
Investigating the Effects of a Focal Muscle Vibration Protocol on Sensorimotor Integration in Healthy Subjects
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Electromyography (EMG)
2.3. STDT
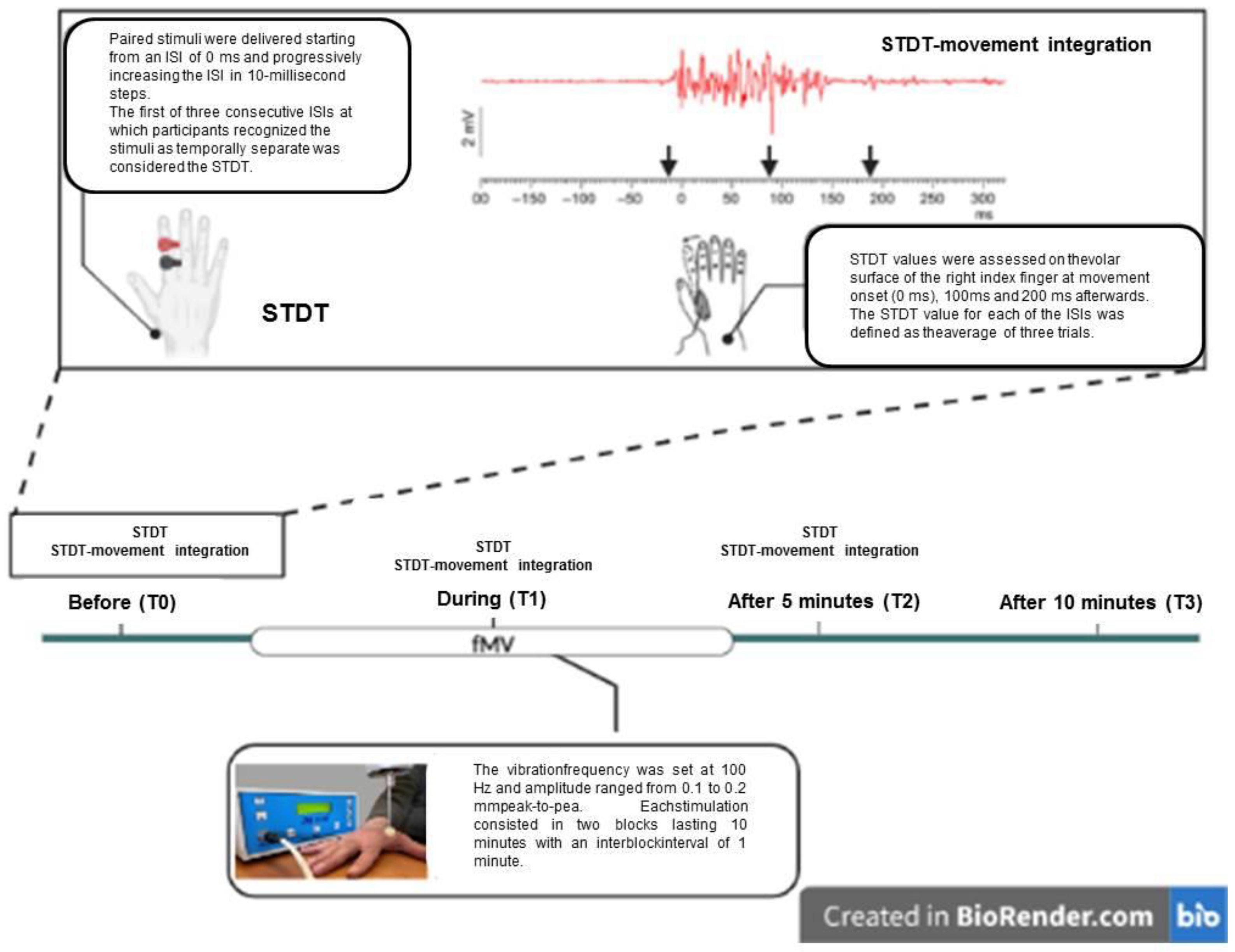
2.4. STDT–Movement Integration Paradigm
2.5. FMV
2.6. Experimental Paradigm
- Experiment 1: Effects of fMV applied over the FDI muscle on STDT and STDT–movement integration
- Experiment 2 (fMV topographic specificity): effects of fMV applied over the abductor pollicis brevis (APB) or the flexor radialis carpi (FRC) muscles on STDT and STDT–movement integration
- Experiment 3: Effects of fMV applied over the APB generating a 0.2 mm muscle displacement
2.7. Statistical Analysis
3. Results
- Experiment 1: Effects of fMV applied over the FDI muscle on STDT and STDT–movement integration
- Experiment 2: fMV topographic specificity effects of fMV applied over the APB or the FRC muscles on STDT–movement integration task
- Experiment 3: Effects of fMV applied over the APB generating a different muscle displacement
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lacruz, F.; Artieda, J.; Pastor, M.A.; Obeso, J.A. The anatomical basis of somaesthetic temporal discrimination in humans. J. Neurol. Neurosurg. Psychiatry 1991, 54, 1077–1081. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; McGovern, E.M.; Narasimham, S.; Beck, R.; Killian, O.; O’Riordan, S.; Reilly, R.B.; Hutchinson, M. Temporal Discrimination: Mechanisms and Relevance to Adult-Onset Dystonia. Front. Neurol. 2017, 8, 625. [Google Scholar] [CrossRef]
- Conte, A.; Belvisi, D.; Tartaglia, M.; Cortese, F.N.; Baione, V.; Battista, E.; Zhu, X.Y.; Fabbrini, G.; Berardelli, A. Abnormal Temporal Coupling of Tactile Perception and Motor Action in Parkinson’s Disease. Front. Neurol. 2017, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Rocchi, L.; Nardella, A.; Dispenza, S.; Scontrini, A.; Khan, N.; Berardelli, A. Theta-burst stimulation-induced plasticity over primary somatosensory cortex changes somatosensory temporal discrimination in healthy humans. PLoS ONE 2012, 7, e32979. [Google Scholar] [CrossRef]
