



Ethnomedicinal Plants with Protective Effects against Beta-Amyloid Peptide (Aβ)1-42 Indicate Therapeutic Potential in a New In Vivo Model of Alzheimer’s Disease
 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Extracts
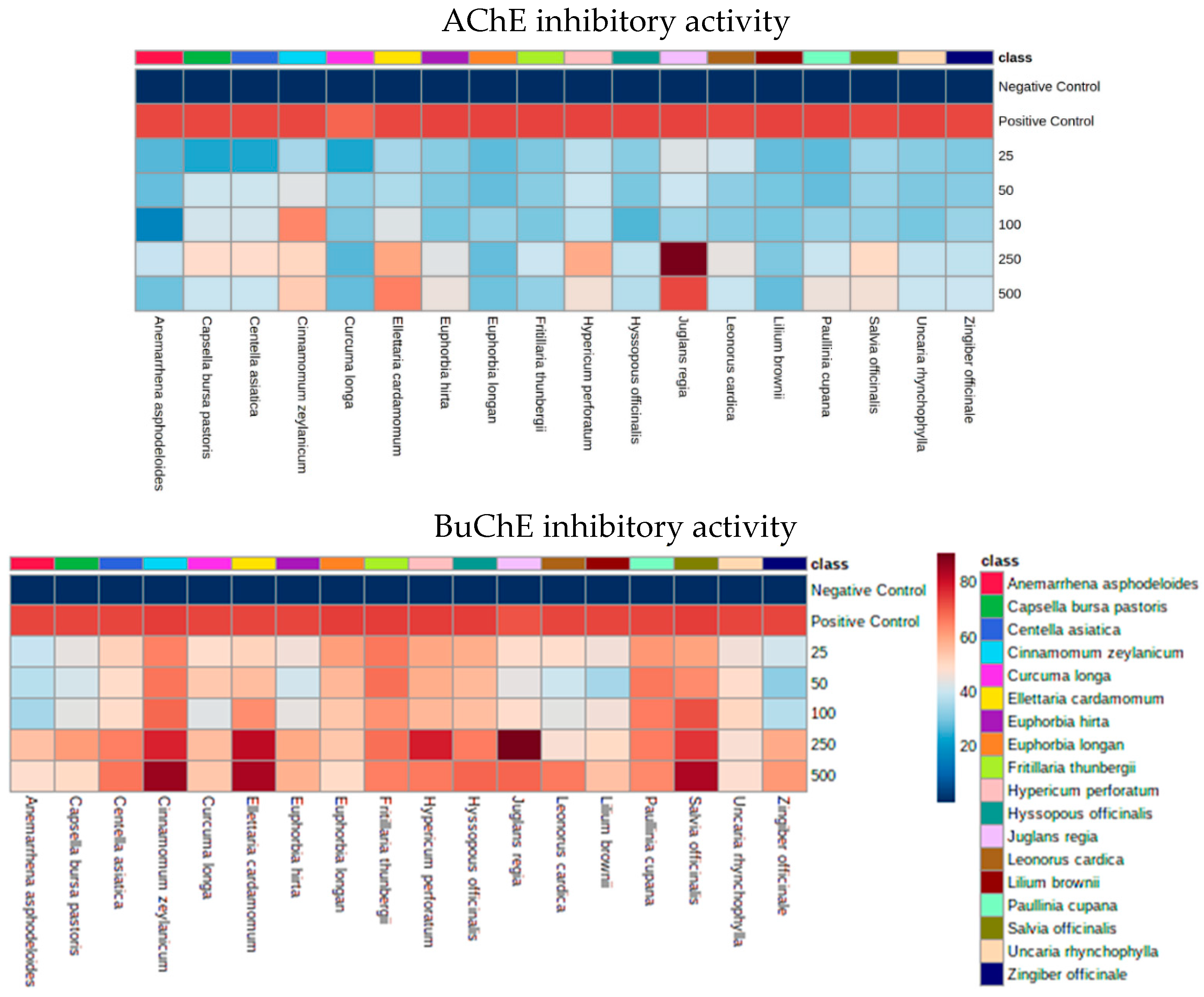
2.3. AChE and BuChE Inhibitory Activities
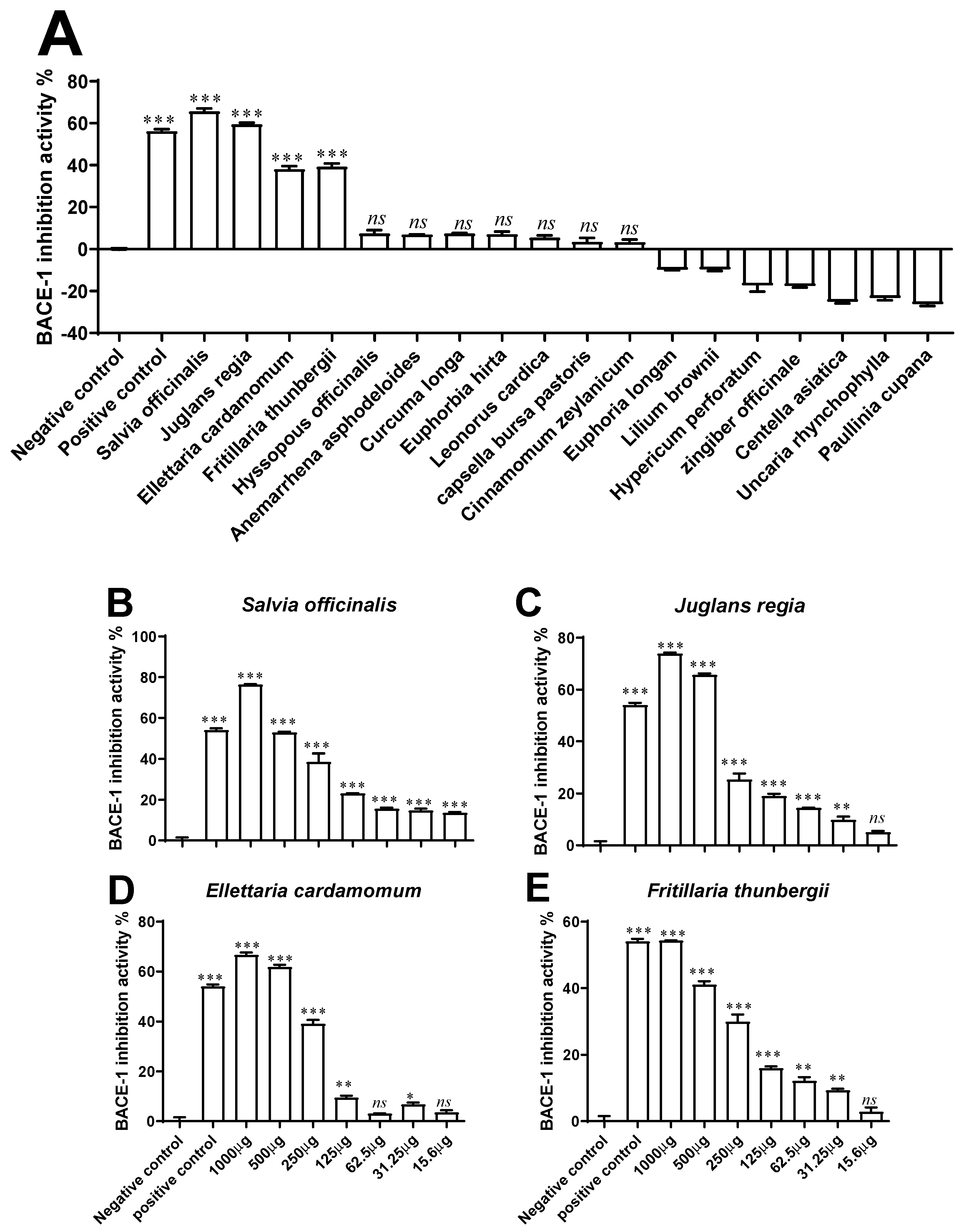
2.4. BACE-1 Inhibitory Activity
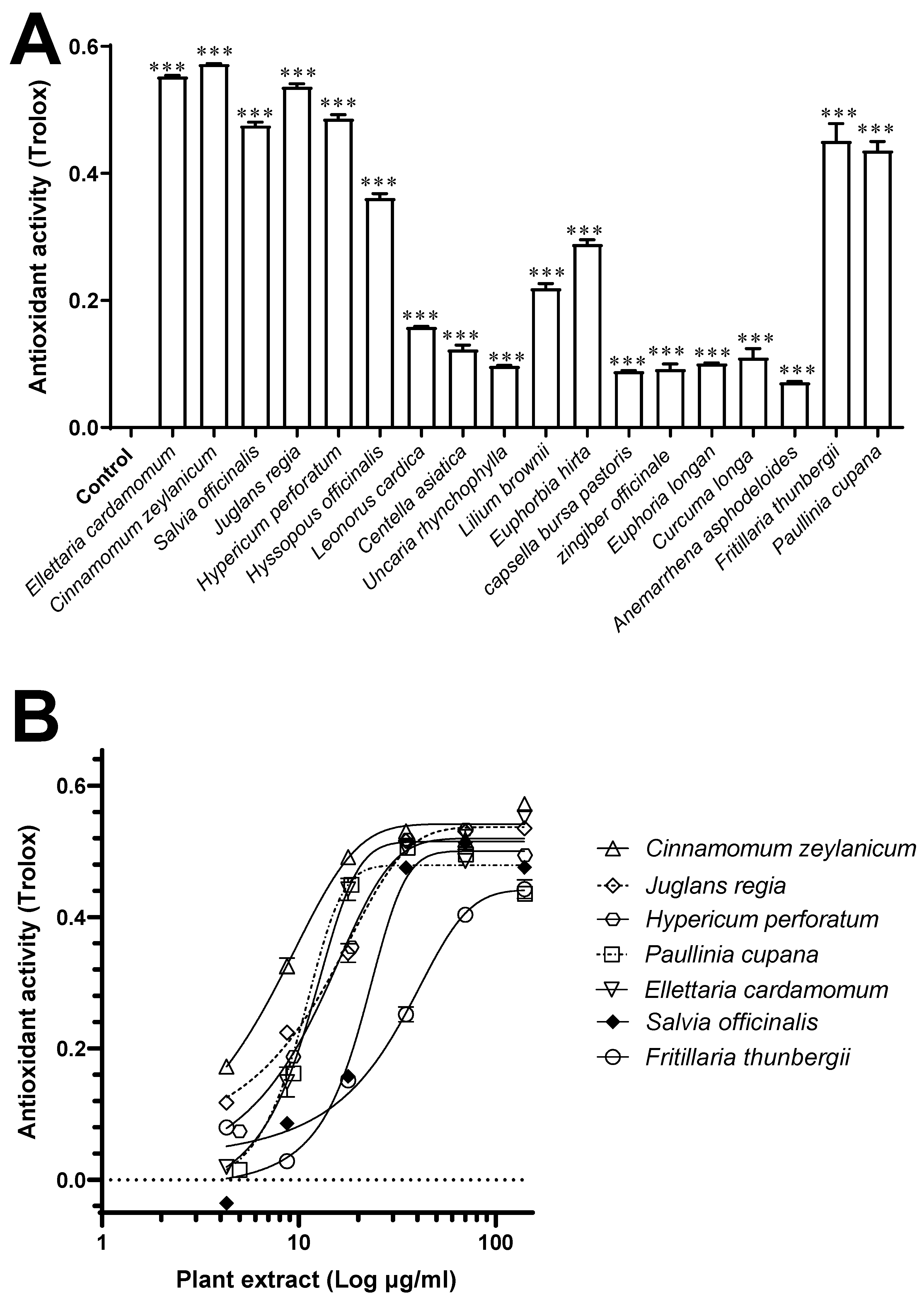
2.5. Antioxidant Activity
2.6. Chemotaxis Studies in G. pallida
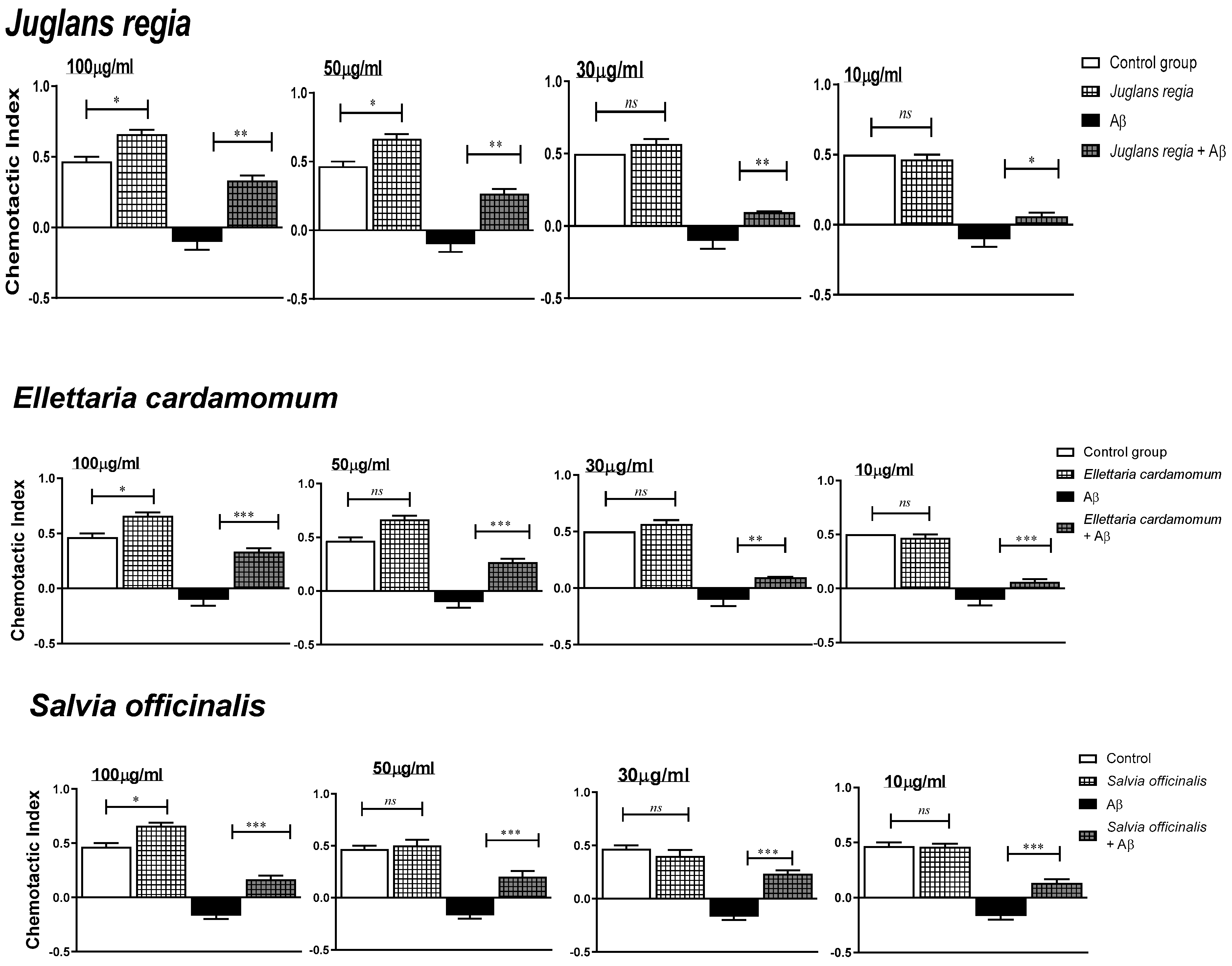
2.7. Phytoprotective Effects against Aβ (1-42)-Induced Impairments in G. pallida
2.8. Statistical Analyses
3. Results
3.1. Cholinesterase Inhibitory Activities
3.2. BACE-1 Inhibitory Activity
3.3. Antioxidant Activity
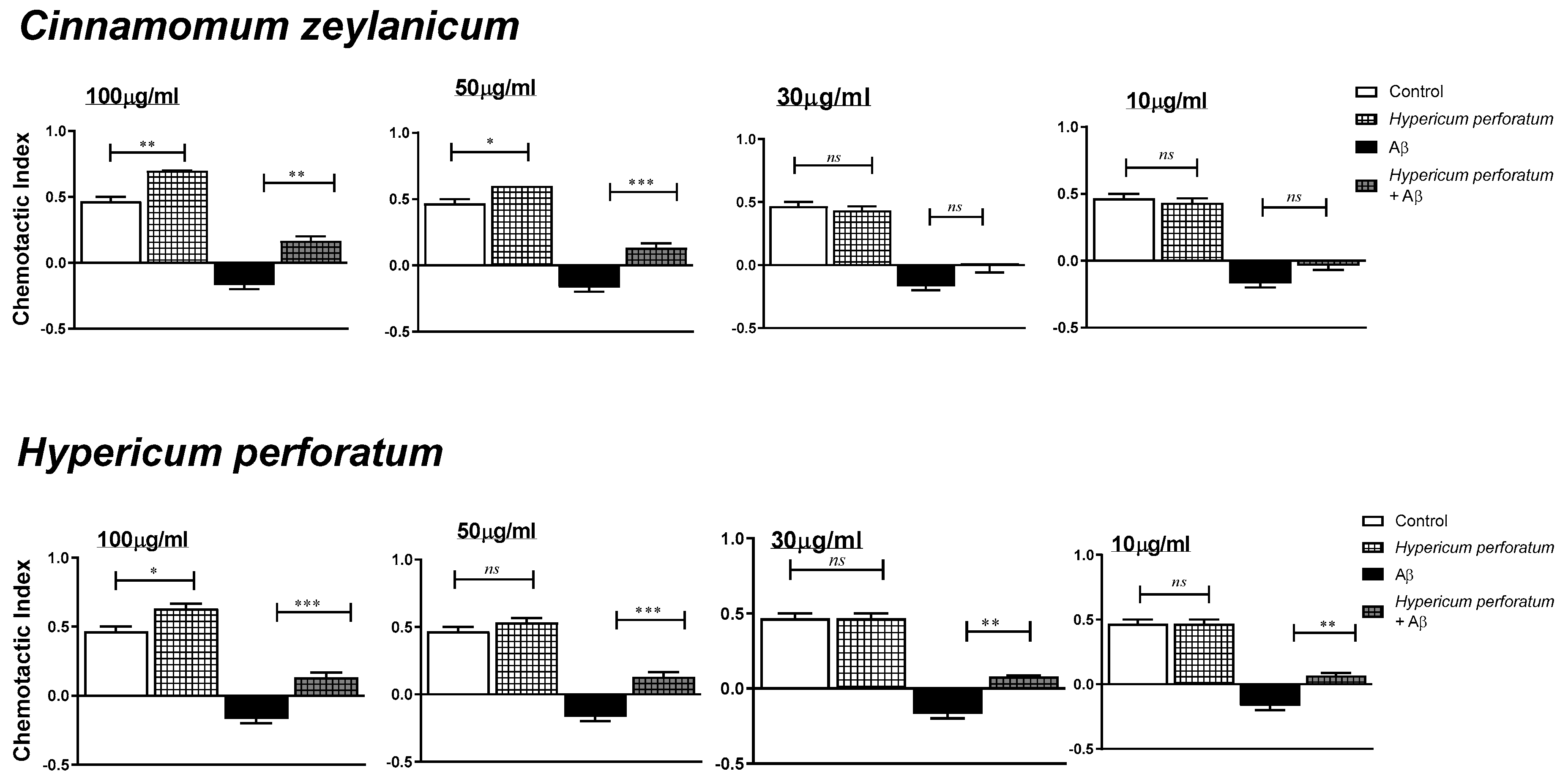
3.4. Effects on Chemotaxis in G. pallida
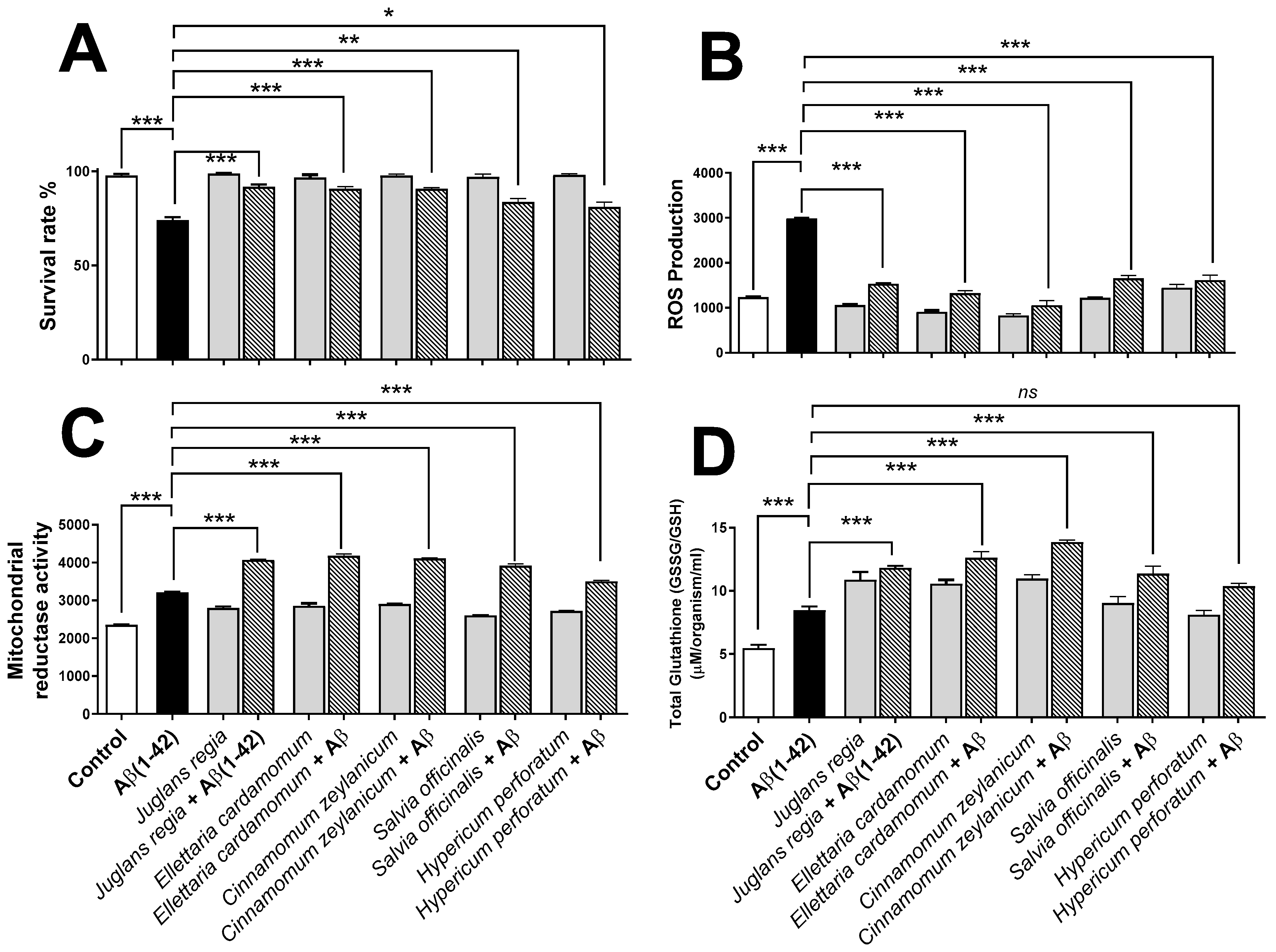
3.5. Phytoprotective Effects against Aβ (1-42)-Induced Impairments in G. pallida
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dey, A.; Bhattacharya, R.; Mukherjee, A.; Pandey, D.K. Natural products against Alzheimer’s disease: Pharmaco-therapeutics and biotechnological interventions. Biotechnol. Adv. 2017, 35, 178–216. [Google Scholar] [CrossRef]
- Sang, Z.; Pan, W.; Wang, K.; Ma, Q.; Yu, L.; Yang, Y.; Bai, P.; Leng, C.; Xu, Q.; Li, X.; et al. Design, synthesis and evaluation of novel ferulic acid-O-alkylamine derivatives as potential multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2017, 130, 379–392. [Google Scholar] [CrossRef]
- Thiratmatrakul, S.; Yenjai, C.; Waiwut, P.; Vajragupta, O.; Reubroycharoen, P.; Tohda, M.; Boonyarat, C. Synthesis, biological evaluation and molecular modeling study of novel tacrine-carbazole hybrids as potential multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2014, 75, 21–30. [Google Scholar] [CrossRef]
- Casey, D.A.; Antimisiaris, D.; O’Brien, J. Drugs for Alzheimer’s disease: Are they effective? P T A Peer-Rev. J. Formul. Manag. 2010, 35, 208–211. [Google Scholar]
- Lleó, A. Current therapeutic options for Alzheimer’s disease. Curr. Genom. 2007, 8, 550–558. [Google Scholar] [CrossRef]
- Butler, M.S. The role of natural product chemistry in drug discovery. J. Nat. Prod. 2004, 67, 2141–2153. [Google Scholar] [CrossRef]
