
Electrophysiological Evidence for Interhemispheric Connectivity and Communication in Young Human Infants
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Advantages of Hemispheric Coordination and Its Developmental Meaningfulness
1.2. Commissural Connectivity
1.3. The Present Study
2. Methods
2.1. Participants
2.2. Procedures
2.3. EEG Recording
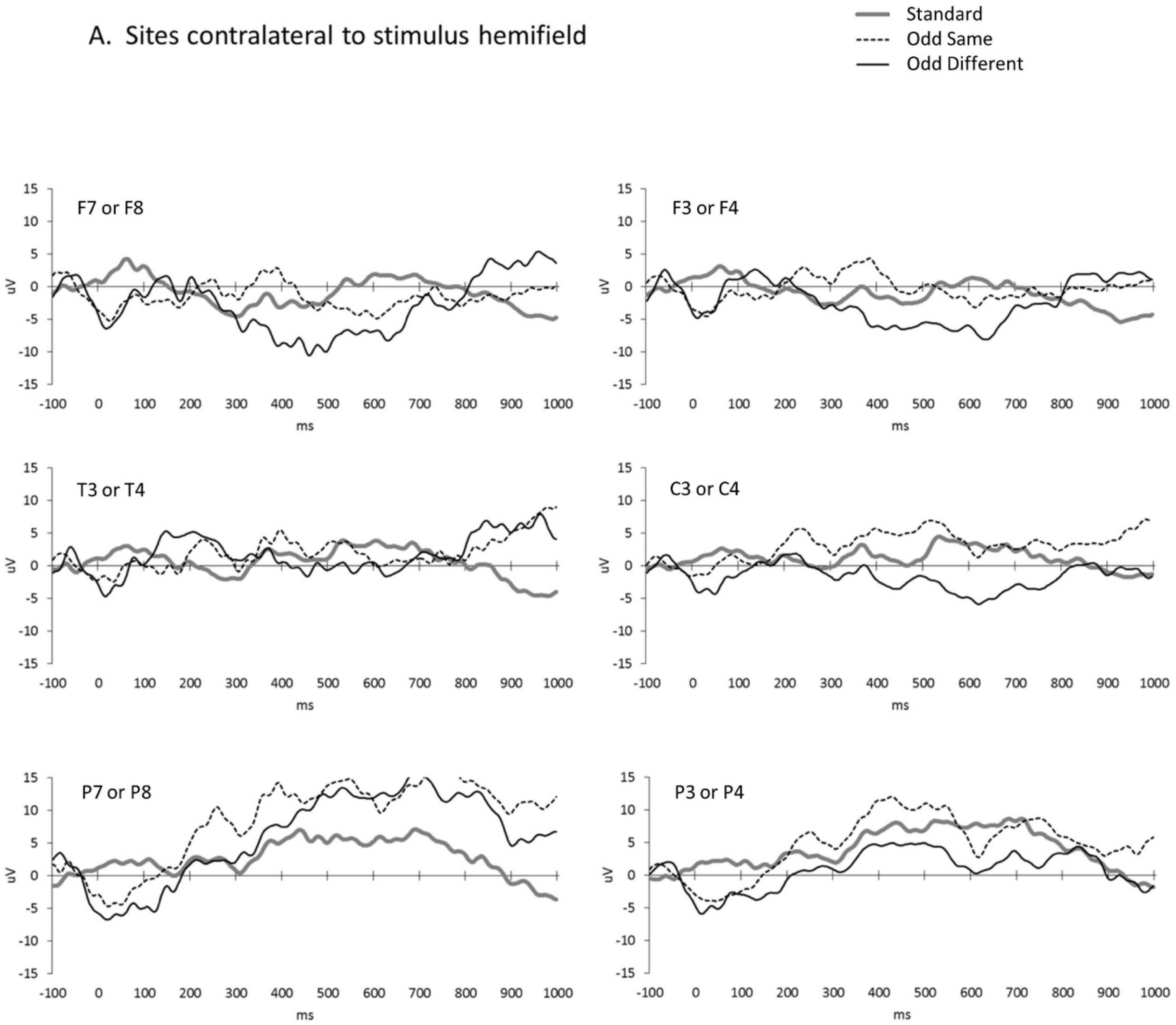
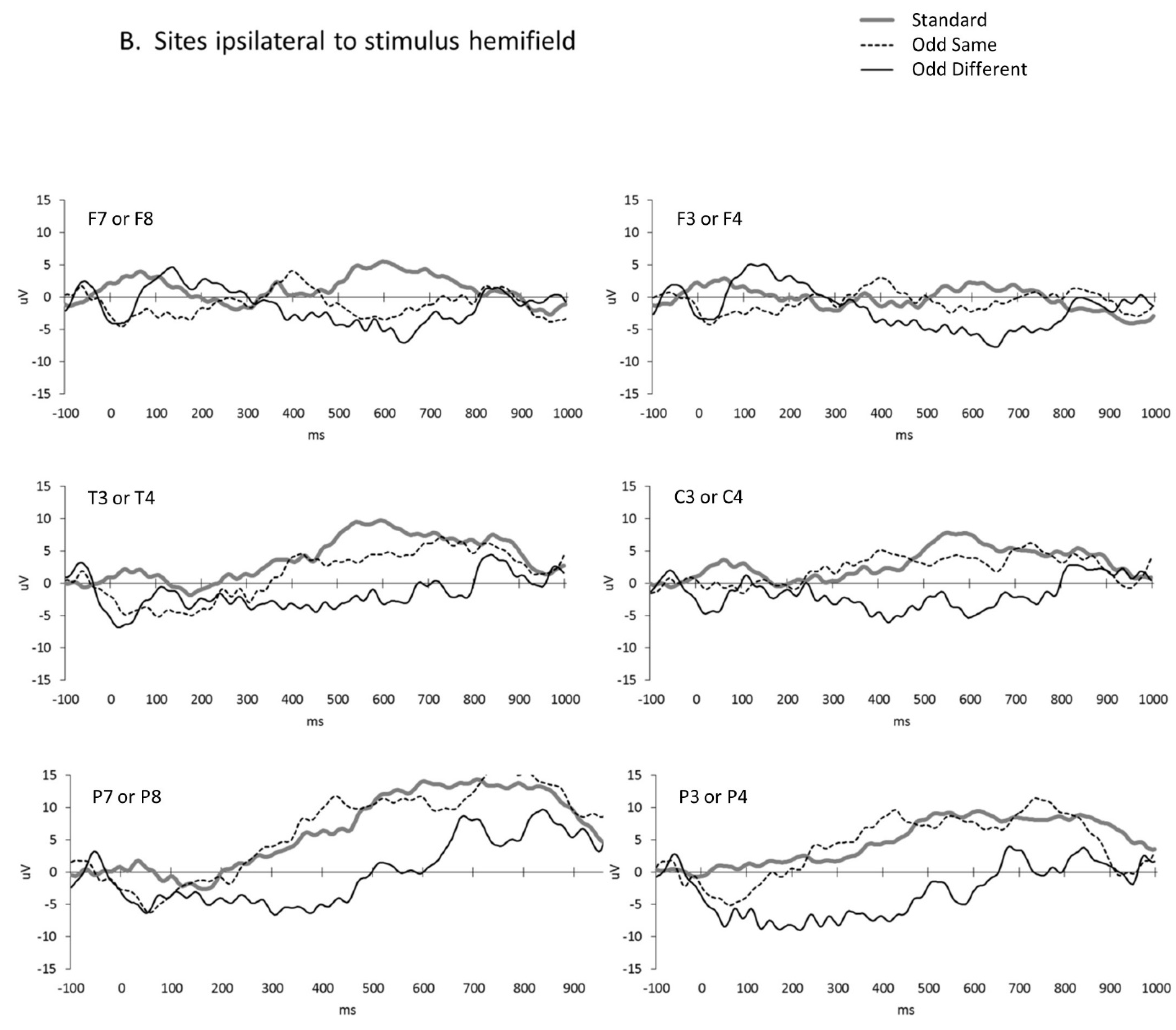
3. Results
4. Discussion
4.1. Limitations
4.2. Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- de Haan, E.H.; Corballis, P.M.; Hillyard, S.A.; Marzi, C.A.; Seth, A.; Lamme, V.A.; Volz, L.; Fabri, M.; Schechter, E.; Bayne, T.; et al. Split-brain: What we know now and why this is important for understanding consciousness. Neuropsychol. Rev. 2020, 30, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Gazzaniga, M.S. The Bisected Brain; Appleton-Century-Crofts: New York, NY, USA, 1970. [Google Scholar]
- Zaidel, E. Language functions in the two hemispheres following complete cerebral commissurotomy and hemispherectomy. In Handbook of Neuropsychology; Nebes, R.D., Corkin, S., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1990; pp. 115–150. [Google Scholar]
- Brysbaert, M. Lateral preferences and visual field asymmetries: Appearances may have been overstated. Cortex 1994, 30, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corballis, M.C. Can commissurotomized subjects compare digits between the visual fields? Neuropsychologia 1994, 32, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Gazzaniga, M.S. Perceptual and attentional processes following callosal section in humans. Neuropsychologia 1987, 25, 119–133. [Google Scholar] [CrossRef]
- Johnson, L.E. Bilateral visual cross-integration by human forebrain commissurotomy subjects. Neuropsychologia 1984, 22, 167–175. [Google Scholar] [CrossRef]
- Sergent, J. Unified response to bilateral hemispheric stimulation by a split-brain patient. Nature 1983, 305, 800–802. [Google Scholar] [CrossRef]
- Sergent, J. Processing of spatial relations within and between the disconnected cerebral hemispheres. Brain 1991, 11, 1025–1043. [Google Scholar] [CrossRef]
- Seymour, S.E.; Reuter-Lorenz, P.A.; Gazzaniga, M.S. The disconnection syndrome: Basic findings reaffirmed. Brain 1994, 117, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Koeda, T.; Knyazeva, M.; Njiokiktjien, C.; Jonkman, E.J.; De Sonneville, L.; Vildavsky, V. The EEG in acallosal children. Coherence values in the resting state: Left hemisphere compensatory mechanism? Electroencephalogr. Clin. Neurophysiol. 1995, 95, 397–407. [Google Scholar] [CrossRef]
- Mancuso, L.; Uddin, L.Q.; Nani, A.; Costa, T.; Cauda, F. Brain functional connectivity in individuals with callosotomy and agenesis of the corpus callosum: A systematic review. Neurosci. Biobehav. Rev. 2019, 105, 231–248. [Google Scholar] [CrossRef]
