
Allysine and α-Aminoadipic Acid as Markers of the Glyco-Oxidative Damage to Human Serum Albumin under Pathological Glucose Concentrations


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals, Reagents, and Materials
2.2. Experimental Design
2.3. Analysis of Samples
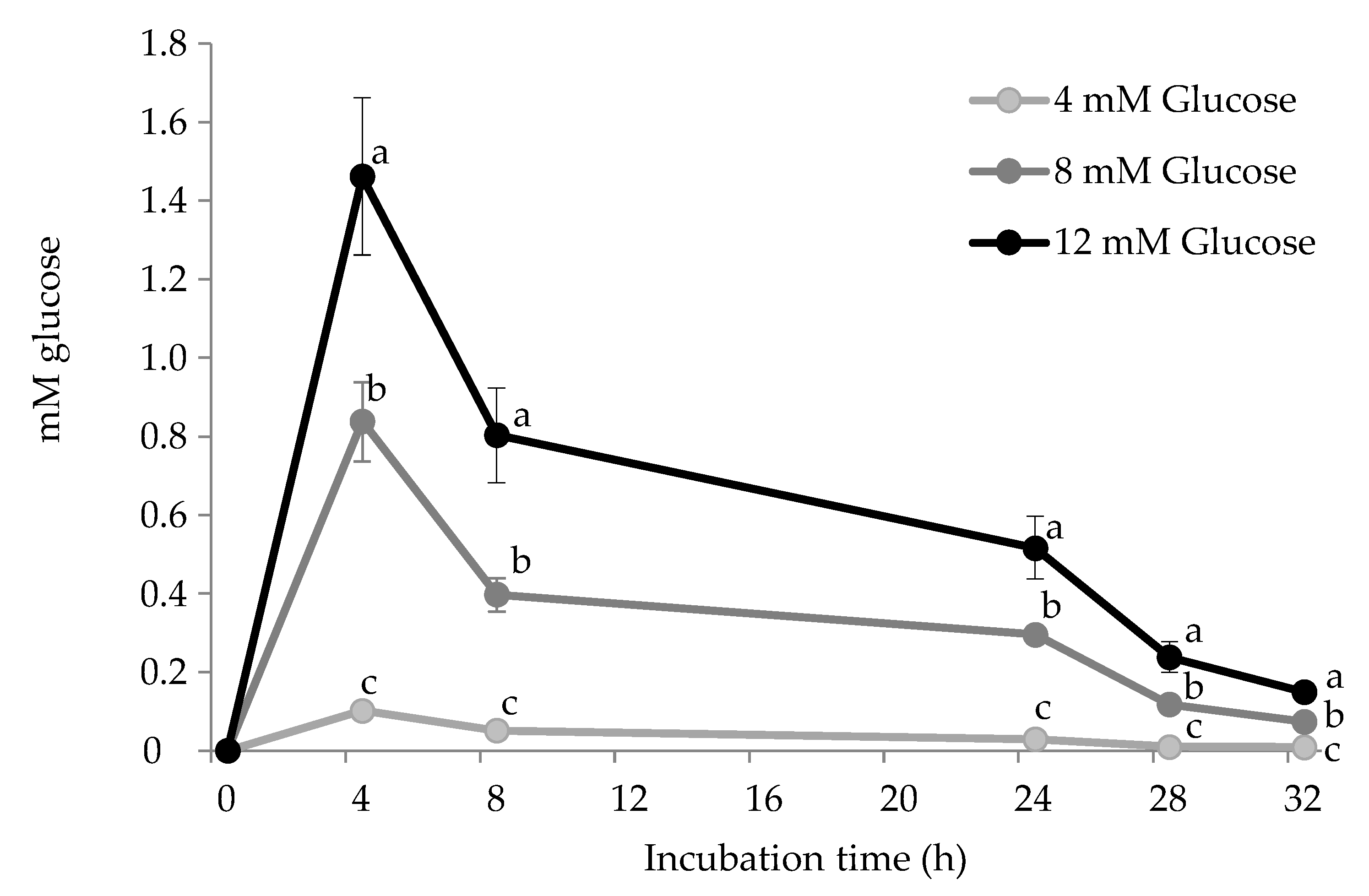
2.3.1. Glucose Analysis by Spectrophotometry
2.3.2. Analysis of Allysine by HPLC
2.3.3. Analysis of α-AA by HPLC
2.3.4. Analysis of Tryptophan by Fluorescence Spectroscopy
2.3.5. Analysis of AGEs by Fluorescence Spectroscopy
2.3.6. Analysis of Yellowness by Spectrophotometry
2.3.7. Statistical Analysis
3. Results and Discussion
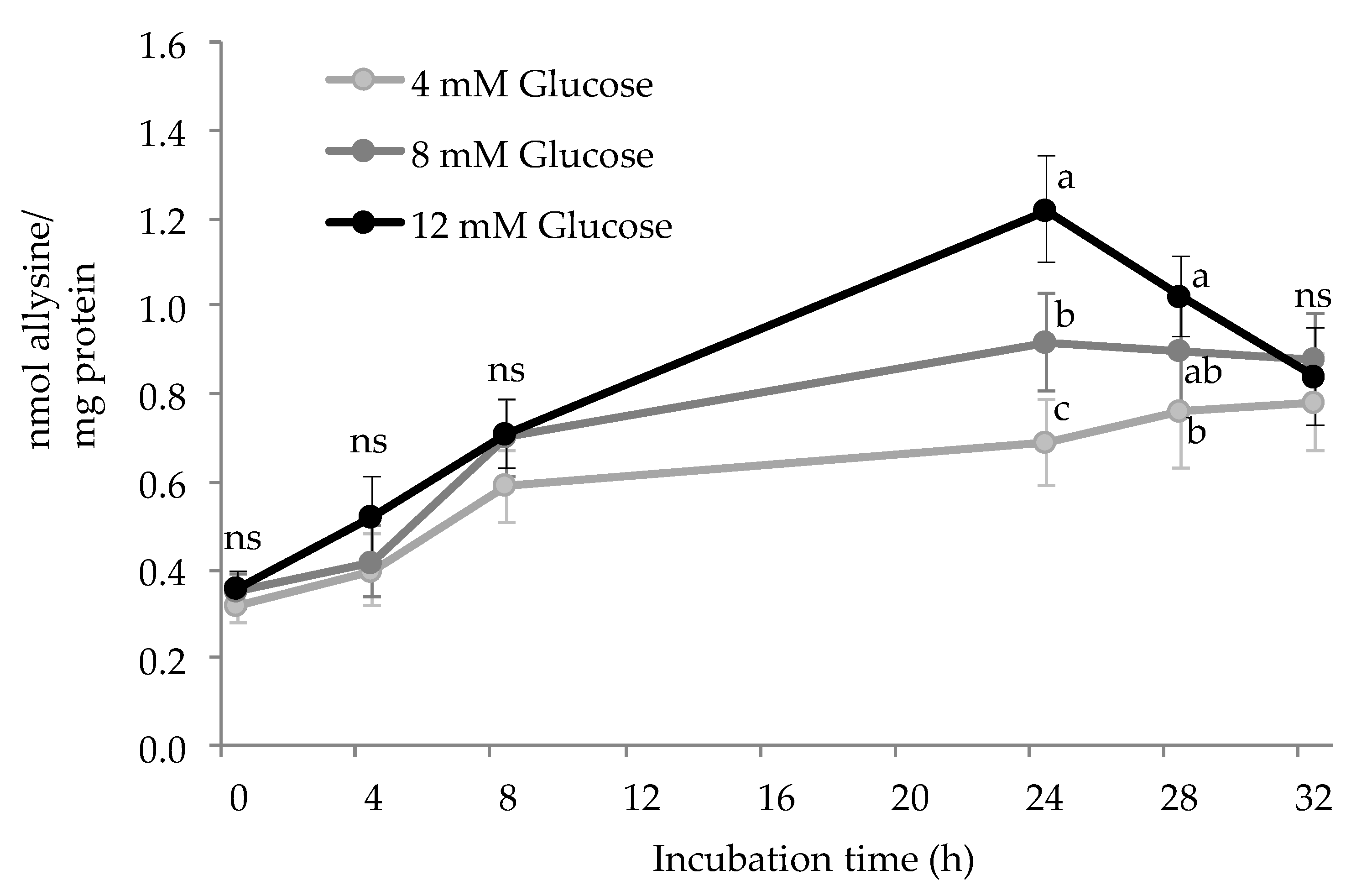
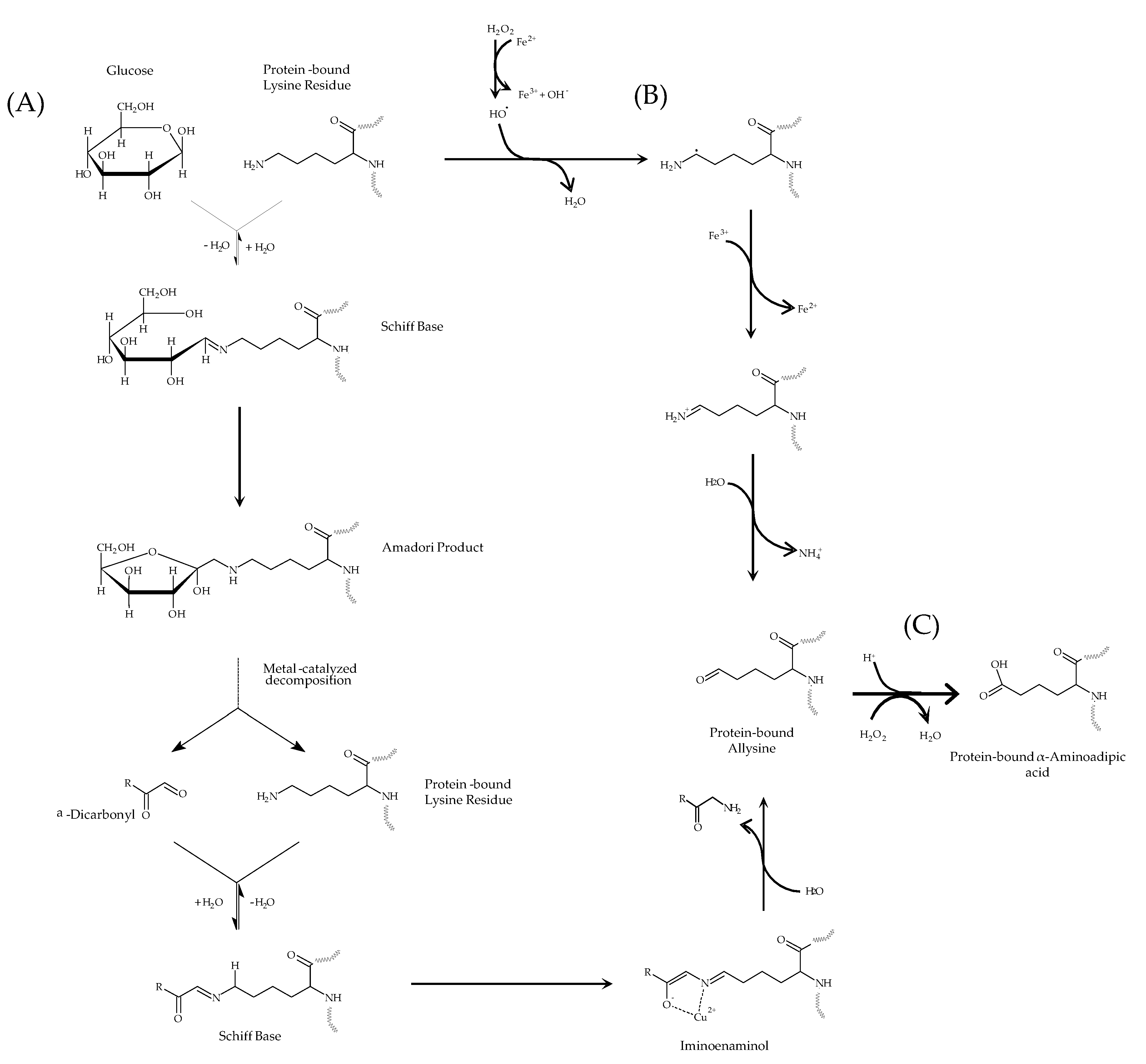
3.1. Carbonylation of HSA by Glucose
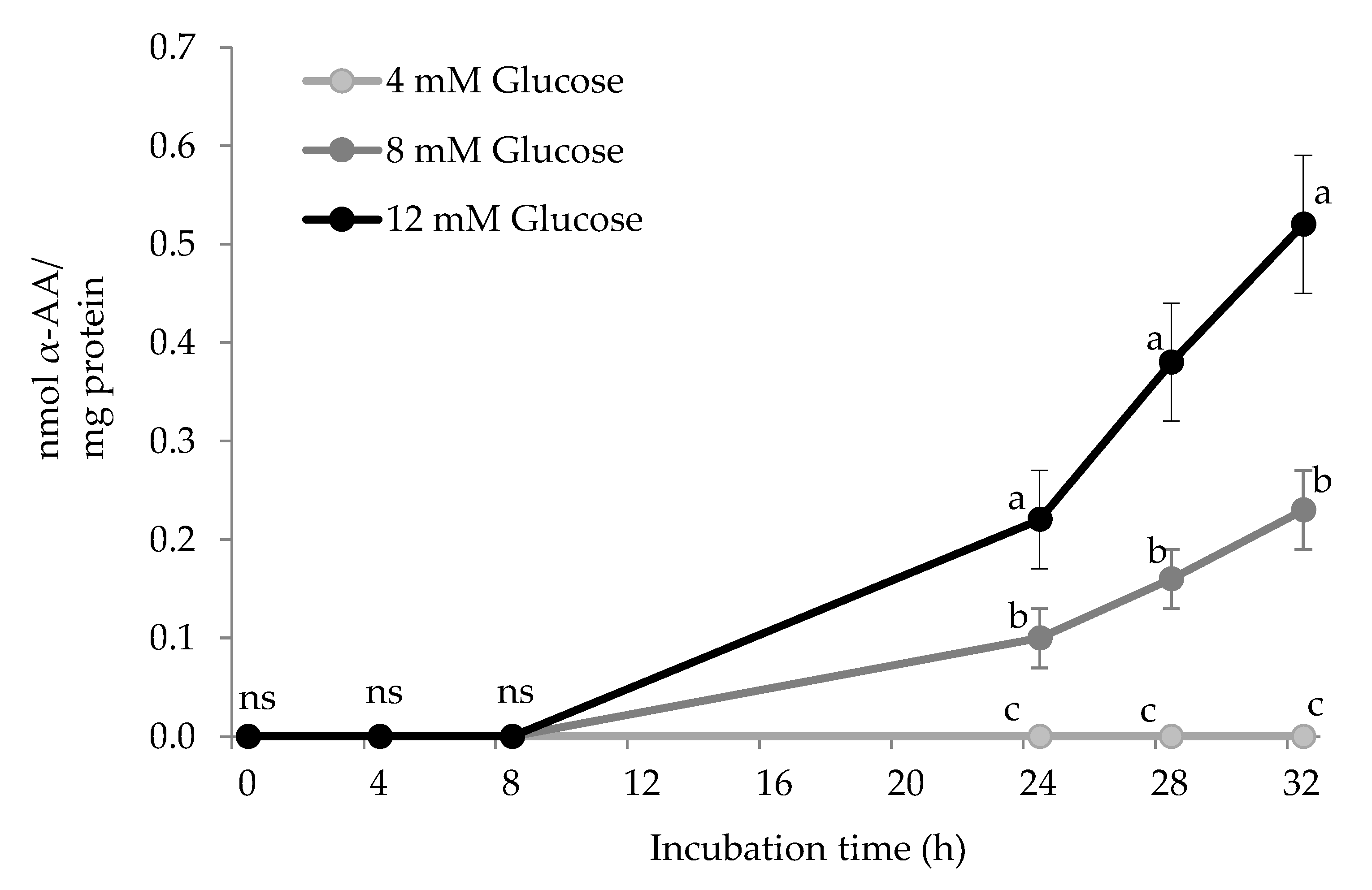
3.2. Formation of α-AA in HSA by Glucose
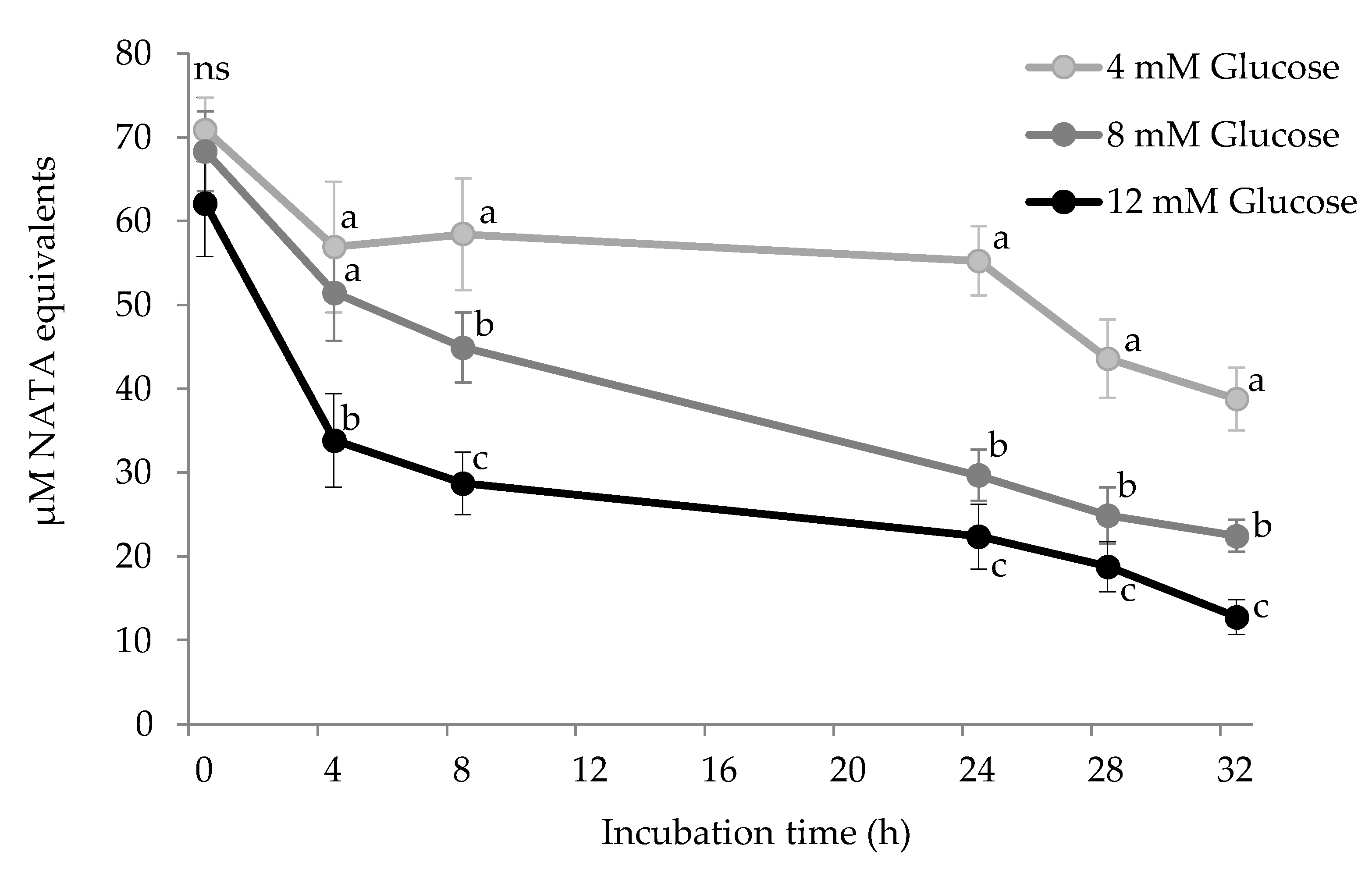
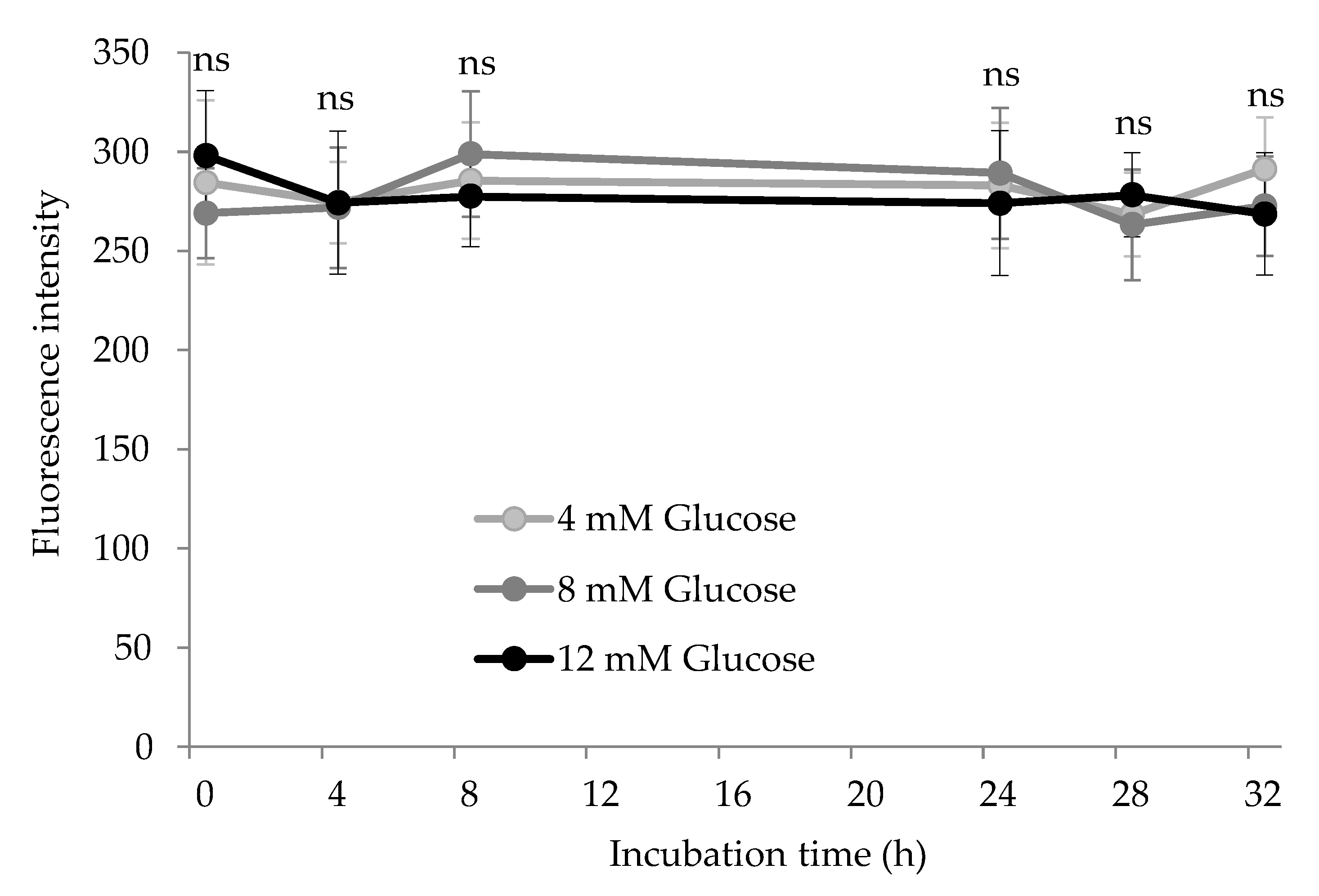
3.3. Tryptophan Depletion by Glucose
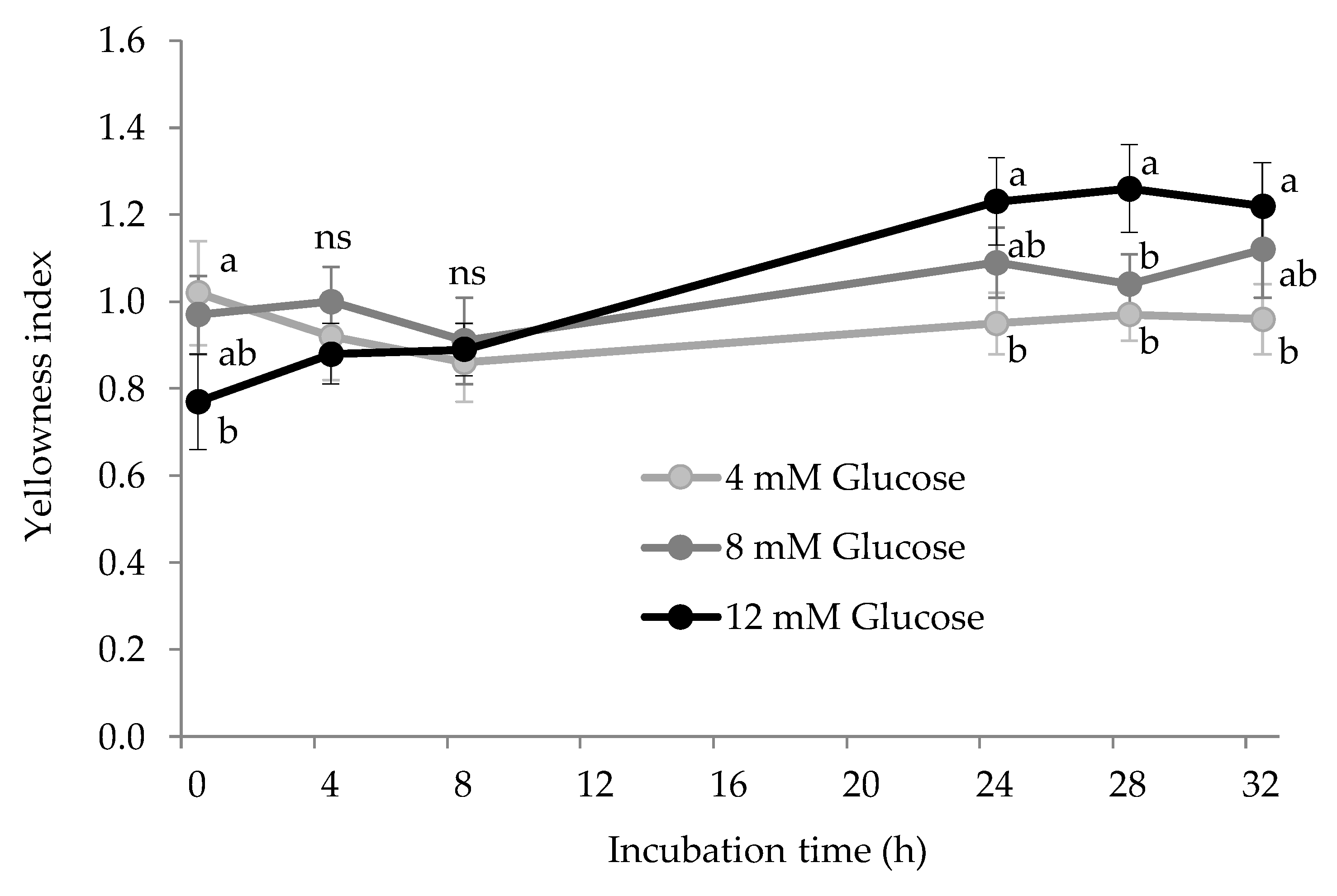
3.4. Other Expressions of Protein Glycoxidation: AGEs and Yellowness
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.J.A. Free radicals and redox regulation in ageing. Free Radic. Biol. Med. 2019, 134, 688–689. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.R.; Chao, C.C.; Levine, R.L.; Stadtman, E.R. Glutamic and aminoadipic semialdehydes are the main carbonyl products of metal-catalyzed oxidation of proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Akagawa, M.; Sasaki, D.; Ishii, Y.; Kurota, Y.; Yotsu-Yamashita, M.; Uchida, K. New methods for the quantitative determination of mayor protein carbonyls, α-aminoadipic and γ-glutamic semialdehydes: Investigation of the formation mechanism and chemical nature in vitro and in vivo. Chem. Res. Toxicol. 2006, 19, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Estévez, M. Formation of allysine in β-lactoglobulin and myofibrillar proteins by glyoxal and methylglyoxal: Impact on water-holding capacity and in vitro digestibility. Food Chem. 2019, 271, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Estévez, M. Oxidative damage to food and human serum proteins: Radical-mediated oxidation vs. glyco-oxidation. Food Chem. 2018, 267, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ruskovska, T.; Bernlohr, D.A. Oxidative stress and protein carbonylation in adipose tissue—Implications for insulin resistance and diabetes mellitus. J. Proteom. 2013, 92, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Jang, H.B.; Kim, W.-H.; Park, K.J.; Kim, K.Y.; Park, S.I.; Lee, H.-J. 2-Aminoadipic acid (2-AAA) as a potential biomarker for insulin resistance in childhood obesity. Sci. Rep. 2019, 9, 13610. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic acid is a biomarker for diabetes risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef]