- World Health Organization. Programme on Traditional Medicine. WHO Traditional Medicine Strategy 2002–2005. World Health Organization. 2002. Available online: https://apps.who.int/iris/handle/10665/67163 (accessed on 22 August 2022).
- Sun, Z.K.; Yang, H.Q.; Chen, S.D. Traditional Chinese medicine: A promising candidate for the treatment of Alzheimer’s disease. Transl. Neurodegener. 2013, 2, 6. [Google Scholar] [CrossRef]
- Ansari, N.; Khodagholi, F. Natural products as promising drug candidates for the treatment of Alzheimer’s disease: Molecular mechanism aspect. Curr. Neuropharmacol. 2013, 11, 414–429. [Google Scholar] [CrossRef]
- Carvajal, F.J.; Inestrosa, N.C. Interactions of AChE with Aβ Aggregates in Alzheimer’s Brain: Therapeutic Relevance of IDN 5706. Front. Mol. Neurosci. 2011, 4, 19. [Google Scholar] [CrossRef]
- Althobaiti, N.A.; Menaa, F.; Albalawi, A.E.; Dalzell, J.J.; Warnock, N.D.; Mccammick, E.M.; Alsolais, A.; Alkhaibari, A.M.; Green, B.D. Assessment and Validation of Globodera pallida as a Novel In Vivo Model for Studying Alzheimer’s Disease. Cells 2021, 10, 2481. [Google Scholar] [CrossRef]
- Althobaiti, N.A.; Menaa, F.; Dalzell, J.J.; Green, B.D. Globodera pallida, a non-transgenic invertebrate as a new model for investigating Alzheimer’s disease (and other proteinopathies)? Neural Regen. Res. 2022, 18, 113–114. [Google Scholar] [CrossRef]
- Folch, J.; Petrov, D.; Ettcheto, M.; Abad, S.; Sánchez-López, E.; García, M.L.; Olloquequi, J.; Beas-Zarate, C.; Auladell, C.; Camins, A. Current Research Therapeutic Strategies for Alzheimer’s Disease Treatment. Neural Plast. 2016, 2016, 8501693. [Google Scholar] [CrossRef]
- Sun, X.; Chen, W.D.; Wang, Y.D. β-Amyloid: The key peptide in the pathogenesis of Alzheimer’s disease. Front. Pharmacol. 2015, 6, 221. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.G.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Zaitoun, S.Y.A.; Qasem, I.B. In-vitro screening of acetylcholinesterase inhibitory activity of extracts from Palestinian indigenous flora in relation to the treatment of Alzheimer’s disease. Funct. Foods Health Dis. 2014, 4, 381–400. [Google Scholar] [CrossRef]
- Ljubenkov, I.; Kri, A.; Juki, M. Antioxidant and acetylcholinesterase inhibiting activity of several aqueous tea infusions in vitro. Food Technol. Biotechnol. 2008, 46, 368–375. [Google Scholar]