- Engel, A.; Konig, P.; Kreiter, A.; Singer, W. Interhemispheric synchronization of oscillatory neuronal responses in cat visual cortex. Science 1991, 252, 1177–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, C.S.; Peltier, S.J.; Wiggins, J.L.; Weng, S.-J.; Carrasco, M.; Risi, S.; Lord, C. Abnormalities of intrinsic functional connectivity in autism spectrum disorders. Neuroimage 2009, 47, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liederman, J.; Meehan, P. When is between-hemisphere division of labor advantageous? Neuropsychologia 1986, 24, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, T.E.; Jeeves, M.A.; Norman, W.D.; DeWitt, R. The bilateral field advantage on a letter-matching task. Cortex 1993, 29, 691–713. [Google Scholar] [CrossRef]
- Norman, W.D.; Jeeves, M.A.; Milne, A.; Ludwig, T. Hemispheric interactions: The bilateral advantage and task difficulty. Cortex 1992, 28, 623–642. [Google Scholar] [CrossRef]
- Liégeois, F.; Bentejac, L.; de Schonen, S. When does inter-hemispheric integration of visual events emerge in infancy? A developmental study on 19- to 28-month-old infants. Neuropsychologia 2000, 38, 1382–1389. [Google Scholar] [CrossRef]
- Gazzaniga, M.S. Cerebral specialization and interhemispheric communication: Does the corpus callosum enable the human condition? Brain 2000, 123, 1293–1326. [Google Scholar] [CrossRef] [Green Version]
- Sperry, R.W.; Gazzaniga, M.S.; Bogen, J.E. Interhemispheric relationships: The neocortical commissures and syndromes of hemisphere disconnection. In Handbook of Clinical Neurology; Vinken, P.J., Bruyn, G.W., Eds.; North-Holland Publishing: Amsterdam, The Netherlands, 1969; pp. 177–184. [Google Scholar]
- Reil, J.C. Untersuchungen über den Bau des grossen Gehirns im Menschen. Arch. Physiol. 1809, 9, 136–208. [Google Scholar]
- Aboitiz, F.; Montiel, J. One hundred million years of interhemispheric communication: The history of the corpus callosum. Braz. J. Med. Biol. Res. 2003, 36, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Pandya, D.N.; Seltzer, B. The topography of commissural fibers. In Two Hemispheres, One Brain: Functions of the Corpus Callosum; Lepore, F., Ptito, M., Jasper, H.H., Alan, R., Eds.; Liss: New York, NY, USA, 1986; pp. 46–53. [Google Scholar]
- Luders, E.; Narr, K.L.; Bilder, R.M.; Thompson, P.M.; Szeszko, P.R.; Hamilton, L.; Toga, A.W. Positive correlations between corpus callosum thickness and intelligence. Neuroimage 2007, 37, 1457–1464. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kim, J.J.; Lee, S.K.; Seok, J.H.; Chun, J.; Kim, D.I.; Lee, J.D. Corpus callosal connection mapping using cortical gray matter parcellation and DT-MRI. Hum. Brain Mapp. 2008, 29, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Berardi, N.; Fiorentini, A. Interhemispheric transfer of visual information in humans: Spatial characteristics. J. Physiol. 1987, 384, 633–647. [Google Scholar] [CrossRef]