- Sell, D.R.; Strauch, C.M.; Shen, W.; Monnier, V.M. 2-Aminoadipic acid is a marker of protein carbonyl oxidation in the aging human skin: Effects of diabetes, renal failure and sepsis. Biochem. J. 2008, 404, 269–277. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, J.; Theves, M.; Strauch, C.; Nemet, I.; Liu, X.; Qian, J.; Giblin, F.J.; Monnier, V.M. Mechanism of lysine oxidation in human lens crystallins during aging and in diabetes. J. Biol. Chem. 2009, 284, 34618–34627. [Google Scholar] [CrossRef] [Green Version]
- Estaras, M.; Ameur, F.Z.; Estévez, M.; Díaz-Velasco, S.; Gonzalez, A. The lysine derivative aminoadipic acid, a biomarker of protein oxidation and diabetes-risk, induces production of reactive oxygen species and impairs trypsin secretion in mouse pancreatic acinar cells. Food Chem. Toxicol. 2020, 145, 111594. [Google Scholar] [CrossRef]
- Díaz-Velasco, S.; González, A.; Peña, F.J.; Estévez, M. Noxious effects of selected food-occurring oxidized amino acids on differentiated CACO-2 intestinal human cells. Food Chem. Toxicol. 2020, 144, 111650. [Google Scholar] [CrossRef]
- Utrera, M.; Estévez, M. Oxidation of myofibrillar proteins and impaired functionality: Underlying mechanisms of the carbonylation pathway. J. Agric. Food Chem. 2012, 60, 8002–8011. [Google Scholar] [CrossRef] [PubMed]
