

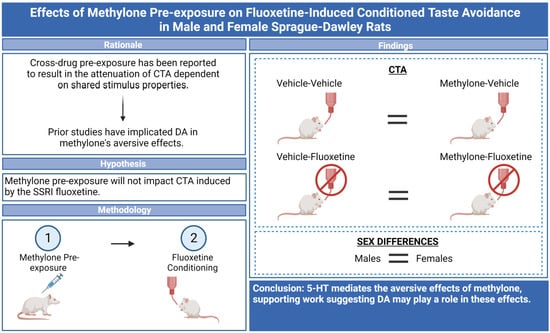
Effects of Methylone Pre-Exposure on Fluoxetine-Induced Conditioned Taste Avoidance in Male and Female Sprague-Dawley Rats

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Subjects
2.2. Drugs and Solutions
2.3. Apparatus
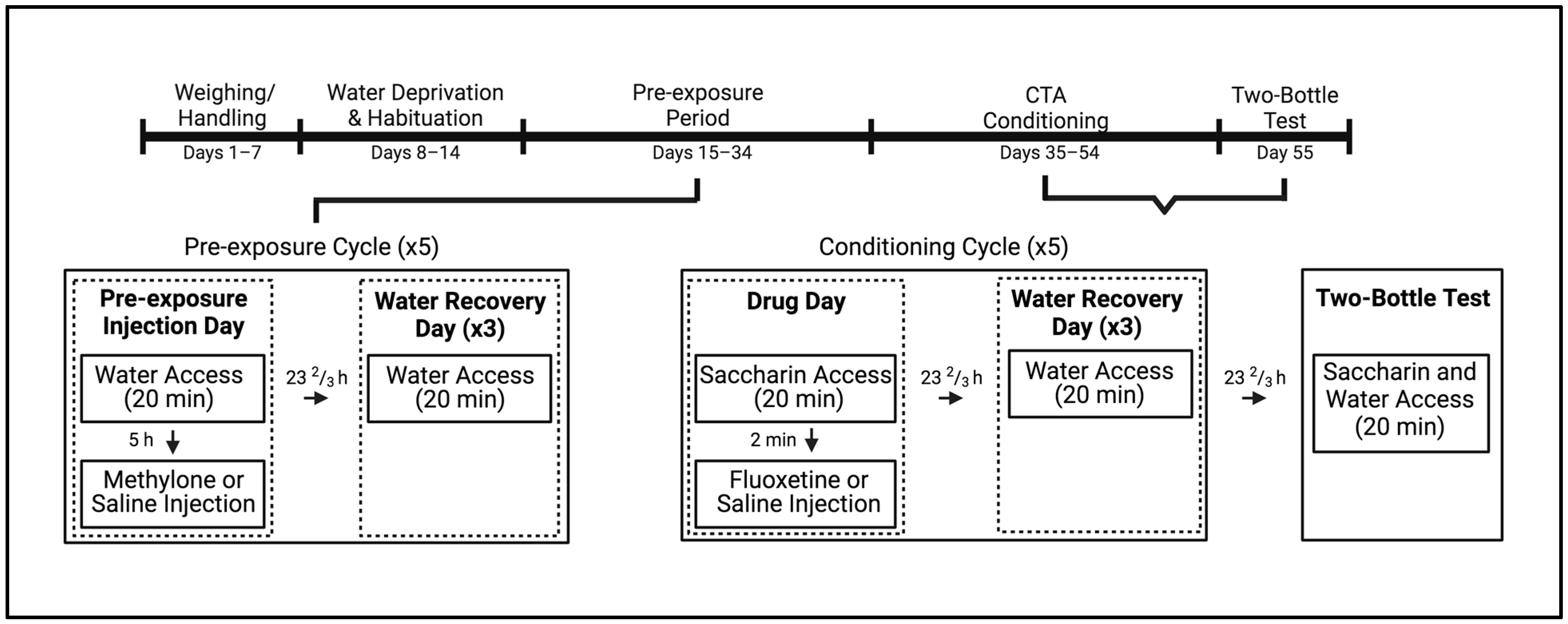
2.4. Procedure
2.4.1. Phase 1: Habituation
2.4.2. Phase 2: Pre-Exposure
2.4.3. Phase 3: Conditioned Taste Avoidance (CTA)
2.5. Statistical Analysis
3. Results
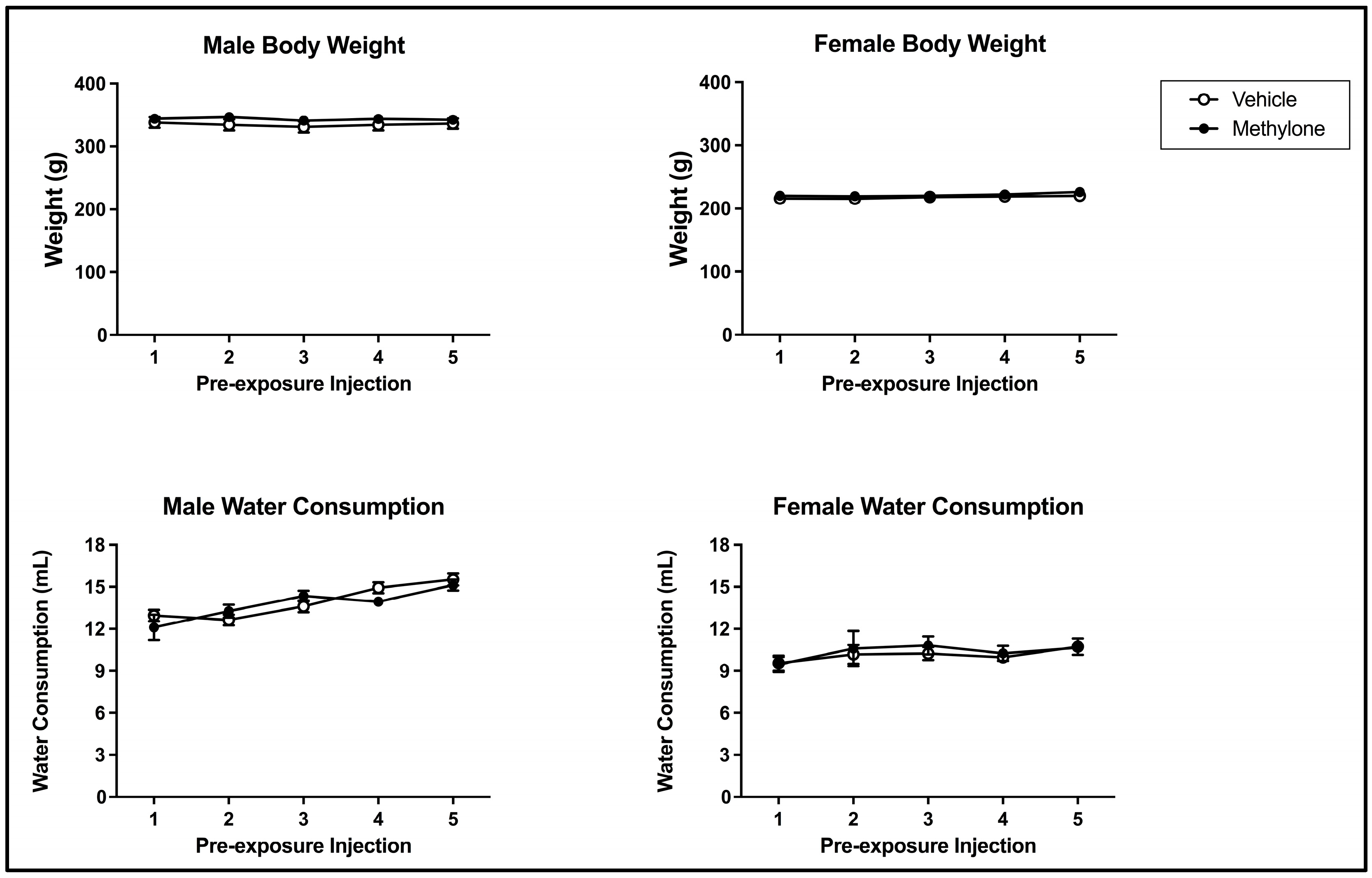
3.1. Bodyweight and Fluid Consumption over Pre-Exposure
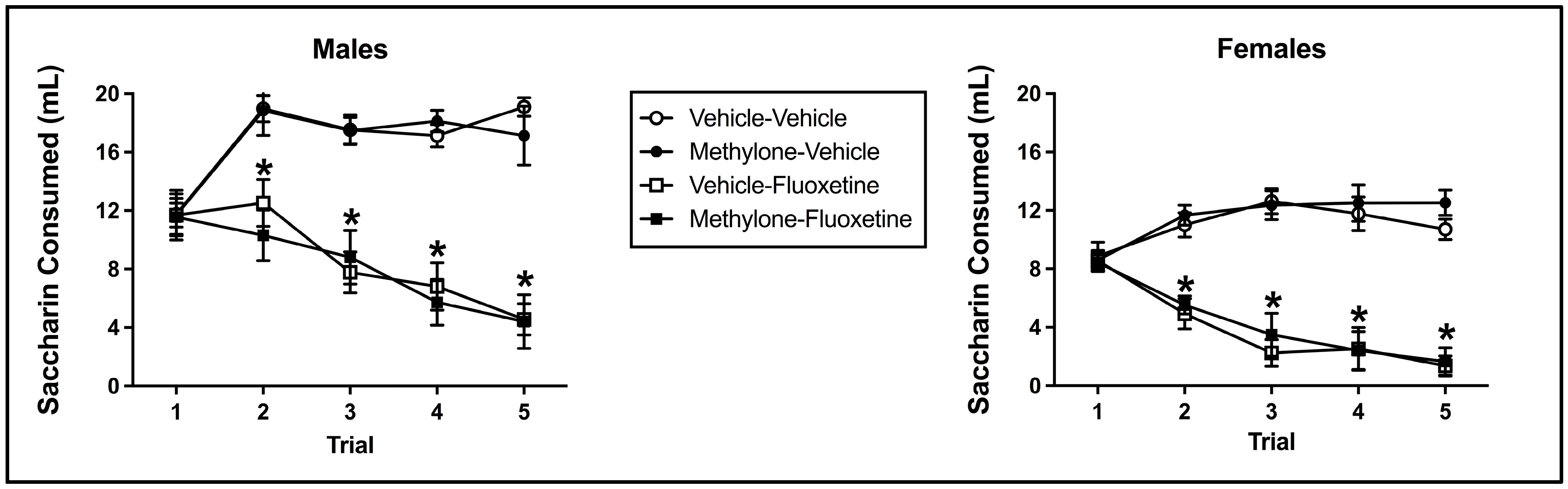
3.2. Conditioned Taste Avoidance
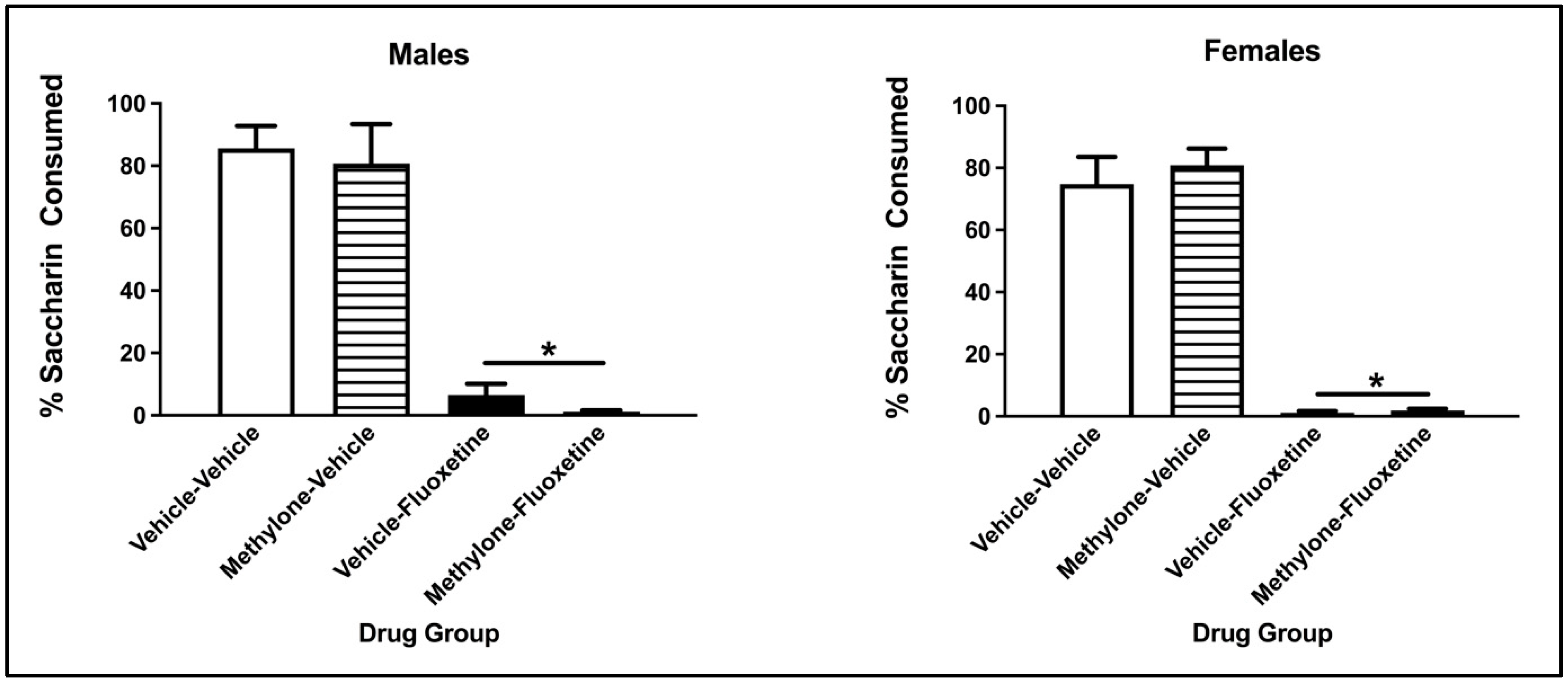
3.3. Two-Bottle Avoidance Test
4. General Discussion
5. Scope
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rzóska, J. Bait shyness, a study in rat behaviour. Brit. J. Anim. Behav. 1953, 1, 128–135. [Google Scholar] [CrossRef]
- Garcia, J.; Kimeldorf, D.J.; Koelling, R.A. Conditioned aversion to saccharin resulting from exposure to gamma radiation. Science 1955, 122, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Freeman, K.B.; Riley, A.L. The origins of conditioned taste aversion learning: A historical analysis. In Conditioned Taste Aversion: Neural and Behavioral Processes; Reilly, S., Schachtman, T.R., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 9–33. [Google Scholar]
- Garcia, J.; Ervin, F.R. Appetites, aversions, and addictions: A model for visceral memory. In Recent Advances in Biological Psychiatry; Wortis, J., Ed.; Springer: Boston, MA, USA, 1968; pp. 284–293. [Google Scholar]
- Revusky, S.; Garcia, J. Learned associations over long delays. In Psychology of Learning and Motivation; Bower, G.H., Ed.; Academic Press: Cambridge, MA, USA, 1970; pp. 1–84. [Google Scholar]
- Rozin, P.; Kalat, J.W. Specific hungers and poison avoidance as adaptive specializations of learning. Psychol. Rev. 1971, 78, 459–486. [Google Scholar] [CrossRef] [PubMed]
- Braveman, N.S. What studies on preexposure to pharmacological agents tell us about the nature of the aversion-inducing agent. In Learning Mechanisms in Food Selection; Barker, L.M., Best, M.R., Domjan, M., Eds.; Baylor University Press: Waco, TX, USA, 1977; pp. 511–530. [Google Scholar]