- De Schonen, S.; De Diaz, M.G.; Mathivet, E. Hemispheric asymmetry in face processing in infancy. In Aspects of Face Processing; Ellis, H.D., Jeeves, M., Newcombe, F., Young, A., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 199–209. [Google Scholar] [CrossRef]
- De Schonen, S.; Mathivet, E. Hemispheric asymmetry in a face discrimination task in infants. Child Dev. 1990, 61, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Deruelle, C.; de Schonen, S. Hemispheric asymmetries in visual pattern processing in infancy. Brain Cogn. 1991, 16, 151–179. [Google Scholar] [CrossRef] [PubMed]
- Deruelle, C.; de Schonen, S. Pattern processing in infancy: Hemispheric differences in the processing of shape and location of visual components. Infant Behav. Dev. 1995, 18, 123–132. [Google Scholar] [CrossRef]
- Rakic, P.; Yakovlev, P.I. Development of the corpus callosum and cavum septi in man. J. Comp. Neurol. 1968, 132, 45–72. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, P.I.; Lecours, A.R. The myelogenetic cycles of regional maturation of the brain. In Regional Development of the Brain in Early Life; de Rosnay, M., Murray, L., Eds.; Blackwell Scientific: Oxford, UK, 1967; pp. 3–70. [Google Scholar]
- Keshavan, M.S.; Diwadkar, V.A.; DeBellis, M.; Dick, E.; Kotwal, R.; Rosenberg, D.R.; Sweeney, J.A.; Minshew, N.; Pettegrew, J.W. Development of the corpus callosum in childhood, adolescence and early adulthood. Life Sci. 2002, 70, 1909–1922. [Google Scholar] [CrossRef]
- Salamy, A. Commissural transmission: Maturational changes in humans. Science 1978, 200, 1409–1411. [Google Scholar] [CrossRef]
- Liederman, J.; Merola, J.L.; Hoffman, C. Longitudinal data indicate that hemispheric independence increases during early adolescence. Dev. Neuropsychol. 1986, 2, 183–201. [Google Scholar] [CrossRef]
- Bornstein, M.H. Intelligence in infancy. In The Cambridge Handbook of Intelligence; Sternberg, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2020; pp. 124–154. [Google Scholar] [CrossRef]
- Thomason, M.E.; Dassanayake, M.T.; Shen, S.; Katkuri, Y.; Alexis, M.; Anderson, A.L.; Yeo, L.; Mody, S.; Hernandez-Andrade, E.; Hassan, S.S.; et al. Cross-hemispheric functional connectivity in the human fetal brain. Sci. Transl. Med. 2013, 5, 173ra24. [Google Scholar] [CrossRef] [Green Version]
