
Mechanistic Basis and Clinical Evidence for the Applications of Nicotinamide (Niacinamide) to Control Skin Aging and Pigmentation


Abstract
:1. Introduction
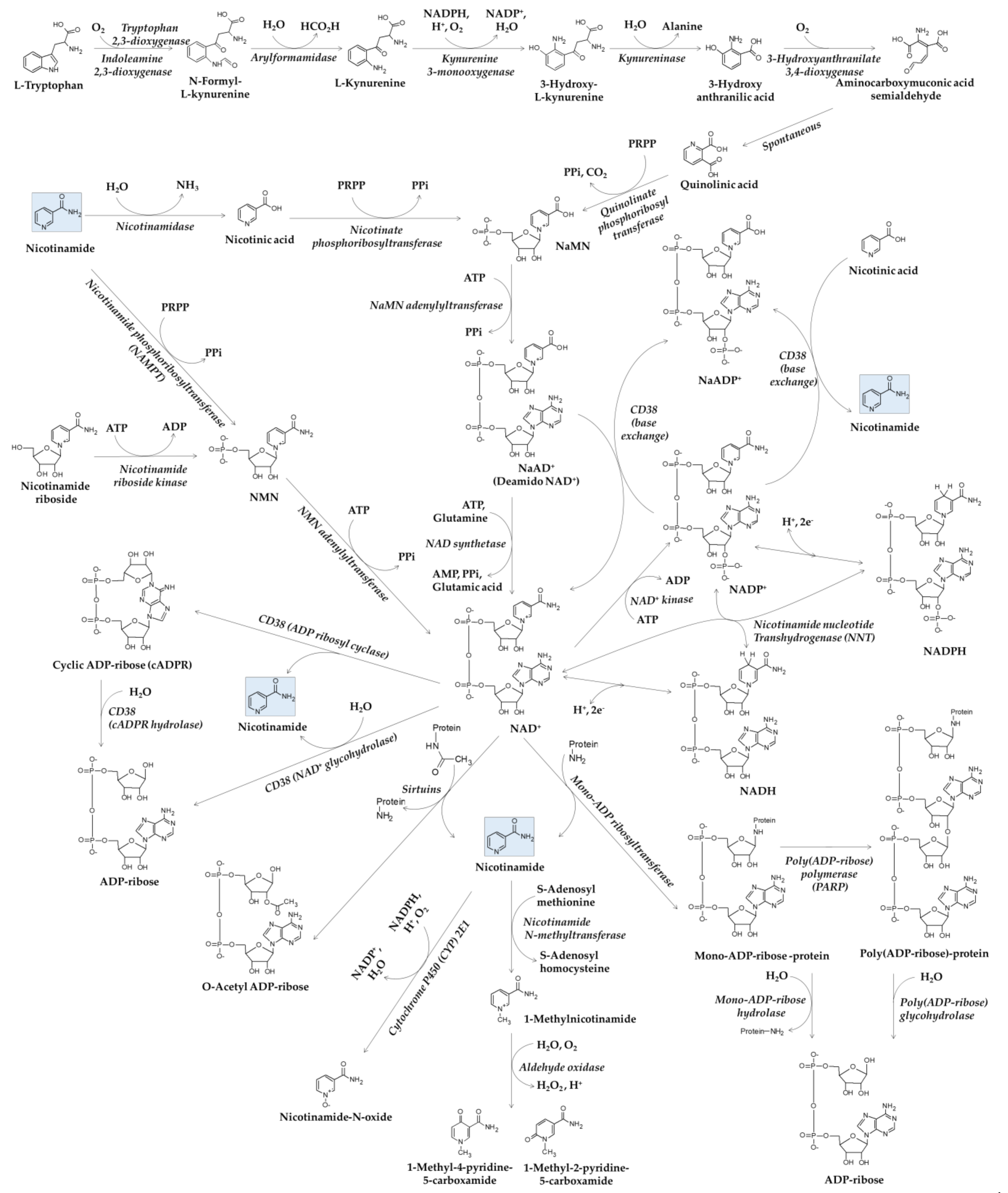
2. Metabolism of Nicotinamide
2.1. Synthesis and Function of NAD(H) and NADP(H)
2.2. Metabolisms of NAD+ and Nicotinamide
3. Antioxidant and Anti-Inflammatory Effects of Nicotinamide
3.1. Antioxidant Properties of Nicotinamide
3.2. Protective Effect of Nicotinamide in Cells Exposed to Environmental Stressors
3.3. Anti-Inflammatory Effects of Nicotinamide
3.4. Anti-Inflammatory Effects of N-Methylnicotinamide and NMN
4. Modulation of Cell Senescence and Epidermal Differentiation by Nicotinamide
4.1. Differential Effects of Nicotinamide on Lifespans of Yeast and Mammalian Cells
4.2. Antisenescence Effects of Nicotinamide
4.3. Epidermal Stem Cells
4.4. Modulation of Epidermal Differentiation by Nicotinamide
5. Enhancement of ECM and Skin Barrier by Nicotinamide
5.1. Changes of ECM by Skin Aging
5.2. Effects of Nicotinamide on Collagen and Other ECMs
5.3. Enhancement of Skin Barrier by Nicotinamide
6. Regulation of Pigmentary Process by Nicotinamide
6.1. Effects of Nicotinamide on Melanogenesis vs. Melanosome Transfer
6.2. Mechanisms for Melanosome Transfer
6.3. Modulation of Melanosome Transfer
7. Clinical Evidence for Skin Antiaging Efficacy of Nicotinamide
8. Clinical Evidence for Skin-Lightening Efficacy of Nicotinamide
8.1. Skin-Lightening Efficacy of Nicotinamide
8.2. Skin-Lightening Efficacy of Combined Formulations of Nicotinamide and Other Active Ingredients
9. Discussion
9.1. Cosmetic Benefits and Side Effects of Topical Nicotinamide
9.2. Mechanisms of Skin Antiaging Action of Nicotinamide
9.3. Mechanisms of Skin Depigmenting Action of Nicotinamide
9.4. Questions to Be Answered in Future Studies
- ●
- Is there any specific molecular target of nicotinamide for the control of skin aging or pigmentation?
- ●
- Is the antiaging effect of nicotinamide is due to its intrinsic property or its metabolites?
- ●
- How does nicotinamide modulate the cell-to-cell interactions in the skin?
- ●
- Whether and how does nicotinamide regulate PAR-2 or NNT involved in skin pigmentation?
- ●
- Does nicotinamide supplementation affect the NADPH/NADP+ ratio and the GSH/GSSG ratio in the skin?
- ●
- Would it be more effective if the dose of nicotinamide is adjusted according to the NAD+ pool level, which varies depending on the individual skin condition?
- ●
- How does nicotinamide affect the self-renewal, proliferation, differentiation, senescence, and eventual exhaustion of epidermal stem cells?
10. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ADP | adenosine diphosphate |
| AMP | adenosine monophosphate |
| ATP | adenosine triphosphate |
| cADPR | cyclic ADP ribose |
| CYP | cytochrome P450 |
| ECM | extracellular matrix |
| GSH | glutathione |
| GSSG | glutathione disulfide |
| IL | interleukin |
| MAPK | mitogen-activated protein kinase |
| MASI | melasma area and severity index |
| MMP | matrix metalloproteinase |
| NaAD + | nicotinic acid adenine dinucleotide |
| NaADP + | nicotinic acid adenine dinucleotide phosphate |
| NAD + | nicotinamide adenine dinucleotide |
| NADH | reduced nicotinamide adenine dinucleotide |
| NADP + | nicotinamide adenine dinucleotide phosphate |
| NADPH | reduced nicotinamide adenine dinucleotide phosphate |
| NaMN | nicotinic acid mononucleotide |
| NAMPT | nicotinamide phosphoribosyltransferase |
| NF-κB | nuclear factor-κB |
| NMN | nicotinamide mononucleotide |
| NNT | nicotinamide nucleotide transhydrogenase |
| PAR-2 | protease-activated receptor-2 |
| PARP | poly(ADP-ribose) polymerase |
| PG | prostaglandin |
| PM | particulate matter |
| PPi | inorganic pyrophosphate |
| PRPP | phosphoribosyl pyrophosphate |
| ROS | reactive oxygen species |
| sir2 | silent information regulator-2 |
| SIRT1 | sirtruin1 |
| SPF | sun protection factor |
| TGF-β | transforming growth factor-β |
| TNF-α | tumor necrosis factor-α |
| UV | ultraviolet |
References
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Rittie, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef] [PubMed]
- Tzaphlidou, M. The role of collagen and elastin in aged skin: An image processing approach. Micron 2004, 35, 173–177. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Shah, A.A.; Sinha, A.A. Oxidative stress and autoimmune skin disease. Eur. J. Dermatol. 2013, 23, 5–13. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative stress in the pathogenesis of skin disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [Green Version]
- Awad, F.; Assrawi, E.; Louvrier, C.; Jumeau, C.; Giurgea, I.; Amselem, S.; Karabina, S.A. Photoaging and skin cancer: Is the inflammasome the missing link? Mech. Ageing Dev. 2018, 172, 131–137. [Google Scholar] [CrossRef]
- Boo, Y.C. Natural Nrf2 Modulators for Skin Protection. Antioxidants 2020, 9, 812. [Google Scholar] [CrossRef]
- Baek, J.; Lee, M.G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef]
- Hegyi, J.; Schwartz, R.A.; Hegyi, V. Pellagra: Dermatitis, dementia, and diarrhea. Int. J. Dermatol. 2004, 43, 1–5. [Google Scholar] [CrossRef]
- Kirkland, J.B. Niacin Status, NAD Distribution and ADP-Ribose Metabolism. Curr. Pharm. Des. 2009, 15, 3–11. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, L.F.; Gossmann, T.I.; Ziegler, M.; Schuster, S. Pathway analysis of NAD+ metabolism. Biochem. J. 2011, 439, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Mattiussi, A.J.; Blais, D. Niacin Versus Niacinamide. Can. Med Assoc. J. 1992, 147, 990–991. [Google Scholar]
- MacKay, D.; Hathcock, J.; Guarneri, E. Niacin: Chemical forms, bioavailability, and health effects. Nutr. Rev. 2012, 70, 357–366. [Google Scholar] [CrossRef]
- Surjana, D.; Damian, D.L. Nicotinamide in dermatology and photoprotection. Skinmed 2011, 9, 360–365. [Google Scholar] [PubMed]
- Forbat, E.; Al-Niaimi, F.; Ali, F.R. Use of nicotinamide in dermatology. Clin. Exp. Dermatol. 2017, 42, 137–144. [Google Scholar] [CrossRef]
- Ballotti, R.; Healy, E.; Bertolotto, C. Nicotinamide as a chemopreventive therapy of skin cancers. Too much of good thing? Pigment. Cell Melanoma Res. 2019, 32, 601–602. [Google Scholar] [CrossRef]
- Snaidr, V.A.; Damian, D.L.; Halliday, G.M. Nicotinamide for photoprotection and skin cancer chemoprevention: A review of efficacy and safety. Exp. Dermatol. 2019, 28 (Suppl. S1), 15–22. [Google Scholar] [CrossRef] [Green Version]
- Hakozaki, T.; Minwalla, L.; Zhuang, J.; Chhoa, M.; Matsubara, A.; Miyamoto, K.; Greatens, A.; Hillebrand, G.G.; Bissett, D.L.; Boissy, R.E. The effect of niacinamide on reducing cutaneous pigmentation and suppression of melanosome transfer. Br. J. Dermatol. 2002, 147, 20–31. [Google Scholar] [CrossRef]
- Bissett, D.L.; Miyamoto, K.; Sun, P.; Li, J.; Berge, C.A. Topical niacinamide reduces yellowing, wrinkling, red blotchiness, and hyperpigmented spots in aging facial skin. Int. J. Cosmet. Sci. 2004, 26, 231–238. [Google Scholar] [CrossRef]
- Bissett, D.L.; Oblong, J.E.; Berge, C.A. Niacinamide: A B vitamin that improves aging facial skin appearance. Dermatol. Surg. 2005, 31, 860–865; discussion 865. [Google Scholar] [CrossRef]
- Otte, N.; Borelli, C.; Korting, H.C. Nicotinamide—Biologic actions of an emerging cosmetic ingredient. Int. J. Cosmet. Sci. 2005, 27, 255–261. [Google Scholar] [CrossRef]
- Braidy, N.; Berg, J.; Clement, J.; Khorshidi, F.; Poljak, A.; Jayasena, T.; Grant, R.; Sachdev, P. Role of Nicotinamide Adenine Dinucleotide and Related Precursors as Therapeutic Targets for Age-Related Degenerative Diseases: Rationale, Biochemistry, Pharmacokinetics, and Outcomes. Antioxid. Redox Signal. 2019, 30, 251–294. [Google Scholar] [CrossRef]
- Fukuwatari, T.; Shibata, K. Nutritional aspect of tryptophan metabolism. Int. J. Tryptophan Res. 2013, 6, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Shibata, K. Organ Co-Relationship in Tryptophan Metabolism and Factors That Govern the Biosynthesis of Nicotinamide from Tryptophan. J. Nutr. Sci. Vitaminol. 2018, 64, 90–98. [Google Scholar] [CrossRef]
- Kennedy, B.E.; Sharif, T.; Martell, E.; Dai, C.; Kim, Y.; Lee, P.W.K.; Gujar, S.A. NAD+ salvage pathway in cancer metabolism and therapy. Pharmacol. Res. 2016, 114, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, P.M.; Bansal, N.; Kerrigan, J.E.; Abali, E.E.; Scotto, K.W.; Bertino, J.R. NAD+ Kinase as a Therapeutic Target in Cancer. Clin. Cancer Res. 2016, 22, 5189–5195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Padayatti, P.S.; Leung, J.H. Proton-Translocating Nicotinamide Nucleotide Transhydrogenase: A Structural Perspective. Front. Physiol. 2017, 8, 1089. [Google Scholar] [CrossRef] [Green Version]
- Chini, C.C.S.; Zeidler, J.D.; Kashyap, S.; Warner, G.; Chini, E.N. Evolving concepts in NAD+ metabolism. Cell Metab. 2021, 33, 1076–1087. [Google Scholar] [CrossRef]
- Schweiker, S.S.; Tauber, A.L.; Sherry, M.E.; Levonis, S.M. Structure, Function and Inhibition of Poly(ADP-ribose)polymerase, Member 14 (PARP14). Mini Rev. Med. Chem. 2018, 18, 1659–1669. [Google Scholar] [CrossRef]