- Cappell, H.; LeBlanc, A.E. Parametric investigations of the effects of prior exposure to amphetamine and morphine on conditioned gustatory aversion. Psychopharmacology 1977, 51, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.; Amit, Z. Conditioned taste aversion induced by self-administered drugs: Paradox revisited. Neurosci. Biobehav. Rev. 1987, 11, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Riley, A.L.; Manke, H.M.; Huang, S. Impact of the aversive effects of drugs on their use and abuse. Behav. Neurol. 2022, 8634176. [Google Scholar] [CrossRef]
- Klosterhalfen, S.; Klosterhalfen, W. Conditioned taste aversion and traditional learning. Psychol. Res. 1985, 47, 71–94. [Google Scholar] [CrossRef]
- Randich, A.; LoLordo, V.M. Associative and nonassociative theories of the UCS preexposure phenomenon: Implications for Pavlovian conditioning. Psychol. Bull. 1979, 86, 523–548. [Google Scholar] [CrossRef]
- Riley, A.L.; Simpson, G.R. The attenuating effects of drug preexposure on taste aversion conditioning: Generality, experimental parameters, underlying mechanisms, and implications for drug use and abuse. In Handbook of Contemporary Learning Theories; Mowrer, R.R., Klein, S.B., Eds.; Lawrence Erlbaum Associates Inc.: Mahawah, NJ, USA, 2001; pp. 505–559. [Google Scholar]
- Berman, R.F.; Cannon, D.S. The effect of prior ethanol experience on ethanol induced saccharin aversions. Physiol. Behav. 1974, 12, 1041–1044. [Google Scholar] [CrossRef]
- Cappell, H.; LeBlanc, A.E. Conditioned aversion by amphetamine: Rates of acquisition and loss of the attenuating effects of prior exposure. Psychopharmacologia 1975, 43, 157–162. [Google Scholar] [CrossRef]
- Domjan, M.; Siegel, S. Attenuation of the aversive and analgesic effects of morphine by repeated administration: Different mechanisms. Physiol. Psychol. 1983, 11, 155–158. [Google Scholar] [CrossRef] [Green Version]
- DeBeun, R.; Rijk, H.W.; Broekkamp, C.L.E. Cross-familiarisation conditioned taste aversion procedure as a method to reveal stimulus resemblance between drugs: Studies on the 5-HT1A agonist 8-OHDPAT. Psychopharmacologia 1993, 112, 121–128. [Google Scholar] [CrossRef]
