
Dehydroepiandrosterone Attenuates Astroglial Activation, Neuronal Loss and Dendritic Degeneration in Iron-Induced Post-Traumatic Epilepsy



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Epilepsy Model
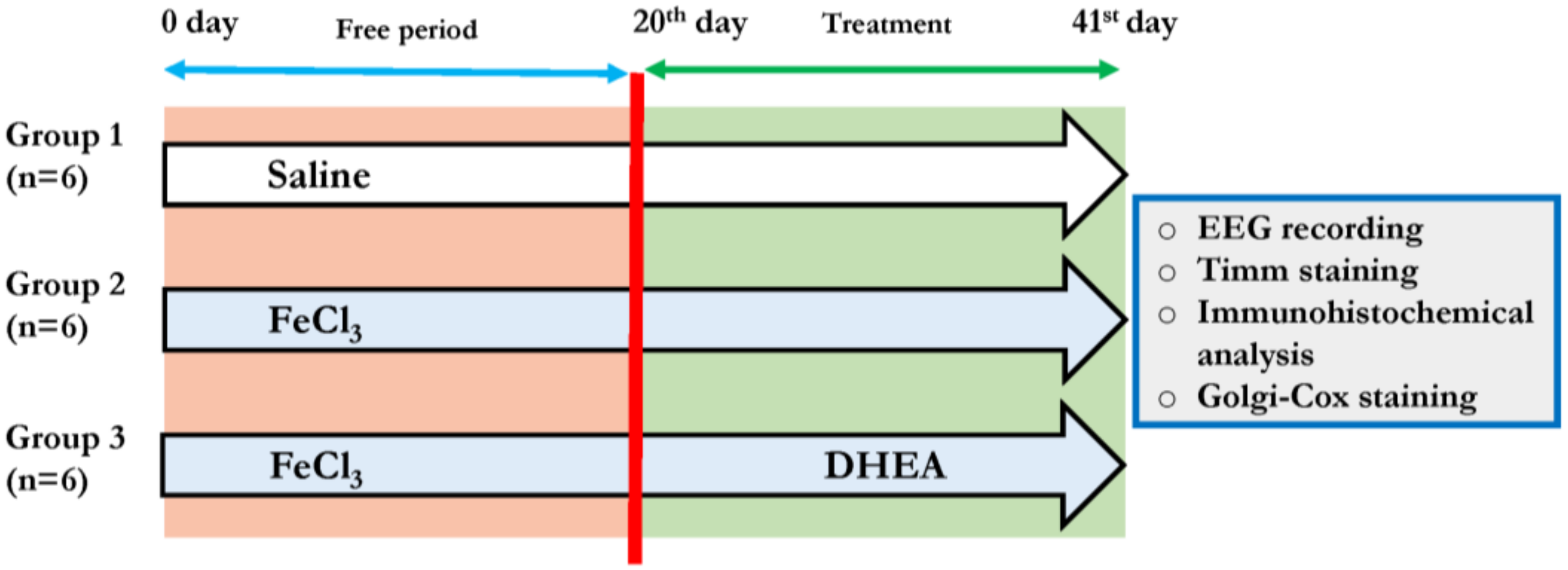
2.3. Experimental Design
2.4. EEG Recordings and Analysis
2.5. Tissue Preparation for Histopathology and Immunofluorescence Analysis
2.6. Timm Staining
2.7. Immunofluorescence Staining
2.8. Golgi-Cox Staining
2.9. Analysis of Dendritic Morphology and Spine Density
2.10. Statistical Analysis
3. Results
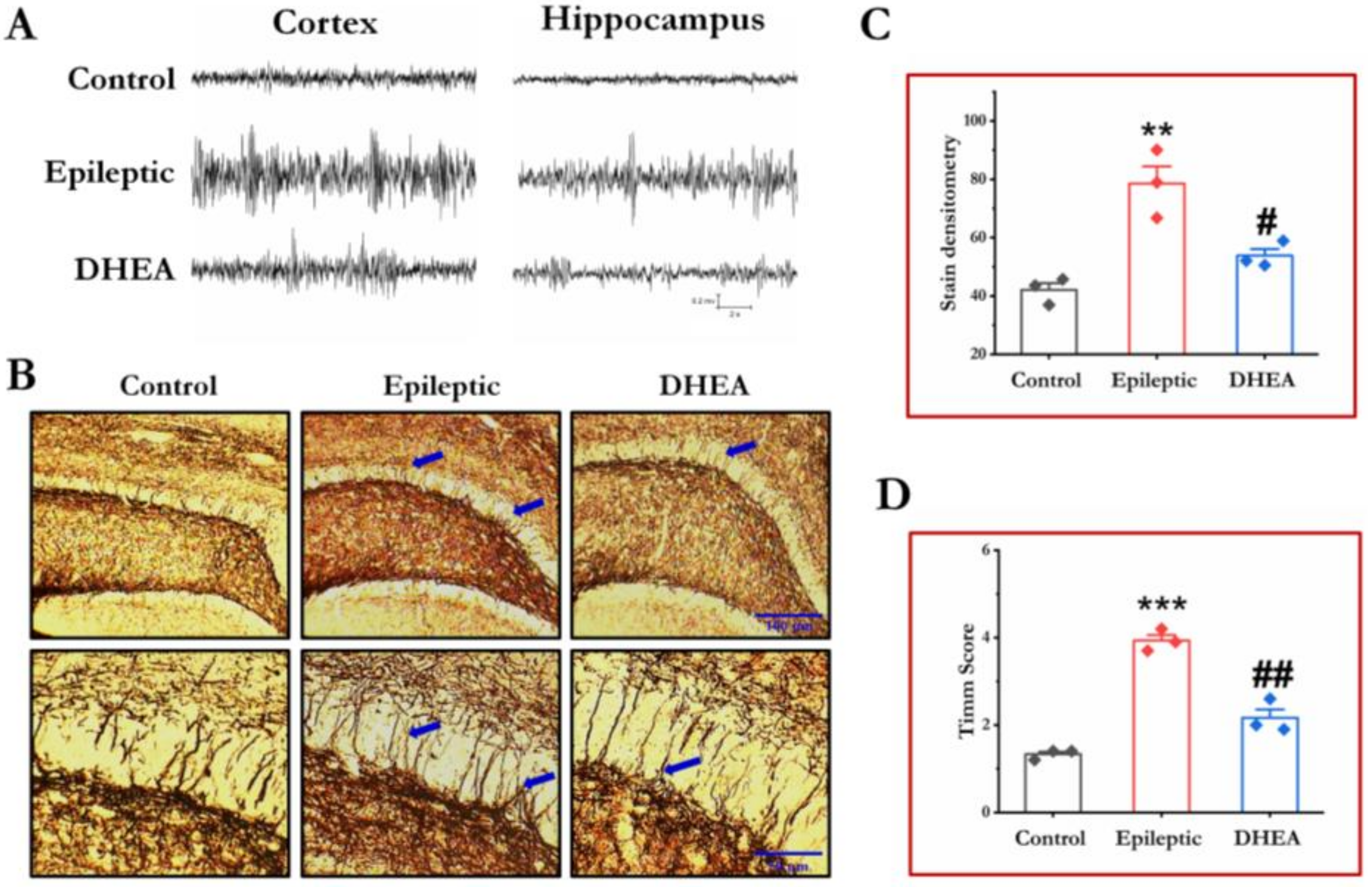
3.1. DHEA Treatment Alleviates Epileptiform Seizures in Epileptic Rats
3.2. DHEA Treatment Reduces MFs Sprouting in Epileptic Rats
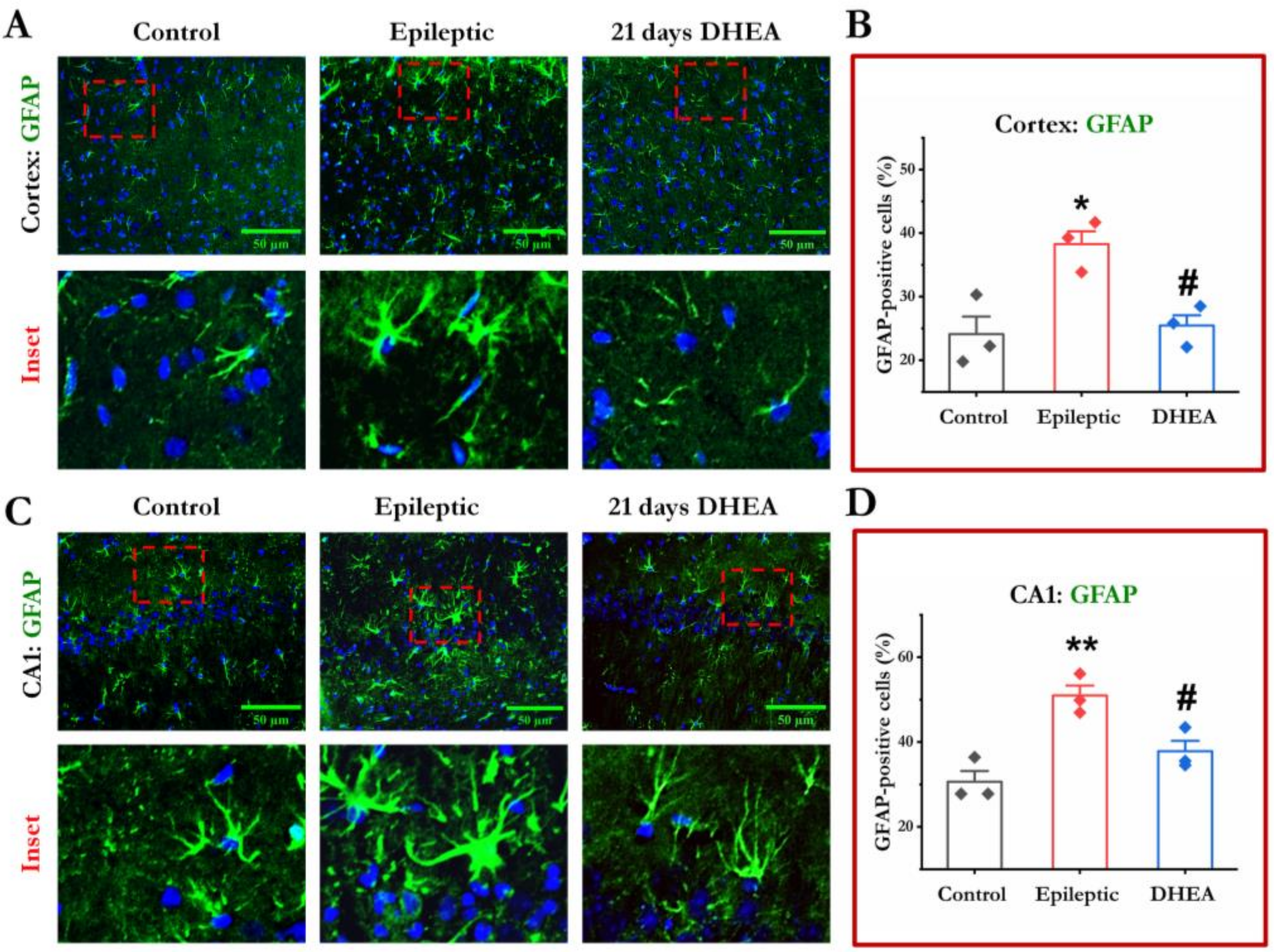
3.3. DHEA Treatment Attenuates Activation of Astrocytes
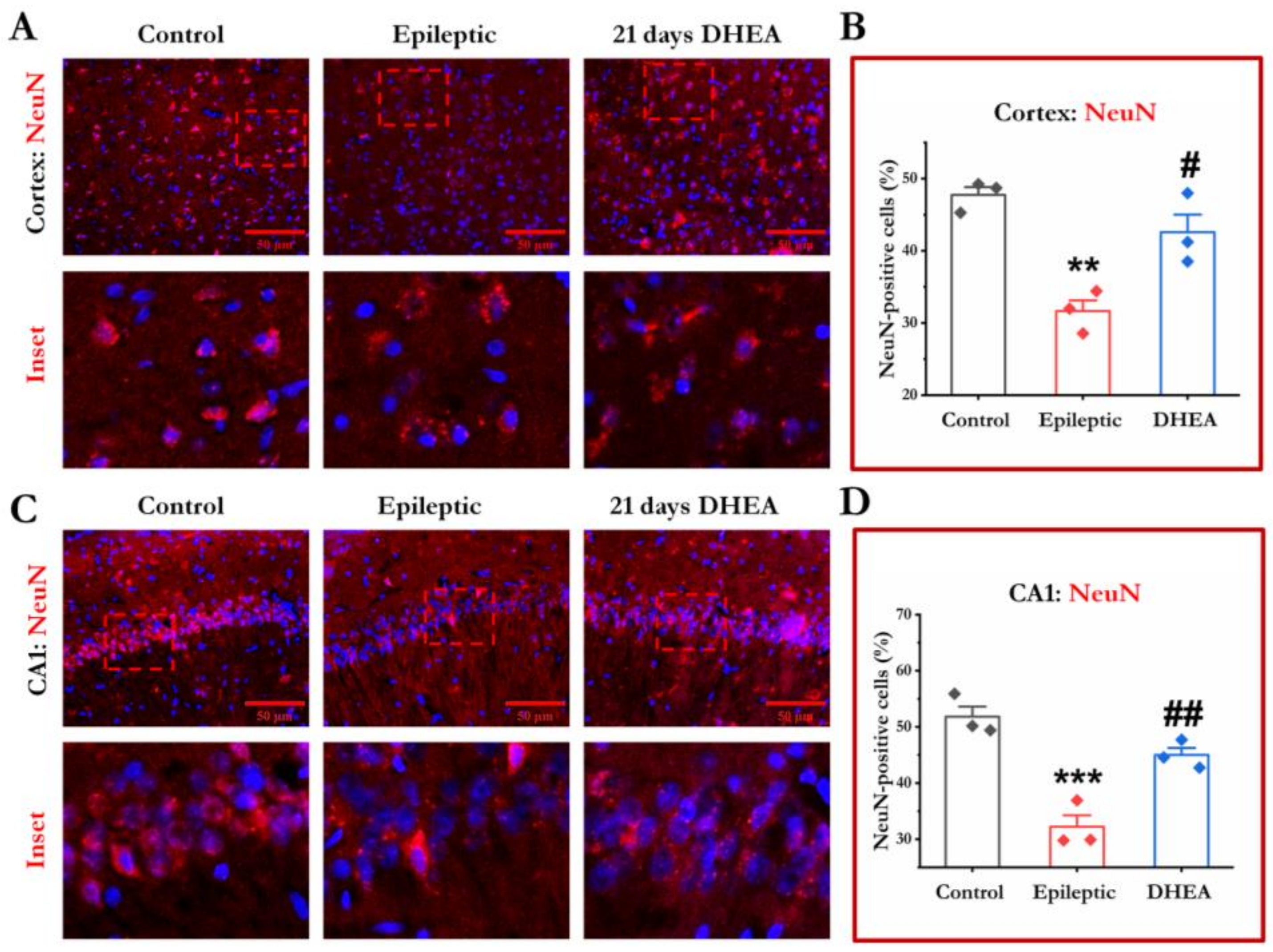
3.4. DHEA Treatment Protects Loss of Neurons
3.5. DHEA Treatment Rescues Dendritic Arborization
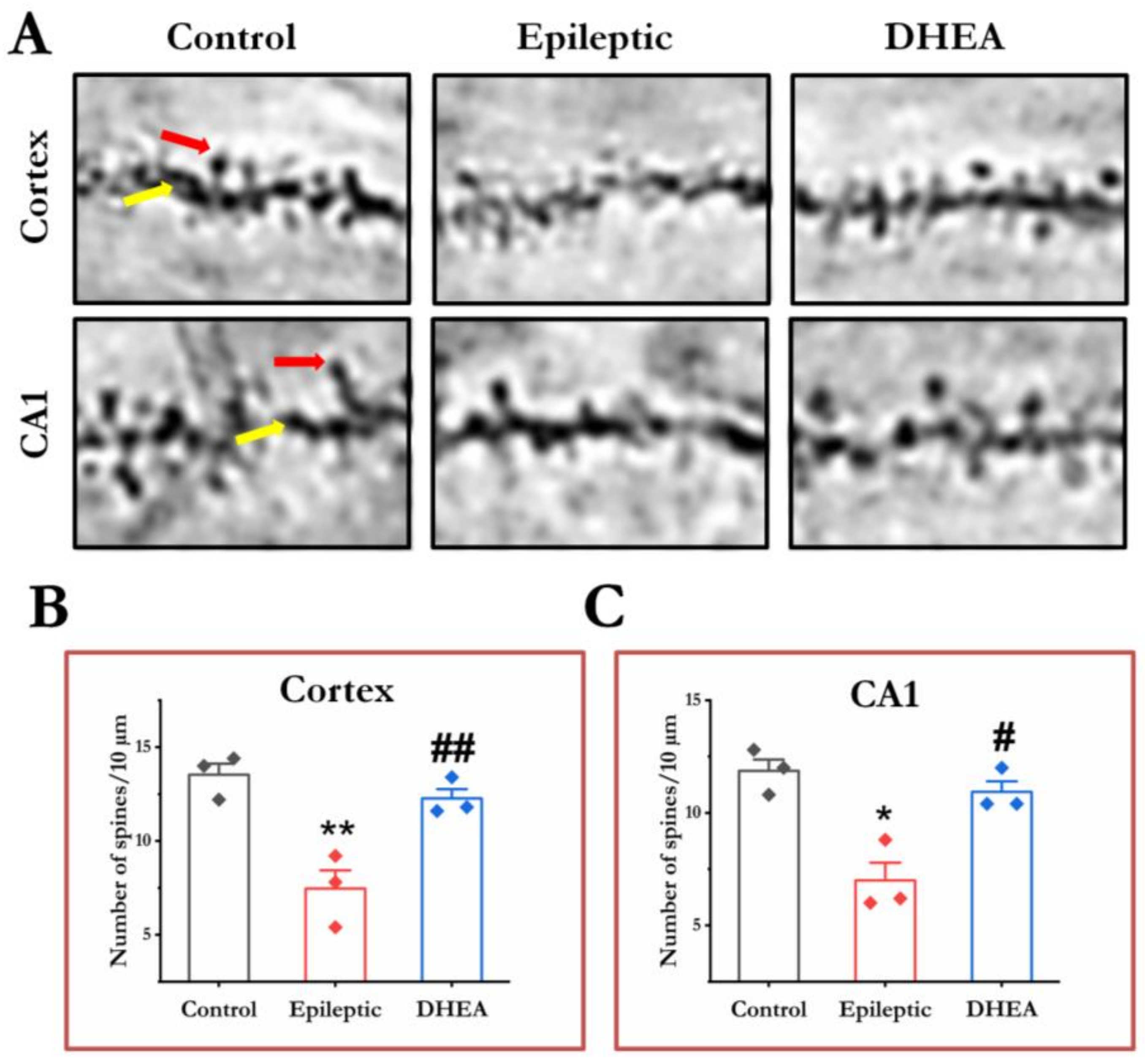
3.6. DHEA Restores Dendritic Spines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pearson-Smith, J.; Patel, M. Metabolic Dysfunction and Oxidative Stress in Epilepsy. Int. J. Mol. Sci. 2017, 18, 2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boison, D.; Steinhäuser, C. Epilepsy