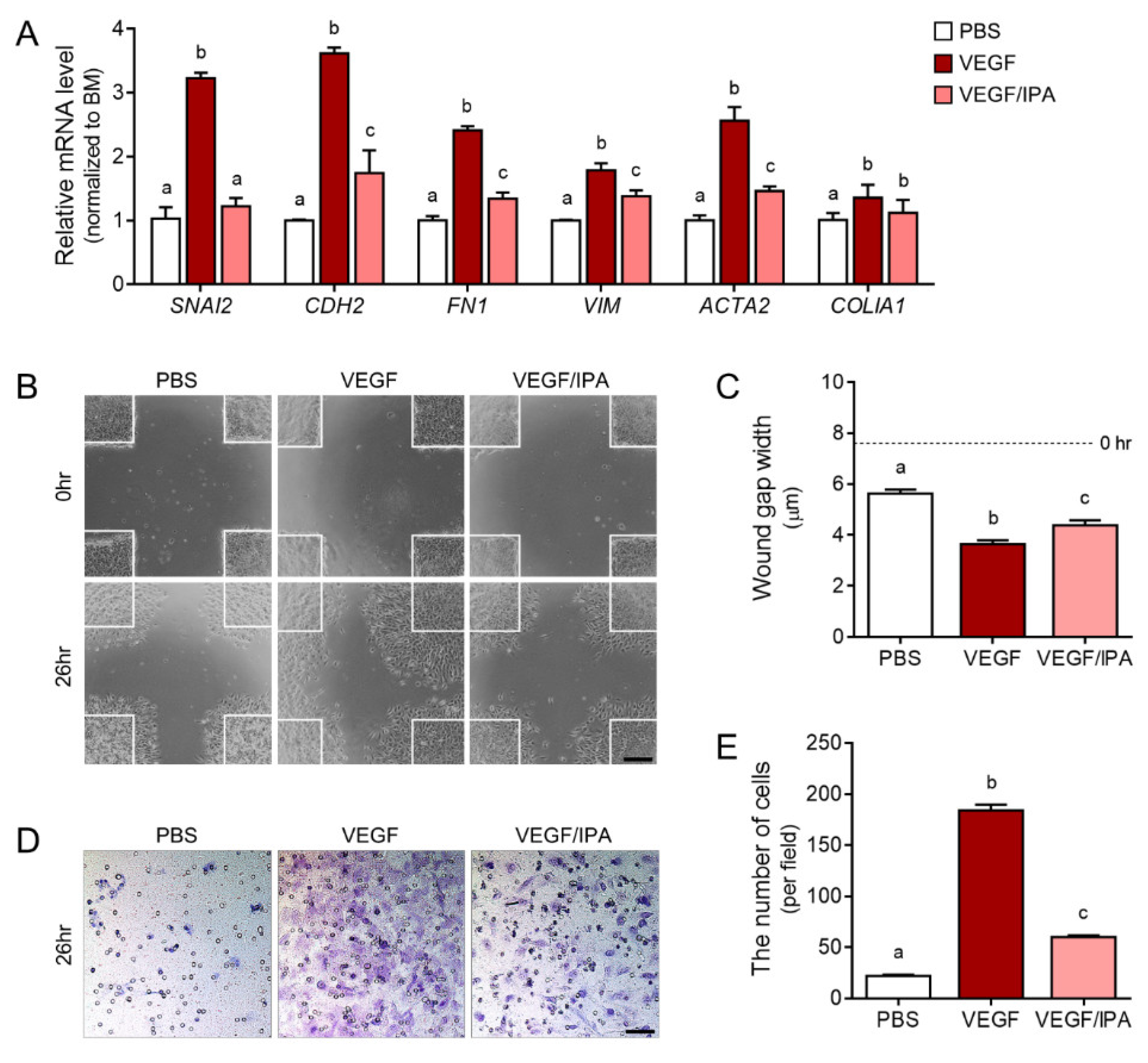
3.5. IPA Reduced Abnormal Migration and EMT in VEGF-Treated ARPE19 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keyak, J.H.; Koyama, A.K.; LeBlanc, A.; Lu, Y.; Lang, T.F. Reduction in proximal femoral strength due to long-duration spaceflight. Bone 2009, 44, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, R.; Genc, K.O.; Rice, A.J.; Lee, S.M.; Evans, H.J.; Maender, C.C.; Ilaslan, H.; Cavanagh, P.R. Muscle volume, strength, endurance, and exercise loads during 6-month missions in space. Aviat. Space Env. Med 2010, 81, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.E.; Stowe, R.P.; Mehta, S.K.; Yetman, D.L.; Leal, M.J.; Quiriarte, H.D.; Pierson, D.L.; Sams, C.F. Immune status, latent viral reactivation, and stress during long-duration head-down bed rest. Aviat. Space Env. Med. 2009, 80, A37–A44. [Google Scholar] [CrossRef]
- Nicogossian, A.E.; Parker, J.F. Space Physiology and Medicine. In NASA SP-447; NASA Special Publication: Washington, DC, USA, 1982; p. 447. [Google Scholar]
- Bock, O.; Weigelt, C.; Bloomberg, J.J. The cognitive demand of human sensorimotor performance during an extended space mission: A dual-task study. Aviat. Space Env. Med. 2010, 81, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.G.; Mader, T.H.; Gibson, C.R.; Tarver, W. Space flight–associated neuro-ocular syndrome. JAMA Ophthalmol. 2017, 135, 992–994. [Google Scholar] [CrossRef]
- Weiter, J.J.; Delori, F.C.; Wing, G.L.; Fitch, K.A. Retinal pigment epithelial lipofuscin and melanin and choroidal melanin in human eyes. Invest. Ophthalmol. Vis. Sci. 1986, 27, 145–152. [Google Scholar]
- Sparrrow, J.R.; Hicks, D.; Hamel, C.P. The retinal pigment epithelium in health and disease. Curr. Mol. Med. 2010, 10, 802–823. [Google Scholar] [CrossRef]
- Mannermaa, E.; Vellonen, K.S.; Urtti, A. Drug transport in corneal epithelium and blood–retina barrier: Emerging role of transporters in ocular pharmacokinetics. Adv. Drug Deliv. Rev. 2006, 58, 1136–1163. [Google Scholar] [CrossRef]
- Cunha-Vaz, J.; Bernardes, R.; Lobo, C. Blood-retinal barrier. Eur. J. Ophthalmol. 2011, 21 (Suppl. 6), S3–S9. [Google Scholar] [CrossRef]
- Kim, L.A.; D’Amore, P.A. A brief history of anti-VEGF for the treatment of ocular angiogenesis. Am J Pathol 2012, 181, 376–379. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Invest. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Neilson, E.G. Biomarkers for epithelial-mesenchymal transitions. J. Clin. Invest. 2009, 119, 1429–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Invest. 2009, 119, 1417–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, D.Y.; Lovicu, F.J. Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog. Retin. Eye Res. 2017, 60, 44–65. [Google Scholar] [CrossRef]
- Kaufhold, S.; Bonavida, B. Central role of Snail1 in the regulation of EMT and resistance in cancer: A target for therapeutic intervention. J. Exp. Clin. Cancer Res. 2014, 33, 62. [Google Scholar] [CrossRef]
