
EEG Dynamics of Error Processing and Associated Behavioral Adjustments in Preschool Children
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Sociodemographic Scale
2.3. Task
2.4. Electrophysiological Recording and Processing
2.4.1. Event-Related Potentials (ERPs)
2.4.2. Event-Related Spectral Perturbations (ERSPs)
2.5. Data Analysis
2.5.1. Behavior
2.5.2. Neural Activity
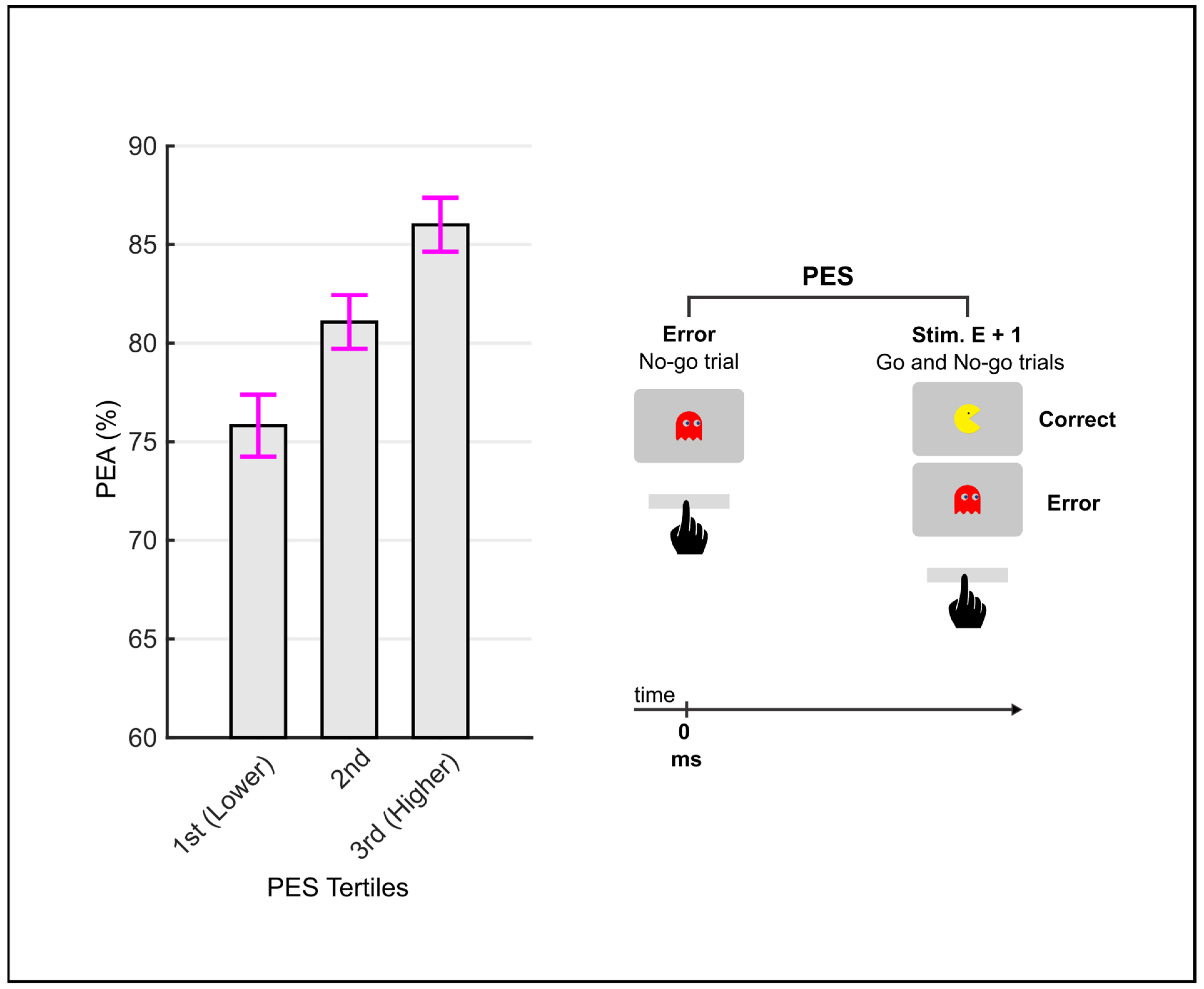
2.5.3. Response–Stimulus Interval Analysis
3. Results
3.1. Sociodemographic Characteristics of the Sample
3.2. Behavior
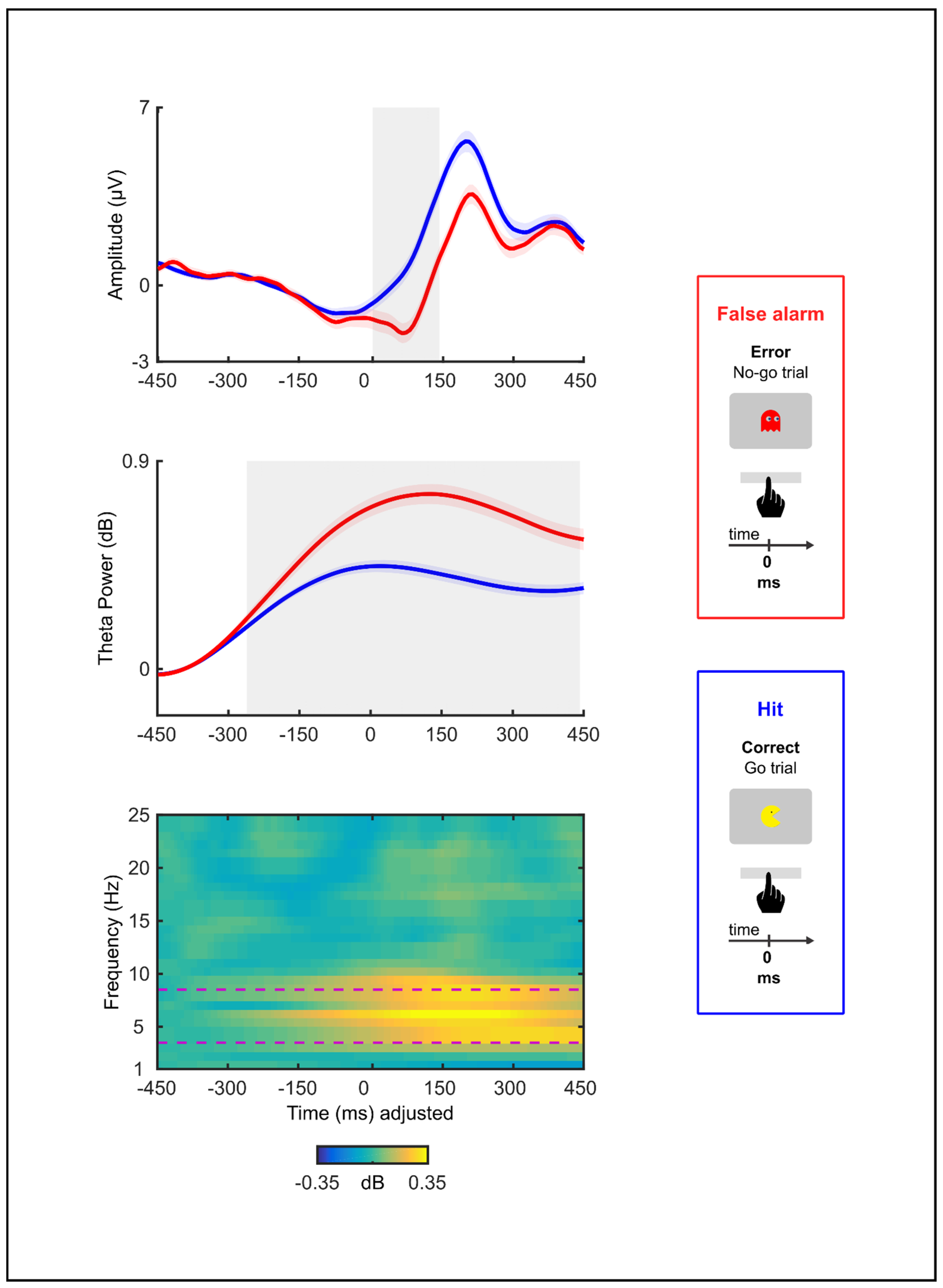
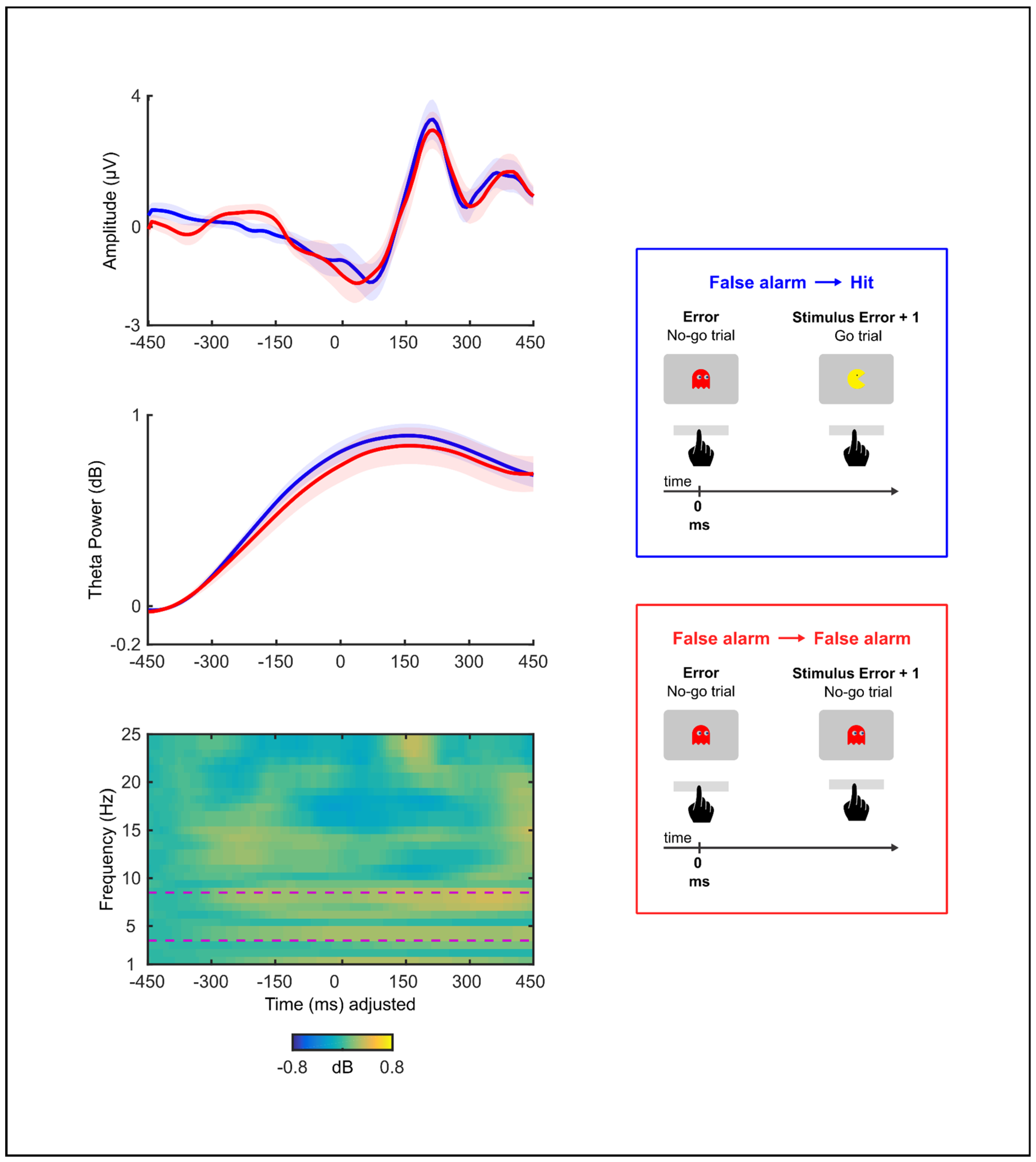
3.3. Electrophysiology
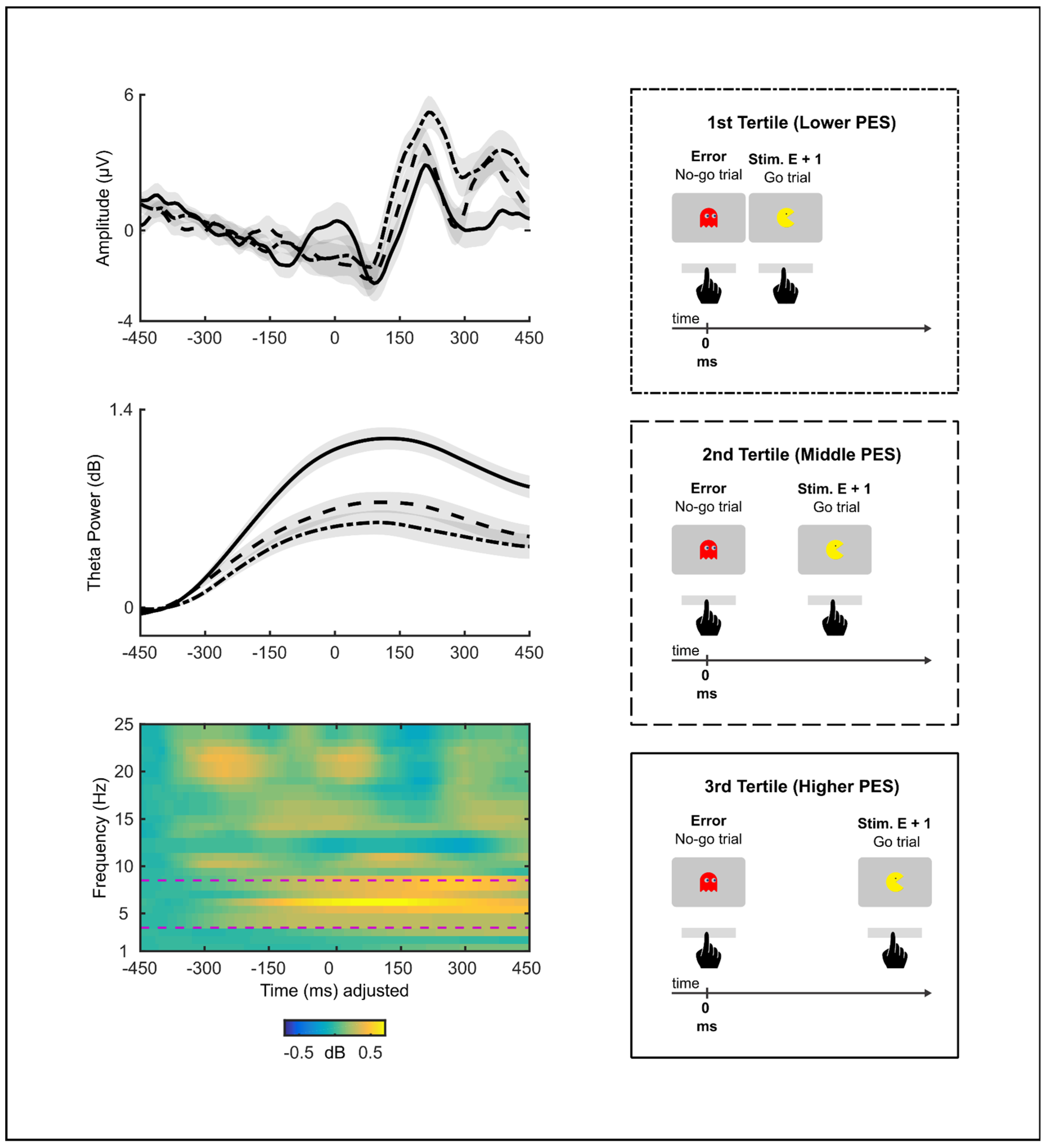
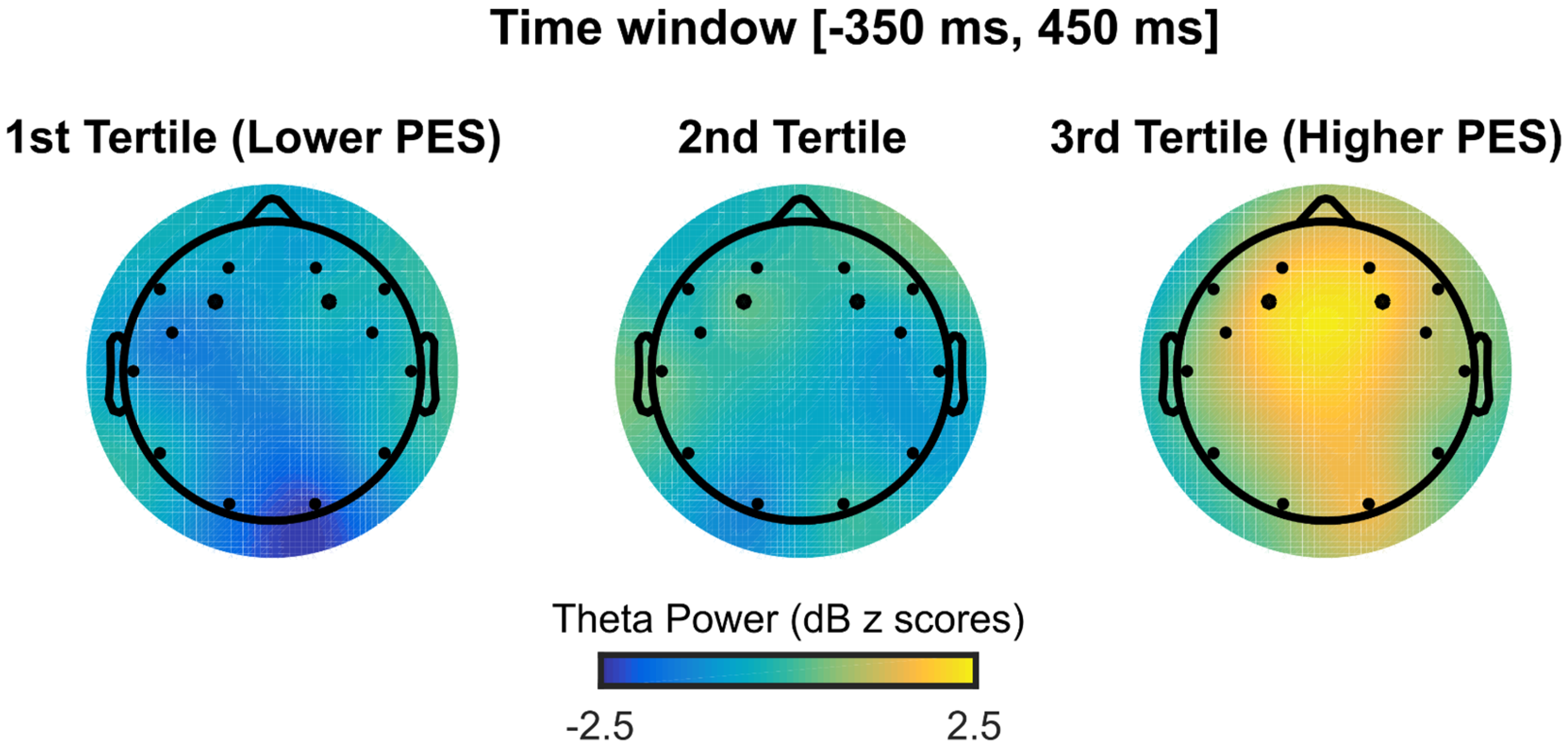
3.4. RSI
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botvinick, M.M.; Braver, T.S.; Barch, D.M.; Carter, C.S.; Cohen, J.D. Conflict Monitoring and Cognitive Control. Psychol. Rev. 2001, 108, 624–652. [Google Scholar] [CrossRef]
- Ullsperger, M.; Fischer, A.G.; Nigbur, R.; Endrass, T. Neural Mechanisms and Temporal Dynamics of Performance Monitoring. Trends Cogn. Sci. 2014, 18, 259–267. [Google Scholar] [CrossRef]
- Ullsperger, M.; Danielmeier, C.; Jocham, G. Neurophysiology of Performance Monitoring and Adaptive Behavior. Physiol. Rev. 2014, 94, 35–79. [Google Scholar] [CrossRef] [Green Version]
- Shenhav, A.; Botvinick, M.M.; Cohen, J.D. The Expected Value of Control: An Integrative Theory of Anterior Cingulate Cortex Function. Neuron 2013, 79, 217–240. [Google Scholar] [CrossRef] [Green Version]
- Wessel, J.R. An Adaptive Orienting Theory of Error Processing. Psychophysiology 2018, 55, e13041. [Google Scholar] [CrossRef] [Green Version]
