
Prolonged Intrinsic Neural Timescales Dissociate from Phase Coherence in Schizophrenia



Abstract
:1. Introduction
Aims
2. Materials and Methods
2.1. Participants
2.2. EEG Recordings
2.3. Preprocessing
2.4. Analysis
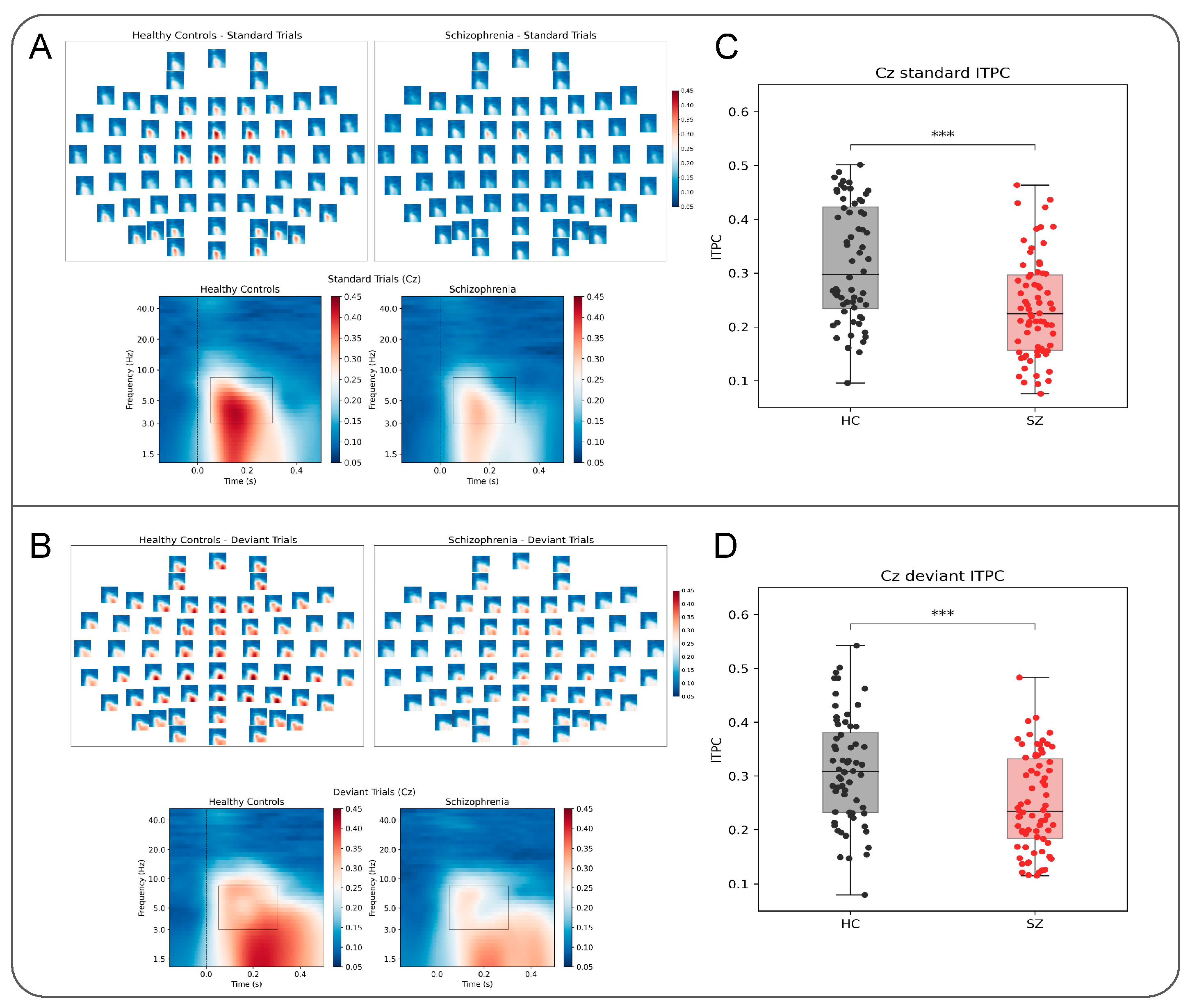
2.4.1. Intertrial Phaser Coherence
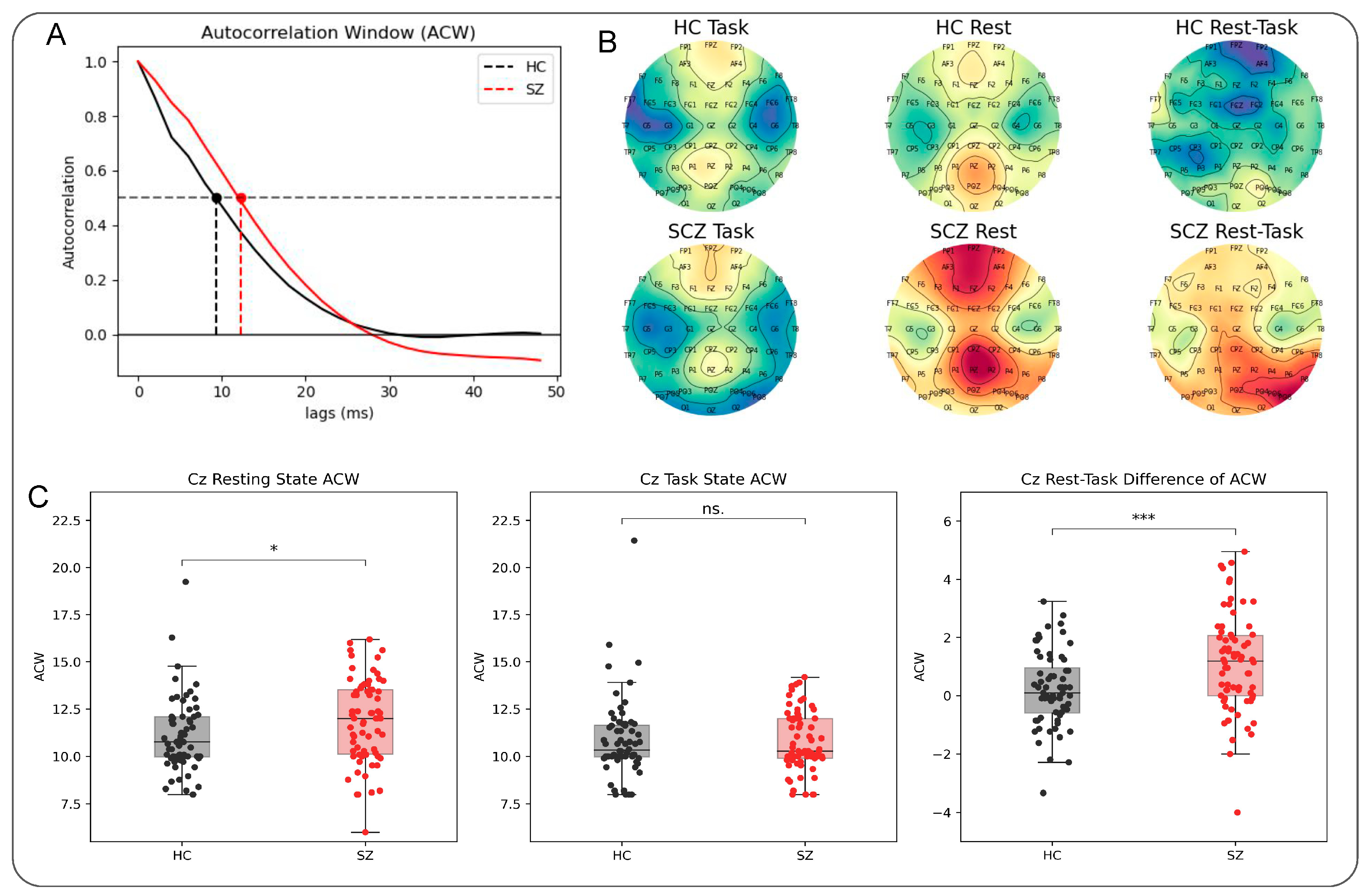
2.4.2. Autocorrelation Window
2.4.3. Statistical Analysis
3. Results
3.1. Decreased ITPC in Schizophrenia
3.2. Longer ACW during the Resting State in Schizophrenia
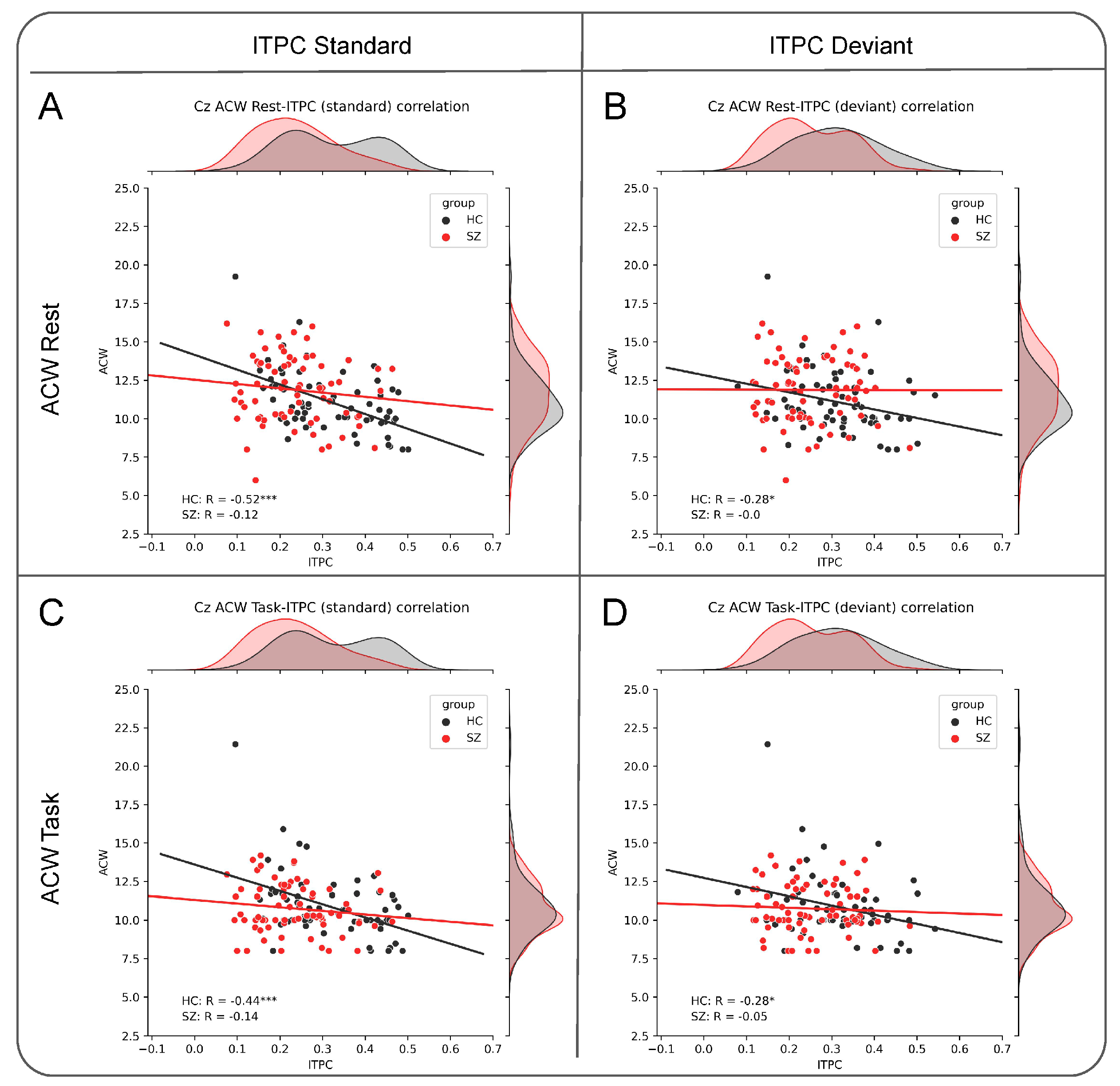
3.3. Negative Relationship between ACW and ITPC
4. Discussion
Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Javitt, D.C.; Freedman, R. Sensory Processing Dysfunction in the Personal Experience and Neuronal Machinery of Schizophrenia. Am. J. Psychiatry 2015, 172, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Javitt, D.C.; Sweet, R.A. Auditory Dysfunction in Schizophrenia: Integrating Clinical and Basic Features. Nat. Rev. Neurosci. 2015, 16, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Karanikolaou, M.; Limanowski, J.; Northoff, G. Does Temporal Irregularity Drive Prediction Failure in Schizophrenia? Temporal Modelling of ERPs. Schizophrenia 2022, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.; Gomez-pilar, J.; Zhang, J.; Choueiry, J.; Salle, S.; De Knott, V.; Northoff, G. It’s in the Timing: Reduced Temporal Precision in Neural Activity of Schizophrenia. Cereb. Cortex 2022, 32, 1–16. [Google Scholar] [CrossRef]
- Shin, K.S.; Kim, J.S.; Kim, S.N.; Hong, K.S.; O’Donnell, B.F.; Chung, C.K.; Kwon, J.S. Intraindividual Neurophysiological Variability in Ultra-Highrisk for Psychosis and Schizophrenia Patients: Single-Trial Analysis. NPJ Schizophr. 2015, 1, 1–7. [Google Scholar] [CrossRef]