- Palazzo, L.; Mikolcevic, P.; Mikoc, A.; Ahel, I. ADP-ribosylation signalling and human disease. Open Biol. 2019, 9, 190041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassa, P.O.; Haenni, S.S.; Elser, M.; Hottiger, M.O. Nuclear ADP-ribosylation reactions in mammalian cells: Where are we today and where are we going? Microbiology and Molecular Biology Reviews 2006, 70, 789-+. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, J.C.; Li, L.S.; Fattah, F.J.; Dong, Y.; Bey, E.A.; Patel, M.; Geo, J.M.; Boothman, D.A. Review of Poly (ADP-ribose) Polymerase (PARP) Mechanisms of Action and Rationale for Targeting in Cancer and Other Diseases. Crit. Rev. Eukaryot. Gene Expr. 2014, 24, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Watroba, M.; Dudek, I.; Skoda, M.; Stangret, A.; Rzodkiewicz, P.; Szukiewicz, D. Sirtuins, epigenetics and longevity. Ageing Res. Rev. 2017, 40, 11–19. [Google Scholar] [CrossRef]
- Klein, M.A.; Denu, J.M. Biological and catalytic functions of sirtuin 6 as targets for small-molecule modulators. J. Biol. Chem. 2020, 295, 11021–11041. [Google Scholar] [CrossRef]
- Chang, H.C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.; Mehrotra, S.; Chatterjee, S. CD38: T Cell Immuno-Metabolic Modulator. Cells 2020, 9, 1716. [Google Scholar] [CrossRef]
- Aarhus, R.; Graeff, R.M.; Dickey, D.M.; Walseth, T.F.; Lee, H.C. ADP-ribosyl cyclase and CD38 catalyze the synthesis of a calcium-mobilizing metabolite from NADP+. J. Biol. Chem. 1995, 270, 30327–30333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, T.S.; Park, D.R.; Rah, S.Y.; Woo, T.G.; Chung, H.T.; Brenner, C.; Kim, U.H. Interleukin-8 drives CD38 to form NAADP from NADP+ and NAAD in the endolysosomes to mobilize Ca2+ and effect cell migration. FASEB J. 2020, 34, 12565–12576. [Google Scholar] [CrossRef]
- Yu, P.L.; Cai, X.B.; Liang, Y.; Wang, M.X.; Yang, W. Roles of NAD+ and Its Metabolites Regulated Calcium Channels in Cancer. Molecules 2020, 25, 4826. [Google Scholar] [CrossRef]
- Gul, R.; Park, D.R.; Shawl, A.I.; Im, S.Y.; Nam, T.S.; Lee, S.H.; Ko, J.K.; Jang, K.Y.; Kim, D.; Kim, U.H. Nicotinic Acid Adenine Dinucleotide Phosphate (NAADP) and Cyclic ADP-Ribose (cADPR) Mediate Ca2+ Signaling in Cardiac Hypertrophy Induced by beta-Adrenergic Stimulation. PLoS ONE 2016, 11, e0149125. [Google Scholar] [CrossRef]
- Pissios, P. Nicotinamide N-Methyltransferase: More Than a Vitamin B3 Clearance Enzyme. Trends Endocrinol. Metab. 2017, 28, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Felsted, R.L.; Chaykin, S. N1-methylnicotinamide oxidation in a number of mammals. J. Biol. Chem. 1967, 242, 1274–1279. [Google Scholar] [CrossRef]
- Real, A.M.; Hong, S.Y.; Pissios, P. Nicotinamide N-Oxidation by CYP2E1 in Human Liver Microsomes. Drug Metab. Dispos. 2013, 41, 550–553. [Google Scholar] [CrossRef] [Green Version]
- Nadzhimutdinov, K.N.; Mavlianov, I.R.; Umarov, E.F.; Mutalov, N.K. The effect of alpha-tocopherol and nicotinamide on lipid peroxidation and the activity of the antioxidant system in the lung tissue of premature rat pups. Eksp. Klin. Farmakol. 1993, 56, 28–30. [Google Scholar]
- Legon’kova, L.F.; Bushma, M.I.; Zverinskii, I.V.; Abakumov, G.Z.; Zavodnik, L.V. The effect of nicotinamide, methionine and alpha-tocopherol on the liver conjugating and mono-oxygenase systems and on lipid peroxidation in hepatosis-hepatitis in rats. Eksp. Klin. Farmakol. 1997, 60, 68–71. [Google Scholar]
- Velykyi, M.M.; Burda, V.A.; Biront, N.V.; Oliiarnyk, O.D.; Velykyi, A.M. The effect of nicotinamide on the enzymatic activity of the antioxidant defense in experimental diabetes. Ukr. Biokhimicheskii Zhurnal (1978) 1996, 68, 109–114. [Google Scholar]
- Kamat, J.P.; Devasagayam, T.P. Methylene blue plus light-induced lipid peroxidation in rat liver microsomes: Inhibition by nicotinamide (vitamin B3) and other antioxidants. Chem. Biol. Interact. 1996, 99, 1–16. [Google Scholar] [CrossRef]
- Kamat, J.P.; Devasagayam, T.P.A. Nicotinamide (vitamin B-3) as an effective antioxidant against oxidative damage in rat brain mitochondria. Redox Rep. 1999, 4, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Izdebska, M.; Halas-Wisniewska, M.; Adamczyk, I.; Lewandowska, I.; Kwiatkowska, I.; Gagat, M.; Grzanka, A. The protective effect of niacinamide on CHO AA8 cell line against ultraviolet radiation in the context of main cytoskeletal proteins. Adv. Clin. Exp. Med. 2018, 27, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, G.; Garvey, D.R.; Singh, C.K.; Mintie, C.A.; Ahmad, N. Effects and Mechanism of Nicotinamide Against UVA- and/or UVB-mediated DNA Damages in Normal Melanocytes. Photochem. Photobiol. 2019, 95, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Zhen, A.X.; Piao, M.J.; Kang, K.A.; Fernando, P.D.S.M.; Kang, H.K.; Koh, Y.S.; Yi, J.M.; Hyun, J.W. Niacinamide Protects Skin Cells from Oxidative Stress Induced by Particulate Matter. Biomol. Ther. 2019, 27, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Mi, T.Y.; Dong, Y.Y.; Santhanam, U.; Huang, N. Niacinamide and 12-hydroxystearic acid prevented benzo(a)pyrene and squalene peroxides induced hyperpigmentation in skin equivalent. Exp. Dermatol. 2019, 28, 742–746. [Google Scholar] [CrossRef]
- Grange, P.A.; Raingeaud, J.; Calvez, V.; Dupin, N. Nicotinamide inhibits Propionibacterium acnes-induced IL-8 production in keratinocytes through the NF-kappaB and MAPK pathways. J. Dermatol. Sci. 2009, 56, 106–112. [Google Scholar] [CrossRef]
- Monfrecola, G.; Gaudiello, F.; Cirillo, T.; Fabbrocini, G.; Balato, A.; Lembo, S. Nicotinamide downregulates gene expression of interleukin-6, interleukin-10, monocyte chemoattractant protein-1, and tumour necrosis factor-alpha gene expression in HaCaT keratinocytes after ultraviolet B irradiation. Clin. Exp. Dermatol. 2013, 38, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Bierman, J.C.; Laughlin, T.; Tamura, M.; Hulette, B.; Mack, C.E.; Sherrill, J.D.; Tan, C.Y.R.; Morenc, M.; Bellanger, S.; Oblong, J.E. Niacinamide mitigates SASP-related inflammation induced by environmental stressors in human epidermal keratinocytes and skin. Int. J. Cosmet. Sci. 2020, 42, 501–511. [Google Scholar] [CrossRef]
- Bryniarski, K.; Biedron, R.; Jakubowski, A.; Chlopicki, S.; Marcinkiewicz, J. Anti-inflammatory effect of 1-methylnicotinamide in contact hypersensitivity to oxazolone in mice; involvement of prostacyclin. Eur. J. Pharmacol. 2008, 578, 332–338. [Google Scholar] [CrossRef]
- Biedron, R.; Ciszek, M.; Tokarczyk, M.; Bobek, M.; Kurnyta, M.; Slominska, E.M.; Smolenski, R.T.; Marcinkiewicz, J. 1-Methylnicotinamide and nicotinamide: Two related anti-inflammatory agents that differentially affect the functions of activated macrophages. Arch. Immunol. Ther. Exp. 2008, 56, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Wozniacka, A.; Wieczorkowska, M.; Gebicki, J.; Sysa-Jedrzejowska, A. Topical application of 1-methylnicotinamide in the treatment of rosacea: A pilot study. Clin. Exp. Dermatol. 2005, 30, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, L.; Mogielnicki, A.; Buczko, W. Nicotinamide and its metabolite N-methylnicotinamide increase skin vascular permeability in rats. Clin. Exp. Dermatol. 2009, 34, 380–384. [Google Scholar] [CrossRef]