- Adibpour, P.; Dubois, J.; Dehaene-Lambertz, G. Right but not left hemispheric discrimination of faces in infancy. Nat. Hum. Behav. 2017, 2, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreassi, J.L.; Okamura, H.; Stern, M. Hemispheric asymmetries in the visual cortical evoked potential as a function of stimulus location. Psychophysiology 1975, 12, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Srebro, R. The topography of scalp potentials evoked by pattern pulse stimuli. Vis. Res. 1987, 27, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Saron, C.D.; Davidson, R.J. Visual evoked potential measures of interhemispheric transfer time in humans. Behav. Neurosci. 1989, 103, 115–1138. [Google Scholar] [CrossRef]
- Ionta, S. Visual neuropsychology in development: Anatomo-functional brain mechanisms of action/perception binding in health and disease. Front. Hum. Neurosci. 2021, 15, 689912. [Google Scholar] [CrossRef]
- Johnson, M.H. Developmental neuroscience psychophysiology, and genetics. In Developmental Science: An Advanced Textbook, 7th ed.; Bornstein, M.H., Lamb, M.E., Eds.; Psychology Press: New York, NY, USA, 2015; pp. 217–260. [Google Scholar]
- Johnson, M.H.; Morton, J. Biology and Cognitive Development; Blackwell: Oxford, UK, 1991. [Google Scholar]
- Haith, M.M. Progress and standardization in eye movement work with human infants. Infancy 2004, 6, 257–265. [Google Scholar] [CrossRef]
- Johnson, M.H.; Dziurawiec, S.; Ellis, H.; Morton, J. Newborn’s preferential tracking of face-like stimuli and its subsequent decline. Cognition 1991, 40, 1–19. [Google Scholar] [CrossRef]
- Quinn, P.C.; Kelly, D.J.; Lee, K.; Pascalis, O.; Slater, A.M. Preference for attractive faces in human infants extends beyond conspecifics. Dev. Sci. 2008, 11, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, I.W. Mother’s face recognition in newborn infants: Learning and memory. Infant Child Dev. 2001, 10, 67–74. [Google Scholar] [CrossRef]
- Pascalis, O.; de Schonen, S.; Morton, J.; Deruelle, C.; Fabre-Grenet, M. Mother’s face recognition by neonates: A replication and an extension. Infant Behav. Dev. 1995, 18, 79–85. [Google Scholar] [CrossRef]
- Bornstein, M.H.; Arterberry, M.E. Recognition, discrimination, and categorization of smiling by 5-month-old infants. Dev. Sci. 2003, 6, 585–599. [Google Scholar] [CrossRef]
- Kuchuk, A.; Vibbert, M.; Bornstein, M.H. The perception of smiling and its experiential correlates in three-month-old infants. Child Dev. 1986, 57, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- De Heering, A.; Rossion, B. Rapid categorization of natural face images in the infant right hemisphere. eLife 2015, 4, e06564. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, R.S.; Bertin, E.; Hayden, A.; Reed, A. Face processing in infancy: Developmental changes in the use of different kinds of relational information. Child Dev. 2005, 76, 169–181. [Google Scholar] [CrossRef]
- Brooks, R.; Meltzoff, A.N. The development of gaze following and its relation to language. Dev. Sci. 2005, 8, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farroni, T.; Massaccesi, S.; Pividori, D.; Johnson, M.H. Gaze following in newborns. Infancy 2004, 5, 39–60. [Google Scholar] [CrossRef]