- Ko, D.; Kwon, S.; Lee, G.T.; Im, C.H.; Kim, K.H.; Jung, K.Y. Theta Oscillation Related to the Auditory Discrimination Process in Mismatch Negativity: Oddball versus Control Paradigm. J. Clin. Neurol. 2012, 8, 35–42. [Google Scholar] [CrossRef]
- Koh, Y.; Shin, K.S.; Kim, J.S.; Choi, J.S.; Kang, D.H.; Jang, J.H.; Cho, K.H.; O’Donnell, B.F.; Chung, C.K.; Kwon, J.S. An MEG Study of Alpha Modulation in Patients with Schizophrenia and in Subjects at High Risk of Developing Psychosis. Schizophr. Res. 2011, 126, 36–42. [Google Scholar] [CrossRef]
- Lakatos, P.; Schroeder, C.E.; Leitman, D.I.; Javitt, D.C. Predictive Suppression of Cortical Excitability and Its Deficit in Schizophrenia. J. Neurosci. 2013, 33, 11692–11702. [Google Scholar] [CrossRef]
- Ramsay, I.S.; Pokorny, V.J.; Lynn, P.A.; Klein, S.D.; Sponheim, S.R. Limited Consistency and Strength of Neural Oscillations during Sustained Visual Attention in Schizophrenia. Biol. Psychiatry Cogn. Neurosci. Neuroimaging, 2023; in Press. [Google Scholar] [CrossRef]
- Parker, D.A.; Hamm, J.P.; Mcdowell, J.E.; Keedy, S.K.; Gershon Elliot, S.; Ivleva, E.I.; Pearlson, G.D.; Keshavan, M.S.; Tamminga, C.A.; Sweeney, J.A.; et al. Auditory Steady-State EEG Response Across the Schizo-Bipolar Spectrum. Schizophr. Res. 2019, 209, 218–226. [Google Scholar] [CrossRef]
- Jefsen, O.H.; Shtyrov, Y.; Larsen, K.M.; Dietz, M.J. The 40-Hz Auditory Steady-State Response in Bipolar Disorder: A Meta-Analysis. Clin. Neurophysiol. 2022, 141, 53–61. [Google Scholar] [CrossRef]
- Makeig, S.; Debener, S.; Onton, J.; Delorme, A. Mining Event-Related Brain Dynamics. Trends Cogn. Sci. 2004, 8, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, P.; Gross, J.; Thut, G. A New Unifying Account of the Roles of Neuronal Entrainment. Curr. Biol. 2019, 29, R890–R905. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.; Berberian, N.; Zilio, F.; Goleoskrhi, M.; Gomez-Pillar, J.; Northoff, G. Intrinsic Neural Timescales: Temporla Integration and Segregation. Trends Cogn. Sci. 2022, 2247, 159–173. [Google Scholar] [CrossRef]