- Boen, R.; Quintana, D.S.; Ladouceur, C.D.; Tamnes, C.K. Age-related Differences in the Error-related Negativity and Error Positivity in Children and Adolescents Are Moderated by Sample and Methodological Characteristics: A Meta-analysis. Psychophysiology 2022, 59, e14003. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Frank, M.J. Frontal Theta as a Mechanism for Cognitive Control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.L. A Meta-Analytic Review of the Event-Related Potentials (ERN and N2) in Childhood and Adolescence: Providing a Developmental Perspective on the Conflict Monitoring Theory. Dev. Rev. 2018, 48, 82–112. [Google Scholar] [CrossRef]
- Danielmeier, C.; Ullsperger, M. Post-Error Adjustments. Front. Psychol. 2011, 2, 233. [Google Scholar] [CrossRef] [Green Version]
- Beatty, P.J.; Buzzell, G.A.; Roberts, D.M.; McDonald, C.G. Contrasting Time and Frequency Domains: ERN and Induced Theta Oscillations Differentially Predict Post-Error Behavior. Cogn. Affect. Behav. Neurosci. 2020, 20, 636–647. [Google Scholar] [CrossRef]
- Marco-Pallarés, J.; Camara, E.; Münte, T.F.; Rodríguez-Fornells, A. Neural Mechanisms Underlying Adaptive Actions after Slips. J. Cogn. Neurosci. 2008, 20, 1595–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadez, E.A.; Simons, R.F. The Power of Frontal Midline Theta and Post-Error Slowing to Predict Performance Recovery: Evidence for Compensatory Mechanisms. Psychophysiology 2018, 55, e13010. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.J.; Buzzell, G.A.; Roberts, D.M.; McDonald, C.G. Speeded Response Errors and the Error-Related Negativity Modulate Early Sensory Processing. NeuroImage 2018, 183, 112–120. [Google Scholar] [CrossRef]
- Jentzsch, I.; Dudschig, C. Short Article: Why Do We Slow down after an Error? Mechanisms Underlying the Effects of Posterror Slowing. Q. J. Exp. Psychol. 2009, 62, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ullsperger, M.; Danielmeier, C. Reducing Speed and Sight: How Adaptive Is Post-Error Slowing? Neuron 2016, 89, 430–432. [Google Scholar] [CrossRef] [Green Version]
