

In Vivo Assessment of the Effects of Mono-Carrier Encapsulated Fucoxanthin Nanoparticles on Type 2 Diabetic C57 Mice and Their Oxidative Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Fabrication of the Nanocomplex
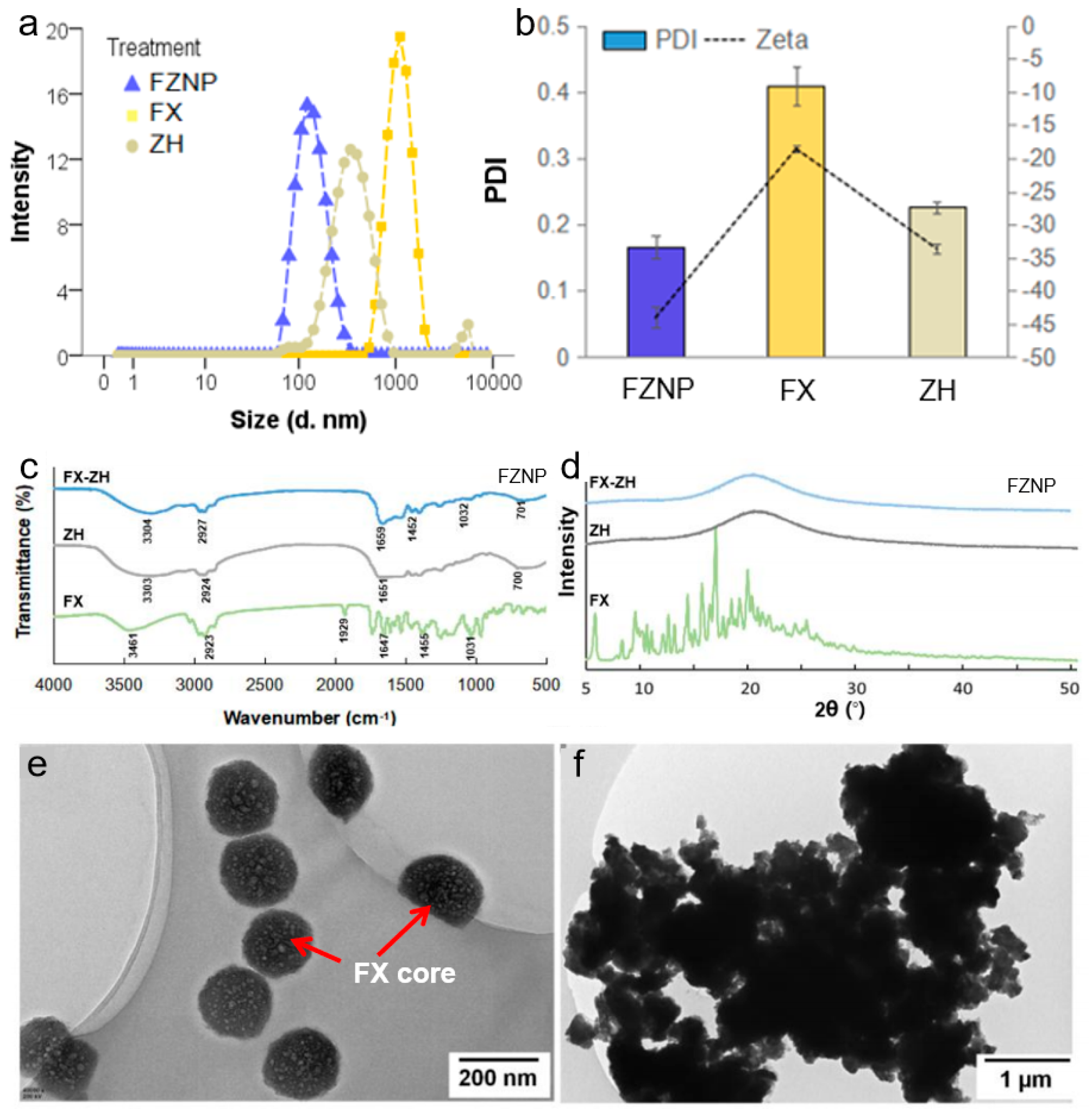
2.3. Characterization of FZNP
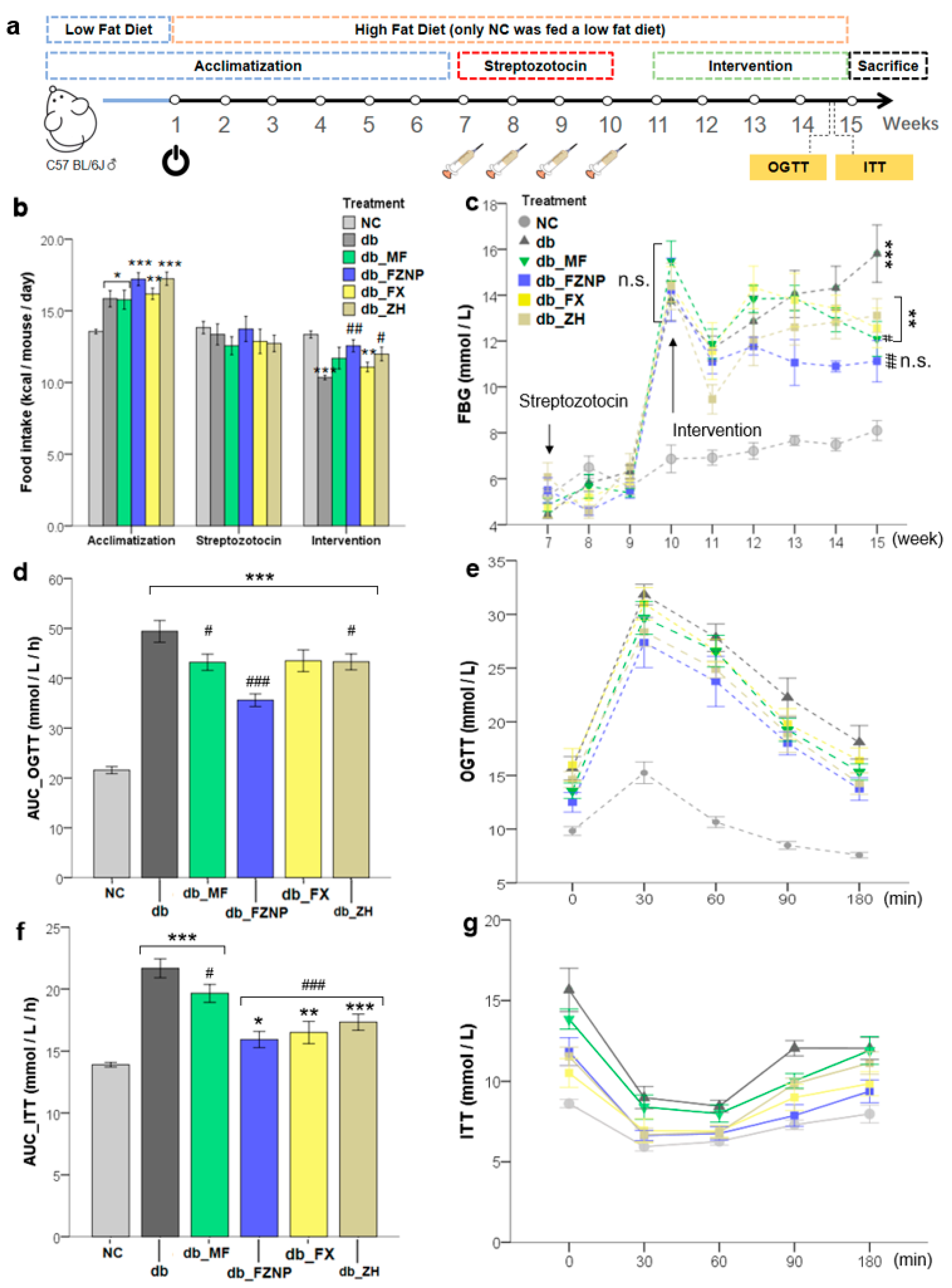
2.4. In Vivo Experiment
2.4.1. Animals and Experimental Protocols
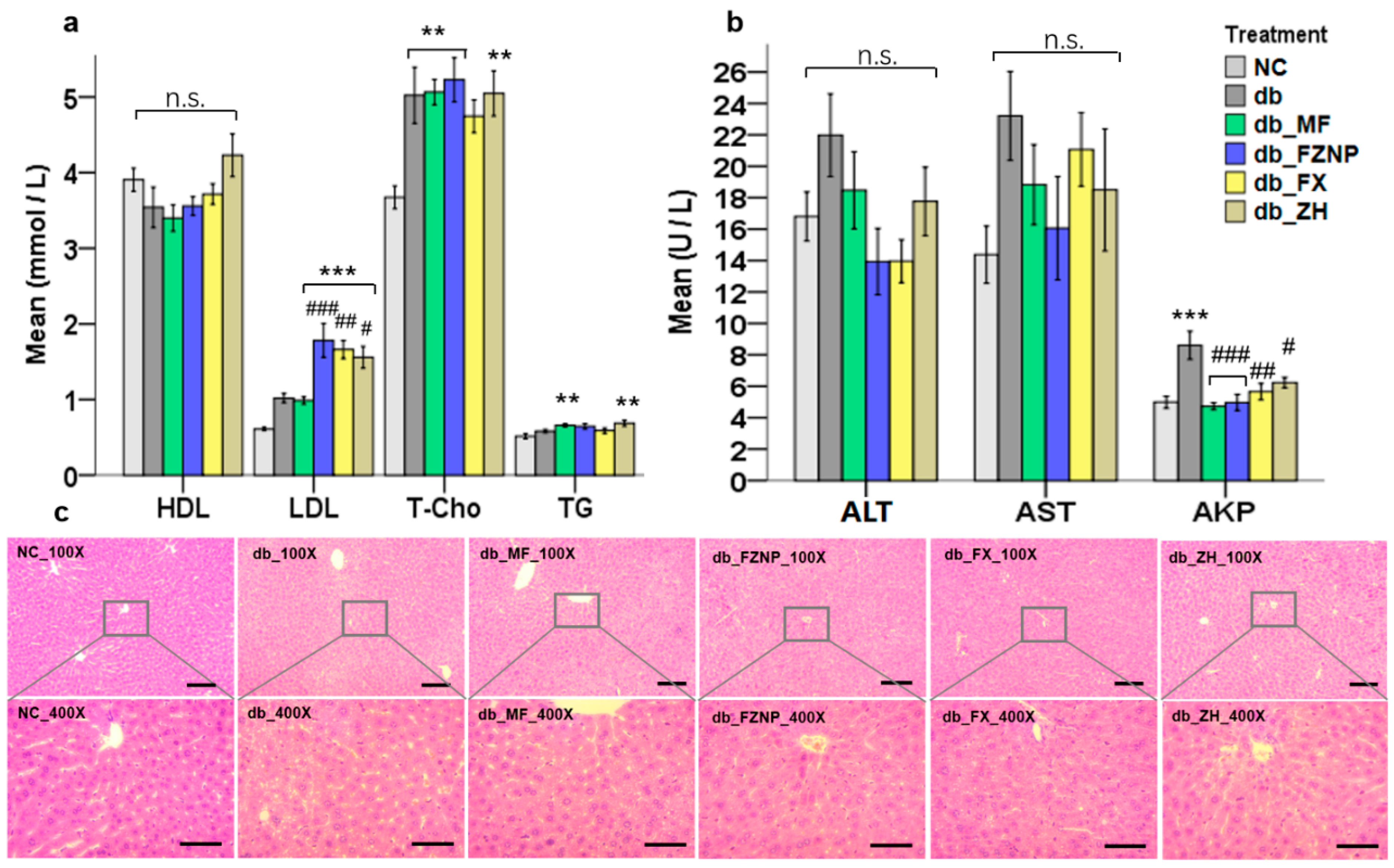
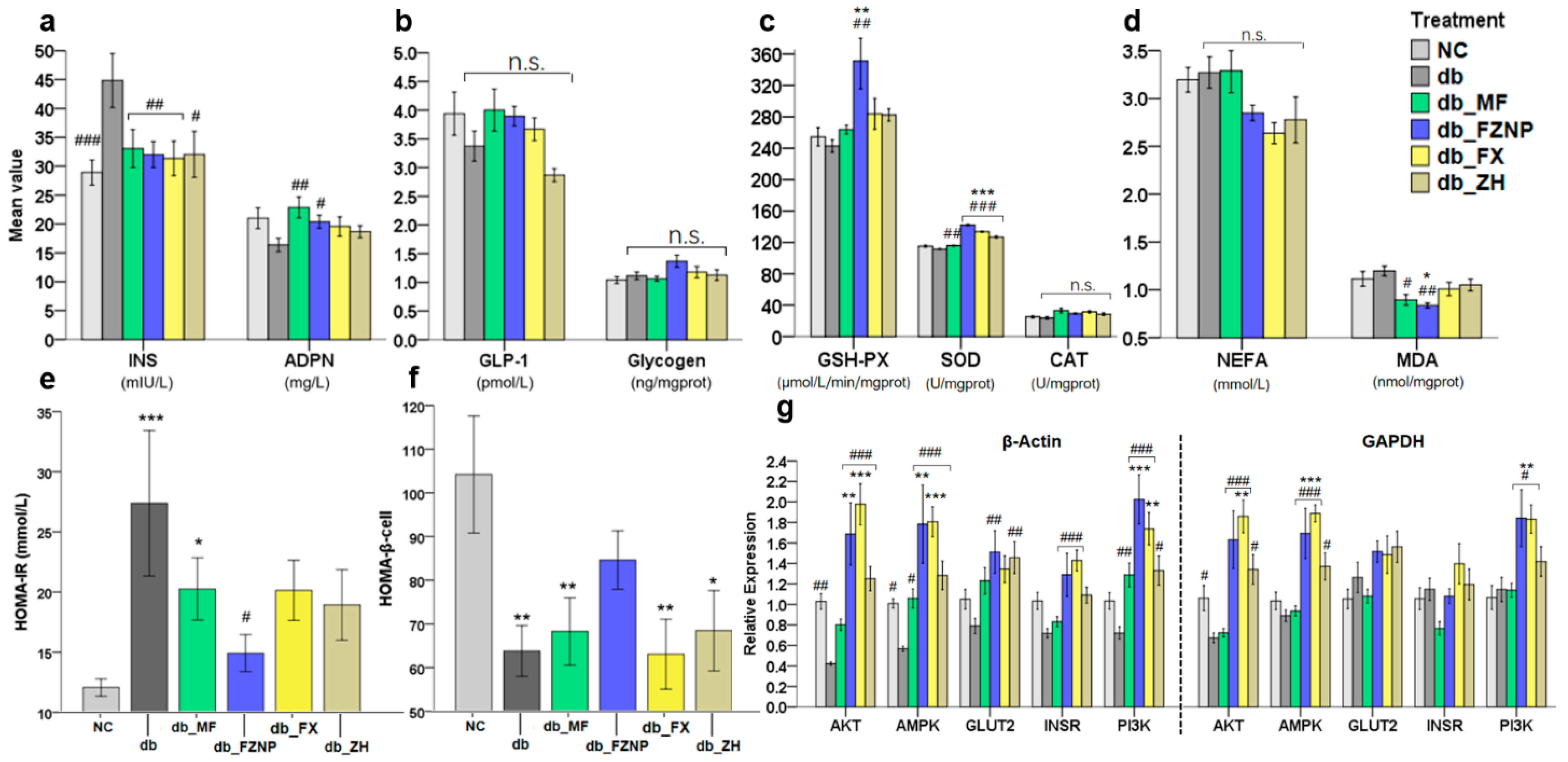
2.4.2. Biochemical Tests in Serum and Tissue
2.4.3. Histology
2.4.4. Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction Assay (Real-Time RT-qPCR)
2.4.5. Western Blot Assay
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characteristic of the FZNP
3.2. Blood Glucose Indicators
3.3. Biochemical Parameters of T2DM Mice
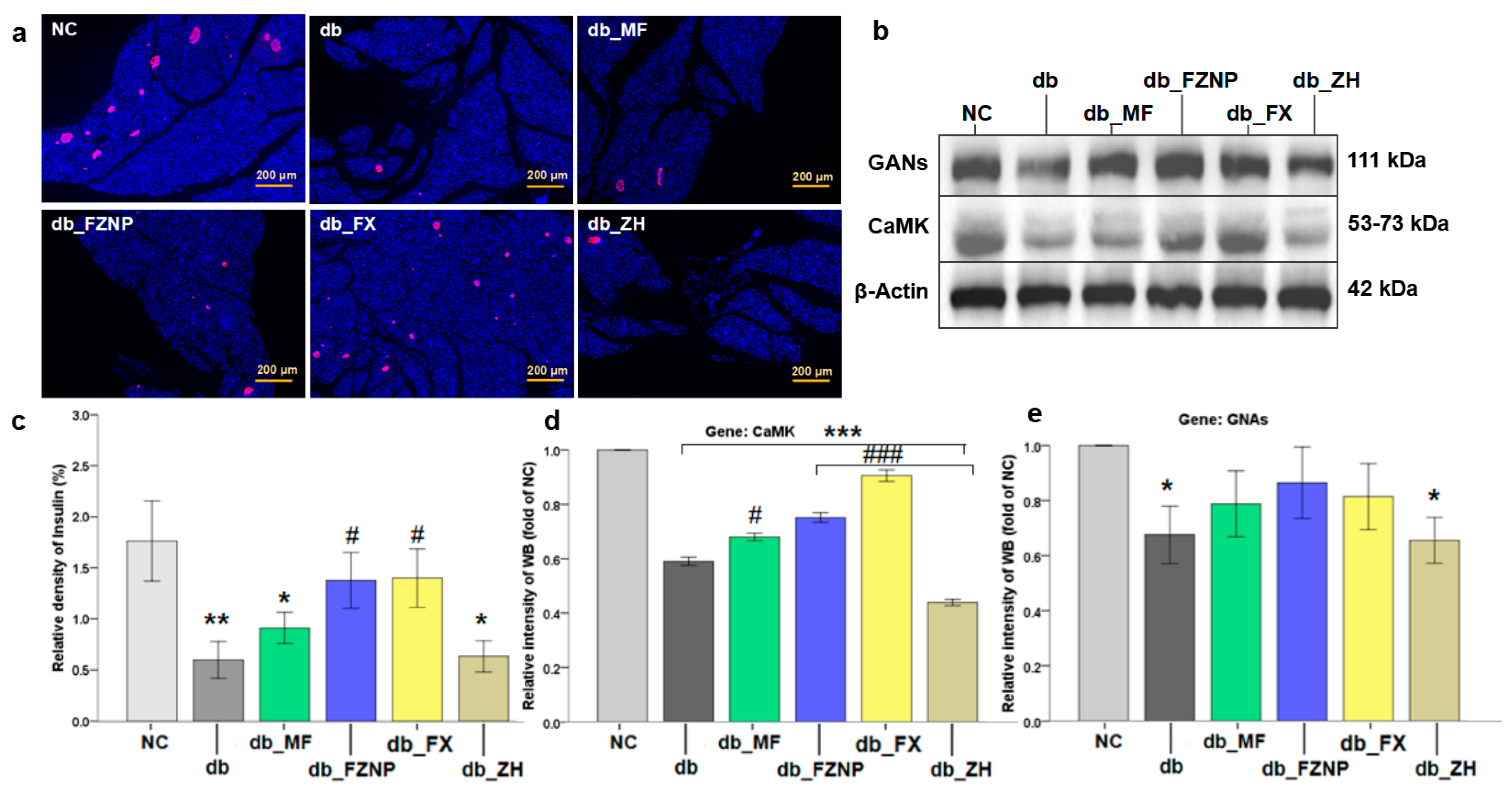
3.4. Mechanisms Affecting Diabetes
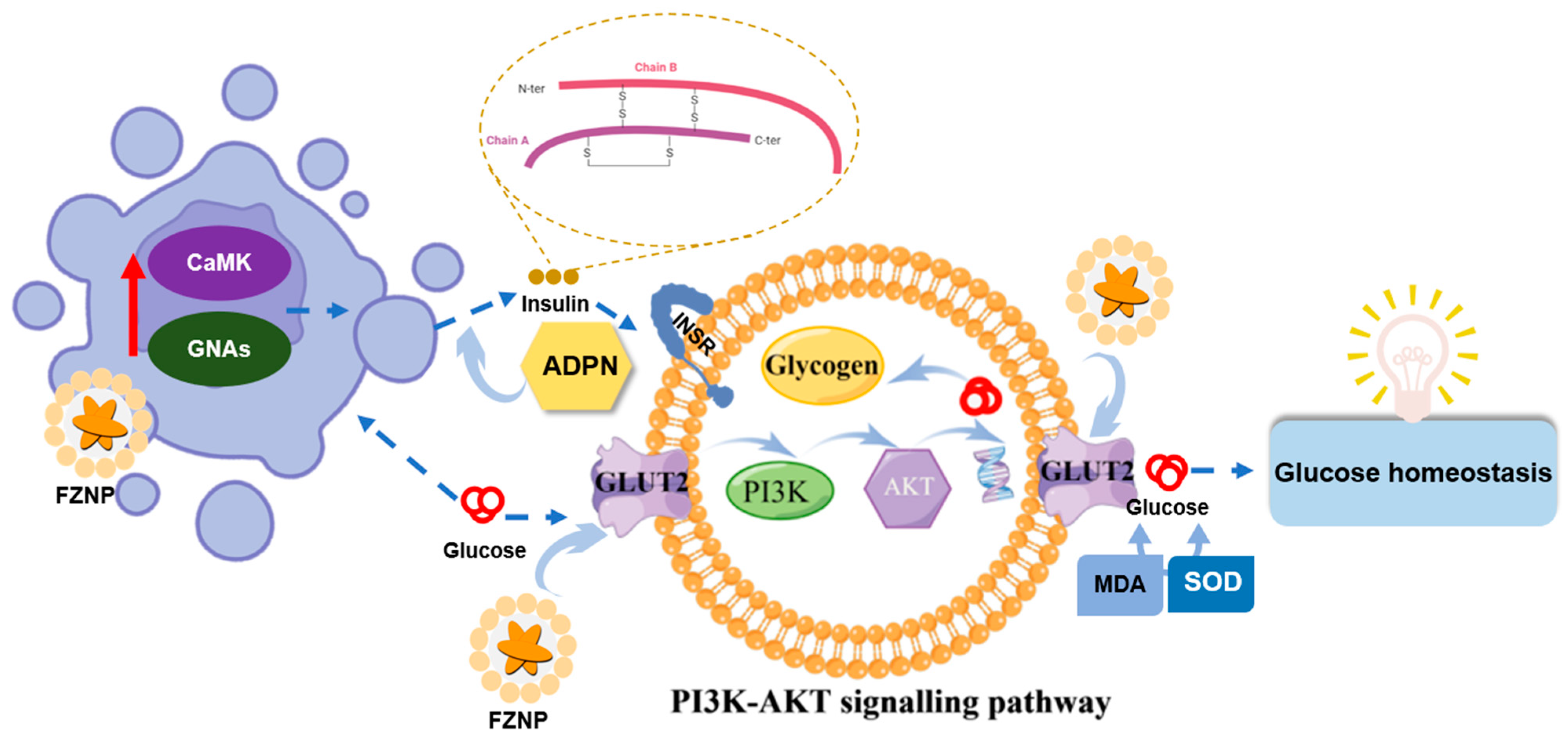
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association. Economic Costs of Diabetes in the US in 2002. Diabetes Care 2003, 26, 917–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A. Banting Lecture. From the Triumvirate to the Ominous Octet: A New Paradigm for the Treatment of Type 2 Diabetes Mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lencioni, C.; Lupi, R.; Del Prato, S. β-Cell Failure in Type 2 Diabetes Mellitus. Curr. Diabetes Rep. 2008, 8, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Kim, Y.