- DeBeun, R.; Peeters, B.W.; Broekkamp, C.L. Stimulus characterization of estradiol applying a cross-familiarization taste aversion procedure in female mice. Physiol. Behav. 1993, 53, 715–719. [Google Scholar] [CrossRef] [PubMed]
- DeBeun, R.; Lohmann, A.; Schneider, R.; De Vry, J. Ethanol intake-reducing effects of ipsapirone in rats are not due to simple stimulus substitution. Pharmacol. Biochem. Behav. 1996, 53, 891–898. [Google Scholar] [CrossRef]
- Serafine, K.M.; Riley, A.L. Preexposure to cocaine attenuates aversions induced by both cocaine and fluoxetine: Implications for the basis of cocaine-induced conditioned taste aversions. Pharmacol. Biochem. Behav. 2010, 95, 230–234. [Google Scholar] [CrossRef] [PubMed]
- De Brugada, I.; Hall, G.; Symonds, M. The US-preexposure effect in lithium-induced flavor-aversion conditioning is a consequence of blocking by injection cues. J. Exp. Psychol. Anim. Behav. Process. 2004, 30, 58–66. [Google Scholar] [CrossRef] [Green Version]
- De Brugada, I.; González, F.; Gil, M.; Hall, G. The role of habituation of the response to LiCl in the US-preexposure effect. Learn. Behav. 2005, 33, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Manke, H.N.; Nelson, K.H.; Huang, S.; Bailey, J.M.; Bowman, S.K.; Jones, R.A.; Cerveny, S.E.; Rice, K.C.; Riley, A.L. Methylone pre-exposure differentially impacts the aversive effects of MDPV and MDMA in male and female Sprague-Dawley rats: Implications for abuse vulnerability. Pharmacol. Biochem. Behav. 2022, 220, 173470. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.H.; Manke, H.N.; Bailey, J.M.; Vlachos, A.; Maradiaga, K.J.; Huang, S.; Weiss, T.D.; Rice, K.C.; Riley, A.L. Ethanol pre-exposure differentially impacts the rewarding and aversive effects of α-pyrrolidinopentiophenone (α-PVP): Implications for drug use and abuse. Pharmacol. Biochem. Behav. 2021, 211, 173286. [Google Scholar] [CrossRef]
- Woloshchuk, C.J.; Nelson, K.H.; Rice, K.C.; Riley, A.L. Effects of 3, 4-methylenedioxypyrovalerone (MDPV) pre-exposure on the aversive effects of MDPV, cocaine and lithium chloride: Implications for abuse vulnerability. Drug Alcohol Depend. 2016, 167, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Glennon, R.A.; Young, R. Neurobiology of 3, 4-methylenedioxypyrovalerone (MDPV) and α-pyrrolidinovalerophenone (α-PVP). Brain Res. Bull. 2016, 126, 111–126. [Google Scholar] [PubMed] [Green Version]