and Astrocyte Energy Metabolism. Glia 2018, 66, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Timothy, J.; Pandit, L.; Manju, M. Post-Traumatic Epilepsy: An Overview. Clin. Neurol. Neurosurg. 2006, 108, 433–439. [Google Scholar] [CrossRef]
- Jensen, F.E. Introduction Posttraumatic Epilepsy: Treatable Epileptogenesis. Epilepsia 2009, 50, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, M.P.; Leblanc, G.G.; Brooks-Kayal, A.; Jensen, F.E.; Lowenstein, D.H.; Noebels, J.L.; Spencer, D.D.; Swann, J.W. Curing Epilepsy: Progress and Future Directions. Epilepsy Behav. 2009, 14, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avoli, M.; de Curtis, M.; Gnatkovsky, V.; Gotman, J.; Köhling, R.; Lévesque, M.; Manseau, F.; Shiri, Z.; Williams, S. Specific Imbalance of Excitatory/Inhibitory Signaling Establishes Seizure Onset Pattern in Temporal Lobe Epilepsy. J. Neurophysiol. 2016, 115, 3229–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmore, L.J.; Sypert, G.W.; Munson, J.B. Recurrent Seizures Induced by Cortical Iron Injection: A Model of Posttraumatic Epilepsy. Ann. Neurol. 1978, 4, 329–336. [Google Scholar] [CrossRef]
- Prakash, C.; Mishra, M.; Kumar, P.; Kumar, V.; Sharma, D. Dehydroepiandrosterone Alleviates Oxidative Stress and Apoptosis in Iron-Induced Epilepsy via Activation of Nrf2/ARE Signal Pathway. Brain. Res. Bull. 2019, 153, 181–190. [Google Scholar] [CrossRef]