-B. Molecular Mechanism of Insulin Resistance in Obesity and Type 2 Diabetes. Korean J. Intern. Med. 2010, 25, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Ferrannini, E.; Muscelli, E.; Frascerra, S.; Baldi, S.; Mari, A.; Heise, T.; Broedl, U.C.; Woerle, H.-J. Metabolic Response to Sodium-Glucose Cotransporter 2 Inhibition in Type 2 Diabetic Patients. J. Clin. Investig. 2014, 124, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, A.; Gan, R.-Y.; Xu, X.-Y.; Mao, Q.-Q.; Zhang, P.-Z.; Li, H.-B. Effects and Mechanisms of Edible and Medicinal Plants on Obesity: An Updated Review. Crit. Rev. Food Sci. Nutr. 2020, 61, 2061–2077. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, W.; Chen, Y.; Wan, X.; Wang, J. Fucoxanthin: A Promising Compound for Human Inflammation-Related Diseases. Life Sci. 2020, 255, 117850. [Google Scholar] [CrossRef]
- Rajauria, G. In-Vitro Antioxidant Properties of Lipophilic Antioxidant Compounds from 3 Brown Seaweed. Antioxidants 2019, 8, 596. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Kanno, S.; Kodate, M.; Hosokawa, M.; Miyashita, K. Fucoxanthinol, Metabolite of Fucoxanthin, Improves Obesity-Induced Inflammation in Adipocyte Cells. Mar. Drugs 2015, 13, 4799–4813. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xu, W.; Huang, X.; Zhao, Y.; Ren, Q.; Hong, Z.; Hong, Z.; Huang, M.; Xing, X. Fucoxanthin Ameliorates Hyperglycemia, Hyperlipidemia and Insulin Resistance in Diabetic Mice Partially Through IRS-1/PI3K/Akt and AMPK Pathways. J. Funct. Foods 2018, 48, 515–524. [Google Scholar] [CrossRef]