- Simmler, L.D.; Buser, T.A.; Donzelli, M.; Schramm, Y.; Dieu, L.H.; Huwyler, J.; Chaboz, S.; Hoener, M.C.; Liechti, M.E. Pharmacology of cathinones. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, S.M.; Grady, M.M.; Muntner, N. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications; Cambridge University Press & Assessment: Cambridge, UK, 2021. [Google Scholar]
- Ervin, G.N.; Birkemo, L.S.; Johnson, M.F.; Conger, L.K.; Mosher, J.T.; Menius, J.A. The effects of anorectic and aversive agents on deprivation-induced feeding and taste aversion conditioning in rats. J. Pharmacol. Exp. Ther. 1995, 273, 1203–1210. [Google Scholar] [PubMed]
- Prendergast, M.A.; Hendricks, S.E.; Yells, D.P.; Balogh, S. Conditioned taste aversion induced by fluoxetine. Physiol. Behav. 1996, 60, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.H.; Broekkamp, C.L. Comparison of stimulus properties of fluoxetine and 5-HT receptor agonists in a conditioned taste aversion procedure. Eur. J. Pharmacol. 1994, 253, 83–89. [Google Scholar] [CrossRef]
- Tancer, M.; Johanson, C.E. The effects of fluoxetine on the subjective and physiological effects of 3, 4-methylenedioxymethamphetamine (MDMA) in humans. Psychopharmacologia 2007, 189, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.R.; Li, K.M.; Clemens, K.J.; Gurtman, C.G.; Hunt, G.E.; Cornish, J.L.; McGregor, I.S. Chronic fluoxetine treatment partly attenuates the long-term anxiety and depressive symptoms induced by MDMA (‘ecstasy’) in rats. Neuropsychopharmacology 2004, 29, 694–704. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.K.; Huang, S.; Vasquez, M.E.; Rice, K.C.; Riley, A.L. Impact of fluoxetine preexposure on MDMA-induced taste avoidance in male and female rats. Exp. Clin. Psychopharmacol. 2022; online ahead of print. [Google Scholar]
- Fernández-Guasti, A.; Olivares-Nazario, M.; Reyes, R.; Martínez-Mota, L. Sex and age differences in the antidepressant-like effect of fluoxetine in the forced swim test. Pharmacol. Biochem. Behav. 2017, 152, 81–89. [Google Scholar]