- Jiang, N.; Wang, M.; Song, J.Y.; Liu, Y.G.; Chen, H.; Mu, D.; Xia, M. N-methylnicotinamide protects against endothelial dysfunction and attenuates atherogenesis in apolipoprotein E-deficient mice. Mol. Nutr. Food Res. 2016, 60, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Huynh, P.K.; Wilder, J.; Hiller, S.; Hagaman, J.; Takahashi, N.; Maeda-Smithies, N.; Li, F. Beneficial effects of nicotinamide on hypertensive mice with impaired endothelial nitric oxide function. J. Exp. Nephrol. 2020, 1, 1–8. [Google Scholar] [PubMed]
- Zhou, X.R.; Du, H.H.; Ni, L.Y.; Ran, J.; Hu, J.; Yu, J.J.; Zhao, X. Nicotinamide Mononucleotide Combined with Lactobacillus fermentum TKSN041 Reduces the Photoaging Damage in Murine Skin by Activating AMPK Signaling Pathway. Front. Pharmacol. 2021, 12, 643089. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Bitterman, K.J.; Wood, J.G.; Medvedik, O.; Sinclair, D.A. Nicotinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature 2003, 423, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Matuoka, K.; Chen, K.Y.; Takenawa, T. Rapid reversion of aging phenotypes by nicotinamide through possible modulation of histone acetylation. Cell. Mol. Life Sci. 2001, 58, 2108–2116. [Google Scholar] [CrossRef]
- Lim, C.S.; Potts, M.; Helm, R.E. Nicotinamide extends the replicative life span of primary human cells. Mech. Ageing Dev. 2006, 127, 511–514. [Google Scholar] [CrossRef]
- Porcu, M.; Chiarugi, A. The emerging therapeutic potential of sirtuin-interacting drugs: From cell death to lifespan extension. Trends Pharmacol. Sci. 2005, 26, 94–103. [Google Scholar] [CrossRef]
- Adams, J.D.; Klaidman, L.K. Sirtuins, nicotinamide and aging: A critical review. Lett. Drug Des. Discov. 2007, 4, 44–48. [Google Scholar] [CrossRef]
- Massudi, H.; Grant, R.; Braidy, N.; Guest, J.; Farnsworth, B.; Guillemin, G.J. Age-Associated Changes In Oxidative Stress and NAD+ Metabolism In Human Tissue. PLoS ONE 2012, 7, e42357. [Google Scholar] [CrossRef]
- Oblong, J.E. The evolving role of the NAD plus/nicotinamide metabolome in skin homeostasis, cellular bioenergetics, and aging. DNA Repair 2014, 23, 59–63. [Google Scholar] [CrossRef]
- Kang, H.T.; Il Lee, H.; Hwang, E.S. Nicotinamide extends replicative lifespan of human cells. Aging Cell 2006, 5, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y.; Ham, H.J.; Kim, C.M.; Hwang, E.S. Nicotinamide exerts antioxidative effects on senescent cells. Mol. Cells 2015, 38, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Oblong, J.E.; Bowman, A.; Rovito, H.A.; Jarrold, B.B.; Sherrill, J.D.; Black, M.R.; Nelson, G.; Kimball, A.B.; Birch-Machin, M.A. Metabolic dysfunction in human skin: Restoration of mitochondrial integrity and metabolic output by nicotinamide (niacinamide) in primary dermal fibroblasts from older aged donors. Aging Cell 2020, 19, e13248. [Google Scholar] [CrossRef]
- Borradaile, N.M.; Pickering, J.G. Nicotinamide phosphoribosyltransferase imparts human endothelial cells with extended replicative lifespan and enhanced angiogenic capacity in a high glucose environment. Aging Cell 2009, 8, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Rovito, H.A.; Oblong, J.E. Nicotinamide preferentially protects glycolysis in dermal fibroblasts under oxidative stress conditions. Br. J. Dermatol. 2013, 169, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Khaidizar, F.D.; Nakahata, Y.; Kume, A.; Sumizawa, K.; Kohno, K.; Matsui, T.; Bessho, Y. Nicotinamide phosphoribosyltransferase delays cellular senescence by upregulating SIRT1 activity and antioxidant gene expression in mouse cells. Genes Cells 2017, 22, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.L.; Chin, T.; Tan, C.Y.R.; Rovito, H.A.; Quek, L.S.; Oblong, J.E.; Bellanger, S. Nicotinamide Metabolism Modulates the Proliferation/Differentiation Balance and Senescence of Human Primary Keratinocytes. J. Investig. Dermatol. 2019, 139, 1638–1647.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taub, A.F.; Pham, K. Stem Cells in Dermatology and Anti-aging Care of the Skin. Facial Plast. Surg. Clin. N. Am. 2018, 26, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Adjaye, J.; Akamatsu, H.; Moe-Behrens, G.; Niemann, C. Human skin stem cells and the ageing process. Exp. Gerontol. 2008, 43, 986–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Matsumura, H.; Kato, T.; Ichinose, S.; Takada, A.; Namiki, T.; Asakawa, K.; Morinaga, H.; Mohri, Y.; De Arcangelis, A.; et al. Stem cell competition orchestrates skin homeostasis and ageing. Nature 2019, 568, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Gannon, H.S.; Donehower, L.A.; Lyle, S.; Jones, S.N. Mdm2-p53 signaling regulates epidermal stem cell senescence and premature aging phenotypes in mouse skin. Dev. Biol. 2011, 353, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chu, G.Y.; Chen, Y.F.; Chen, H.Y.; Chan, M.H.; Gau, C.S.; Weng, S.M. Stem cell therapy on skin: Mechanisms, recent advances and drug reviewing issues. J. Food Drug Anal. 2018, 26, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Mayack, B.K.; Sippl, W.; Ntie-Kang, F. Natural Products as Modulators of Sirtuins. Molecules 2020, 25, 3287. [Google Scholar] [CrossRef]
- Meng, Y.; Ren, Z.; Xu, F.; Zhou, X.; Song, C.; Wang, V.Y.; Liu, W.; Lu, L.; Thomson, J.A.; Chen, G. Nicotinamide Promotes Cell Survival and Differentiation as Kinase Inhibitor in Human Pluripotent Stem Cells. Stem Cell Rep. 2018, 11, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [Green Version]
- Quan, T.H.; Fisher, G.J. Role of Age-Associated Alterations of the Dermal Extracellular Matrix Microenvironment in Human Skin Aging: A Mini-Review. Gerontology 2015, 61, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghahary, A.; Marcoux, Y.; Karimi-Busheri, F.; Li, Y.; Tredget, E.E.; Kilani, R.T.; Lam, E.; Weinfeld, M. Differentiated keratinocyte-releasable stratifin (14-3-3 sigma) stimulates MMP-1 expression in dermal fibroblasts. J. Investig. Dermatol. 2005, 124, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Lam, E.; Kilani, R.T.; Li, Y.; Tredget, E.E.; Ghahary, A. Stratifin-induced matrix metalloproteinase-1 in fibroblast is mediated by c-fos and p38 mitogen-activated protein kinase activation. J. Investig. Dermatol. 2005, 125, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, H.; Murakami, Y.; Tanaka, H.; Nakata, S. Increase of stratifin triggered by ultraviolet irradiation is possibly related to premature aging of human skin. Exp. Dermatol. 2014, 23 (Suppl. S1), 32–36. [Google Scholar] [CrossRef]
- Seok, J.K.; Boo, Y.C. p-Coumaric Acid Attenuates UVB-Induced Release of Stratifin from Keratinocytes and Indirectly Regulates Matrix Metalloproteinase 1 Release from Fibroblasts. Korean J. Physiol. Pharmacol. 2015, 19, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.R.; Gallo, R.L. Glycosaminoglycans and their proteoglycans: Host-associated molecular patterns for initiation and modulation of inflammation. FASEB J. 2006, 20, 9–22. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Oh, J.H.; Chung, J.H. Glycosaminoglycan and proteoglycan in skin aging. J. Dermatol. Sci. 2016, 83, 174–181. [Google Scholar] [CrossRef]