- Prakash, C.; Mishra, M.; Kumar, P.; Kumar, V.; Sharma, D. Response of Voltage-Gated Sodium and Calcium Channels Subtypes on Dehydroepiandrosterone Treatment in Iron-Induced Epilepsy. Cell. Mol. Neurobiol. 2021, 41, 279–292. [Google Scholar] [CrossRef]
- Kumar, V.; Prakash, C.; Singh, R.; Sharma, D. Curcumin’s Antiepileptic Effect, and Alterations in Nav1.1 and Nav1.6 Expression in Iron-Induced Epilepsy. Epilepsy Res. 2019, 150, 7–16. [Google Scholar] [CrossRef]
- Das, J.; Singh, R.; Sharma, D. Antiepileptic Effect of Fisetin in Iron-Induced Experimental Model of Traumatic Epilepsy in Rats in the Light of Electrophysiological, Biochemical, and Behavioral Observations. Nutr. Neurosci. 2017, 20, 255–264. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, D. Ameliorative Effect of Curcumin on Altered Expression of CACNA1A and GABRD in the Pathogenesis of FeCl3-Induced Epilepsy. Mol. Biol. Rep. 2020, 47, 5699–5710. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Guo, M.; Shi, C.; Wang, H.; Yao, L.; Liu, L.; Xie, C.; Pu, S.; LaChaud, G.; Shen, J.; et al. Protection against Cognitive Impairment and Modification of Epileptogenesis with Curcumin in a Post-Status Epilepticus Model of Temporal Lobe Epilepsy. Neuroscience 2015, 310, 362–371. [Google Scholar] [CrossRef]