- Golesorkhi, M.; Gomez-Pilar, J.; Zilio, F.; Berberian, N.; Wolff, A.; Yagoub, M.C.E.; Northoff, G. The Brain and Its Time: Intrinsic Neural Timescales Are Key for Input Processing. Commun. Biol. 2021, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hasson, U.; Chen, J.; Honey, C.J. Hierarchical Process Memory: Memory as an Integral Component of Information Processing. Trends Cogn. Sci. 2015, 19, 304–313. [Google Scholar] [CrossRef]
- Chen, J.; Leong, Y.C.; Honey, C.J.; Yong, C.H.; Norman, K.A.; Hasson, U. Shared Memories Reveal Shared Structure in Neural Activity across Individuals. Nat. Neurosci. 2016, 20, 115–125. [Google Scholar] [CrossRef]
- Golesorkhi, M.; Gomez-Pilar, J.; Tumati, S.; Fraser, M.; Northoff, G. Temporal Hierarchy of Intrinsic Neural Timescales Converges with Spatial Core-Periphery Organization. Commun. Biol. 2021, 4, 277. [Google Scholar] [CrossRef]
- Himberger, K.D.; Chien, H.Y.; Honey, C.J. Principles of Temporal Processing Across the Cortical Hierarchy. Neuroscience 2018, 389, 161–174. [Google Scholar] [CrossRef]
- Honey, C.J.; Thesen, T.; Donner, T.H.; Silbert, L.J.; Carlson, C.E.; Devinsky, O.; Doyle, W.K.; Rubin, N.; Heeger, D.J.; Hasson, U. Slow Cortical Dynamics and the Accumulation of Information over Long Timescales. Neuron 2012, 76, 423–434. [Google Scholar] [CrossRef]
- Murray, J.D.; Bernacchia, A.; Freedman, D.J.; Romo, R.; Wallis, J.D.; Cai, X.; Padoa-Schioppa, C.; Pasternak, T.; Seo, H.; Lee, D.; et al. A Hierarchy of Intrinsic Timescales across Primate Cortex. Nat. Neurosci. 2014, 17, 1661–1663. [Google Scholar] [CrossRef]