- Essa, M.M.; Subash, S.; Dhanalakshmi, C.; Manivasagam, T.; Al-Adawi, S.; Guillemin, G.J.; Thenmozhi, A.J. Dietary supplementation of walnut partially reverses 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine induced neurodegeneration in a mouse model of Parkinson’s disease. Neurochem. Res. 2015, 40, 1283–1293. [Google Scholar] [CrossRef]
- Fisher, D.R.; Poulose, S.M.; Bielinski, D.F.; Shukitt-Hale, B. Serum metabolites from walnut-fed aged rats attenuate stress-induced neurotoxicity in BV-2 microglial cells. Nutr. Neurosci. 2017, 20, 103–109. [Google Scholar] [CrossRef]
- Shabani, M.; Nazeri, M.; Parsania, S.; Razavinasab, M.; Zangiabadi, N.; Esmaeilpour, K.; Abareghi, F. Walnut consumption protects rats against cisplatin-induced neurotoxicity. Neurotoxicology 2012, 33, 1314–1321. [Google Scholar] [CrossRef]
- Rath, B.; Pradhan, D. Antidepressant Activity of Juglans regia L. fruit extract. Int. J. Toxicol. Pharmacol. Res. 2009, 1, 24–26. [Google Scholar]
- Willis, L.M.; Shukitt-Hale, B.; Cheng, V.; Joseph, J.A. Dose-dependent effects of walnuts on motor and cognitive function in aged rats. Br. J. Nutr. 2009, 101, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Taha, N.A.; Al-wadaan, M.A. Utility and importance of walnut, Juglans regia Linn: A review. Afr. J. Microbiol. Res. 2011, 5, 5796–5805. [Google Scholar]
- Nag, G.; De, B. Acetylcholinesterase inhibitory activity of Terminalia chebula, Terminalia bellerica and Emblica officinalis and some phenolic compounds. Int. J. Pharm. Pharm. Sci. 2011, 3, 121–124. [Google Scholar]
- Orhan, I.E.; Suntar, I.P.; Akkol, E.K. In vitro neuroprotective effects of the leaf and fruit extracts of Juglans regia L. (walnut) through enzymes linked to Alzheimer’s disease and antioxidant activity. Int. J. Food Sci. Nutr. 2011, 62, 781–786. [Google Scholar] [CrossRef]
- Chauhan, N.; Wang, K.C.; Wegiel, J.; Malik, M.N. Walnut extract inhibits the fibrillization of amyloid beta-protein, and also defibrillizes its preformed fibrils. Curr. Alzheimer Res. 2004, 1, 183–188. [Google Scholar] [CrossRef]
- Muthaiyah, B.; Essa, M.M.; Chauhan, V.; Chauhan, A. Protective effects of walnut extract against amyloid beta peptide-induced cell death and oxidative stress in PC12 cells. Neurochem. Res. 2011, 36, 2096–2103. [Google Scholar] [CrossRef]
- Danysz, W.; Parsons, C.G. Alzheimer’s disease, β-amyloid, glutamate, NMDA receptors and memantine--searching for the connections. Br. J. Pharmacol. 2012, 167, 324–352. [Google Scholar] [CrossRef]