- Zamudio, S.R.; Pichardo-Macías, L.A.; Díaz-Villegas, V.; Flores-Navarrete, I.L.; Guzmán-Velázquez, S. Subchronic Cerebrolysin Treatment Alleviates Cognitive Impairments and Dendritic Arborization Alterations of Granular Neurons in the Hippocampal Dentate Gyrus of Rats with Temporal Lobe Epilepsy. Epilepsy Behav. 2019, 97, 96–104. [Google Scholar] [CrossRef]
- Hashemian, M.; Anissian, D.; Ghasemi-Kasman, M.; Akbari, A.; Khalili-Fomeshi, M.; Ghasemi, S.; Ahmadi, F.; Moghadamnia, A.A.; Ebrahimpour, A. Curcumin-Loaded Chitosan-Alginate-STPP Nanoparticles Ameliorate Memory Deficits and Reduce Glial Activation in Pentylenetetrazol-Induced Kindling Model of Epilepsy. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, S.R.; Ghasemi-Kasman, M.; Pouramir, M.; Sadeghi, F. Arbutin Attenuates Cognitive Impairment and Inflammatory Response in Pentylenetetrazol-Induced Kindling Model of Epilepsy. Neuropharmacology 2019, 146, 117–127. [Google Scholar] [CrossRef]
- Cao, J.; Tang, C.; Gao, M.; Rui, Y.; Zhang, J.; Wang, L.; Wang, Y.; Xu, B.; Yan, B.C. Hyperoside Alleviates Epilepsy-Induced Neuronal Damage by Enhancing Antioxidant Levels and Reducing Autophagy. J. Ethnopharmacol. 