- Oakes, L.M. Sample size, statistical power, and false conclusions in infant looking-time research. Infancy 2017, 22, 436–469. [Google Scholar] [CrossRef]
- Knyazeva, M.G.; Kiper, D.C.; Vildavski, V.Y.; Despland, P.A.; Maeder-Ingvar, M.; Innocenti, G.M. Visual stimulus–dependent changes in interhemispheric EEG coherence in humans. J. Neurophysiol. 1999, 82, 3095–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paus, T.; Jech, R.; Thompson, C.J.; Comeau, R.; Peters, T.; Evans, A.C. Transcranial magnetic stimulation during positron emission tomography: A new method for studying connectivity of the human cerebral cortex. J. Neurosci. 1997, 17, 3178–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berardi, N.; Bodis-Wollner, I.; Fiorentini, A.; Giuffré, G.; Morelli, M. Electrophysiological evidence for interhemispheric transmission of visual information in man. J. Physiol. 1989, 411, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Fendrich, R.; Wessinger, C.M.; Gazzaniga, M. Nasotemporal overlap at the retinal vertical meridian: Investigations with a callosotomy patient. Neuropsychologia 1996, 34, 637–646. [Google Scholar] [CrossRef]
- Clarke, S.; Miklossy, J. Occipital cortex in man: Organization of callosal connections, related myelo-and cytoarchitecture, and putative boundaries of functional visual areas. J. Comp. Neurol. 1990, 298, 188–214. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2020, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Bunt, A.H.; Minckler, D.S.; Johanson, G.W. Demonstration of bilateral projection of the central retina of the monkey with horseradish peroxidase neuronography. J. Comp. Neurol. 1977, 171, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, Y.; Sawai, H.; Watanabe, M.; Wakakuwa, K.; Morigiwa, K. Nasotemporal overlap of crossed and uncrossed retinal ganglion cell projections in the Japanese monkey (Macaca fuscata). J. Neurosci. 1989, 9, 2353–2373. [Google Scholar] [CrossRef]
- Leventhal, A.; Ault, S.; Vitek, D. The nasotemporal division in primate retina: The neural bases of macular sparing and splitting. Science 1988, 240, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.; Leicester, J.; Sherman, S.M. The naso-temporal division of the monkey’s retina. J. Comp. Neurol. 1973, 150, 333–348. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Mognon, A.; Jovicich, J.; Bruzzone, L.; Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology 2011, 48, 229–240. [Google Scholar] [CrossRef]
- Richards, J.E.; Reynolds, G.D.; Courage, M.L. The neural bases of infant attention. Curr. Dir. Psychol. Sci. 2010, 19, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmaso, M.; Castelli, L.; Galfano, G. Social modulators of gaze-mediated orienting of attention: A review. Psychon. Bull. Rev. 2020, 27, 833–855. [Google Scholar] [CrossRef] [PubMed]
