
D2 Receptors and Sodium Ion Channel Blockades of the Basolateral Amygdala Attenuate Lithium Chloride-Induced Conditioned Taste Aversion Applying to Cancer Chemotherapy Nausea and Vomiting

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Microinjections
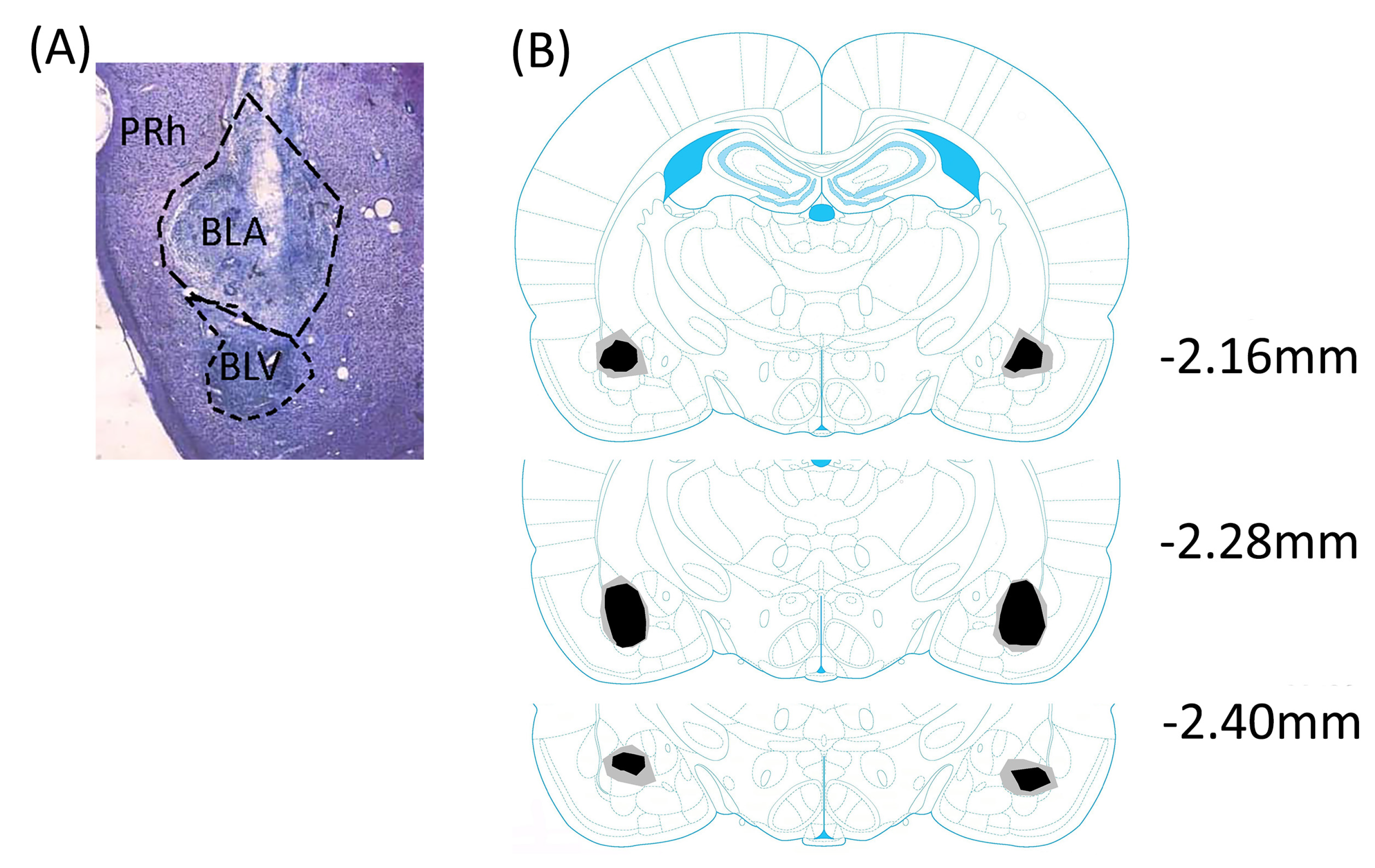
2.3. Histology: Thionin Staining
2.4. Apparatus
2.5. Procedure
2.6. Drugs
2.7. Statistical Analysis
3. Results
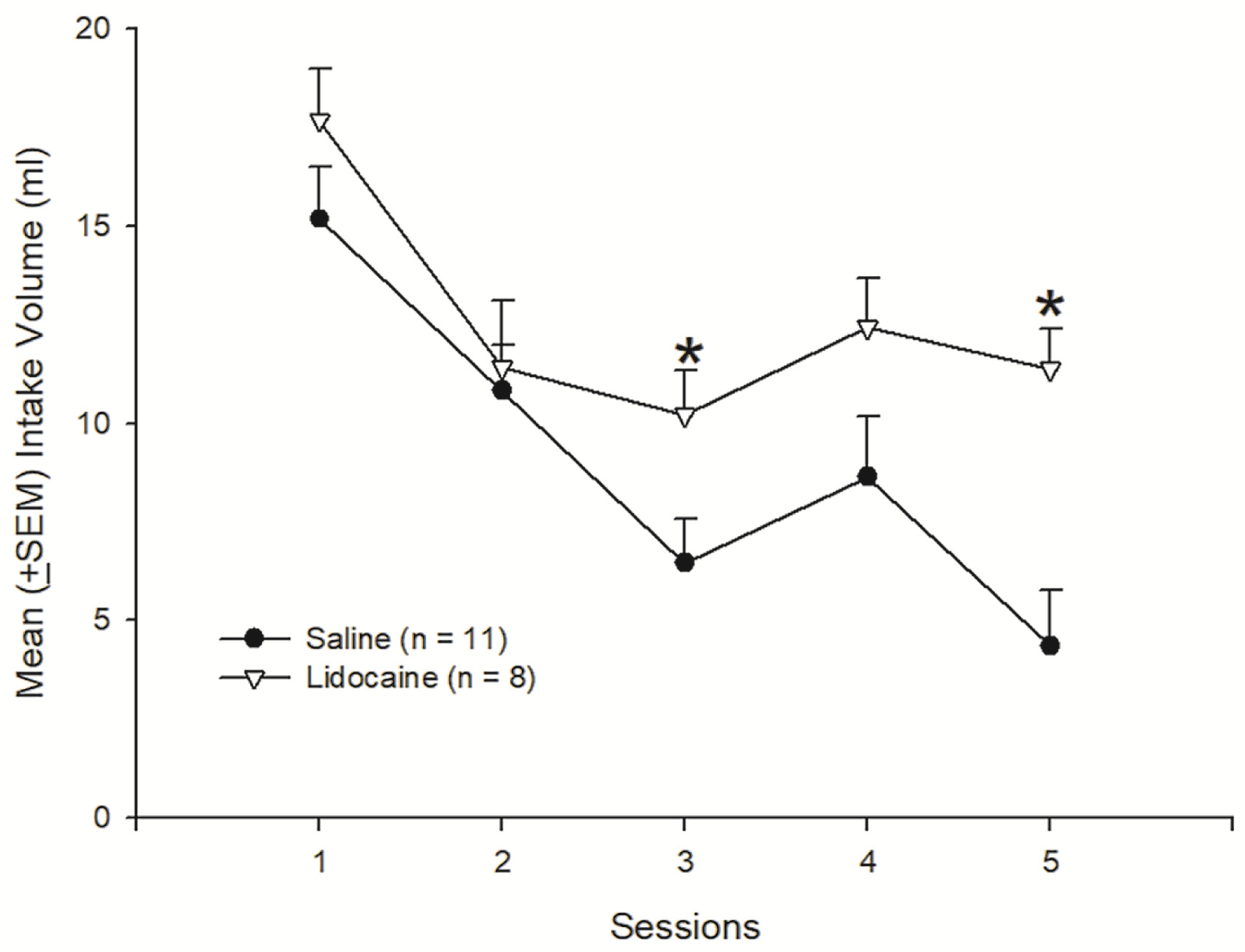
3.1. Experiment 1: Attenuated Effects of Sodium Ion Channel Blocker Lidocaine in the Basolateral Amygdala to LiCl-Induced CTA Learning
3.2. Experiment 2: Blunted Effects of D2 Receptor Antagonist Haloperidol in the Basolateral Amygdala to LiCl-Induced CTA Learning
4. Discussion
4.1. The Basolateral Amygdala and Conditioned Taste Aversion
4.2. Clinical Implications: Behavioral Interventions for Cancer Chemotherapy-Induced Anticipatory Nausea and Vomiting
4.3. Emerged Issues and Further Studies
4.4. Experimental Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stockhorst, U.; Steingrueber, H.-J.; Enck, P.; Klosterhalfen, S. Pavlovian conditioning of nausea and vomiting. Auton. Neurosci. 2006, 129, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, P.B.; Bovbjerg, D.H.; Schwartz, M.D.; Andrykowski, M.A.; Futterman, A.D.; Gilewski, T.; Norton, L.; Redd, W.H. Formation of food aversions in cancer patients receiving repeated infusions of chemotherapy. Behav. Res. Ther. 1993, 31, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Mattes, R.D.; Arnold, C.; Boraas, M. Learned food aversions among cancer chemotherapy patients. Incidence, nature, and clinical implications. Cancer 1987, 60, 2576–2580. [Google Scholar] [CrossRef] [PubMed]
- Scalera, G.; Bavieri, M. Role of conditioned taste aversion on the side effects of chemotherapy in cancer patients. In Conditioned Taste Aversions: Behavioral and Neural Mechanisms; Reilly, S., Schachtman, T.R., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 513–541. [Google Scholar]
- Bovbjerg, D.H.; Redd, W.H.; Jacobsen, P.B.; Manne, S.L.; Taylor, K.L.; Surbone, A.; Crown, J.P.; Norton, L.; Gilewski, T.A.; Hudis, C.A. An experimental analysis of classically conditioned nausea during cancer chemotherapy. Psychosom. Med. 1992, 54, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Greene, P.G.; Seime, R.J. Stimulus control of anticipatory nausea in cancer chemotherapy. J. Behav. Ther. Exp. Psychiatry 1987, 18, 61–64. [Google Scholar] [CrossRef]