2020, 257, 112884. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Vezzani, A.; Najjar, S.; de Lanerolle, N.C.; Rogawski, M.A. Glia and Epilepsy: Excitability and Inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef]
- Shimada, T.; Takemiya, T.; Sugiura, H.; Yamagata, K. Role of Inflammatory Mediators in the Pathogenesis of Epilepsy. Mediat. Inflamm. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tailby, C.; Kowalczyk, M.A.; Jackson, G.D. Cognitive Impairment in Epilepsy: The Role of Reduced Network Flexibility. Ann. Clin. Transl. Neurol. 2018, 5, 29–40. [Google Scholar] [CrossRef]
- Swann, J.W.; Al-Noori, S.; Jiang, M.; Lee, C.L. Spine Loss and Other Dendritic Abnormalities in Epilepsy. Hippocampus 2000, 10, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Li, T.; Song, X.; Sun, H.; Liu, J.; Yang, J.; Zhao, W.; Cheng, L.; Chen, H.; Liu, B.; et al. Dynamic Alteration of Dendrites and Dendritic Spines in the Hippocampus and Microglia in Mouse Brain Tissues after Kainate-Induced Status Epilepticus. Int. J. Neurosci. 2021, 131, 1045–1057. [Google Scholar] [CrossRef]
- Musto, A.E.; Rosencrans, R.F.; Walker, C.P.; Bhattacharjee, S.; Raulji, C.M.; Belayev, L.; Fang, Z.; Gordon, W.C.; Bazan, N.G. Dysfunctional Epileptic Neuronal Circuits and Dysmorphic Dendritic Spines