- Saito, A. EMT and EndMT: Regulated in similar ways? J. Bio. 2013, 153, 493–495. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Shang, P.; Terasaki, H.; Stepicheva, N.; Hose, S.; Yazdankhah, M.; Weiss, J.; Sakamoto, T.; Bhutto, I.A.; Xia, S.; et al. A Role for βA3/A1-Crystallin in Type 2 EMT of RPE Cells Occurring in Dry Age-Related Macular Degeneration. Invest. Ophthalmol. Vis. Sci. 2018, 59, AMD104–AMD113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuttfeld, E.; Ballmer-Hofer, K. Structure and function of VEGF receptors. IUBMB Life 2009, 61, 915–922. [Google Scholar] [CrossRef]
- Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Biochem. Soc. Trans. 2003, 31, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yu, C.; Peng, S.; Xu, H.; Wright, E.; Zhang, X.; Huo, X.; Cheng, E.; Pham, T.H.; Asanuma, K.; et al. Autocrine VEGF signaling promotes proliferation of neoplastic Barrett’s epithelial cells through a PLC-dependent pathway. Gastroenterology 2014, 146, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Zhang, Y.; Peng, H.; Ke, Z.; Xu, L.; Su, T.; Tsung, A.; Tohme, S.; Huang, H.; Zhang, Q.; et al. Intracellular autocrine VEGF signaling promotes EBDC cell proliferation, which can be inhibited by Apatinib. Cancer Lett. 2016, 373, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Chen, Q.; Rao, W.; Zhang, R.; Wang, Y.; Ge, H.; Wei, Q. OVA66 promotes tumour angiogenesis and progression through enhancing autocrine VEGF-VEGFR2 signalling. EBioMedicine 2019, 41, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzaneh Behelgardi, M.; Zahri, S.; Gholami Shahvir, Z.; Mashayekhi, F.; Mirzanejad, L.; Asghari, S.M. Targeting signaling pathways of VEGFR1 and VEGFR2 as a potential target in the treatment of breast cancer. Mol. Biol. Rep. 2020, 47, 2061–2071. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Yang, E.C.; Coyer, J.A.; Zuccarello, G.C.; Wang, W.L.; Choi, C.G.; Boo, S.M. Phylogeography of the seaweed Ishige okamurae (Phaeophyceae): Evidence for glacial refugia in the northwest Pacific region. Mar. Biochem. 2012, 159, 1021–1028. [Google Scholar] [CrossRef]
- Fernando, K.H.N.; Yang, H.-W.; Jiang, Y.; Jeon, Y.-J.; Ryu, B. Ishige okamurae extract and its constituent ishophloroglucin an attenuated in vitro and in vivo high glucose-induced angiogenesis. Int. J. Mol. Sci. 2019, 20, 5542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, G.; Zhang, J.; Tang, P.; Yang, C.; Wang, G.; Chen, J.; Liu, J.; Zhang, L.; Ouyang, L. Discovery, Synthesis, and Evaluation of Highly Selective Vascular Endothelial Growth Factor Receptor 3 (VEGFR3) Inhibitor for the Potential Treatment of Metastatic Triple-Negative Breast Cancer. J. Med. Chem. 2021, 64, 12022–12048. [Google Scholar] [CrossRef]
