
Neural Processing of Odors with Different Well-Being Associations—Findings from Two Consecutive Neuroimaging Studies
 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Study Design and Participants
2.2. Study 1
2.3. Study 2
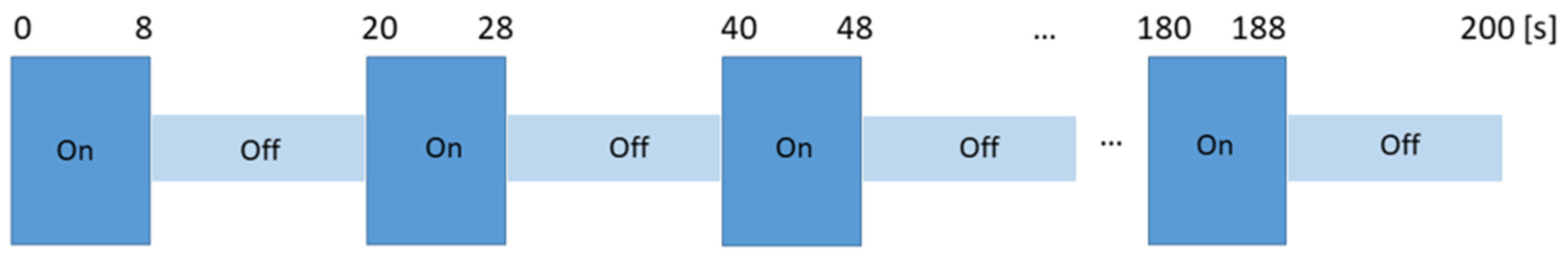
2.4. Odor Presentation in the Scanner
2.5. fMRI Acquisition
2.6. MRI Data Preprocessing and GLM Analysis
2.7. Statistical Analysis
3. Results
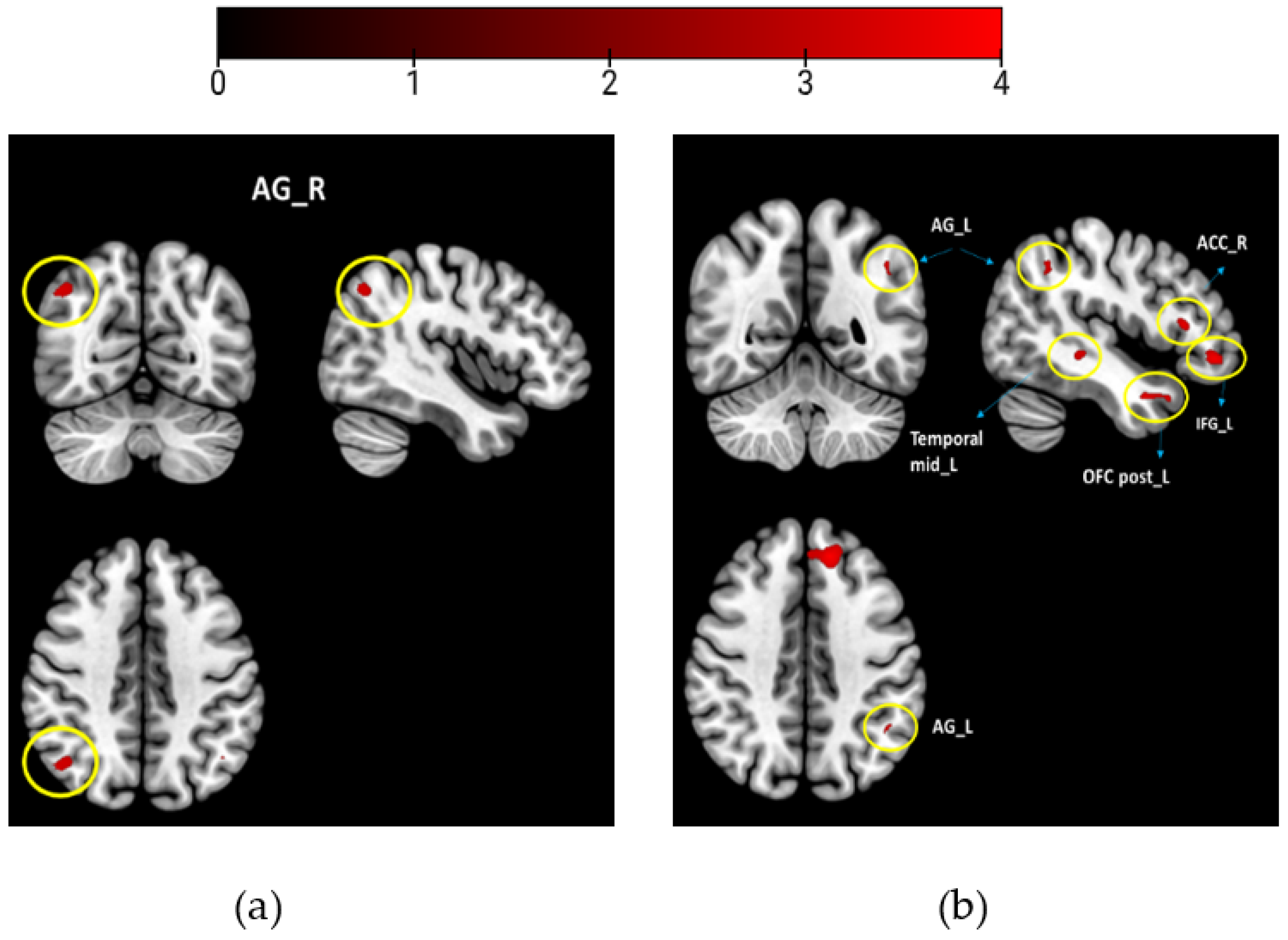
fMRI Results
- (1)
- WB-associated odors > neutral odors
- (2)
- WB-associated odors < neutral odors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conversano, C.; Rotondo, A.; Lensi, E.; Della Vista, O.; Arpone, F.; Reda, M.A. Optimism and Its Impact on Mental and Physical Well-Being. Clin. Pract. Epidemiol. Ment. Health 2010, 6, 25–29. [Google Scholar] [CrossRef]
- Kühn, S.; Rieger, U.M. Health is a state of complete physical, mental and social well-being and not merely absence of disease or infirmity. Surg. Obes. Relat. Dis. 2017, 13, 887. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.D.; Drummond, P.D. Olfaction in migraine. Cephalalgia 1997, 17, 729–732. [Google Scholar] [CrossRef] [PubMed]
- Villemure, C.; Slotnick, B.M.; Bushnell, M.C. Effects of odors on pain perception: Deciphering the roles of emotion and attention. Pain 2003, 106, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.; Hewitt, S.; Moss, L.; Wesnes, K. Modulation of cognitive performance and mood by aromas of peppermint and ylang-ylang. Int. J. Neurosci. 2008, 118, 59–77. [Google Scholar] [CrossRef]
- Schifferstein, H.N.J.; Smeets, M.A.M.; Hallensleben, R. Stimulus sets can induce shifts in descriptor meanings in product evaluation tasks. Acta Psychol. 2011, 138, 237–243. [Google Scholar] [CrossRef]
- Heuberger, E.; Hongratanaworakit, T.; Buchbauer, G. East Indian Sandalwood and alpha-santalol odor increase physiological and self-rated arousal in humans. Planta Med. 2006, 72, 792–800. [Google Scholar] [CrossRef]
- Gottfried, J.A.; Deichmann, R.; Winston, J.S.; Dolan, R.J. Functional heterogeneity in human olfactory cortex: An event-related functional magnetic resonance imaging study. J. Neurosci. 2002, 22, 10819–10828. [Google Scholar] [CrossRef]
- Gottfried, J.A.; O’Doherty, J.; Dolan, R.J. Appetitive and aversive olfactory learning in humans studied using event-related functional magnetic resonance imaging. J. Neurosci. 2002, 22, 10829–10837. [Google Scholar] [CrossRef]
- Chen, B.; Benzien, C.; Faria, V.; Ning, Y.; Cuevas, M.; Linke, J.; Croy, I.; Haehner, A.; Hummel, T. Symptoms of Depression in Patients with Chemosensory Disorders. ORL J. Otorhinolaryngol. Relat. Spec. 2021, 83, 135–143. [Google Scholar] [CrossRef]