- Terasaki, M.; Iida, T.; Kikuchi, F.; Tamura, K.; Endo, T.; Kuramitsu, Y.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M. Fucoxanthin Potentiates Anoikis in Colon Mucosa and Prevents Carcinogenesis in AOM/DSS Model Mice. J. Nutr. Biochem. 2018, 64, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Kim, M.B.; Park, Y.K.; Lee, J.Y. Health Benefits of Fucoxanthin in the Prevention of Chronic Diseases. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-J.; Lin, T.-B.; Peng, H.-Y.; Lin, C.-H.; Lee, A.-S.; Liu, H.-J.; Li, C.-C.; Tseng, K.-W. Protective Effects of Fucoxanthin Dampen Pathogen-Associated Molecular Pattern (PAMP) Lipopolysaccharide-Induced Inflammatory Action and Elevated Intraocular Pressure by Activating Nrf2 Signaling and Generating Reactive Oxygen Species. Antioxidants 2021, 10, 1092. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin Promotes Translocation and Induction of Glucose Transporter 4 in Skeletal Muscles of Diabetic/Obese KK-Ay Mice. Phytomedicine 2012, 19, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Giri, S.; Dutta, P.; Giri, T.K. Inulin-Based Carriers for Colon Drug Targeting. J. Drug Deliv. Sci. Technol. 2021, 64, 102595. [Google Scholar] [CrossRef]
- Zhong, Q.; Jin, M. Zein Nanoparticles Produced by Liquid–Liquid Dispersion. Food Hydrocoll. 2009, 23, 2380–2387. [Google Scholar] [CrossRef]