- van den Brink, R.L.; Wynn, S.C.; Nieuwenhuis, S. Post-Error Slowing as a Consequence of Disturbed Low-Frequency Oscillatory Phase Entrainment. J. Neurosci. 2014, 34, 11096–11105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutilh, G.; Vandekerckhove, J.; Forstmann, B.U.; Keuleers, E.; Brysbaert, M.; Wagenmakers, E.-J. Testing Theories of Post-Error Slowing. Atten. Percept. Psychophys. 2012, 74, 454–465. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.A.C. Information Theory of Choice-Reaction Times. By D. R. J. Laming. Br. J. Math. Stat. Psychol. 1969, 22, 103–104. [Google Scholar] [CrossRef]
- Ridderinkhof, R. Micro- and Macro-Adjustments of Task Set: Activation and Suppression in Conflict Tasks. Psychol. Res. 2002, 66, 312–323. [Google Scholar] [CrossRef]
- Notebaert, W.; Houtman, F.; Opstal, F.V.; Gevers, W.; Fias, W.; Verguts, T. Post-Error Slowing: An Orienting Account. Cognition 2009, 111, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Purcell, B.A.; Kiani, R. Neural Mechanisms of Post-Error Adjustments of Decision Policy in Parietal Cortex. Neuron 2016, 89, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannaccone, R.; Hauser, T.U.; Staempfli, P.; Walitza, S.; Brandeis, D.; Brem, S. Conflict Monitoring and Error Processing: New Insights from Simultaneous EEG–FMRI. NeuroImage 2015, 105, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Luu, P.; Tucker, D.M.; Makeig, S. Frontal Midline Theta and the Error-Related Negativity: Neurophysiological Mechanisms of Action Regulation. Clin. Neurophysiol. 2004, 115, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Nigbur, R.; Ivanova, G.; Stürmer, B. Theta Power as a Marker for Cognitive Interference. Clin. Neurophysiol. 2011, 122, 2185–2194. [Google Scholar] [CrossRef]