- Uscătescu, L.C.; Said, S.; Kronbichler, L.; Stelzig, R.; Pearce, B.; Reich, L.A.; Weber, S.; Kronbichler, M. Reduced Intrinsic Neural Timescales in Schizophrenia along Posterior Parietal and Occipital Areas. NPJ Schizophr. 2021, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Wengler, K.; Goldberg, A.T.; Chahine, G.; Horga, G. Distinct Hierarchical Alterations of Intrinsic Neural Timescales Account for Different Manifestations of Psychosis. Elife 2020, 9, e56151. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Sandsten, K.E.; Nordgaard, J.; Kjaer, T.W.; Parnas, J. The Self and Its Prolonged Intrinsic Neural Timescale in Schizophrenia. Schizophr. Bull. 2021, 47, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Hartwich, P.; Wolff, A.; Golesorkhi, M.; Northoff, G. The Self in Art Therapy–Brain-Based Assessment of the Drawing Process. Med. Hypotheses 2020, 138, 109596. [Google Scholar] [CrossRef] [PubMed]
- Zilio, F.; Gomez-Pilar, J.; Cao, S.; Zhang, J.; Zang, D.; Qi, Z.; Tan, J.; Hiromi, T.; Wu, X.; Fogel, S.; et al. Are Intrinsic Neural Timescales Related to Sensory Processing? Evidence from Abnormal Behavioral States. Neuroimage 2021, 226, 117579. [Google Scholar] [CrossRef] [PubMed]
- Tamminga, C.A.; Ivleva, E.I.; Keshavan, M.S.; Pearlson, G.D.; Clementz, B.A.; Witte, B.; Morris, D.W.; Bishop, J.; Thaker, G.K.; Sweeney, J.A. Clinical Phenotypes of Psychosis in the Bipolar-Schizophrenia Network on Intermediate Phenotypes (B-SNIP). Am. J. Psychiatry 2013, 170, 1263–1274. [Google Scholar] [CrossRef]
- Gabard-Durnam, L.J.; Leal, A.S.M.; Wilkinson, C.L.; Levin, A.R. The Harvard Automated Processing Pipeline for Electroencephalography (HAPPE): Standardized Processing Software for Developmental and High-Artifact Data. Front. Neurosci. 2018, 12, 97. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Nolan, H.; Whelan, R.; Reilly, R.B. FASTER: Fully Automated Statistical Thresholding for EEG Artifact Rejection. J. Neurosci. Methods 2010, 192, 152–162. [Google Scholar] [CrossRef]