- Chang, C.; Wang, T.; Hu, Q.; Zhou, M.; Xue, J.; Luo, Y. Pectin Coating Improves Physicochemical Properties of Caseinate/Zein Nanoparticles as Oral Delivery Vehicles for Curcumin. Food Hydrocoll. 2017, 70, 143–151. [Google Scholar] [CrossRef]
- Hu, D.; Lin, C.; Liu, L.; Li, S.; Zhao, Y. Preparation, Characterization, and In Vitro Release Investigation of Lutein/Zein Nanoparticles via Solution Enhanced Dispersion by Supercritical Fluids. J. Food Eng. 2012, 109, 545–552. [Google Scholar] [CrossRef]
- Ji, N.; Hong, Y.; Gu, Z.; Cheng, L.; Li, Z.; Li, C. Chitosan Coating of Zein-Carboxymethylated Short-Chain Amylose Nanocomposites Improves Oral Bioavailability of Insulin In Vitro and In Vivo. J. Control. Release 2019, 313, 1–13. [Google Scholar] [CrossRef]
- Chen, S.; Han, Y.; Huang, J.; Dai, L.; Du, J.; McClements, D.J.; Mao, L.; Liu, J.; Gao, Y. Fabrication and Characterization of Layer-By-Layer Composite Nanoparticles Based on Zein and Hyaluronic Acid for Codelivery of Curcumin and Quercetagetin. ACS Appl. Mater. Interfaces 2019, 11, 16922–16933. [Google Scholar] [CrossRef]
- Penalva, R.; Esparza, I.; Larraneta, E.; González-Navarro, C.J.; Gamazo, C.; Irache, J.M. Zein-Based Nanoparticles Improve the Oral Bioavailability of Resveratrol and Its Anti-inflammatory Effects in a Mouse Model of Endotoxic Shock. J. Agric. Food Chem. 2015, 63, 5603–5611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, E.; Su, Z.; Yang, C.; Ji, Y.; Liu, B.; Meng, X. Fabrication, Characterization and Properties of DHA-Loaded Nanoparticles Based on Zein and PLGA. Food Chem. 2021, 360, 129957. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Wang, J.-M.; Yang, X.-Q.; Guo, J.; Lin, Y. Amphiphilic Zein Hydrolysate as a Novel Nano-Delivery Vehicle for Curcumin. Food Funct. 2015, 6, 2636–2645. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fan, M.; Yuan, Y.; Dong, J.; Yin, H.; He, Y.; Ma, L.; Wang, D.; Yu, J. Investigation of the Optimal Fabrication of a Single-Carrier Encapsulated Fucoxanthin Based on Colloidal Nanoparticles. J. Ind. Eng. Chem. 2022, 114, 96–107. [Google Scholar] [CrossRef]
- Li, H.; Xu, Y.; Sun, X.; Wang, S.; Wang, J.; Zhu, J.; Wang, D.; Zhao, L. Stability, Bioactivity, and Bioaccessibility of Fucoxanthin in Zein-Caseinate Composite Nanoparticles Fabricated at Neutral pH by Antisolvent Precipitation. Food Hydrocoll. 2018, 84, 379–388. [Google Scholar] [CrossRef]
- Xia, X.; Xu, J.; Wang, X.; Wang, H.; Lin, Z.; Shao, K.; Fang, L.; Zhang, C.; Zhao, Y. Jiaogulan Tea (Gpostemma Pentaphyllum) Potentiates the Antidiabetic Effect of White Tea via the AMPK And PI3K Pathways in C57BL/6 Mice. Food Funct. 2020, 11, 4339–4355. [Google Scholar] [CrossRef]
- Hirano, H.; Watanabe, T. Microsequencing of Proteins Electrotransferred onto Immobilizing Matrices from Polyacrylamide Gel Electrophoresis: Application to an Insoluble Protein. Electrophoresis 1990, 11, 573–580. [Google Scholar] [CrossRef]
- Moore, J.; Cerasoli, E. Particle Light Scattering Methods and Applications. In Encyclopedia of Spectroscopy and Spectrometry, 2nd ed.; Lindon, J.C., Ed.; Academic Press: Oxford, UK, 2010; pp. 2077–2088. [Google Scholar]