- Kercmar, J.; Majdic, G. Sex-specific behavioral effects of fluoxetine treatment in animal models of depression and anxiety. Slov. Vet. Res. 2014, 51, 189–200. [Google Scholar]
- Becker, J.B.; Koob, G.F. Sex differences in animal models: Focus on addiction. Pharmacol. Rev. 2016, 68, 242–263. [Google Scholar] [CrossRef] [Green Version]
- Riley, A.L.; Hempel, B.J.; Clasen, M.M. Sex as a biological variable: Drug use and abuse. Physiol. Behav. 2018, 187, 79–96. [Google Scholar] [CrossRef] [PubMed]
- National Research Council US. Guide for the Care and Use of Laboratory Animals; National Academy: Washington, DC, USA, 2011. [Google Scholar]
- National Research Council US. Guidelines for the Care and Use of Mammals in Neuroscience and Behavioral Research; National Academy: Washington, DC, USA, 2003. [Google Scholar]
- Manke, H.N.; Nelson, K.H.; Vlachos, A.; Bailey, J.M.; Maradiaga, K.J.; Weiss, T.D.; Rice, K.C.; Riley, A.L. Assessment of aversive effects of methylone in male and female Sprague-Dawley rats: Conditioned taste avoidance, body temperature and activity/stereotypies. Neurotoxicol. Teratol. 2021, 86, 106977. [Google Scholar] [CrossRef] [PubMed]
- Batsell, W.R.; Best, M.R. One bottle too many? Method of testing determines the detection of overshadowing and retention of taste aversions. Anim. Learn. Behav. 1993, 21, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Dragoin, W.; Mccleary, G.E.; McCleary, P. A comparison of two methods of measuring conditioned taste aversions. Behav. Res. Methods Instrum. 1971, 3, 309–310. [Google Scholar] [CrossRef]
- Grote, F.W.; Brown, R.T. Conditioned taste aversions: Two-stimulus tests are more sensitive than one-stimulus tests. Behav. Res. Methods Instrum. 1971, 3, 311–312. [Google Scholar] [CrossRef] [Green Version]
- Chambers, K.C. Progesterone, estradiol, testosterone and dihydrotestosterone: Effects on rate of extinction of a conditioned taste aversion in rats. Physiol. Behav. 1980, 24, 1061–1065. [Google Scholar] [CrossRef]
- Bano, S.; Akhter, S.; Afridi, M.I. Gender based response to fluoxetine hydrochloride medication in endogenous depression. J. Coll. Physicians Surg. Pak. 2004, 14, 161–165. [Google Scholar]