- Rocchi, L.; Casula, E.; Tocco, P.; Berardelli, A.; Rothwell, J. Somatosensory Temporal Discrimination Threshold Involves Inhibitory Mechanisms in the Primary Somatosensory Area. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Pastor, M.A.; Day, B.L.; Macaluso, E.; Friston, K.J.; Frackowiak, R.S.J. The functional neuroanatomy of temporal discrimination. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 2585–2591. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Belvisi, D.; Manzo, N.; Bologna, M.; Barone, F.; Tartaglia, M.; Upadhyay, N.; Berardelli, A. Understanding the link between somatosensory temporal discrimination and movement execution in healthy subjects. Physiol. Rep. 2016, 4, e12899. [Google Scholar] [CrossRef]
- Yoshida, N.; Suzuki, T.; Ogahara, K.; Higashi, T.; Sugawara, K. Somatosensory temporal discrimination threshold changes during motor learning. Somatosens. Mot. Res. 2020, 37, 313–319. [Google Scholar] [CrossRef]
- Conte, A.; Belvisi, D.; De Bartolo, M.I.; Manzo, N.; Cortese, F.N.; Tartaglia, M.; Ferrazzano, G.; Fabbrini, G.; Berardelli, A. Abnormal sensory gating in patients with different types of focal dystonias. Mov. Disord. Off. J. Mov. Disord. Soc. 2018, 33, 1910–1917. [Google Scholar] [CrossRef]
- Scontrini, A.; Conte, A.; Defazio, G.; Fiorio, M.; Fabbrini, G.; Suppa, A.; Tinazzi, M.; Berardelli, A. Somatosensory temporal discrimination in patients with primary focal dystonia. J. Neurol. Neurosurg. Psychiatry 2009, 80, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Rossini, P.M.; Filippi, M.M.; Vernieri, F. Neurophysiology of sensorimotor integration in Parkinson’s disease. Clin. Neurosci. 1998, 5, 121–130. [Google Scholar]