- van Veen, V.; Carter, C.S. The Timing of Action-Monitoring Processes in the Anterior Cingulate Cortex. J. Cogn. Neurosci. 2002, 14, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Yeung, N.; Botvinick, M.M.; Cohen, J.D. The Neural Basis of Error Detection: Conflict Monitoring and the Error-Related Negativity. Psychol. Rev. 2004, 111, 931–959. [Google Scholar] [CrossRef]
- Falkenstein, M.; Hohnsbein, J.; Hoormann, J.; Blanke, L. Effects of Crossmodal Divided Attention on Late ERP Components. II. Error Processing in Choice Reaction Tasks. Electroencephalogr. Clin. Neurophysiol. 1991, 78, 447–455. [Google Scholar] [CrossRef]
- Gehring, W.J.; Goss, B.; Coles, M.G.H.; Meyer, D.E.; Donchin, E. A Neural System for Error Detection and Compensation. Psychol. Sci. 1993, 4, 385–390. [Google Scholar] [CrossRef]
- Wu, T.; Spagna, A.; Chen, C.; Schulz, K.P.; Hof, P.R.; Fan, J. Supramodal Mechanisms of the Cognitive Control Network in Uncertainty Processing. Cereb. Cortex 2020, 30, 6336–6349. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Shackman, A.J. Frontal Midline Theta Reflects Anxiety and Cognitive Control: Meta-Analytic Evidence. J. Physiol.-Paris 2015, 109, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Garon, N.; Bryson, S.E.; Smith, I.M. Executive Function in Preschoolers: A Review Using an Integrative Framework. Psychol. Bull. 2008, 134, 31–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, M.R.; Conejero, A. Developing Attention and Self-Regulation in Infancy and Childhood. In Neural Circuit and Cognitive Development; Elsevier: Amsterdam, The Netherlands, 2020; pp. 505–522. ISBN 978-0-12-814411-4. [Google Scholar]