- McKay, K.T.; Grainger, S.A.; Coundouris, S.P.; Skorich, D.P.; Phillips, L.H.; Henry, J.D. Visual attentional orienting by eye gaze: A meta-analytic review of the gaze-cueing effect. Psychol. Bull. 2021, 147, 1269–1289. [Google Scholar] [CrossRef] [PubMed]
- Bremer, F.; Brihaye, J.; André-Balisaux, G. Physiologie et pathologie du corps calleux. Schweiz. Arch. Neurol. Psychiatr. 1956, 78, 31–97. [Google Scholar]
- Funnell, M.G.; Corballis, P.M.; Gazzaniga, M.S. Cortical and subcortical interhemispheric interactions following partial and complete callosotomy. Arch. Neurol. 2000, 57, 185–189. [Google Scholar] [CrossRef]
- Berlucchi, G.; Buchtel, E.; Marzi, C.A.; Mascetti, G.G.; Simoni, A. Effects of experience on interocular transfer of pattern discriminations in split-chiasm & split-brain cats. J. Comp. Physiol. Psychol. 1978, 92, 532–543. [Google Scholar]
- Gazzaniga, M.S. Organization of the human brain. Science 1989, 245, 947–952. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, C.R. Mechanisms of interocular equivalence. In Analysis of Visual Behavior; Ingle, D.J., Goodale, M.A., Mansfield, R.J.W., Eds.; MIT Press: Cambridge, MA, USA, 1982; pp. 693–717. [Google Scholar]
- Jeeves, M.A.; Milner, A.D. Specificity and plasticity in interhemispheric integration: Evidence from callosal agenesis. In The Duality and Unity of the Brain: Unified Functioning and Specialization of the Hemispheres; Ottoson, D., Ed.; Palgrave Macmillan: London, UK, 1987; Volume 2, pp. 25–29. [Google Scholar] [CrossRef]
- Leporé, F.; Samson, A.; Ptito, M. Role of the corpus callosum in interhemispheric transfer of visual learning in the Siamese cat. Can. J. Psychol. 1983, 37, 535–546. [Google Scholar] [CrossRef]
- Sidtis, J.J.; Volpe, B.T.; Holtzman, J.D.; Wilson, D.H.; Gazzaniga, M.S. Cognitive interaction after staged callosal section: Evidence for transfer of semantic activation. Science 1981, 212, 344–346. [Google Scholar] [CrossRef]
- Sperry, R.W. Mental unity following surgical disconnection of the cerebral hemispheres. Harvey Lect. 1966, 62, 293–323. [Google Scholar]
- Bremer, F. La physiologie du corps calleux a la umiere de travaux recents. Lav. Med. 1967, 38, 835–843. [Google Scholar]
- Creutzfeldt, O.D.; Maekawa, K.; Hosli, L.C.R. Forms of spontaneous and evoked post-synaptic potentials of cortical nerve cells. Prog. Brain Res. 1969, 31, 265–274. [Google Scholar] [CrossRef]
- Aboitiz, F.; Scheibel, A.B.; Fisher, R.S.; Zaidel, E. Fiber composition of the human corpus callosum. Brain Res. 1993, 598, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Van Valkenburg, C.T. Experimental and pathologico-anatomical researches on the corpus callosum. Brain 1913, 36, 119–165. [Google Scholar] [CrossRef] [Green Version]