- Rysman, T.; Utrera, M.; Morcuende, D.; Royen, G.V.; Weyenberg, S.V.; De Smet, S.; Estévez, M. Apple phenolics as inhibitors of the carbonylation pathway during in vitro metal-catalyzed oxidation of myofibrillar proteins. Food Chem. 2016, 211, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Arcanjo, N.M.O.; Luna, C.; Madruga, M.S.; Estévez, M. Antioxidant and pro-oxidant actions of resveratrol on human serum albumin in the presence of toxic diabetes metabolites: Glyoxal and methyl-glyoxal. Biochim. Biophys. Acta 2018, 1862, 1938–1947. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Rodríguez-Carpena, J.-G.; Estévez, M. Fluorescent HPLC for the detection of specific protein oxidation carbonyls—α-aminoadipic and γ-glutamic semialdehydes—In meat systems. Meat Sci. 2012, 89, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Utrera, M.; Estévez, M. Oxidative damage to poultry, pork, and beef during frozen storage through the analysis of novel protein oxidation markers. J. Agric. Food Chem. 2013, 61, 7987–7993. [Google Scholar] [CrossRef]
- Hunter, R.S. Appearance measurements. Color difference tolerances. Packag. Dev. 1974, 4, 53–60. [Google Scholar]
- Akagawa, M.; Sasaki, D.; Kurota, Y.; Suyama, K. Formation of α-aminoadipic and γ-glutamic semialdehydes in proteins by the Maillard reaction. Ann. N. Y. Acad. Sci. 2005, 1043, 129–134. [Google Scholar] [CrossRef]