- Chevalier, N.; Hadley, L.V.; Balthrop, K. Midfrontal Theta Oscillations and Conflict Monitoring in Children and Adults. Dev. Psychobiol. 2021, 63, e22216. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Alexander, N.; Wessel, J.R.; Wimberger, P.; Nitzsche, K.; Kirschbaum, C.; Li, S.-C. Neurocognitive Development of Novelty and Error Monitoring in Children and Adolescents. Sci. Rep. 2021, 11, 19844. [Google Scholar] [CrossRef]
- Berwid, O.G.; Halperin, J.M.; Johnson, R.; Marks, D.J. Preliminary Evidence for Reduced Posterror Reaction Time Slowing in Hyperactive/Inattentive Preschool Children. Child Neuropsychol. 2014, 20, 196–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.B.; Rothbart, M.K.; Posner, M.I. Development of Executive Attention in Preschool Children. Dev. Sci. 2003, 6, 498–504. [Google Scholar] [CrossRef]
- Smulders, S.F.A.; Soetens, E.; van der Molen, M.W. What Happens When Children Encounter an Error? Brain Cogn. 2016, 104, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Torpey, D.C.; Hajcak, G.; Kim, J.; Kujawa, A.; Klein, D.N. Electrocortical and Behavioral Measures of Response Monitoring in Young Children during a Go/No-Go Task. Dev. Psychobiol. 2012, 54, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Canen, M.J.; Brooker, R.J. ERN, Theta Power, and Risk for Anxiety Problems in Preschoolers. Biol. Psychol. 2017, 123, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Conejero, Á.; Guerra, S.; Abundis-Gutiérrez, A.; Rueda, M.R. Frontal Theta Activation Associated with Error Detection in Toddlers: Influence of Familial Socioeconomic Status. Dev. Sci. 2018, 21, e12494. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Buzzell, G.; Faja, S.; Choi, Y.B.; Thomas, H.R.; Brito, N.H.; Shuffrey, L.C.; Fifer, W.P.; Morrison, F.D.; Lord, C.; et al. Neural Dynamics of Executive Function in Cognitively Able Kindergarteners with Autism Spectrum Disorders as Predictors of Concurrent Academic Achievement. Autism 2020, 24, 780–794. [Google Scholar] [CrossRef]