- Ratcliffe, D.R.; Iqbal, J.; Hussain, M.M.; Cramer, E.B. Fibrillar collagen type I stimulation of apolipoprotein B secretion in Caco-2 cells is mediated by beta1 integrin. Biochim. Biophys. Acta 2009, 1791, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Chalensouk-Khaosaat, J.; Gonzalez, S. Simulation of the Elastin and Fibrillin in Non-Irradiated or UVA Radiated Fibroblasts, and Direct Inhibition of Elastase or Matrix Metalloptoteinases Activity by Nicotinamide or Its Derivatives. J. Cosmet. Sci. 2018, 69, 47–56. [Google Scholar]
- Wessels, Q.; Pretorius, E.; Smith, C.M.; Nel, H. The potential of a niacinamide dominated cosmeceutical formulation on fibroblast activity and wound healing in vitro. Int. Wound J. 2014, 11, 152–158. [Google Scholar] [CrossRef]
- Ashkani Esfahani, S.; Khoshneviszadeh, M.; Namazi, M.R.; Noorafshan, A.; Geramizadeh, B.; Nadimi, E.; Razavipour, S.T. Topical Nicotinamide Improves Tissue Regeneration in Excisional Full-Thickness Skin Wounds: A Stereological and Pathological Study. Trauma Mon. 2015, 20, e18193. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.H. Aging of the skin barrier. Clin. Dermatol. 2019, 37, 336–345. [Google Scholar] [CrossRef]
- Tanno, O.; Ota, Y.; Kitamura, N.; Katsube, T.; Inoue, S. Nicotinamide increases biosynthesis of ceramides as well as other stratum corneum lipids to improve the epidermal permeability barrier. Br. J. Dermatol. 2000, 143, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Bissett, D. Topical niacinamide and barrier enhancement. Cutis 2002, 70, 8–12; discussion 21–23. [Google Scholar] [PubMed]
- Draelos, Z.D.; Ertel, K.; Berge, C. Niacinamide-containing facial moisturizer improves skin barrier and benefits subjects with rosacea. Cutis 2005, 76, 135–141. [Google Scholar]
- Jacobson, E.L.; Kim, H.; Kim, M.; Williams, J.D.; Coyle, D.L.; Coyle, W.R.; Grove, G.; Rizer, R.L.; Stratton, M.S.; Jacobson, M.K. A topical lipophilic niacin derivative increases NAD, epidermal differentiation and barrier function in photodamaged skin. Exp. Dermatol. 2007, 16, 490–499. [Google Scholar] [CrossRef]
- Virador, V.M.; Kobayashi, N.; Matsunaga, J.; Hearing, V.J. A standardized protocol for assessing regulators of pigmentation. Anal. Biochem. 1999, 270, 207–219. [Google Scholar] [CrossRef]
- Lei, T.C.; Virador, V.M.; Vieira, W.D.; Hearing, V.J. A melanocyte-keratinocyte coculture model to assess regulators of pigmentation in vitro. Anal. Biochem. 2002, 305, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Greatens, A.; Hakozaki, T.; Koshoffer, A.; Epstein, H.; Schwemberger, S.; Babcock, G.; Bissett, D.; Takiwaki, H.; Arase, S.; Wickett, R.R.; et al. Effective inhibition of melanosome transfer to keratinocytes by lectins and niacinamide is reversible. Exp. Dermatol. 2005, 14, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Seiberg, M. Keratinocyte-melanocyte interactions during melanosome transfer. Pigment Cell Res. 2001, 14, 236–242. [Google Scholar] [CrossRef]
- Boissy, R.E. Melanosome transfer to and translocation in the keratinocyte. Exp. Dermatol. 2003, 12, 5–12. [Google Scholar] [CrossRef]
- Hearing, V.J. Regulating melanosome transfer: Who’s driving the bus? Pigment Cell Res. 2007, 20, 334–335. [Google Scholar] [CrossRef]
- Wu, X.F.; Hammer, J.A. Melanosome transfer: It is best to give and receive. Curr. Opin. Cell Biol. 2014, 29, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.; Leopardi, S.; Printup, S.; Madden, B.C. Filopodia are conduits for melanosome transfer to keratinocytes. J. Cell Sci. 2002, 115, 1441–1451. [Google Scholar] [CrossRef]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes Are Transferred from Melanocytes to Keratinocytes through the Processes of Packaging, Release, Uptake, and Dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.F.S.; Masedunskas, A.; Weigert, R.; Copeland, N.G.; Jenkins, N.A.; Hammer, J.A. Melanoregulin regulates a shedding mechanism that drives melanosome transfer from melanocytes to keratinocytes. Proc. Natl. Acad. Sci. USA 2012, 109, E2101–E2109. [Google Scholar] [CrossRef] [Green Version]
- Tarafder, A.K.; Bolasco, G.; Correia, M.S.; Pereira, F.J.C.; Iannone, L.; Hume, A.N.; Kirkpatrick, N.; Picardo, M.; Torrisi, M.R.; Rodrigues, I.P.; et al. Rab11b Mediates Melanin Transfer between Donor Melanocytes and Acceptor Keratinocytes via Coupled Exo/Endocytosis. J. Investig. Dermatol. 2014, 134, 1056–1066. [Google Scholar] [CrossRef] [Green Version]
- Santulli, R.J.; Derian, C.K.; Darrow, A.L.; Tomko, K.A.; Eckardt, A.J.; Seiberg, M.; Scarborough, R.M.; Andradegordon, P. Evidence for the Presence of a Protease-Activated Receptor Distinct from the Thrombin Receptor in Human Keratinocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 9151–9155. [Google Scholar] [CrossRef] [Green Version]
- Derian, C.K.; Eckardt, A.J.; Andrade-Gordon, P. Differential regulation of human keratinocyte growth and differentiation by a novel family of protease-activated receptors. Cell Growth Differ. 1997, 8, 743–749. [Google Scholar]
- Seiberg, M.; Paine, C.; Sharlow, E.; Andrade-Gordon, P.; Costanzo, M.; Eisinger, M.; Shapiro, S.S. The protease-activated receptor 2 regulates pigmentation via keratinocyte-melanocyte interactions. Exp. Cell Res. 2000, 254, 25–32. [Google Scholar] [CrossRef]
- Seiberg, M.; Paine, C.; Sharlow, E.; Andrade-Gordon, P.; Costanzo, M.; Eisinger, M.; Shapiro, S.S. Inhibition of melanosome transfer results in skin lightening. J. Investig. Dermatol. 2000, 115, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.; Deng, A.; Rodriguez-Burford, C.; Seiberg, M.; Han, R.J.; Babiarz, L.; Grizzle, W.; Bell, W.; Pentland, A. Protease-activated receptor 2, a receptor involved in melanosome transfer, is upregulated in human skin by ultraviolet irradiation. J. Investig. Dermatol. 2001, 117, 1412–1420. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.Y.; Li, J.; Lin, X.; Wu, W.Y.; Kang, K.F.; Fu, W.W. Oxidation Levels Differentially Impact Melanocytes: Low versus High Concentration of Hydrogen Peroxide Promotes Melanin Synthesis and Melanosome Transfer. Dermatology 2012, 224, 145–153. [Google Scholar] [CrossRef]
- Scott, G.; Leopardi, S.; Printup, S.; Malhi, N.; Seiberg, M.; Lapoint, R. Proteinase-activated receptor-2 stimulates prostaglandin production in keratinocytes: Analysis of prostaglandin receptors on human melanocytes and effects of PGE2 and PGF2alpha on melanocyte dendricity. J. Investig. Dermatol. 2004, 122, 1214–1224. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.J.; Ma, H.Y.; Yang, Y.; Li, P.C.; Zi, S.X.; Jia, C.Y.; Chen, R. alpha-Melanocyte stimulating hormone (MSH) and prostaglandin E2 (PGE2) drive melanosome transfer by promoting filopodia delivery and shedding spheroid granules: Evidences from atomic force microscopy observation. J. Dermatol. Sci. 2014, 76, 222–230. [Google Scholar] [CrossRef]