- Croy, I.; Nordin, S.; Hummel, T. Olfactory disorders and quality of life—An updated review. Chem. Senses 2014, 39, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.; Menzel, S.; Cuevas, M.; Haehner, A.; Hummel, T. Well-being in patients with olfactory dysfunction. Physiol. Behav. 2022, 254, 113899. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, K.; Hudson, J.; Haviland-Jones, J.; Wilson, P. Does exposure to ambient odors influence the emotional content of memories? Am. J. Psychol. 2010, 123, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Pennebaker, J.W. The Psychology of Physical Symptoms; Springer: New York, NY, USA, 1982; ISBN 978-1-4613-8198-3. [Google Scholar]
- Paulus, M.P.; Stein, M.B. An Insular View of Anxiety. Biol. Psychiatry 2006, 60, 383–387. [Google Scholar] [CrossRef]
- Soudry, Y.; Lemogne, C.; Malinvaud, D.; Consoli, S.-M.; Bonfils, P. Olfactory system and emotion: Common substrates. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2011, 128, 18–23. [Google Scholar] [CrossRef]
- Hooker, C.I.; Germine, L.T.; Knight, R.T.; D’Esposito, M. Amygdala Response to Facial Expressions Reflects Emotional Learning. J. Neurosci. 2006, 26, 8915–8922. [Google Scholar] [CrossRef]
- O’Doherty, J.; Rolls, E.T.; Francis, S.; Bowtell, R.; McGlone, F.; Kobal, G.; Renner, B.; Ahne, G. Sensory-specific satiety-related olfactory activation of the human orbitofrontal cortex. Neuroreport 2000, 11, 399–403. [Google Scholar] [CrossRef]
- Welge-Luessen, A.; Leopold, D.A.; Miwa, T. Smell and taste disorders-diagnostic and clinical work-up. In Management of Smell and Taste Disorders—A Practical Guide for Clinicians; Thieme: Stuttgart, Germany, 2013; pp. 49–57. [Google Scholar]
- Hummel, T.; Sekinger, B.; Wolf, S.R.; Pauli, E.; Kobal, G. “Sniffin” sticks’: Olfactory performance assessed by the combined testing of odor identification, odor discrimination and olfactory threshold. Chem. Senses 1997, 22, 39–52. [Google Scholar] [CrossRef]
- Wydra, G. FAHW: Fragebogen zum Allgemeinen Habituellen Wohlbefinden. Germany, 2003. Available online: https://www.sportpaedagogik-sb.de/pdf/FAHW%202020.pdf (accessed on 15 February 2021).
- Topp, C.W.; Østergaard, S.D.; Søndergaard, S.; Bech, P. The WHO-5 Well-Being Index: A Systematic Review of the Literature. Psychother. Psychosom. 2015, 84, 167–176. [Google Scholar] [CrossRef]
- Lyubomirsky, S.; Lepper, H.S. A measure of subjective happiness: Preliminary reliability and construct validation. Soc. Indic. Res. 1999, 46, 137–155. [Google Scholar] [CrossRef]
- Bartram, D.J.; Yadegarfar, G.; Sinclair, J.M.A.; Baldwin, D.S. Validation of the Warwick–Edinburgh Mental Well-being Scale (WEMWBS) as an overall indicator of population mental health and well-being in the UK veterinary profession. Vet. J. 2011, 187, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Bradley, C. The 12-Item Well-Being Questionnaire: Origins, current stage of development, and availability. Diabetes Care 2000, 23, 875. [Google Scholar] [CrossRef] [PubMed]