- Hodes, G.E.; Hill-Smith, T.E.; Suckow, R.F.; Cooper, T.B.; Lucki, I. Sex-specific effects of chronic fluoxetine treatment on neuroplasticity and pharmacokinetics in mice. J. Pharmacol. Exp. Ther. 2010, 332, 266–273. [Google Scholar] [CrossRef]
- Cailhol, S.; Mormède, P. Conditioned taste aversion and alcohol drinking: Strain and gender differences. J. Stud. Alcohol. 2002, 63, 91–99. [Google Scholar] [CrossRef]
- Sherrill, L.K.; Berthold, C.; Koss, W.A.; Juraska, J.M.; Gulley, J.M. Sex differences in the effects of ethanol pre-exposure during adolescence on ethanol-induced conditioned taste aversion in adult rats. Behav. Brain Res. 2011, 225, 104–109. [Google Scholar] [CrossRef] [Green Version]
- Torres, O.V.; Natividad, L.A.; Tejeda, H.A.; van Weelden, S.A.; O’Dell, L.E. Female rats display dose-dependent differences to the rewarding and aversive effects of nicotine in an age-, hormone-, and sex-dependent manner. Psychopharmacologia 2009, 206, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, G.J.; Vail, B.J. Preexposure to delta-9-THC blocks THC-induced conditioned taste aversion in rats. Behav. Neural Biol. 1980, 30, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Cappell, H.; LeBlanc, A.E.; Endrenyi, L. Aversive conditioning by psychoactive drugs: Effects of morphine, alcohol and chlordiazepoxide. Psychopharmacologia 1973, 29, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Serafine, K.M.; Riley, A.L. Cocaine-induced conditioned taste aversions: Role of monoamine reuptake inhibition. In Serotonin: Biosynthesis, Regulation and Health Implications; Hall, F.S., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2013; pp. 257–291. [Google Scholar]
- Goudie, A.J.; Leathley, M.; McNally, J.; West, C.R. Individual differences in cocaine discrimination. Drug Dev. Res. 1989, 16, 123–131. [Google Scholar] [CrossRef]
- Marona-Lewicka, D.; Nichols, D.E. Drug discrimination studies of the interoceptive cues produced by selective serotonin uptake inhibitors and selective serotonin releasing agents. Psychopharmacologia 1998, 138, 67–75. [Google Scholar] [CrossRef]