- Lewis, G.N.; Byblow, W.D. Altered sensorimotor integration in Parkinson’s disease. Brain J. Neurol. 2002, 125, 2089–2099. [Google Scholar] [CrossRef] [Green Version]
- Abbruzzese, G.; Berardelli, A. Sensorimotor integration in movement disorders. Mov. Disord. Off. J. Mov. Disord. Soc. 2003, 18, 231–240. [Google Scholar] [CrossRef]
- Rosenkranz, K.; Rothwell, J.C. Modulation of proprioceptive integration in the motor cortex shapes human motor learning. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 9000–9006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, K.; Pesenti, A.; Paulus, W.; Tergau, F. Focal reduction of intracortical inhibition in the motor cortex by selective proprioceptive stimulation. Exp. Brain Res. 2003, 149, 9–16. [Google Scholar] [CrossRef]
- Buchner, R.; Van der Glas, H.W.; Brouwers, J.E.; Bosman, F. Electromyographic parameters related to clenching level and jaw-jerk reflex in patients with a simple type of myogenous cranio-mandibular disorder. J. Oral Rehabil. 1992, 19, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Lapole, T.; Tindel, J. Acute effects of muscle vibration on sensorimotor integration. Neurosci. Lett. 2015, 587, 46–50. [Google Scholar] [CrossRef]
- Souron, R.; Baudry, S.; Millet, G.Y.; Lapole, T. Vibration-induced depression in spinal loop excitability revisited. J. Physiol. 2019, 597, 5179–5193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, B. Sensory mechanisms in dentine. Proc. R. Soc. Med. 1972, 65, 493–495. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, K.; Rothwell, J.C. Differential effect of muscle vibration on intracortical inhibitory circuits in humans. J. Physiol. 2003, 551, 649–660. [Google Scholar] [CrossRef]