- Link, P.; Roth, K.; Sporer, F.; Wink, M. Carlina acaulis Exhibits Antioxidant Activity and Counteracts Aβ Toxicity in Caenorhabditis elegans. Molecules 2016, 21, 871. [Google Scholar] [CrossRef]
- Demuro, A.; Parker, I.; Stutzmann, G.E. Calcium signaling and amyloid toxicity in Alzheimer disease. J. Biol. Chem. 2010, 285, 12463–12468. [Google Scholar] [CrossRef]
- Yu, Y.B.; Dosanjh, L.; Lao, L.; Tan, M.; Shim, B.S.; Luo, Y. Cinnamomum cassia bark in two herbal formulas increases life span in Caenorhabditis elegans via insulin signaling and stress response pathways. PLoS ONE 2010, 5, e9339. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Brijeshlata, D.S.; Dixit, S. Screening of traditional Indian spices for inhibitory activity of acetylcholinesterase and butyrylcholinesterase enzymes. Int. J. Pharma Bio Sci. 2012, 3, P59–P65. [Google Scholar]
- Masoumi-Ardakani, Y.; Mahmoudvand, H.; Mirzaei, A.; Esmaeilpour, K.; Ghazvini, H.; Khalifeh, S.; Sepehri, G. The effect of Elettaria cardamomum extract on anxiety-like behavior in a rat model of post-traumatic stress disorder. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 87, 489–495. [Google Scholar] [CrossRef]
- Yadav, A.S.; Bhatnagar, D. Free radical scavenging activity, metal chelation and antioxidant power of some of the Indian spices. BioFactors 2007, 31, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Wszelaki, N.; Kuciun, A.; Kiss, A.K. Screening of traditional European herbal medicines for acetylcholinesterase and butyrylcholinesterase inhibitory activity. Acta Pharm. 2010, 60, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.F.; Falé, P.L.; Araújo, M.E.M.; Serralheiro, M.L.M. Acetylcholinesterase inhibition and antioxidant activity of the water extracts of several Hypericum species. Food Chem. 2010, 120, 1076–1082. [Google Scholar] [CrossRef]
- Altun, M.L.; Yılmaz, B.S.; Orhan, I.E.; Citoglu, G.S. Assessment of cholinesterase and tyrosinase inhibitory and antioxidant effects of Hypericum perforatum L.(St. John’s wort). Ind. Crops Prod. 2013, 43, 87–92. [Google Scholar] [CrossRef]