- Sommer, J.U.; Maboshe, W.; Griebe, M.; Heiser, C.; Hörmann, K.; Stuck, B.A.; Hummel, T. A mobile olfactometer for fMRI-studies. J. Neurosci. Methods 2012, 209, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Penny, W.D.; Friston, K.J.; Ashburner, J.T.; Kiebel, S.J.; Nichols, T.E. Statistical Parametric Mapping: The Analysis of Functional Brain Images; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Geschwind, N. Disconnexion syndromes in animals and man. Brain 1965, 88, 237. [Google Scholar] [CrossRef] [PubMed]
- Seghier, M.L. The angular gyrus: Multiple functions and multiple subdivisions. Neuroscientist 2013, 19, 43–61. [Google Scholar] [CrossRef]
- Singh-Curry, V.; Husain, M. The functional role of the inferior parietal lobe in the dorsal and ventral stream dichotomy. Neuropsychologia 2009, 47, 1434–1448. [Google Scholar] [CrossRef]
- Carlson, H.; Leitão, J.; Delplanque, S.; Cayeux, I.; Sander, D.; Vuilleumier, P. Sustained effects of pleasant and unpleasant smells on resting state brain activity. Cortex 2020, 132, 386–403. [Google Scholar] [CrossRef]
- Kohn, N.; Eickhoff, S.B.; Scheller, M.; Laird, A.R.; Fox, P.T.; Habel, U. Neural network of cognitive emotion regulation—An ALE meta-analysis and MACM analysis. Neuroimage 2014, 87, 345–355. [Google Scholar] [CrossRef]
- Binder, J.R.; Desai, R.H. The neurobiology of semantic memory. Trends Cogn. Sci. 2011, 15, 527–536. [Google Scholar] [CrossRef]
- Royet, J.-P.; Koenig, O.; Gregoire, M.-C.; Cinotti, L.; Lavenne, F.; Bars, D.L.; Costes, N.; Vigouroux, M.; Farget, V.; Sicard, G.; et al. Functional Anatomy of Perceptual and Semantic Processing for Odors. J. Cogn. Neurosci. 1999, 11, 94–109. [Google Scholar] [CrossRef]
- Olson, I.R.; McCoy, D.; Klobusicky, E.; Ross, L.A. Social cognition and the anterior temporal lobes: A review and theoretical framework. Soc. Cogn. Affect. Neurosci. 2013, 8, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sasabe, T.; Kobayashi, M.; Kondo, Y.; Onoe, H.; Matsubara, S.; Yamamoto, S.; Tsukada, H.; Onoe, K.; Watabe, H.; Iida, H.; et al. Activation of the Anterior Cingulate Gyrus by ‘Green Odor’: A Positron Emission Tomography Study in the Monkey. Chem. Senses 2003, 28, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Bechara, A.; Damasio, H.; Damasio, A.R. Emotion, Decision Making and the Orbitofrontal Cortex. Cereb. Cortex 2000, 10, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, J.; Hummel, C.; Bojanowski, V.; Warr, J.; Gerber, J.; Hummel, T. Food-Related Odors and the Reward Circuit: Functional MRI. Chemosens. Percept. 2015, 4, 192–200. [Google Scholar] [CrossRef]