- Toscano, M.; Celletti, C.; Viganò, A.; Altarocca, A.; Giuliani, G.; Jannini, T.B.; Mastria, G.; Ruggiero, M.; Maestrini, I.; Vicenzini, E.; et al. Short-Term Effects of Focal Muscle Vibration on Motor Recovery After Acute Stroke: A Pilot Randomized Sham-Controlled Study. Front. Neurol. 2019, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia. 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Caliandro, P.; Celletti, C.; Padua, L.; Minciotti, I.; Russo, G.; Granata, G.; La Torre, G.; Granieri, E.; Camerota, F. Focal muscle vibration in the treatment of upper limb spasticity: A pilot randomized controlled trial in patients with chronic stroke. Arch. Phys. Med. Rehabil. 2012, 93, 1656–1661. [Google Scholar] [CrossRef]
- Marconi, B.; Filippi, G.M.; Koch, G.; Giacobbe, V.; Pecchioli, C.; Versace, V.; Camerota, F.; Saraceni, V.M.; Caltagirone, C. Long-term effects on cortical excitability and motor recovery induced by repeated muscle vibration in chronic stroke patients. Neurorehabil. Neural Repair 2011, 25, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Pujari, A.N.; Neilson, R.D.; Cardinale, M. Effects of different vibration frequencies, amplitudes and contraction levels on lower limb muscles during graded isometric contractions superimposed on whole body vibration stimulation. J. Rehabil. Assist. Technol. Eng. 2019, 6, 2055668319827466. [Google Scholar] [CrossRef] [Green Version]
- Tanji, J.; Wise, S.P. Submodality distribution in sensorimotor cortex of the unanesthetized monkey. J. Neurophysiol. 1981, 45, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Johnson, K.O.; Hsiao, S.S. Spatial pattern representation and transformation in monkey somatosensory cortex. Proc. Natl. Acad. Sci. USA 1988, 85, 1317–1321. [Google Scholar] [CrossRef] [Green Version]
- Colder, B. The basal ganglia select the expected sensory input used for predictive coding. Front. Comput. Neurosci. 2015, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Crutcher, M.D.; DeLong, M.R. Single cell studies of the primate putamen. I. Functional organization. Exp. Brain Res. 1984, 53, 233–243. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.R.; Vernon, W.B.; Bollinger, R.R. Sensitivity and specificity of a monitoring test. Biometrics 1985, 41, 947–958. [Google Scholar] [CrossRef]