- Innocenti, G.M. General organization of callosal connections in the cerebral cortex. In Cerebral Cortex; Jones, G., Peters, A., Eds.; Plenum: New York, NY, USA, 1986; Volume 5, pp. 291–353. [Google Scholar] [CrossRef]
- Zilles, K.; Clarke, S. Architecture, connectivity and transmitter receptors of human extrastriate visual cortex: Comparison with non-human primates. In Cerebral Cortex; Jones, G., Peters, A., Rockland, K.S., Kaas, J.H., Peters, A., Eds.; Springer: Boston, MA, USA, 1997; Volume 12. [Google Scholar] [CrossRef]
- Johnson, S.P.; Hannon, E.E. Perceptual development. In Handbook of Child Psychology and Developmental Science: Cognitive Processes, 7th ed.; Liben, L.S., Müller, U., Lerner, R.M., Eds.; Wiley: Hoboken, NJ, USA, 2015; Volume 2, pp. 63–112. [Google Scholar] [CrossRef]
- Menacker, S.J.; Batshaw, M.L. Vision: Our Window to the World; In Children with Disabilities, 4th ed.; Batshaw, M.L., Paul, H., Eds.; Brookes: Baltimore, MD, USA, 1997; pp. 211–239. [Google Scholar]
- Farah, M.J.; Rabinowitz, C.; Quinn, G.E.; Liu, G.T. Early commitment of neural substrates for face recognition. Cogn. Neuropsychol. 2000, 17, 117–123. [Google Scholar] [CrossRef]
- Gauthier, I.; Nelson, C.A. The development of face expertise. Curr. Opin. Neurobiol. 2001, 11, 219–224. [Google Scholar] [CrossRef]
- Lettvin, J.Y.; Maturana, H.R.; McCulloch, W.S.; Pitts, W.H. What the frog’s eye tells the frog’s brain. Proc. Inst. Radio Eng. 1959, 47, 1940–1951. [Google Scholar] [CrossRef]
- Gross, C.G. The genealogy of the “grandmother cell”. Neuroscientist 2002, 8, 512–518. [Google Scholar] [CrossRef]
- Haxby, J.V.; Ungerleider, L.G.; Horwitz, B.; Rapoport, S.I.; Grady, C.L. Hemispheric differences in neural systems for face working memory: A PET-rCBF study. Hum. Brain Mapp. 1995, 3, 68–82. [Google Scholar] [CrossRef]
- Mash, C.; Bornstein, M.H.; Arterberry, M.E. Brain dynamics in young infants’ recognition of faces. Neuroreport 2013, 24, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Farah, R.; Ionta, S.; Horowitz-Kraus, T. Neuro-behavioral correlates of executive dysfunctions in dyslexia over development from childhood to adulthood. Front. Psychol. 2021, 12, 708863. [Google Scholar] [CrossRef]
- Bornstein, M.H. Human Infancy... and the Rest of the Lifespan. Annu. Rev. Psychol. 2014, 65, 121–158. [Google Scholar] [CrossRef]
- Stewart, A.L.; Rifkin, L.; Amess, P.N.; Kirkbride, V.; Townsend, J.P.; Miller, D.H.; Lewis, S.W.; Kingsley, D.P.; Moseley, I.F.; Foster, O.; et al. Brain structure and neurocognitive and behavioural function in adolescents who were born very preterm. Lancet 1999, 353, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Joseph, R.; Gallagher, R.E.; Holloway, W.; Kahn, J. Two brains, one child: Interhemispheric information transfer deficits and confabulatory responding in children aged 4, 7, 10. Cortex 1984, 20, 317–331. [Google Scholar] [CrossRef]