- Overbye, K.; Walhovd, K.B.; Paus, T.; Fjell, A.M.; Huster, R.J.; Tamnes, C.K. Error Processing in the Adolescent Brain: Age-Related Differences in Electrophysiology, Behavioral Adaptation, and Brain Morphology. Dev. Cogn. Neurosci. 2019, 38, 100665. [Google Scholar] [CrossRef] [PubMed]
- Wiersema, J.R.; van der Meere, J.J.; Roeyers, H. Developmental Changes in Error Monitoring: An Event-Related Potential Study. Neuropsychologia 2007, 45, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Adam, N.; Blaye, A.; Gulbinaite, R.; Delorme, A.; Farrer, C. The Role of Midfrontal Theta Oscillations across the Development of Cognitive Control in Preschoolers and School-age Children. Dev. Sci. 2020, 23, e12936. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Endedijk, H.M.; van Ede, F.; Hunnius, S. Theta Oscillations in 4-Year-Olds Are Sensitive to Task Engagement and Task Demands. Sci. Rep. 2019, 9, 6049. [Google Scholar] [CrossRef] [Green Version]
- Brooker, R.J. Maternal Behavior and Socioeconomic Status Predict Longitudinal Changes in Error-Related Negativity in Preschoolers. Child Dev. 2018, 89, 725–733. [Google Scholar] [CrossRef]
- Mistry-Patel, S.; Brooker, R.J. Associations between Error-Related Negativity and Childhood Anxiety Risk Differ Based on Socioeconomic Status. Dev. Psychol. 2022. [Google Scholar] [CrossRef]
- Pietto, M.L.; Giovannetti, F.; Segretin, M.S.; Rueda, R.; Kamienkowski, J.E.; Lipina, S.J. Conflict-Related Brain Activity after Individualized Cognitive Training in Preschoolers from Poor Homes. J. Cogn. Enhanc. 2022, 6, 74–107. [Google Scholar] [CrossRef]