- Sorokowska, A.; Schoen, K.; Hummel, C.; Han, P.; Warr, J.; Hummel, T. Food-Related Odors Activate Dopaminergic Brain Areas. Front. Hum. Neurosci. 2017, 11, 625. [Google Scholar] [CrossRef]
- Lieberman, M.D.; Cunningham, W.A. Type I and Type II error concerns in fMRI research: Re-balancing the scale. Soc. Cogn. Affect. Neurosci. 2009, 4, 423–428. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ratings (Range of Scale) | WB-Associated Odors | Neutral Odors | |
|---|---|---|---|
| study 1 | musk flower + orange | coffee + grass | p-value |
| Intensity (0 to 10) | 6.79 ± 1.98 | 6.62 ± 2.07 | n.s. |
| Valence (−5 to +5) | 3.00 ± 1.72 | −0.42 ± 2.49 | <0.001 |
| Well-being (0 to 10) | 7.25 ± 1.78 | 4.63 ± 2.15 | <0.001 |
| study 2 | vanilla + white soap | sheets + grass | |
| Intensity (0 to 10) | 6.59 ± 1.84 | 7.12 ± 1.87 | n.s. |
| Valence (−5 to +5) | 3.21 ± 1.85 | −0.18 ± 2.69 | <0.001 |
| Well-being (0 to 10) | 6.72 ± 2.11 | 3.4 ± 2.13 | <0.001 |
| Study 1 | k | T Value | x y z | Region |
|---|---|---|---|---|
| WB-associated odors > neutral odors | 23 | 3.40 | 46 −68 44 | Angular gyrus R |
| WB-associated odors < neutral odors | - | - | - | - |
| study 2 | ||||
| WB-associated odors > neutral odors | 111 | 4.51 | −52 6 −26 | Mid temporal L |
| 303 | 4.35 | −10 50 24 | Frontal superior medial L | |
| 194 | 4.27 | −14 38 40 | Frontal superior L | |
| 45 | 4.11 | −8 24 −24 | Gyrus Rectus L | |
| 141 | 4.07 | −42 42 −8 | Inferior frontal gyrus L | |
| 22 | 3.94 | 60 −4 −26 | ||
| 80 | 3.90 | −52 24 10 | Inferior frontal gyrus L | |
| 76 | 3.78 | −60 −16 −16 | Mid temporal L | |
| 58 | 3.66 | 14 38 4 | Anterior cingulate R | |
| 32 | 3.65 | −34 30 −18 | Posterior OFC L | |
| 13 | 3.57 | −46 −54 44 | Angular gyrus L | |
| WB-associated odors < neutral odors | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, A.; Hornstein, H.; Thaploo, D.; Faria, V.; Warr, J.; Hummel, T. Neural Processing of Odors with Different Well-Being Associations—Findings from Two Consecutive Neuroimaging Studies. Brain Sci. 2023, 13, 576. https://doi.org/10.3390/brainsci13040576
Joshi A, Hornstein H, Thaploo D, Faria V, Warr J, Hummel T. Neural Processing of Odors with Different Well-Being Associations—Findings from Two Consecutive Neuroimaging Studies. Brain Sciences. 2023; 13(4):576. https://doi.org/10.3390/brainsci13040576
Chicago/Turabian StyleJoshi, Akshita, Henriette Hornstein, Divesh Thaploo, Vanda Faria, Jonathan Warr, and Thomas Hummel. 2023. "Neural Processing of Odors with Different Well-Being Associations—Findings from Two Consecutive Neuroimaging Studies" Brain Sciences 13, no. 4: 576. https://doi.org/10.3390/brainsci13040576