- Ahmado, A.; Carr, A.J.; Vugler, A.A.; Semo, M.; Gias, C.; Lawrence, J.M.; Chen, L.L.; Chen, F.K.; Turowski, P.; da Cruz, L.; et al. Induction of differentiation by pyruvate and DMEM in the human retinal pigment epithelium cell line ARPE-19. Investig. Opthalmology Vis. Sci. 2011, 52, 7148–7159. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, T.; Tanimoto, K.; Fukazawa, T.; Imura, T.; Kawahara, Y.; Yuge, L. Simulated microgravity attenuates myogenic differentiation via epigenetic regulations. NPJ Microgravity 2018, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Manabe, T.; Matsumoto, M.; Kajiume, T.; Matsumoto, M.; Yuge, L. LIF-free embryonic stem cell culture in simulated microgravity. PLoS ONE 2009, 4, e6343. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhara, T.; Takeda, M.; Yamaguchi, S.; Manabe, T.; Matsumoto, M.; Kawahara, Y.; Yuge, L.; Kurisu, K. Simulated microgravity facilitates cell migration and neuroprotection after bone marrow stromal cell transplantation in spinal cord injury. Stem. Cell Res. Ther. 2013, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Louis, F.; Bouleftour, W.; Rattner, A.; Linossier, M.T.; Vico, L.; Guignandon, A. RhoGTPase stimulation is associated with strontium chloride treatment to counter simulated microgravity-induced changes in multipotent cell commitment. NPJ Microgravity 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Corydon, T.J.; Kopp, S.; Wehland, M.; Braun, M.; Schütte, A.; Mayer, T.; Hülsing, T.; Oltmann, H.; Schmitz, B.; Hemmersbach, R.; et al. Alterations of the cytoskeleton in human cells in space proved by life-cell imaging. Sci. Rep. 2016, 6, 20043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated microgravity inhibits cell focal adhesions leading to reduced melanoma cell proliferation and metastasis via FAK/RhoA-regulated mTORC1 and AMPK pathways. Sci. Rep. 2018, 8, 3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Nishigaki, N.; Washio, Y.; Kano, K.; Harris, P.A.; Sato, H.; Mori, I.; West, R.I.; Shibahara, M.; Toyoda, H.; et al. Discovery of novel benzimidazoles as potent inhibitors of TIE-2 and VEGFR-2 tyrosine kinase receptors. J. Med. Chem. 2007, 50, 4453–4470. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Tugues, S.; Li, X.; Gualandi, L.; Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Biochem. J. 2011, 437, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Byeon, S.H.; Lee, S.C.; Choi, S.H.; Lee, H.K.; Lee, J.H.; Chu, Y.K.; Kwon, O.W. Vascular Endothelial Growth Factor as an Autocrine Survival Factor for Retinal Pigment Epithelial Cells under Oxidative Stress via the VEGF-R2/PI3K/Akt. Invest. Ophthalmol. Vis. Sci. 2010, 51, 1190–1197. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.L.; Chen, Y.H.; Tai, M.C.; Liang, C.M.; Lu, D.W.; Chen, J.T. Resveratrol inhibits transforming growth factor-β2-induced epithelial-to-mesenchymal transition in human retinal pigment epithelial cells by suppressing the Smad pathway. Drug Des. Dev. Ther. 2017, 11, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis induction and alteration of cell adherence in human lung cancer cells under simulated microgravity. Int. J. Mol. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [Green Version]