- Naito, E.; Morita, T.; Amemiya, K. Body representations in the human brain revealed by kinesthetic illusions and their essential contributions to motor control and corporeal awareness. Neurosci. Res. 2016, 104, 16–30. [Google Scholar] [CrossRef]
- Rocchi, L.; Suppa, A.; Leodori, G.; Celletti, C.; Camerota, F.; Rothwell, J.; Berardelli, A. Corrigendum: Plasticity Induced in the Human Spinal Cord by Focal Muscle Vibration. Front. Neurol. 2018, 9, 1170. [Google Scholar] [CrossRef] [PubMed]
- Lopez, S.; Bini, F.; Del Percio, C.; Marinozzi, F.; Celletti, C.; Suppa, A.; Ferri, R.; Staltari, E.; Camerota, F.; Babiloni, C. Electroencephalographic sensorimotor rhythms are modulated in the acute phase following focal vibration in healthy subjects. Neuroscience 2017, 352, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Aviles, M.; Sánchez-Reyes, L.-M.; Fuentes-Aguilar, R.Q.; Toledo-Pérez, D.C.; Rodríguez-Reséndiz, J. A Novel Methodology for Classifying EMG Movements Based on SVM and Genetic Algorithms. Micromachines 2022, 13, 2108. [Google Scholar] [CrossRef] [PubMed]
- Antelmi, E.; Erro, R.; Rocchi, L.; Liguori, R.; Tinazzi, M.; Di Stasio, F.; Berardelli, A.; Rothwell, J.C.; Bhatia, K.P. Neurophysiological correlates of abnormal somatosensory temporal discrimination in dystonia. Mov. Disord. Off. J. Mov. Disord. Soc. 2017, 32, 141–148. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Protocol | Experiment 1 | Experiment 2 | Experiment 3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Muscle fMV | First Dorsali Interosseus | Abductor Pollicis Brevis | Flexor Radialis Carpi | Abductor Pollicis Brevis | ||||||||||
| Time | T0 | T1 | T2 | T3 | T0 | T1 | T2 | T0 | T1 | T2 | T0 | T1 | T2 | |
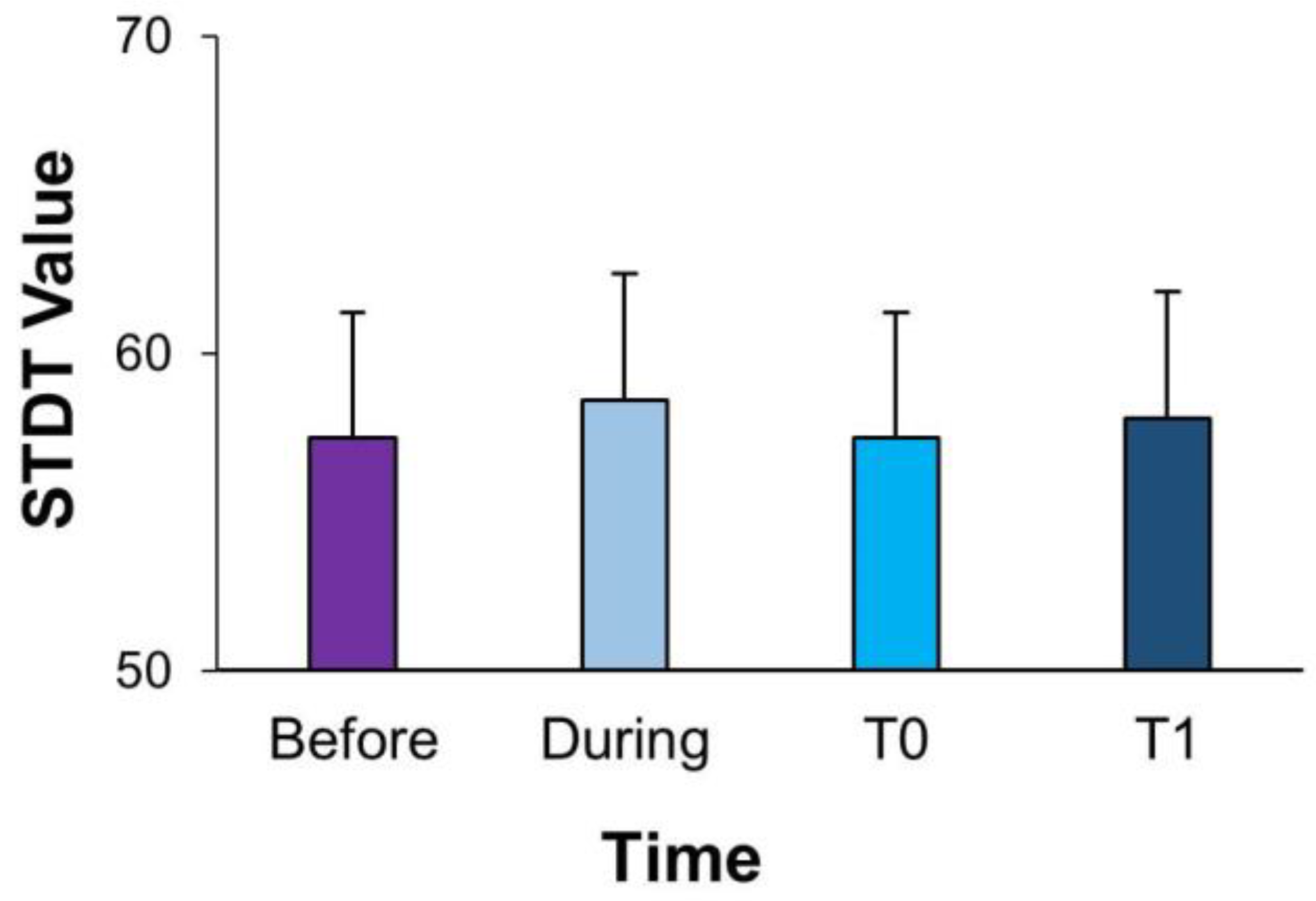