- Prus, A.J.; Porter, J.H. The discriminative stimulus properties of drugs used to treat depression and anxiety. In The Behavioral Neuroscience of Drug Discrimination; Porter, J., Prus, A., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 213–241. [Google Scholar]
- Webster, J.I.; Harper, D.N.; Schenk, S. Generalization of serotonin and dopamine ligands to the discriminative stimulus effects of different doses of ±3, 4-methylenedioxymethamphetamine. Behav. Pharmacol. 2017, 28, 245–254. [Google Scholar] [CrossRef]
- Wolff, M.C.; Leander, J.D. The discriminative stimulus properties of LY233708, a selective serotonin reuptake inhibitor, in the pigeon. Psychopharmacologia 1999, 146, 275–279. [Google Scholar] [CrossRef]
- Smith, R.L.; Gresch, P.J.; Barrett, R.J.; Sanders-Bush, E. Stimulus generalization by fenfluramine in a quipazine–ketanserin drug discrimination is not dependent on indirect serotonin release. Pharmacol. Biochem. Behav. 2002, 72, 77–85. [Google Scholar] [CrossRef]
- Cunningham, K.A.; Callahan, P.M. Monoamine reuptake inhibitors enhance the discriminative state induced by cocaine in the rat. Psychopharmacologia 1991, 104, 177–180. [Google Scholar] [CrossRef]
- Simon, B.; Appel, J.B. Dopaminergic and serotonergic properties of fluoxetine. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1997, 21, 169–181. [Google Scholar] [CrossRef]
- Schama, K.F.; Howell, L.L.; Byrd, L.D. Serotonergic modulation of the discriminative-stimulus effects of cocaine in squirrel monkeys. Psychopharmacologia 1997, 132, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.E.; Riddle, E.E.; Saunders, R.B.; Appel, J.B. The role of monoamine uptake in the discriminative stimulus effects of cocaine and related compounds. Behav. Pharmacol. 1993, 4, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Barrett, R.L.; Appel, J.B. Effects of stimulation and blockade of dopamine receptor subtypes on the discriminative stimulus properties of cocaine. Psychopharmacologia 1989, 99, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Callahan, P.M.; Appel, J.B.; Cunningham, K.A. Dopamine D1 and D2 mediation of the discriminative stimulus properties of d-amphetamine and cocaine. Psychopharmacologia 1991, 103, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Loney, G.C.; Meyer, P.J. Nicotine pre-treatment reduces sensitivity to the interoceptive stimulus effects of commonly abused drugs as assessed with taste conditioning paradigms. Drug Alcohol Depend. 