- Buken, C.; Sahana, J.; Corydon, T.J.; Melnik, D.; Bauer, J.; Wehland, M.; Krüger, M.; Balk, S.; Abuagela, N.; Infanger, M.; et al. Morphological and molecular changes in juvenile normal human fibroblasts exposed to simulated microgravity. Sci. Rep. Nat. Publ. Group 2019, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Tauber, S.; Seebacher, C.; Schropp, M.; Uhl, R.; Lauber, B.; Polzer, J.; Neelam, S.; Zhang, Y.; Ullrich, O. Real-time 3D high-resolution microscopy of human cells on the international space station. Int. J. Mol. Sci. 2019, 20, 2033. [Google Scholar] [CrossRef] [Green Version]
- Overbey, E.G.; da Silveira, W.A.; Stanbouly, S.; Nishiyama, N.C.; Roque-Torres, G.D.; Pecaut, M.J.; Zawieja, D.C.; Wang, C.; Willey, J.S.; Delp, M.D.; et al. Spaceflight influences gene expression, photoreceptor integrity, and oxidative stress-related damage in the murine retina. Sci. Rep. 2019, 9, 13304. [Google Scholar] [CrossRef] [PubMed]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced expression of cytoskeletal and extracellular matrix genes in human adult retinal pigment epithelium cells exposed to simulated microgravity. Cell Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Thiel, C.S.; Tauber, S.; Lauber, B.; Polzer, J.; Seebacher, C.; Uhl, R.; Neelam, S.; Zhang, Y.; Levine, H.; Ullrich, O. Rapid morphological and cytoskeletal response to microgravity in human primary macrophages. Int. J. Mol. Sci. 2019, 20, 2402. [Google Scholar] [CrossRef] [Green Version]
- Crawford-Young, S.J. Effects of microgravity on cell cytoskeleton and embryogenesis. International journal of developmental biology. Int. J. Dev. Biol. 2006, 50, 183–191. [Google Scholar] [CrossRef]
- Gruener, R.; Roberts, R.; Reitstetter, R. Reduced receptor aggregation and altered cytoskeleton in cultured myocytes after space-flight. Biol. Sci. Space 1994, 8, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Garrett, T.A.; van Buul, J.D.; Burridge, K. VEGF-induced Rac1 activation in endothelial cells is regulated by the guanine nucleotide exchange factor Vav2. Exp. Cell Res. 2007, 313, 3285–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Toledo, M.; Anguille, C.; Roger, L.; Roux, P.; Gadea, G. Cooperative anti-invasive effect of Cdc42/Rac1 activation and ROCK inhibition in SW620 colorectal cancer cells with elevated blebbing activity. PLoS ONE 2012, 7, e48344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijman, M.N.A.; van Nieuw Amerongen, G.P.; Laurens, N.; van Hinsbergh, V.W.M.; Boven, E. Microtubule-targeting agents inhibit angiogenesis at subtoxic concentrations, a process associated with inhibition of Rac1 and Cdc42 activity and changes in the endothelial cytoskeleton. Mol. Cancer Ther. 2006, 5, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Cascone, I.; Giraudo, E.; Caccavari, F.; Napione, L.; Bertotti, E.; Collard, J.G.; Serini, G.; Bussolino, F. Temporal and spatial modulation of Rho GTPases during in vitro formation of capillary vascular network: Adherens junctions and myosin light chain as targets of Rac1 and RhoA. J. Biol. Chem. 2003, 278, 50702–50713. [Google Scholar] [CrossRef] [Green Version]