| STDT-At rest | 57.35 ± 3.93 | 58.53 ± 3.97 | 57.35 ± 3.93 | 57.94 ± 4.0 | 49.77 ± 3.23 | 52.44 ± 2.75 | 52.00 ± 2.96 | 52.33 ± 2.80 | 53.00 ± 3.44 | 52.67 ± 3.30 | 49.77 ± 3.23 | 52.44 ± 2.75 | 52.00 ± 2.96 | |
| STDT-movement integration | ||||||||||||||
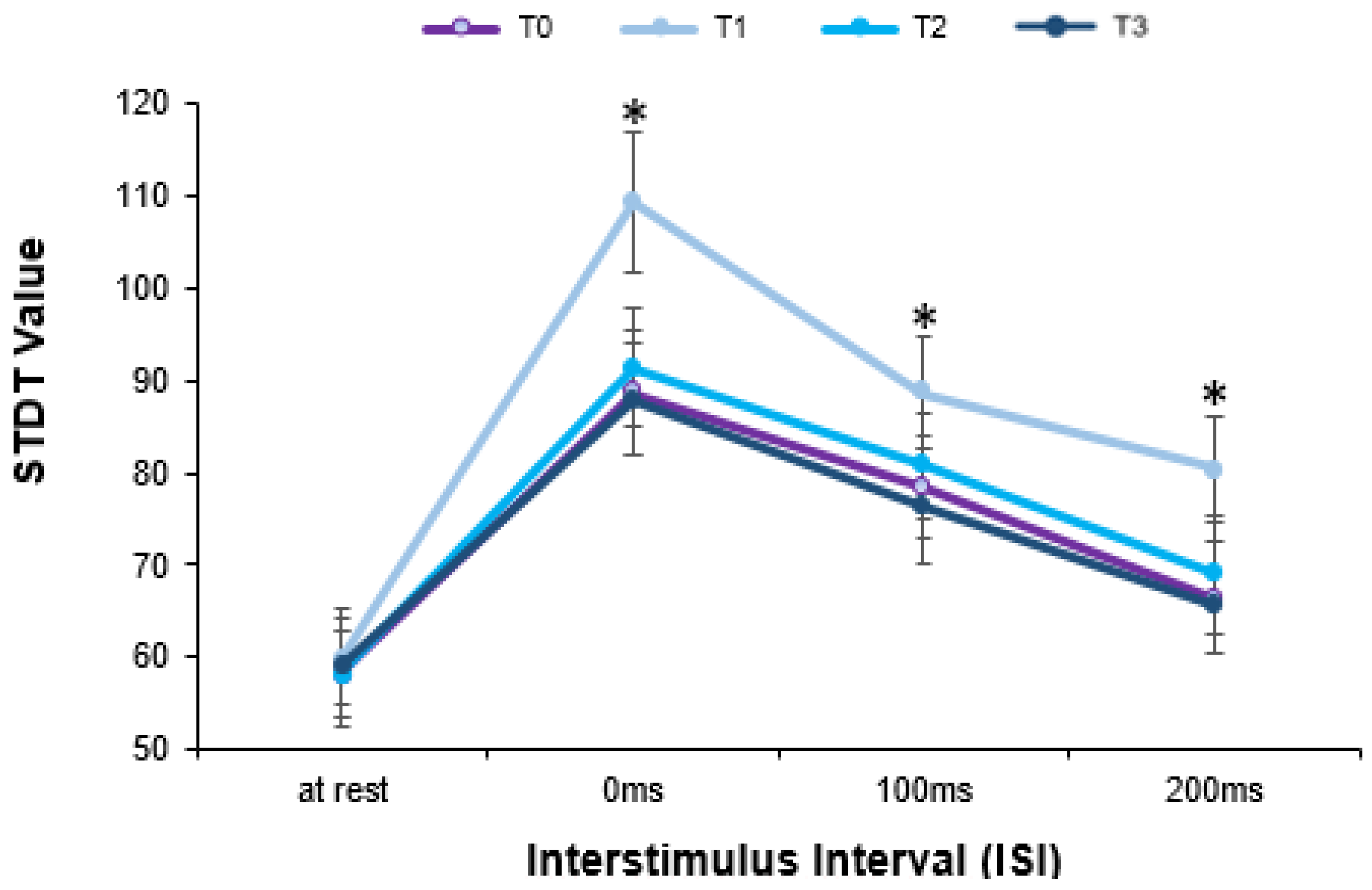
| 0 ms | 87.65 ± 5.79 | 107.06 ± 6.57 | 90.59 ± 5.52 | 85.88 ± 5.56 | 88.33 ± 5.91 | 88.00 ± 6.38 | 86.25 ± 6.59 | 83.67 ± 5.08 | 84.00 ± 4.34 | 83.46 ± 4.84 | 88.33 ± 5.91 | 88.00 ± 6.38 | 86.25 ± 6.59 | |
| 100 ms | 78.24 ± 4.72 | 89.41 ± 5.11 | 80.00 ± 5.0 | 77.65 ± 5.32 | 75.44 ± 4.43 | 78.07 ± 3.56 | 76.11 ± 3.62 | 76.0 ± 5.24 | 78.0 ± 5.21 | 72.68 ± 4.40 | 75.44 ± 4.43 | 78.07 ± 3.56 | 76.11 ± 3.62 | |
| 200 ms | 65.29 ± 5.21 | 80.29 ± 4.90 | 67.35 ± 5.46 | 64.71 ± 5.41 | 61.00 ± 4.12 | 65.33 ± 4.84 | 64.17 ± 4.50 | 61.67 ± 5.06 | 64.00 ± 5.76 | 60.38 ± 5.12 | 61.00 ± 4.12 | 65.33 ± 4.84 | 64.17 ± 4.50 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzo, N.; Ginatempo, F.; Belvisi, D.; Arcara, G.; Parrotta, I.; Leodori, G.; Deriu, F.; Celletti, C.; Camerota, F.; Conte, A. Investigating the Effects of a Focal Muscle Vibration Protocol on Sensorimotor Integration in Healthy Subjects. Brain Sci. 2023, 13, 664. https://doi.org/10.3390/brainsci13040664
Manzo N, Ginatempo F, Belvisi D, Arcara G, Parrotta I, Leodori G, Deriu F, Celletti C, Camerota F, Conte A. Investigating the Effects of a Focal Muscle Vibration Protocol on Sensorimotor Integration in Healthy Subjects. Brain Sciences. 2023; 13(4):664. https://doi.org/10.3390/brainsci13040664
Chicago/Turabian StyleManzo, Nicoletta, Francesca Ginatempo, Daniele Belvisi, Giorgio Arcara, Ilaria Parrotta, Giorgio Leodori, Franca Deriu, Claudia Celletti, Filippo Camerota, and Antonella Conte. 2023. "Investigating the Effects of a Focal Muscle Vibration Protocol on Sensorimotor Integration in Healthy Subjects" Brain Sciences 13, no. 4: 664. https://doi.org/10.3390/brainsci13040664