- Montgomery, G.H.; Bovbjerg, D.H. The Development of Anticipatory Nausea in Patients Receiving Adjuvant Chemotherapy for Breast Cancer. Physiol. Behav. 1997, 61, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, G.H.; Tomoyasu, N.; Bovbjerg, D.H.; Andrykowski, M.A.; Currie, V.E.; Jacobsen, P.B.; Redd, W.H. Patients’ pretreatment expectations of chemotherapy-related nausea are an independent predictor of anticipatory nausea. Ann. Behav. Med. 1998, 20, 104–108. [Google Scholar] [CrossRef]
- Bernstein, I.L. Learned Food Aversions in the Progression of Cancer and Its Treatment. Ann. N. Y. Acad. Sci. 1985, 443, 365–380. [Google Scholar] [CrossRef]
- Jacobsen, P.B.; Bovbjerg, D.H.; Schwartz, M.D.; Hudis, C.A.; Gilewski, T.A.; Norton, L. Conditioned emotional distress in women receiving chemotherapy for breast cancer. J. Consult. Clin. Psychol. 1995, 63, 108–114. [Google Scholar] [CrossRef]
- Alejandro Borja, G.-P.; Alejandro Navarro, E.; Beatriz, G.-C.; Ignacio, M.; Milagros, G. Accumbens and amygdala in taste recognition memory: The role of d1 dopamine receptors. Neurobiol. Learn. Mem. 2020, 174, 107277. [Google Scholar] [CrossRef]
- St Andre, J.; Reilly, S. Effects of central and basolateral amygdala lesions on conditioned taste aversion and latent inhibition. Behav. Neurosci. 2007, 121, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Fujimoto, Y.; Shimura, T.; Sakai, N. Conditioned taste aversion in rats with excitotoxic brain lesions. Neurosci. Res. 1995, 22, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Shimura, T.; Sako, N.; Yasoshima, Y.; Sakai, N. Neural substrates for conditioned taste aversion in the rat. Behav. Brain Res. 1994, 65, 123–137. [Google Scholar] [CrossRef]
- Inui, T.; Inui-Yamamoto, C.; Yoshioka, Y.; Ohzawa, I.; Shimura, T. Activation of efferents from the basolateral amygdala during the retrieval of conditioned taste aversion. Neurobiol. Learn. Mem. 2013, 106, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Shionoya, K.; Datiche, F. Inactivation of the basolateral amygdala impairs the retrieval of recent and remote taste-potentiated odor aversion memory. Neurobiol. Learn. Mem. 2009, 92, 590–596. [Google Scholar] [CrossRef]