- Sallam, A.; Mira, A.; Ashour, A.; Shimizu, K. Acetylcholine esterase inhibitors and melanin synthesis inhibitors from Salvia officinalis. Phytomed. Int. J. Phytother. Phytopharm. 2016, 23, 1005–1011. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Scholey, A.B. The psychopharmacology of European herbs with cognition-enhancing properties. Curr. Pharm. Des. 2006, 12, 4613–4623. [Google Scholar] [CrossRef]
- Savelev, S.U.; Okello, E.J.; Perry, E.K. Butyryl- and acetyl-cholinesterase inhibitory activities in essential oils of Salvia species and their constituents. Phytother. Res. PTR 2004, 18, 315–324. [Google Scholar] [CrossRef]
- Tildesley, N.T.; Kennedy, D.O.; Perry, E.K.; Ballard, C.G.; Savelev, S.; Wesnes, K.A.; Scholey, A.B. Salvia lavandulaefolia (Spanish sage) enhances memory in healthy young volunteers. Pharmacol. Biochem. Behav. 2003, 75, 669–674. [Google Scholar] [CrossRef]
- Smach, M.A.; Hafsa, J.; Charfeddine, B.; Dridi, H.; Limem, K. Effects of sage extract on memory performance in mice and acetylcholinesterase activity. Ann. Pharm. Fr. 2015, 73, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Frustaci, A.; Del Bufalo, A.; Fini, M.; Cesario, A. From traditional European medicine to discovery of new drug candidates for the treatment of dementia and Alzheimer’s disease: Acetylcholinesterase inhibitors. Curr. Med. Chem. 2013, 20, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; De Filippis, D.; Esposito, G.; D’Amico, A.; Izzo, A.A. The spice sage and its active ingredient rosmarinic acid protect PC12 cells from amyloid-beta peptide-induced neurotoxicity. J. Pharmacol. Exp. Ther. 2006, 317, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Imanshahidi, M.; Hosseinzadeh, H. The pharmacological effects of Salvia species on the central nervous system. Phytother. Res. PTR 2006, 20, 427–437. [Google Scholar] [CrossRef]
- Akhondzadeh, S.; Noroozian, M.; Mohammadi, M.; Ohadinia, S.; Jamshidi, A.H.; Khani, M. Salvia officinalis extract in the treatment of patients with mild to moderate Alzheimer’s disease: A double blind, randomized and placebo-controlled trial. J. Clin. Pharm. Ther. 2003, 28, 53–59. [Google Scholar] [CrossRef]
- Vladimir-Knežević, S.; Blažeković, B.; Kindl, M.; Vladić, J.; Lower-Nedza, A.D.; Brantner, A.H. Acetylcholinesterase inhibitory, antioxidant and phytochemical properties of selected medicinal plants of the Lamiaceae family. Molecules 2014, 19, 767–782. [Google Scholar] [CrossRef]
- Lopresti, A.L. Salvia (Sage): A Review of its Potential Cognitive-Enhancing and Protective Effects. Drugs RD 2017, 17, 53–64. [Google Scholar] [CrossRef]