- Koo, S.Y.; Mok, I.-K.; Pan, C.-H.; Kim, S.M. Preparation of Fucoxanthin-Loaded Nanoparticles Composed of Casein and Chitosan with Improved Fucoxanthin Bioavailability. J. Agric. Food Chem. 2016, 64, 9428–9435. [Google Scholar] [CrossRef]
- Wang, C.; Ren, J.; Song, H.; Chen, X.; Qi, H. Characterization of Whey Protein-Based Nanocomplex to Load Fucoxanthin and The Mechanism of Action on Glial Cells PC12. LWT 2021, 151, 112208. [Google Scholar] [CrossRef]
- Song, Q.; Liu, X.; Wang, A.; Wang, Y.; Zhou, Y.; Zhou, W.; Wang, X. Associations Between Non-Traditional Lipid Measures and Risk for Type 2 Diabetes Mellitus in a Chinese Community Population: A Cross-Sectional Study. Lipids Health Dis. 2016, 15, 1–6. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic Syndrome: A Comprehensive Perspective Based on Interactions Between Obesity, Diabetes, and Inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Liu, D.; Chen, Y.; Wu, J.; Wang, S. Antioxidant Activity of Sulfated Polysaccharide Fractions Extracted from Undaria Pinnitafida In Vitro. Int. J. Biol. Macromol. 2010, 46, 193–198. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; De Girolamo, M.; Cusenza, S.; Riccioni, G. Marine Bioactives: Pharmacological Properties and Potential Applications against Inflammatory Diseases. Marine Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busato, S.; Bionaz, M. The