- Suyama, K.; Akagawa, M.; Sasaki, T. Oxidative deamination of lysine residue in plasma protein from diabetic rat: α-dicarbonyl-mediated mechanism. Int. Conf. Ser. 2002, 1245, 243–248. [Google Scholar] [CrossRef]
- Akagawa, M.; Suyama, K. Oxidative deamination by hydrogen peroxide in the presence of metals. Free Radic. Res. 2002, 36, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Durand, E.; Elias, R.J.; Tikekar, R.V. Generation of reactive oxidative species from thermal treatment of sugar solutions. Food Chem. 2016, 196, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Terrón, G.; Dueñas, C.; Arjona, A.; Ruiz, J.; Estévez, M. Glucose boosts protein oxidation/nitration during simulated gastric digestion of myofibrillar proteins by creating a severe pro-oxidative environment. Food Chem. 2021, in press. [Google Scholar]
- Özyurt, H.; Luna, C.; Estévez, M. Redox chemistry of the molecular interactions between tea catechins and human serum proteins under simulated hyperglycemic conditions. Food Funct. 2016, 7, 1390–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollineni, R.C.; Fedorova, M.; Blüher, M.; Hoffmann, R. Carbonylated plasma proteins as potential biomarkers of obesity induced type 2 diabetes mellitus. J. Proteome Res. 2014, 13, 5081–5093. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Wagner, A.H. Role of protein carbonylation in diabetes. J. Inherit. Metab. Dis. 2018, 41, 29–38. [Google Scholar] [CrossRef]