- Lee, Y.W.; Kim, D.H.; Jeon, S.J.; Park, S.J.; Kim, J.M.; Jung, J.M.; Lee, H.E.; Bae, S.G.; Oh, H.K.; Son, K.H.H. Neuroprotective effects of salvianolic acid B on an Aβ25–35 peptide-induced mouse model of Alzheimer’s disease. Eur. J. Pharmacol. 2013, 704, 70–77. [Google Scholar] [CrossRef]
- Rasoolijazi, H.; Azad, N.; Joghataei, M.T.; Kerdari, M.; Nikbakht, F.; Soleimani, M. The protective role of carnosic acid against beta-amyloid toxicity in rats. Sci. World J. 2013, 2013, 917082. [Google Scholar] [CrossRef]
- Patil, C.S.; Singh, V.P.; Satyanarayan, P.S.; Jain, N.K.; Singh, A.; Kulkarni, S.K. Protective effect of flavonoids against aging- and lipopolysaccharide-induced cognitive impairment in mice. Pharmacology 2003, 69, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.J.; Perry, N.S.; Houghton, P.J. Plants with traditional uses and activities, relevant to the management of Alzheimer’s disease and other cognitive disorders. Phytother. Res. PTR 2003, 17, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin has potent anti-amyloidogenic effects for Alzheimer’s beta-amyloid fibrils in vitro. J. Neurosci. Res. 2004, 75, 742–750. [Google Scholar] [CrossRef]
- Jiang, P.; Li, C.; Xiang, Z.; Jiao, B. Tanshinone IIA reduces the risk of Alzheimer’s disease by inhibiting iNOS, MMP-2 and NF-κBp65 transcription and translation in the temporal lobes of rat models of Alzheimer’s disease. Mol. Med. Rep. 2014, 10, 689–694. [Google Scholar] [CrossRef]
- Teng, Y.; Zhang, M.Q.; Wang, W.; Liu, L.T.; Zhou, L.M.; Miao, S.K.; Wan, L.H. Compound danshen tablet ameliorated aβ25-35-induced spatial memory impairment in mice via rescuing imbalance between cytokines and neurotrophins. BMC Complement. Altern. Med. 2014, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Tulini, F.L.; Souza, V.B.; Thomazini, M.; Silva, M.P.; Massarioli, A.P.; Alencar, S.M.; Pallone, E.M.; Genovese, M.I.; Favaro-Trindade, C.S. Evaluation of the release profile, stability and antioxidant activity of a proanthocyanidin-rich cinnamon (Cinnamomum zeylanicum) extract co-encapsulated with α-tocopherol by spray chilling. Food Res. Int. 2017, 95, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Frydman-Marom, A.; Levin, A.; Farfara, D.; Benromano, T.; Scherzer-Attali, R.; Peled, S.; Vassar, R.; Segal, D.; Gazit, E.; Frenkel, D.; et al. Orally administrated cinnamon extract reduces β-amyloid oligomerization and corrects cognitive impairment in Alzheimer’s disease animal models. PLoS ONE 2011, 6, e16564. [Google Scholar] [CrossRef]