Interplay between Non-Esterified Fatty Acids and Bovine Peroxisome Proliferator-Activated Receptors: Results of an In Vitro Hybrid Approach. J. Anim. Sci. Biotechnol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Zhou, S.S.; He, F.; Chen, A.H.; Hao, P.Y.; Song, X.D. Suppression of Rat Frizzled-2 Attenuates Hypoxia/Reoxygenation-Induced Ca2+ Accumulation in Rat H9c2 Cells. Exp. Cell Res. 2012, 318, 1480–1491. [Google Scholar] [CrossRef]
- Krook, A.; Kawano, Y.; Song, X.M.; Efendić, S.; Roth, R.A.; Wallberg-Henriksson, H.; Zierath, J.R. Improved Glucose Tolerance Restores Insulin-Stimulated Akt Kinase Activity and Glucose Transport in Skeletal Muscle from Diabetic Goto-Kakizaki Rats. Diabetes 1997, 46, 2110–2114. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Cantley, L.C. AKT/PKB Signaling: Navigating Downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tang, Y.; Shi, S.; Gao, S.; Wang, Y.; Xiao, D.; Chen, T.; He, Q.; Zhang, J.; Lin, Y. Tetrahedral Framework Nucleic Acids Ameliorate Insulin Resistance in Type 2 Diabetes Mellitus via the PI3K/Akt Pathway. ACS Appl. Mater. Interfaces 2021, 13, 40354–40364. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Q.; Ji, H.; Lu, H.; Yang, Q.; Yin, J.; Guan, W. Porcine Pancreas Mesenchymal Cell Characterization and Functional Differentiation into Insulin-Producing Cells In Vitro. Mol. Med. Rep. 2021, 24, 1–9. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | |
|---|---|---|
| * β-Actin | Forward | 5′-GTGCTATGTTGCTCTAGACTTCG-3′ |