- Choi, E.J.; Kang, Y.G.; Kim, J.; Hwang, J.K. Macelignan Inhibits Melanosome Transfer Mediated by Protease-Activated Receptor-2 in Keratinocytes. Biol. Pharm. Bull. 2011, 34, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Wang, N.; Gao, L.L.; Li, L.L.; Zheng, S.W.; Liu, Y.J.; Ozukum, M.; Nikiforova, A.; Zhao, G.M.; Song, Z.Q. The effect of the NMDA receptor-dependent signaling pathway on cell morphology and melanosome transfer in melanocytes. J. Dermatol. Sci. 2016, 84, 296–304. [Google Scholar] [CrossRef]
- Hu, Q.M.; Yi, W.J.; Su, M.Y.; Jiang, S.; Xu, S.Z.; Lei, T.C. Induction of retinal-dependent calcium influx in human melanocytes by UVA or UVB radiation contributes to the stimulation of melanosome transfer. Cell Prolif. 2017, 50, e12372. [Google Scholar] [CrossRef]
- Koike, S.; Yamasaki, K.; Yamauchi, T.; Inoue, M.; Shimada-Ohmori, R.; Tsuchiyama, K.; Aiba, S. Toll-like receptors 2 and 3 enhance melanogenesis and melanosome transport in human melanocytes. Pigment Cell Melanoma Res. 2018, 31, 570–584. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Chiu, P.C.; Chan, C.C.; Lin, H.M.; Chiu, H.C. The clinical anti-aging effects of topical kinetin and niacinamide in Asians: A randomized, double-blind, placebo-controlled, split-face comparative trial. J. Cosmet. Dermatol. 2007, 6, 243–249. [Google Scholar] [CrossRef]
- Kawada, A.; Konishi, N.; Oiso, N.; Kawara, S.; Date, A. Evaluation of anti-wrinkle effects of a novel cosmetic containing niacinamide. J. Dermatol. 2008, 35, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.J.J.; Hillebrand, G.G.; Raleigh, P.; Li, J.; Marmor, M.J.; Bertucci, V.; Grimes, P.E.; Mandy, S.H.; Perez, M.I.; Weinkle, S.H.; et al. A randomized, controlled comparative study of the wrinkle reduction benefits of a cosmetic niacinamide/peptide/retinyl propionate product regimen vs. a prescription 0.02% tretinoin product regimen. Br. J. Dermatol. 2010, 162, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berardesca, E.; Ardigo, M.; Cameli, N.; Mariano, M.; Agozzino, M.; Matts, P.J. Randomized, double-blinded, vehicle-controlled, split-face study to evaluate the effects of topical application of a Gold Silk Sericin/Niacinamide/Signaline complex on biophysical parameters related to skin ageing. Int. J. Cosmet. Sci. 2015, 37, 606–612. [Google Scholar] [CrossRef]
- Farris, P.; Zeichner, J.; Berson, D. Efficacy and Tolerability of a Skin Brightening/Anti-Aging Cosmeceutical Containing Retinol 0.5%, Niacinamide, Hexylresorcinol, and Resveratrol. J. Drugs Dermatol. 2016, 15, 863–868. [Google Scholar] [PubMed]
- Lee, Y.I.; Kim, S.; Kim, J.; Kim, J.; Chung, K.B.; Lee, J.H. Randomized controlled study for the anti-aging effect of human adipocyte-derived mesenchymal stem cell media combined with niacinamide after laser therapy. J. Cosmet. Dermatol. 2021, 20, 1774–1781. [Google Scholar] [CrossRef] [PubMed]
- Navarrete-Solis, J.; Castanedo-Cazares, J.P.; Torres-Alvarez, B.; Oros-Ovalle, C.; Fuentes-Ahumada, C.; Gonzalez, F.J.; Martinez-Ramirez, J.D.; Moncada, B. A Double-Blind, Randomized Clinical Trial of Niacinamide 4% versus Hydroquinone 4% in the Treatment of Melasma. Dermatol. Res. Pract. 2011, 2011, 379173. [Google Scholar] [CrossRef]
- Castanedo-Cazares, J.P.; Larraga-Pinones, G.; Ehnis-Perez, A.; Fuentes-Ahumada, C.; Oros-Ovalle, C.; Smoller, B.R.; Torres-Alvarez, B. Topical niacinamide 4% and desonide 0.05% for treatment of axillary hyperpigmentation: A randomized, double-blind, placebo-controlled study. Clin. Cosmet. Investig. Dermatol. 2013, 6, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Pierard, G.E. EEMCO guidance for the assessment of skin colour. J Eur Acad Dermatol Venereol 1998, 10, 1–11. [Google Scholar] [CrossRef]
- Hakozaki, T.; Takiwaki, H.; Miyamoto, K.; Sato, Y.; Arase, S. Ultrasound enhanced skin-lightening effect of vitamin C and niacinamide. Skin Res. Technol. 2006, 12, 105–113. [Google Scholar] [CrossRef]
- Bissett, D.L.; Robinson, L.R.; Raleigh, P.S.; Miyamoto, K.; Hakozaki, T.; Li, J.; Kelm, G.R. Reduction in the appearance of facial hyperpigmentation by topical N-undecyl-10-enoyl-L-phenylalanine and its combination with niacinamide. J. Cosmet. Dermatol. 2009, 8, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Kimball, A.B.; Kaczvinsky, J.R.; Li, J.; Robinson, L.R.; Matts, P.J.; Berge, C.A.; Miyamoto, K.; Bissett, D.L. Reduction in the appearance of facial hyperpigmentation after use of moisturizers with a combination of topical niacinamide and N-acetyl glucosamine: Results of a randomized, double-blind, vehicle-controlled trial. Br. J. Dermatol. 2010, 162, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Viyoch, J.; Tengamnuay, I.; Phetdee, K.; Tuntijarukorn, P.; Waranuch, N. Effects of Trans-4-(Aminomethyl) Cyclohexanecarboxylic Acid/Potassium Azeloyl Diglycinate/Niacinamide Topical Emulsion in Thai Adults With Melasma: A Single-Center, Randomized, Double-Blind, Controlled Study. Curr. Ther. Res. Clin. Exp. 2010, 71, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Oh, I.Y.; Koo, K.T.; Suk, J.M.; Jung, S.W.; Park, J.O.; Kim, B.J.; Choi, Y.M. Reduction in facial hyperpigmentation after treatment with a combination of topical niacinamide and tranexamic acid: A randomized, double-blind, vehicle-controlled trial. Skin Res. Technol. 2014, 20, 208–212. [Google Scholar] [CrossRef]
- Desai, S.; Ayres, E.; Bak, H.; Manco, M.; Lynch, S.; Raab, S.; Du, A.; Green, D.; Skobowiat, C.; Wangari-Talbot, J.; et al. Effect of a Tranexamic Acid, Kojic Acid, and Niacinamide Containing Serum on Facial Dyschromia: A Clinical Evaluation. J. Drugs Dermatol. 2019, 18, 454–459. [Google Scholar]
- Kalasho, B.D.; Minokadeh, A.; Zhang-Nunes, S.; Zoumalan, R.A.; Shemirani, N.L.; Waldman, A.R.; Pletzer, V.; Zoumalan, C.I. Evaluating the Safety and Efficacy of a Topical Formulation Containing Epidermal Growth Factor, Tranexamic Acid, Vitamin C, Arbutin, Niacinamide and Other Ingredients as Hydroquinone 4% Alternatives to Improve Hyperpigmentation: A Prospective, Randomized, Controlled Split Face Study. J. Cosmet. Sci. 2020, 71, 263–290. [Google Scholar]
- Kim, J.H.; Seok, J.K.; Kim, Y.M.; Boo, Y.C. Identification of small peptides and glycinamide that inhibit melanin synthesis using a positional scanning synthetic peptide combinatorial library. Br. J. Dermatol. 2019, 181, 128–137. [Google Scholar] [CrossRef]
- Boo, Y.C. Up- or Downregulation of Melanin Synthesis Using Amino Acids, Peptides, and Their Analogs. Biomedicines 2020, 8, 322. [Google Scholar] [CrossRef]