- Radeke, M.J.; Radeke, C.M.; Shih, Y.H.; Hu, J.; Bok, D.; Johnson, L.V.; Coffey, P.J. Restoration of mesenchymal retinal pigmented epithelial cells by TGFβ pathway inhibitors: Implications for age-related macular degeneration. Genome. Med. 2015, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Moreno, O.; Lecanda, J.; Green, J.E.; Segura, V.; Catena, R.; Serrano, D.; Calvo, A. VEGF elicits epithelial-mesenchymal transition (EMT) in prostate intraepithelial neoplasia (PIN)-like cells via an autocrine loop. Exp. Cell Res. 2010, 316, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Lois, N.; Mcbain, V.; Abdelkader, E.; Scott, N.W.; Kumari, R. Retinal pigment epithelial atrophy in patients with exudative age-related macular degeneration undergoing anti–vascular endothelial growth factor therapy. Retina 2013, 33, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, N.S.; Zhang, H.; Boyer, D.S.; Sadda, S.R. Progression of macular atrophy in patients with neovascular age-related macular degeneration undergoing antivascular endothelial growth factor therapy. Retina 2016, 36, 1843–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.; Chui, L.; Fallah, N.; Or, C.; Merkur, A.B.; Kirker, A.W.; Albiani, D.A.; Forooghian, F. Exacerbation of choroidal and retinal pigment epithelial atrophy after anti–vascular endothelial growth factor treatment in neovascular age-related macular degeneration. Retina 2014, 34, 1308–1315. [Google Scholar] [CrossRef]
- Ewing, T.J.A.; Makino, S.; Skillman, A.G.; Kuntz, I.D. DOCK 4.0: Search strategies for automated molecular docking of flexible molecule databases. J. Comput. Aided. Mol. Des. 2001, 15, 411–428. [Google Scholar] [CrossRef]
- Hwang, J.; Yang, H.-W.; Lu, Y.A.; Je, J.G.; Lee, H.G.; Fernando, K.H.N.; Jeon, Y.-J.; Ryu, B. Phloroglucinol and dieckol isolated from Ecklonia cava suppress impaired diabetic angiogenesis; A study of in-vitro and in-vivo. Biomed. Pharm. 2021, 138, 111431. [Google Scholar] [CrossRef]
- Ryu, B.; Jiang, Y.; Kim, H.S.; Hyun, J.M.; Lim, S.B.; Li, Y.; Jeon, Y.-J. Ishophloroglucin A, a Novel Phlorotannin for Standardizing the Anti-α-Glucosidase Activity of Ishige okamurae. Mar. Drugs 2018, 16, 436. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, M.; Ryu, B.; Je, J.-G.; Jeon, Y.-J.; Kim, D.Y. Ishophloroglucin A Ameliorates VEGF-Induced Epithelial-Mesenchymal Transition via VEGFR2 Pathway Inhibition in Microgravity-Stimulated Human Retinal Pigment Epithelial Cells. Antioxidants 2022, 11, 2212. https://doi.org/10.3390/antiox11112212
Son M, Ryu B, Je J-G, Jeon Y-J, Kim DY. Ishophloroglucin A Ameliorates VEGF-Induced Epithelial-Mesenchymal Transition via VEGFR2 Pathway Inhibition in Microgravity-Stimulated Human Retinal Pigment Epithelial Cells. Antioxidants. 2022; 11(11):2212. https://doi.org/10.3390/antiox11112212
Chicago/Turabian StyleSon, Myeongjoo, Bomi Ryu, Jun-Geon Je, You-Jin Jeon, and Dae Yu Kim. 2022. "Ishophloroglucin A Ameliorates VEGF-Induced Epithelial-Mesenchymal Transition via VEGFR2 Pathway Inhibition in Microgravity-Stimulated Human Retinal Pigment Epithelial Cells" Antioxidants 11, no. 11: 2212. https://doi.org/10.3390/antiox11112212