2019, 194, 341–350. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, M.L.; Escarabajal, M.D.; Agüero, Á. Sex differences in adult Wistar rats in the voluntary consumption of ethanol after pre-exposure to ethanol-induced flavor avoidance learning. Pharmacol. Biochem. Behav. 2015, 137, 7–15. [Google Scholar] [CrossRef]
- Lin, J.Y.; Arthurs, J.; Reilly, S. Conditioned taste aversions: From poisons to pain to drugs of abuse. Psychon. Bull. Rev. 2017, 24, 335–351. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Genetic influences on conditioned taste aversion. In Conditioned Taste Aversion: Behavioral and Neural Processes; Reilly, S., Schachtman, T.R., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 387–421. [Google Scholar]
- Fenu, S.; Bassareo, V.; di Chiara, G. A role for dopamine D1 receptors of the nucleus accumbens shell in conditioned taste aversion learning. J. Neurosci. 2001, 21, 6897–6904. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.G.; Alva, H.; Blednov, Y.A.; Cunningham, C.L. Reduced ethanol-induced conditioned taste aversion and conditioned place preference in GIRK2 null mutant mice. Psychopharmacologia 2003, 169, 108–114. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manke, H.N.; Rice, K.C.; Riley, A.L. Effects of Methylone Pre-Exposure on Fluoxetine-Induced Conditioned Taste Avoidance in Male and Female Sprague-Dawley Rats. Brain Sci. 2023, 13, 585. https://doi.org/10.3390/brainsci13040585
Manke HN, Rice KC, Riley AL. Effects of Methylone Pre-Exposure on Fluoxetine-Induced Conditioned Taste Avoidance in Male and Female Sprague-Dawley Rats. Brain Sciences. 2023; 13(4):585. https://doi.org/10.3390/brainsci13040585
Chicago/Turabian StyleManke, Hayley N., Kenner C. Rice, and Anthony L. Riley. 2023. "Effects of Methylone Pre-Exposure on Fluoxetine-Induced Conditioned Taste Avoidance in Male and Female Sprague-Dawley Rats" Brain Sciences 13, no. 4: 585. https://doi.org/10.3390/brainsci13040585