- Mirmosayyeb, O.; Tanhaei, A.; Sohrabi, H.R.; Martins, R.N.; Tanhaei, M.; Najafi, M.A.; Safaei, A.; Meamar, R. Possible Role of Common Spices as a Preventive and Therapeutic Agent for Alzheimer’s Disease. Int. J. Prev. Med. 2017, 8, 5. [Google Scholar] [CrossRef]
- Mashhadi, N.S.; Ghiasvand, R.; Hariri, M.; Askari, G.; Feizi, A.; Darvishi, L.; Hajishafiee, M.; Barani, A. Effect of ginger and cinnamon intake on oxidative stress and exercise performance and body composition in Iranian female athletes. Int. J. Prev. Med. 2013, 4, S31–S35. [Google Scholar]
- Moselhy, S.S.; Ali, H.K. Hepatoprotective effect of cinnamon extracts against carbon tetrachloride induced oxidative stress and liver injury in rats. Biol. Res. 2009, 42, 93–98. [Google Scholar] [CrossRef]
- Amin, K.A.; Abd El-Twab, T.M. Oxidative markers, nitric oxide and homocysteine alteration in hypercholesterolimic rats: Role of atorvastatine and cinnamon. Int. J. Clin. Exp. Med. 2009, 2, 254–265. [Google Scholar]
- Chen, B.J.; Fu, C.S.; Li, G.H.; Wang, X.N.; Lou, H.X.; Ren, D.M.; Shen, T. Cinnamaldehyde Analogues as Potential Therapeutic Agents. Mini Rev. Med. Chem. 2017, 17, 33–43. [Google Scholar] [CrossRef]
- Stavinoha, R.C.; Vattem, D.A. Potential neuroprotective effects of cinnamon. Int. J. Appl. Res. Nat. Prod. 2015, 8, 24–46. [Google Scholar]
- Sheean, P.; Rout, M.K.; Head, R.J.; Bennett, L.E. Modulation of in vitro activity of zymogenic and mature recombinant human β-secretase by dietary plants. FEBS J. 2012, 279, 1291–1305. [Google Scholar] [CrossRef]
- Gu, T.; Wu, W.Y.; Dong, Z.X.; Yu, S.P.; Sun, Y.; Zhong, Y.; Lu, Y.T.; Li, N.G. Development and Structural Modification of BACE1 Inhibitors. Molecules 2016, 22, 4. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls | Screened Plant Extract (n = 5) ± Std. Error | |||||||
|---|---|---|---|---|---|---|---|---|
| Kinetics Properties | NC | PC | J. r | E. c | C. z | S. o | H. p | |
| AChE | Vmax (mM/min) | 0.008651 ± 0.0005743 | 0.00869 ± 0.0004418 | 0.00849 ± 0.0003554 | 0.003548 ± 0.0002209 | 0.004507 ± 0.0004567 | 0.00434 ± 0.0004683 | 0.00469 ± 0.0004768 |