- DuPuis, D.; Ram, N.; Willner, C.J.; Karalunas, S.; Segalowitz, S.J.; Gatzke-Kopp, L.M. Implications of Ongoing Neural Development for the Measurement of the Error-Related Negativity in Childhood. Dev. Sci. 2015, 18, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Grammer, J.K.; Carrasco, M.; Gehring, W.J.; Morrison, F.J. Age-Related Changes in Error Processing in Young Children: A School-Based Investigation. Dev. Cogn. Neurosci. 2014, 9, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Grammer, J.K.; Gehring, W.J.; Morrison, F.J. Associations between Developmental Changes in Error-Related Brain Activity and Executive Functions in Early Childhood. Psychophysiology 2018, 55, e13040. [Google Scholar] [CrossRef]
- Ip, K.I.; Liu, Y.; Moser, J.; Mannella, K.; Hruschak, J.; Bilek, E.; Muzik, M.; Rosenblum, K.; Fitzgerald, K. Moderation of the Relationship between the Error-Related Negativity and Anxiety by Age and Gender in Young Children: A Preliminary Investigation. Dev. Cogn. Neurosci. 2019, 39, 100702. [Google Scholar] [CrossRef]
- Kessel, E.M.; Meyer, A.; Hajcak, G.; Dougherty, L.R.; Torpey-Newman, D.C.; Carlson, G.A.; Klein, D.N. Transdiagnostic Factors and Pathways to Multifinality: The Error-Related Negativity Predicts Whether Preschool Irritability Is Associated with Internalizing versus Externalizing Symptoms at Age 9. Dev. Psychopathol. 2016, 28, 913–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olvet, D.M.; Hajcak, G. The Stability of Error-Related Brain Activity with Increasing Trials. Psychophysiology 2009, 46, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Abeyá Gilardon, E.; Calvo, E.; Durán, P.; Longo, E.; Mazza, C. Evaluación del Estado Nutricional de Niñas, Niños y Embarazadas Mediante Antropometría. 2009. Available online: https://bancos.salud.gob.ar/recurso/evaluacion-del-estado-nutricional-de-ninas-ninos-y-embarazadas-mediante-antropometria (accessed on 26 March 2023).