- Boo, Y.C. p-Coumaric Acid as An Active Ingredient in Cosmetics: A Review Focusing on its Antimelanogenic Effects. Antioxidants 2019, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C. Arbutin as a Skin Depigmenting Agent with Antimelanogenic and Antioxidant Properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.Y.; Choi, S.R.; Moon, I.J.; Park, C.W.; Kim, Y.H.; Chang, S.E. The Glutathione Derivative, GSH Monoethyl Ester, May Effectively Whiten Skin but GSH Does Not. Int. J. Mol. Sci. 2016, 17, 629. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Ha, J.W.; Hwang, Y.J.; Boo, Y.C. Identification of L-Cysteinamide as a Potent Inhibitor of Tyrosinase-Mediated Dopachrome Formation and Eumelanin Synthesis. Antioxidants 2021, 10, 1202. [Google Scholar] [CrossRef]
- Hwang, E.S.; Song, S.B. Possible Adverse Effects of High-Dose Nicotinamide: Mechanisms and Safety Assessment. Biomolecules 2020, 10, 687. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tian, Y.J.; Guo, J.; Sun, W.P.; Lun, Y.Z.; Guo, M.; Luo, N.; Cao, Y.; Cao, J.M.; Gong, X.J.; et al. Nicotinamide supplementation induces detrimental metabolic and epigenetic changes in developing rats. Br. J. Nutr. 2013, 110, 2156–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosmetic Ingredient Review Expert Panel. Final report of the safety assessment of niacinamide and niacin. Int. J. Toxicol. 2005, 24 (Suppl. 5), 1–31. [Google Scholar]
- Wohlrab, J.; Kreft, D. Niacinamide—Mechanisms of Action and Its Topical Use in Dermatology. Skin Pharmacol. Physiol. 2014, 27, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.Y.; Lin, P.; Knoll, E.; Chakrabarti, R. Mechanism of Inhibition of the Human Sirtuin Enzyme SIRT3 by Nicotinamide: Computational and Experimental Studies. PLoS ONE 2014, 9, e107729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banasik, M.; Stedeford, T.; Strosznajder, R.P. Natural Inhibitors of Poly(ADP-ribose) Polymerase-1. Mol. Neurobiol. 2012, 46, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Mathialagan, S.; Bi, Y.A.; Costales, C.; Kalgutkar, A.S.; Rodrigues, A.D.; Varma, M.V.S. Nicotinic acid transport into human liver involves organic anion transporter 2 (SLC22A7). Biochem. Pharmacol. 2020, 174, 113829. [Google Scholar] [CrossRef] [PubMed]
- Grolla, A.A.; Torretta, S.; Gnemmi, I.; Amoruso, A.; Orsomando, G.; Gatti, M.; Caldarelli, A.; Lim, D.; Penengo, L.; Brunelleschi, S.; et al. Nicotinamide phosphoribosyltransferase (NAMPT/PBEF/visfatin) is a tumoural cytokine released from melanoma. Pigment Cell Melanoma Res. 2015, 28, 718–729. [Google Scholar] [CrossRef]
- Grozio, A.; Mills, K.F.; Yoshino, J.; Bruzzone, S.; Sociali, G.; Tokizane, K.; Lei, H.C.; Cunningham, R.; Sasaki, Y.; Migaud, M.E.; et al. Slc12a8 is a nicotinamide mononucleotide transporter. Nat. Metab. 2019, 1, 47–57. [Google Scholar] [CrossRef]
- Khaidizar, F.D.; Bessho, Y.; Nakahata, Y. Nicotinamide Phosphoribosyltransferase as a Key Molecule of the Aging/Senescence Process. Int. J. Mol. Sci. 2021, 22, 3709. [Google Scholar] [CrossRef]
- Kim, H.J.; Kazi, J.U.; Lee, Y.R.; Nguyen, D.H.; Lee, H.B.; Shin, J.H.; Soh, J.W.; Kim, E.K. Visualization of the melanosome transfer-inhibition in a mouse epidermal cell co-culture model. Int. J. Mol. Med. 2010, 25, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Hwang, J.S.; Kim, H.S. N-Nicotinoyl dopamine inhibits skin pigmentation by suppressing of melanosome transfer. Eur. J. Pharmacol. 2015, 769, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Allouche, J.; Rachmin, I.; Adhikari, K.; Pardo, L.M.; Lee, J.H.; McConnell, A.M.; Kato, S.; Fan, S.; Kawakami, A.; Suita, Y.; et al. NNT mediates redox-dependent pigmentation via a UVB- and MITF-independent mechanism. Cell 2021, 184, 4268–4283.e4220. [Google Scholar] [CrossRef]


{kind=link}
| Literature | Study Format | No. of Subjects | Compared Formulations | Treatment | Key Findings |
|---|---|---|---|---|---|
| [23] | A double-blind, placebo-controlled, split-face, left–right randomized clinical study | 50 | An oil-in-water moisturizer (placebo control | To each side of the face was applied each product, twice daily for 12 weeks. | Improved fine lines/wrinkles, hyperpigmentation spots, texture, red blotchiness, and skin yellowing (sallowness) compared to the control in endpoints. |
| 5% nicotinamide | |||||
| [24] | A double-blind, placebo formulation–controlled, split-face study with left–right randomization | 50 | An oil-in-water moisturizer (placebo control | To each side of the face was applied each product, twice daily for 12 weeks. | Reduced fine lines, wrinkles, hyperpigmented spots, red blotchiness, and skin sallowness (yellowing), and increased elasticity (as measured via cutometry). |
| 5% nicotinamide | |||||
| [133] | A randomized, double-blind, placebo-controlled, split-face comparative study | 27 | An aqueous serum | Test serum was applied evenly to one side of the face and vehicle to the other side twice daily for 12 weeks. | Combination of kinetin and nicotinamide reduced pore, wrinkle, unevenness, erythema, and spot at weeks 8 and 12 and increased corneal moisture at week 12. Nicotinamide alone reduced pore and unevenness at week 8 and wrinkle at week 12. |
| 0.03% kinetin + 4% nicotinamide | |||||
| 25 | An aqueous serum | ||||
| 4% nicotinamide | |||||
| [134] | A randomized, placebo-controlled, split-face study | 30 | A vehicle lotion | The test product was applied on wrinkles of one side and a control product on the other side for 8 weeks. | Test product reduced wrinkle grades and average roughness of skin surface (Ra value) in the tested skin area to lower levels compared to pre-application (p < 0.001) and the vehicle control (p < 0.001) in endpoints. |
| 4% nicotinamide | |||||
| [135] | A randomized, parallel-group facial appearance study | 99 | A daytime lotion (SPF 30) containing 5% nicotinamide and peptides; a night cream containing nicotinamide and peptides; a wrinkle treatment containing nicotinamide, peptides, and 0.3% retinyl propionate. | Subjects applied a wrinkle treatment twice daily, a daytime lotion, and the night cream daily. | The cosmetic regimen significantly improved wrinkle appearance after 8 weeks relative to tretinoin in the total population, with comparable benefits in subject cohorts (n = 25) who continued treatment for an additional 16 weeks. |
| 97 | 0.02% tretinoin in an emollient base; a sunscreen (SPF 30) | ||||
| [136] | A randomized, double-blinded, vehicle-controlled, split-face study | 40 | A simple oil-in-water emulsion | Subjects applied the products twice daily to either the left- or right-hand side of their face at 2 mg cm−2. | Test product improved stratum corneum hydration, barrier function, elasticity, and surface topography compared with the vehicle control in endpoints. |
| 2% gold silk sericin, 5% nicotinamide, 0.1% signalineTM (diacylglycerol and fatty alcohols) | |||||
| [137] | An open-label, single-center study | 25 | 0.5% retinol, 4.4% nicotinamide, 1% resveratrol, and 1.1% hexylresorcinol | Treatment at night for 10 weeks. | The formulation improved hyperpigmentation, overall skin clarity, evenness of skin tone, and wrinkles compared to baseline at week 4 and through week 10. |