| Reverse | 5′-ATGCCACAGGATTCCATACC-3′ | |
| INSR | Forward | 5′-CGTGTTGCGGTTAAGACTGTCAATG-3′ |
| Reverse | 5′-CCAAGAAGGCGGACCACATGATG-3′ | |
| PI3K | Forward | 5′-GCTTTGCCGAGCCCTACAAC-3′ |
| Reverse | 5′-GTCATTGTGCTGCACGAGGG-3′ | |
| AKT | Forward | 5′-TCAGGATGTGGATCAGCGAGAGTC-3′ |
| Reverse | 5′-AGGCAGCGGATGATAAAGGTGTTG-3′ | |
| AMPK | Forward | 5′-CGAGTGTTCGGAGGAGGAGGTC-3′ |
| Reverse | 5′-GTGGGCTGGTTGCTAGGTAGAAATC-3′ | |
| GLUT2 | Forward | 5′-ACAGTCACACCAGCATACACAACAC-3′ |
| Reverse | 5′-CCGAGCCACCCACCAAAGAATG-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Fan, M.; Luo, K.; Xu, W.; Dong, J.; Wang, D.; Chen, L.; Yu, J. In Vivo Assessment of the Effects of Mono-Carrier Encapsulated Fucoxanthin Nanoparticles on Type 2 Diabetic C57 Mice and Their Oxidative Stress. Antioxidants 2022, 11, 1976. https://doi.org/10.3390/antiox11101976
Zhang X, Fan M, Luo K, Xu W, Dong J, Wang D, Chen L, Yu J. In Vivo Assessment of the Effects of Mono-Carrier Encapsulated Fucoxanthin Nanoparticles on Type 2 Diabetic C57 Mice and Their Oxidative Stress. Antioxidants. 2022; 11(10):1976. https://doi.org/10.3390/antiox11101976
Chicago/Turabian StyleZhang, Xin, Minghao Fan, Ke Luo, Wanying Xu, Jianjun Dong, Dongfeng Wang, Lu Chen, and Junhong Yu. 2022. "In Vivo Assessment of the Effects of Mono-Carrier Encapsulated Fucoxanthin Nanoparticles on Type 2 Diabetic C57 Mice and Their Oxidative Stress" Antioxidants 11, no. 10: 1976. https://doi.org/10.3390/antiox11101976