- Gramfort, A.; Luessi, M.; Larson, E.; Engemann, D.; Strohmeier, D.; Brodbeck, C.; Parkkonen, L.; Hämäläinen, M. MNE Software for Processing MEG and EEG Data. Neuroimage 2014, 86, 446–460. [Google Scholar] [CrossRef]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and Statistical Modeling with Python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; pp. 92–96. [Google Scholar] [CrossRef]
- Ding, N.; Simon, J.Z. Cortical Entrainment to Continuous Speech: Functional Roles and Interpretations. Front. Hum. Neurosci. 2014, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Calderone, D.J.; Lakatos, P.; Butler, P.D.; Castellanos, F.X. Entrainment of Neural Oscillations as a Modifiable Substrate of Attention. Trends Cogn. Sci. 2014, 18, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.K.R.; Debener, S.; Nobre, A.C. Synchronisation of Neural Oscillations and Cross-Modal Influences. Trends Cogn. Sci. 2020, 24, 481–495. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, B.F.; Vohs, J.L.; Krishnan, G.P.; Rass, O.; Hetrick, W.P.; Morzorati, S.L. The Auditory Steady-State Response (ASSR): A Translational Biomarker for Schizophrenia. Suppl. Clin. Neurophysiol. 2013, 62, 101–112. [Google Scholar] [CrossRef]
- Watanabe, T.; Rees, G.; Masuda, N. Atypical Intrinsic Neural Timescale in Autism. Elife 2019, 8, e42256. [Google Scholar] [CrossRef]
- Northoff, G.; Gomez-Pilar, J. Overcoming Rest-Task Divide-Abnormal Temporospatial Dynamics and Its Cognition in Schizophrenia. Schizophr. Bull. 2021, 47, 751–765. [Google Scholar] [CrossRef]