Are Mitigated by Platelet-Activating Factor Receptor Antagonism. Sci. Rep. 2016, 6, 30298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossini, L.; de Santis, D.; Mauceri, R.R.; Tesoriero, C.; Bentivoglio, M.; Maderna, E.; Maiorana, A.; Deleo, F.; de Curtis, M.; Tringali, G.; et al. Dendritic Pathology, Spine Loss and Synaptic Reorganization in Human Cortex from Epilepsy Patients. Brain 2021, 144, 251–265. [Google Scholar] [CrossRef]
- Thom, M.; Sisodiya, S.; Harkness, W.; Scaravilli, F. Microdysgenesis in Temporal Lobe Epilepsy: A Quantitative and Immunohistochemical Study of White Matter Neurones. Brain 2001, 124, 2299–2309. [Google Scholar] [CrossRef] [Green Version]
- Freiman, T.M.; Eismann-Schweimler, J.; Frotscher, M. Granule Cell Dispersion in Temporal Lobe Epilepsy Is Associated with Changes in Dendritic Orientation and Spine Distribution. Exp. Neurol. 2011, 229, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Strzelczyk, A.; Schubert-Bast, S. Psychobehavioural and Cognitive Adverse Events of Anti-Seizure Medications for the Treatment of Developmental and Epileptic Encephalopathies. CNS Drugs 2022, 36, 1079–1111. [Google Scholar] [CrossRef]
- Pitkänen, A.; Sutula, T.P. Is Epilepsy a Progressive Disorder? Prospects for New Therapeutic Approaches in Temporal-Lobe Epilepsy. Lancet Neurol. 2002, 1, 173–181. [Google Scholar] [CrossRef]