| Km (mM) | 0.003523 ± 0.001149 | 0.01441 ± 0.002464 | 0.01092 ± 0.001659 | 0.001344 ± 0.0006181 | 0.01951 ± 0.006144 | 0.01644 ± 0.005772 | 0.00246 ± 0.001398 | |
| Proposed Inhibitory type | N/A | competitive | competitive | mixed | mixed | mixed | uncompetitive | |
| BuChE | Vmax (mM/min) | 0.00753 ± 0.0007588 | 0.00565 ± 0.001249 | 0.00391 ± 0.0004516 | 0.002214 ± 0.0002146 | 0.005219 ± 0.0008111 | 0.00372 ± 0.0003888 | 0.00383 ± 0.0006241 |
| Km (mM) | 0.09534 ± 0.03577 | 0.43380 ± 0.2367 | 0.0938 ± 0.04096 | 0.02058 ± 0.01200 | 0.3486 ± 0.1430 | 0.14000 ± 0.04964 | 0.09475 ± 0.05747 | |
| Proposed Inhibitory type | N/A | mixed | uncompetitive | mixed | mixed | mixed | uncompetitive | |
| Controls | Screened Plant Extract (n = 4) | ||||||
|---|---|---|---|---|---|---|---|
| Kinetics Properties | NC | PC | J. r | E. c | F. t | S. o | |
| BACE-1 | Vmax (mM/min) | 361.9 | 361.7 | 276.5 | 351.0 | 361.3 | 120.3 |
| Km (mM) | 2.881 | 6.062 | 6.893 | 4.627 | 4.82 | 0.9230 | |
| Proposed Inhibitory type | N/A | competitive | mixed | mixed | competitive | uncompetitive | |
| IC50 (mg/mL) | N/A | 25 µM | 0.55 | 0.60 | 0.81 | 0.53 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Althobaiti, N.A.; Menaa, F.; Dalzell, J.J.; Albalawi, A.E.; Ismail, H.; Alghuthaymi, M.A.; Aldawsari, R.D.; Iqbal, H.; McAlinney, C.; Green, B.D. Ethnomedicinal Plants with Protective Effects against Beta-Amyloid Peptide (Aβ)1-42 Indicate Therapeutic Potential in a New In Vivo Model of Alzheimer’s Disease. Antioxidants 2022, 11, 1865. https://doi.org/10.3390/antiox11101865
Althobaiti NA, Menaa F, Dalzell JJ, Albalawi AE, Ismail H, Alghuthaymi MA, Aldawsari RD, Iqbal H, McAlinney C, Green BD. Ethnomedicinal Plants with Protective Effects against Beta-Amyloid Peptide (Aβ)1-42 Indicate Therapeutic Potential in a New In Vivo Model of Alzheimer’s Disease. Antioxidants. 2022; 11(10):1865. https://doi.org/10.3390/antiox11101865
Chicago/Turabian StyleAlthobaiti, Norah A., Farid Menaa, Johnathan J. Dalzell, Aishah E. Albalawi, Hammad Ismail, Mousa A. Alghuthaymi, Reem D. Aldawsari, Haroon Iqbal, Claire McAlinney, and Brian D. Green. 2022. "Ethnomedicinal Plants with Protective Effects against Beta-Amyloid Peptide (Aβ)1-42 Indicate Therapeutic Potential in a New In Vivo Model of Alzheimer’s Disease" Antioxidants 11, no. 10: 1865. https://doi.org/10.3390/antiox11101865