- Inui, T.; Sugishita, T.; Inui-Yamamoto, C.; Yasoshima, Y.; Shimura, T. The Basolateral Nucleus of the Amygdala Executes the Parallel Processes of Avoidance and Palatability in the Retrieval of Conditioned Taste Aversion in Male Rats. eNeuro 2019, 6, ENEURO.0004-19.2019. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2007; ISBN 9780123919496. [Google Scholar]
- Molero-Chamizo, A.; Rivera-Urbina, G.N. Effects of lesions in different nuclei of the amygdala on conditioned taste aversion. Exp. Brain Res. 2017, 235, 3517–3526. [Google Scholar] [CrossRef]
- Reilly, S.; Bornovalova, M.A. Conditioned taste aversion and amygdala lesions in the rat: A critical review. Neurosci. Biobehav. Rev. 2005, 29, 1067–1088. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.C.; Hsiao, S. Haloperidol attenuates rewarding and aversively conditioned suppression of saccharin solution intake: Reevaluation of the anhedonia hypothesis of dopamine blocking. Behav. Neurosci. 2002, 116, 646–650. [Google Scholar] [CrossRef]
- Stevenson, C.W.; Gratton, A. Role of basolateral amygdala dopamine in modulating prepulse inhibition and latent inhibition in the rat. Psychopharmacology 2004, 176, 139–145. [Google Scholar] [CrossRef]
- Keshavarzian, E.; Ghasemzadeh, Z.; Rezayof, A. The basolateral amygdala dopaminergic system contributes to the improving effect of nicotine on stress-induced memory impairment in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 86, 30–35. [Google Scholar] [CrossRef] [PubMed]
- LaLumiere, R.T.; Nawar, E.M.; McGaugh, J.L. Modulation of memory consolidation by the basolateral amygdala or nucleus accumbens shell requires concurrent dopamine receptor activation in both brain regions. Learn. Mem. 2005, 12, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Lalumiere, R.T.; Nguyen, L.T.; McGaugh, J.L. Post-training intrabasolateral amygdala infusions of dopamine modulate consolidation of inhibitory avoidance memory: Involvement of noradrenergic and cholinergic systems. Eur. J. Neurosci. 2004, 20, 2804–2810. [Google Scholar] [CrossRef] [PubMed]
- Macedo, C.E.; Martinez, R.C.; Albrechet-Souza, L.; Molina, V.A.; Brandão, M.L. 5-HT2- and D1-mechanisms of the basolateral nucleus of the amygdala enhance conditioned fear and impair unconditioned fear. Behav. Brain Res. 2007, 177, 100–108. [Google Scholar] [CrossRef]