| [138] | A double-blind, randomized, split-face, vehicle-controlled study | 24 | 2% human adipocyte-derived mesenchymal stem cell-conditioned medium and 2% nicotinamide | Applied twice daily for 3 weeks after fractional ablative CO2 laser treatment. | The formulation reduced the wrinkle index (p = 0.036) and melanin index (p = 0.043) compared to the control group. |
| A vehicle cream |
| Literature | Study Format | No. of Subjects | Compared Formulations | Treatment | Key Findings |
|---|---|---|---|---|---|
| [22] | A randomized, split-face, double-blind, paired clinical study | 18 | 5% nicotinamide | Subjects applied a test or a control moisturizer to each side of the face twice daily for 8 weeks. | The side of the face receiving the test moisturizer showed a significant decrease in the total hyperpigmented area measured by image analysis and a reduction in visually assessed hyperpigmentation degree compared to the side receiving the control moisturizer after 4 weeks or 8 weeks of treatment. |
| A control moisturizer | |||||
| A randomized, split-face, double-blind, round-robin design | 120 | A vehicle moisturizer | Applied two of three different products, each product to each side of their face twice daily for 8 weeks. | After 4 weeks, L* value of the treated sides was highest with a test sunscreen moisturizer, followed by a control sunscreen moisturizer, and a vehicle moisturizer. | |
| A control sunscreen moisturizer (SPF 15) | |||||
| A test sunscreen moisturizer containing 2% nicotinamide | |||||
| [111] | A double-blinded, randomized, vehicle-controlled, split-face design human clinical trial | 39 | 5% nicotinamide | Subjects applied either test or control product to the assigned sides of their faces twice a day for 8 weeks. | 5% Nicotinamide-containing moisturizer demonstrated a higher reduction in hyperpigmented spot than the vehicle moisturizer, after 4 and 8 weeks of treatment. 2% Nicotinamide did not show a statistically significant effect compared to the vehicle moisturizer. |
| A vehicle moisturizer | |||||
| 40 | 2% nicotinamide | ||||
| A vehicle moisturizer | |||||
| [139] | A double-blind, randomized, clinical trial | 27 | 4% nicotinamide | Melasma patients applied a product on the left side of the face and the other on the right side for 8 weeks. | After 8 weeks of treatment, MASI score was decreased, L* value was increased and a* value was unchanged by both treatments compared to the baseline values. Good to excellent improvement was observed in 44% of patients receiving nicotinamide and 55% of patients receiving hydroquinone. |
| 4% hydroquinone | |||||
| [140] | A randomized, double-blind, left–right axilla, placebo-controlled trial | 24 | 4% nicotinamide (n = 16 axillae) | Treatment at night for 9 weeks. | At 9 weeks, L* values in the nicotinamide and desonide groups were increased more compared with the placebo group. Desonide was more effective than nicotinamide (p = 0.002). |
| 0.05% desonide (n = 16 axillae) | |||||
| A placebo cream (n = 16 axillae) |
| Literature | Study Format | No. of Subjects | Compared Formulations | Treatment | Key Findings |
|---|---|---|---|---|---|
| [142] | A randomized, split-face design | 30 | No treatment | Subjects used the ultrasound device for 10 min with or without a gel every night for 4 weeks. | Use of ultrasound treatment with a gel reduced hyperpigmentation compared with no treatment or treatment of a gel alone after 4 weeks. |
| Ultrasound treatment | |||||
| 30 | A gel containing 2% ascorbyl glucoside, and 3.5% nicotinamide | ||||
| Ultrasound treatment with a gel | |||||
| [143] | Double-blind, left–right, randomized, split-face clinical studies | 40 | A vehicle emulsion | Treatment in the morning and evening before bedtime for 8 weeks. | Combination formulation and nicotinamide alone reduced the appearance of hyperpigmentation after 8 weeks. The combination was more effective than nicotinamide alone (p = 0.0003). |
| 5% nicotinamide | |||||
| 40 | 5% nicotinamide | ||||
| 5% nicotinamide plus 1% n-undecylenoyl phenylalanine | |||||
| [144] | A double-blind, vehicle-controlled, full-face, parallel-group clinical study | 101 | 4% nicotinamide plus 2% N-acetyl glucosamine | Treatment of a sunscreen lotion in the morning and test creams in the evening for 8 weeks. | The formulation reduced the area of facial spots and the appearance of irregular pigmentation at weeks 6 (p = 0.0270 and weeks 8 (p = 0.037). |
| 101 | A vehicle cream | ||||
| [145] | A single-center, randomized, double-blind, controlled study | 30 | trans-4-(amino methyl) cyclohexanecarboxylic acid, potassium azeloyl diglycinate, and nicotinamide | Treatment in the morning and before bedtime for 8 weeks. | The formulation reduced the relative melanin value at week 6 (p = 0.006); Reduced MASI scores at week 4 (p = 0.005). |
| 30 | Emulsion-based control | ||||
| [146] | A prospective, randomized, double-blind, vehicle-controlled clinical study | 21 | 2% nicotinamide plus 2% tranexamic acid | Treatment in the morning and evening for 8 weeks. | The formulation reduced melanin index from baseline at weeks 4 (p < 0.001) and 8 (p < 0.001). It reduced the mean pigment intensity score compared with the vehicle control formulation (p = 0.015). |
| 21 | A vehicle cream | ||||
| [147] | A clinical study | 55 | 3% tranexamic acid, 1% kojic acid, 5% nicotinamide, and 5% hydroxyethylpiperazineethane sulfonic acid | Treatment in the morning and evening for 12 weeks. | The formulation reduced melanin index and improved the appearance of hyperpigmentation compared to both pre-treatment baselines. |
| [148] | A prospective, randomized, controlled split-face study | 18 | SKNB19 formulation containing epidermal growth factor, tranexamic acid, vitamin C, arbutin, nicotinamide, and other ingredients | Treatment in the morning and night for 8 weeks. Hydroquinone application only nightly. | SKNB19 improved the appearance of hyperpigmentation when compared with 4% hydroquinone. |
| Standard formulation containing 4% hydroquinone |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boo, Y.C. Mechanistic Basis and Clinical Evidence for the Applications of Nicotinamide (Niacinamide) to Control Skin Aging and Pigmentation. Antioxidants 2021, 10, 1315. https://doi.org/10.3390/antiox10081315
Boo YC. Mechanistic Basis and Clinical Evidence for the Applications of Nicotinamide (Niacinamide) to Control Skin Aging and Pigmentation. Antioxidants. 2021; 10(8):1315. https://doi.org/10.3390/antiox10081315
Chicago/Turabian StyleBoo, Yong Chool. 2021. "Mechanistic Basis and Clinical Evidence for the Applications of Nicotinamide (Niacinamide) to Control Skin Aging and Pigmentation" Antioxidants 10, no. 8: 1315. https://doi.org/10.3390/antiox10081315