- Lipina, S.; Segretin, S.; Hermida, J.; Prats, L.; Fracchia, C.; Camelo, J.L.; Colombo, J. Linking Childhood Poverty and Cognition: Environmental Mediators of Non-Verbal Executive Control in an Argentine Sample. Dev. Sci. 2013, 16, 697–707. [Google Scholar] [CrossRef]
- Pietto, M.L.; Gatti, M.; Raimondo, F.; Lipina, S.J.; Kamienkowski, J.E. Electrophysiological Approaches in the Study of Cognitive Development Outside the Lab. PLoS ONE 2018, 13, e0206983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, M.; Quian Quiroga, R. Automatic Denoising of Single-Trial Evoked Potentials. NeuroImage 2013, 66, 672–680. [Google Scholar] [CrossRef]
- Navajas, J.; Ahmadi, M.; Quian Quiroga, R. Uncovering the Mechanisms of Conscious Face Perception: A Single-Trial Study of the N170 Responses. J. Neurosci. 2013, 33, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Quian Quiroga, R. Obtaining Single Stimulus Evoked Potentials with Wavelet Denoising. Phys. Nonlinear Phenom. 2000, 145, 278–292. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.X. Analyzing Neural Time Series Data: Theory and Practice; Issues in clinical and cognitive neuropsychology; MIT Press: Cambridge, MA, USA, 2014; ISBN 978-0-262-01987-3. [Google Scholar]
- Checa, P.; Castellanos, M.C.; Abundis-Gutiérrez, A.; Rosario Rueda, M. Development of Neural Mechanisms of Conflict and Error Processing during Childhood: Implications for Self-Regulation. Front. Psychol. 2014, 5, 326. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, M.; Tomczak, E. The Need to Report Effect Size Estimates Revisited. An Overview of Some Recommended Measures of Effect Size. Trends Sport Sci. 2014, 1, 19–25. [Google Scholar]
- Brooker, R.J.; Buss, K.A. Toddler Fearfulness Is Linked to Individual Differences in Error-Related Negativity During Preschool. Dev. Neuropsychol. 2014, 39, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairweather, H. Choice Reaction Times in Children: Error and Post-Error Responses, and the Repetition Effect. J. Exp. Child Psychol. 1978, 26, 407–418. [Google Scholar] [CrossRef]
- Trujillo, L.T.; Allen, J.J.B. Theta EEG Dynamics of the Error-Related Negativity. Clin. Neurophysiol. 2007, 118, 645–668. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.R.; Kovacevic, N.; Itier, R.J. Increased Brain Signal Variability Accompanies Lower Behavioral Variability in Development. PLoS Comput. Biol. 2008, 4, e1000106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermida, M.J.; Shalom, D.E.; Segretin, M.S.; Goldin, A.P.; Abril, M.C.; Lipina, S.J.; Sigman, M. Risks for child cognitive development in rural contexts. Front. Psychol. 2019, 9, 2735. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.J.; Segretin, M.S.; Prats, L.M.; Fracchia, C.S.; Colombo, J.A.; Lipina, S.J. Cognitive Neuroscience, Developmental Psychology, and Education: Interdisciplinary Development of an Intervention for Low Socioeconomic Status Kindergarten Children. Trends Neurosci. Educ. 2015, 4, 15–25. [Google Scholar] [CrossRef]
- Segretin, M.S.; Lipina, S.J.; Hermida, M.J.; Sheffield, T.D.; Nelson, J.M.; Espy, K.A.; Colombo, J.A. Predictors of Cognitive Enhancement after Training in Preschoolers from Diverse Socioeconomic Backgrounds. Front. Psychol. 2014, 5, 205. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of UBN Indicator | Proportion of the Sample |
|---|---|
| UBN 1 | 96.4% |
| UBNHEAD 2 | 9.6% |
| UBNEDU 3 | 2.7% |
| UBNHOUSE 4 | 26.2% |
| UBNWATER 5 | 0% |
| UBNBATHR 6 | 26.5% |
| UBNOVERC 7 | 14.3% |
| NEIGHBORHOOD 8 | 95.2% |
| Measure | Median | Q25 | Q75 | n |
|---|---|---|---|---|
| Hits (%) | 73.8 | 64.8 | 82.9 | 93 |
| False Alarms (%) | 36.1 | 26.1 | 50.9 | 93 |
| Overall RT (ms) | 561.1 | 518.8 | 607.4 | 93 |
| Correct-trial RT (ms) | 587.4 | 544.6 | 633.0 | 93 |
| Error-trial RT (ms) | 481.2 | 427.5 | 532.6 | 93 |
| Post-correct slowing (ms) | 10.3 | −9.0 | 37.8 | 91 |
| Post-error slowing (ms) | 148.3 | 88.8 | 197.9 | 91 |
| Post-correct accuracy (%) | 83.5 | 76.1 | 88.8 | 91 |
| Post-error accuracy (%) | 83.6 | 76.2 | 89.6 | 91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietto, M.L.; Giovannetti, F.; Segretin, M.S.; Lipina, S.J.; Kamienkowski, J.E. EEG Dynamics of Error Processing and Associated Behavioral Adjustments in Preschool Children. Brain Sci. 2023, 13, 575. https://doi.org/10.3390/brainsci13040575
Pietto ML, Giovannetti F, Segretin MS, Lipina SJ, Kamienkowski JE. EEG Dynamics of Error Processing and Associated Behavioral Adjustments in Preschool Children. Brain Sciences. 2023; 13(4):575. https://doi.org/10.3390/brainsci13040575
Chicago/Turabian StylePietto, Marcos Luis, Federico Giovannetti, María Soledad Segretin, Sebastián Javier Lipina, and Juan Esteban Kamienkowski. 2023. "EEG Dynamics of Error Processing and Associated Behavioral Adjustments in Preschool Children" Brain Sciences 13, no. 4: 575. https://doi.org/10.3390/brainsci13040575