- Rosen, L.G.; Rushlow, W.J.; Laviolette, S.R. Opiate exposure state controls dopamine D3 receptor and cdk5/calcineurin signaling in the basolateral amygdala during reward and withdrawal aversion memory formation. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 59–66. [Google Scholar] [CrossRef]
- Aapro, M.S.; Molassiotis, A.; Olver, I. Anticipatory nausea and vomiting. Support. Care Cancer 2005, 13, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Burish, T.G.; Carey, M.P.; Krozely, M.G.; Greco, F.A. Conditioned side effects induced by cancer chemotherapy: Prevention through behavioral treatment. J. Consult Clin. Psychol. 1987, 55, 42–48. [Google Scholar] [CrossRef]
- Kamen, C.; Tejani, M.A.; Chandwani, K.; Janelsins, M.; Peoples, A.R.; Roscoe, J.A.; Morrow, G.R. Anticipatory nausea and vomiting due to chemotherapy. Eur. J. Pharmacol. 2014, 722, 172–179. [Google Scholar] [CrossRef]
- Stockhorst, U.; Klosterhalfen, S.; Klosterhalfen, W.; Winkelmann, M.; Steingrueber, H.-J. Anticipatory nausea in cancer patients receiving chemotherapy: Classical conditioning etiology and therapeutical implications. Integr. Psychol. Behav. Sci. 1993, 28, 177–181. [Google Scholar] [CrossRef]
- Stockhorst, U.; Wiener, J.A.; Klosterhalfen, S.; Klosterhalfen, W.; Aul, C.; Steingrüber, H.-J. Effects of overshadowing on conditioned nausea in cancer patients: An experimental study. Physiol. Behav. 1998, 64, 743–753. [Google Scholar] [CrossRef]
- Redd, W.H.; Montgomery, G.H.; Duhamel, K.N. Behavioral Intervention for Cancer Treatment Side Effects. J. Natl. Cancer Inst. 2001, 93, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Andresen, G.V.; Birch, L.L.; Johnson, P.A. The scapegoat effect on food aversions after chemotherapy. Cancer 1990, 66, 1649–1653. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, I.L. Aversion conditioning in response to cancer and cancer treatment. Clin. Psychol. Rev. 1991, 11, 185–191. [Google Scholar] [CrossRef]
- He, A.B.H.; Huang, C.L.; Kozłowska, A.; Chen, J.C.; Wu, C.-W.; Huang, A.C.W.; Liu, Y.Q. Involvement of neural substrates in reward and aversion to methamphetamine addiction: Testing the reward comparison hypothesis and the paradoxical effect hypothesis of abused drugs. Neurobiol. Learn. Mem. 2019, 166, 107090. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.C.W.; Yu, Y.H.; He, A.B.H.; Ou, C.Y. Interactions between prelimbic cortex and basolateral amygdala contribute to morphine-induced conditioned taste aversion in conditioning and extinction. Neurobiol. Learn. Mem. 2020, 172, 107248. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.Y.; Yu, Y.H.; Wu, C.-W.; Kozłowska, A.; Shyu, B.-C.; Huang, A.C.W. Neuronal activity of the medial prefrontal cortex, nucleus accumbens, and basolateral amygdala in conditioned taste aversion and conditioned place preference induced by different doses of morphine administrations in rats. Front. Pharmacol. 2023, 14, 1062169. [Google Scholar] [CrossRef]
- Yu, Y.; He, A.B.; Liou, M.; Ou, C.; Kozłowska, A.; Chen, P.; Huang, A.C. The Paradoxical Effect Hypothesis of Abused Drugs in a Rat Model of Chronic Morphine Administration. J. Clin. Med. 2021, 10, 3197. [Google Scholar] [CrossRef]
- Grigson, P.S.; Lyuboslavsky, P.N.; Tanase, D.; Wheeler, R.A. Water-Deprivation Prevents Morphine-, but not LiCl-Induced, Suppression of Sucrose Intake. Physiol. Behav. 1999, 67, 277–286. [Google Scholar] [CrossRef]
- Huang, A.C.; Hsiao, S. Re-examination of amphetamine-induced conditioned suppression of tastant intake in rats: The task-dependent drug effects hypothesis. Behav. Neurosci. 2008, 122, 1207–1216. [Google Scholar] [CrossRef]
- Roma, P.G.; Flint, W.W.; Higley, J.D.; Riley, A.L. Assessment of the aversive and rewarding effects of alcohol in Fischer and Lewis rats. Psychopharmacology 2006, 189, 187–199. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Z.-Y.; Huang, C.M.; Cheng, C.-N.; Huang, A.C.-W. D2 Receptors and Sodium Ion Channel Blockades of the Basolateral Amygdala Attenuate Lithium Chloride-Induced Conditioned Taste Aversion Applying to Cancer Chemotherapy Nausea and Vomiting. Brain Sci. 2023, 13, 697. https://doi.org/10.3390/brainsci13040697
Gao Z-Y, Huang CM, Cheng C-N, Huang AC-W. D2 Receptors and Sodium Ion Channel Blockades of the Basolateral Amygdala Attenuate Lithium Chloride-Induced Conditioned Taste Aversion Applying to Cancer Chemotherapy Nausea and Vomiting. Brain Sciences. 2023; 13(4):697. https://doi.org/10.3390/brainsci13040697
Chicago/Turabian StyleGao, Zhi-Yue, Chung Ming Huang, Cai-N Cheng, and Andrew Chih-Wei Huang. 2023. "D2 Receptors and Sodium Ion Channel Blockades of the Basolateral Amygdala Attenuate Lithium Chloride-Induced Conditioned Taste Aversion Applying to Cancer Chemotherapy Nausea and Vomiting" Brain Sciences 13, no. 4: 697. https://doi.org/10.3390/brainsci13040697