- Lin, H.; Levison, B.S.; Buffa, J.A.; Huang, Y.; Fu, X.; Wang, Z.; Gogonea, V.; DiDonato, J.A.; Hazen, S.L. Myeloperoxidase-mediated protein lysine oxidation generates 2-aminoadipic acid and lysine nitrile in vivo. Free Radic. Biol. Med. 2017, 104, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Razquin, C.; Ruiz-Canela, M.; Clish, C.B.; Li, J.; Toledo, E.; Dennis, C.; Liang, L.; Salas-Huetos, A.; Pierce, K.A.; Guasch-Ferré, M.; et al. Lysine pathway metabolites and the risk of type 2 diabetes and cardiovascular disease in the PREDIMED study: Results from two case-cohort studies. Cardiovasc. Diabetol. 2019, 18, 151. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.W.A.; Rasheed, Z.; Khan, W.A.; Ali, R. Biochemical, biophysical, and thermodynamic analysis of in vitro glycated human serum albumin. Biochemistry 2007, 72, 146–152. [Google Scholar]
- Coussons, P.J.; Jacoby, J.; McKay, A.; Kelly, S.M.; Price, N.C.; Hunt, J.V. Glucose modification of human serum albumin: A structural study. Free Radic. Biol. Med. 1997, 22, 1217–1227. [Google Scholar] [CrossRef]
- Cussimanio, B.L.; Booth, A.A.; Todd, P.; Hudson, B.G.; Khalifah, R.G. Unusual susceptibility of heme proteins to damage by glucose during non-enzymatic glycation. Biophys. Chem. 2003, 105, 743–755. [Google Scholar] [CrossRef]