- Kobow, K.; Auvin, S.; Jensen, F.; Löscher, W.; Mody, I.; Potschka, H.; Prince, D.; Sierra, A.; Simonato, M.; Pitkänen, A.; et al. Finding a Better Drug for Epilepsy: Antiepileptogenesis Targets. Epilepsia 2012, 53, 1868–1876. [Google Scholar] [CrossRef] [Green Version]
- Lazaridis, I.; Charalampopoulos, I.; Alexaki, V.-I.; Avlonitis, N.; Pediaditakis, I.; Efstathopoulos, P.; Calogeropoulou, T.; Castanas, E.; Gravanis, A. Neurosteroid Dehydroepiandrosterone Interacts with Nerve Growth Factor (NGF) Receptors, Preventing Neuronal Apoptosis. PLoS Biol. 2011, 9, e1001051. [Google Scholar] [CrossRef]
- Arbo, B.D.; Ribeiro, F.S.; Ribeiro, M.F. Astrocyte Neuroprotection and Dehydroepiandrosterone. In Vitamins and Hormones; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 108, pp. 175–203. ISBN 9780128143612. [Google Scholar]
- Alexaki, V.I.; Fodelianaki, G.; Neuwirth, A.; Mund, C.; Kourgiantaki, A.; Ieronimaki, E.; Lyroni, K.; Troullinaki, M.; Fujii, C.; Kanczkowski, W.; et al. DHEA Inhibits Acute Microglia-Mediated Inflammation through Activation of the TrkA-Akt1/2-CREB-Jmjd3 Pathway. Mol. Psychiatry 2018, 23, 1410–1420. [Google Scholar] [CrossRef]