- Stanghellini, G.; Ballerini, M.; Presenza, S.; Mancini, M.; Raballo, A.; Blasi, S.; Cutting, J. Psychopathology of Lived Time: Abnormal Time Experience in Persons With Schizophrenia. Schizophr. Bull. 2016, 42, 45–55. [Google Scholar] [CrossRef]
- Arantes-Gonçalves, F.; Wolman, A.; Bastos-Leite, A.J.; Northoff, G. Scale for Space and Time Experience in Psychosis: Converging Phenomenological and Psychopathological Perspectives. Psychopathology 2021, 55, 132–142. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HC | SCZ | ||
|---|---|---|---|
| N | 64 | 70 | |
| Sex (F/M) | 37/27 | 22/48 | χ2 = 9.44, p = 0.002 |
| Age | 37.62 ± 1.41 years | 36.56 ± 1.31 years | t133 = 0.59, p = 0.552 |
| Sub-diagnosis | |||
| Paranoid | 39 | ||
| Undifferentiated | 14 | ||
| Residual | 4 | ||
| No sub-diagnosis | 13 | ||
| ACW REST | 11.02 ± 1.78 ms | 11.87 ± 2.27 ms | t133 = −2.16, p = 0.016 |
| ACW TASK | 1.78 ± 1.66 ms | 1.74 ± 1.57 ms | t133 = 0.365, p = 0.642 |
| ACW REST–TASK DIFFERENCE | 0.24 ± 1.27 ms | 1.13 ± 1.78 ms | t133 = −3.38, p = 0.001 |
| ITPC STANDARD | 0.23 ± 1.27 | 1.13 ± 1.78 | t133 = 4.8, p = 0.0 |
| ITPC DEVIANT | 0.32 ± 0.1 | 0.25 ± 0.08 | t133 = 3.8, p = 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechner, S.; Northoff, G. Prolonged Intrinsic Neural Timescales Dissociate from Phase Coherence in Schizophrenia. Brain Sci. 2023, 13, 695. https://doi.org/10.3390/brainsci13040695
Lechner S, Northoff G. Prolonged Intrinsic Neural Timescales Dissociate from Phase Coherence in Schizophrenia. Brain Sciences. 2023; 13(4):695. https://doi.org/10.3390/brainsci13040695
Chicago/Turabian StyleLechner, Stephan, and Georg Northoff. 2023. "Prolonged Intrinsic Neural Timescales Dissociate from Phase Coherence in Schizophrenia" Brain Sciences 13, no. 4: 695. https://doi.org/10.3390/brainsci13040695