- Yu, E.; Papandreou, C.; Ruiz-Canela, M.; Guasch-Ferre, M.; Clish, C.B.; Dennis, C.; Liang, L.; Corella, D.; Fitó, M.; Razquin, C.; et al. Association of tryptophan metabolites with incident type 2 diabetes in the PREDIMED trial: A case–cohort study. Clin. Chem. 2018, 64, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Rebnord, E.W.; Strand, E.; Midttun, Ø.; Svingen, G.F.T.; Christensen, M.H.E.; Ueland, P.M.; Mellgren, G.; Njølstad, P.R.; Tell, G.S.; Nygård, O.K.; et al. The kynurenine:tryptophan ratio as a predictor of incident type 2 diabetes mellitus in individuals with coronary artery disease. Diabetologia 2017, 60, 1712–1721. [Google Scholar] [CrossRef]
- Matsuoka, K.; Kato, K.; Takao, T.; Ogawa, M.; Ishii, Y.; Shimizu, F.; Masuda, J.; Takada, A. Concentrations of various tryptophan metabolites are higher in patients with diabetes mellitus than in healthy aged male adults. Diabetol. Int. 2017, 8, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.M.S.; Nunes, E.A.; Lago, L.; Pedron, C.N.; Manieri, T.M.; Sato, R.H.; Oliveira, V.X.; Cerchiaro, G. Generation of Advanced Glycation End-Products (AGEs) by glycoxidation mediated by copper and ROS in a human serum albumin (HSA) model peptide: Reaction mechanism and damage in motor neuron cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2017, 824, 42–51. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of glycation inhibitors on aging and age-related diseases. Mech. Ageing Dev. 2016, 160, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ventanas, S.; Estévez, M.; Delgado, C.L.; Ruiz, J. Phospholipid oxidation, non-enzymatic browning development and volatile compounds generation in model systems containing liposomes from porcine Longissimus dorsi and selected amino acids. Eur. Food Res. Technol. 2007, 225, 665–675. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luna, C.; Arjona, A.; Dueñas, C.; Estevez, M. Allysine and α-Aminoadipic Acid as Markers of the Glyco-Oxidative Damage to Human Serum Albumin under Pathological Glucose Concentrations. Antioxidants 2021, 10, 474. https://doi.org/10.3390/antiox10030474
Luna C, Arjona A, Dueñas C, Estevez M. Allysine and α-Aminoadipic Acid as Markers of the Glyco-Oxidative Damage to Human Serum Albumin under Pathological Glucose Concentrations. Antioxidants. 2021; 10(3):474. https://doi.org/10.3390/antiox10030474
Chicago/Turabian StyleLuna, Carolina, Alexis Arjona, Carmen Dueñas, and Mario Estevez. 2021. "Allysine and α-Aminoadipic Acid as Markers of the Glyco-Oxidative Damage to Human Serum Albumin under Pathological Glucose Concentrations" Antioxidants 10, no. 3: 474. https://doi.org/10.3390/antiox10030474