- Lassonde, M.; Sauerwein, H.; Geoffroy, G.; Décarie, M. Effects of early and late transection of the corpus callosum in children. Brain 1986, 109, 953–967. [Google Scholar] [CrossRef]
- Bluhm, R.L.; Miller, J.; Lanius, R.A.; Osuch, E.A.; Boksman, K.; Neufeld, R.W.; Théberge, J.; Schaefer, B.; Williamson, P. Spontaneous low-frequency fluctuations in the BOLD signal in schizophrenic patients: Anomalies in the default network. Schizophr. Bull. 2007, 33, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Bluhm, R.L.; Miller, J.; Lanius, R.A.; Osuch, E.A.; Boksman, K.; Neufeld, R.W.J.; Théberge, J.; Schaefer, B.; Williamson, P.C. Retrosplenial cortex connectivity in schizophrenia. Psychiatry Res. Neuroimaging 2009, 174, 17–23. [Google Scholar] [CrossRef]
- Bluhm, R.L.; Williamson, P.C.; Osuch, E.A.; Frewen, P.A.; Stevens, T.K.; Boksman, K.; Neufeld, R.W.; Théberge, J.; Lanius, R.A. Alterations in default network connectivity in posttraumatic stress disorder related to early-life trauma. J. Psychiatry Neurosci. 2009, 34, 187–194. [Google Scholar] [PubMed]
- Cao, Q.; Zang, Y.; Sun, L.; Sui, M.; Long, X.; Zou, Q.; Wang, Y. Abnormal neural activity in children with attention deficit hyperactivity disorder: A resting-state functional magnetic resonance imaging study. Neuroreport 2006, 17, 1033–1036. [Google Scholar] [CrossRef]
- Tian, L.; Jiang, T.; Wang, Y.; Zang, Y.; He, Y.; Liang, M.; Sui, M.; Cao, Q.; Hu, S.; Peng, M.; et al. Altered resting-state functional connectivity patterns of anterior cingulate cortex in adolescents with attention deficit hyperactivity disorder. Neurosci. Lett. 2006, 400, 39–43. [Google Scholar] [CrossRef]
- Weng, S.-J.; Wiggins, J.L.; Peltier, S.J.; Carrasco, M.; Risi, S.; Lord, C.; Monk, C.S. Alterations of resting state functional connectivity in the default network in adolescents with autism spectrum disorders. Brain Res. 2010, 1313, 202–214. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bornstein, M.H.; Mash, C.; Romero, R.; Gandjbakhche, A.H.; Nguyen, T. Electrophysiological Evidence for Interhemispheric Connectivity and Communication in Young Human Infants. Brain Sci. 2023, 13, 647. https://doi.org/10.3390/brainsci13040647
Bornstein MH, Mash C, Romero R, Gandjbakhche AH, Nguyen T. Electrophysiological Evidence for Interhemispheric Connectivity and Communication in Young Human Infants. Brain Sciences. 2023; 13(4):647. https://doi.org/10.3390/brainsci13040647
Chicago/Turabian StyleBornstein, Marc H., Clay Mash, Roberto Romero, Amir H. Gandjbakhche, and Thien Nguyen. 2023. "Electrophysiological Evidence for Interhemispheric Connectivity and Communication in Young Human Infants" Brain Sciences 13, no. 4: 647. https://doi.org/10.3390/brainsci13040647