- Mishra, M.; Singh, R.; Mukherjee, S.; Sharma, D. Dehydroepiandrosterone’s Antiepileptic Action in FeCl3-Induced Epileptogenesis Involves Upregulation of Glutamate Transporters. Epilepsy Res. 2013, 106, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Liu, L.; Wei, J.-L.; Dai, R.-P. Step by Step Golgi-Cox Staining for Cryosection. Front. Neuroanat. 2019, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Sholl, D.A. Dendritic Organization in the Neurons of the Visual and Motor Cortices of the Cat. J. Anat. 1953, 87, 387–406. [Google Scholar]
- Maiti, P.; Bowers, Z.; Bourcier-Schultz, A.; Morse, J.; Dunbar, G.L. Preservation of Dendritic Spine Morphology and Postsynaptic Signaling Markers after Treatment with Solid Lipid Curcumin Particles in the 5xFAD Mouse Model of Alzheimer’s Amyloidosis. Alzheimer’s Res. Ther. 2021, 13, 37. [Google Scholar] [CrossRef]
- Mishra, M.; Singh, R.; Sharma, D. Antiepileptic Action of Exogenous Dehydroepiandrosterone in Iron-Induced Epilepsy in Rat Brain. Epilepsy Behav. 2010, 19, 264–271. [Google Scholar] [CrossRef]
- Danzer, S. Mossy Fiber Sprouting in the Epileptic Brain: Taking on the Lernaean Hydra. Epilepsy Curr. 2017, 17, 50–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Patro, I.; Tikoo, K.; Sandhir, R. Curcumin Attenuates Inflammatory Response and Cognitive Deficits in Experimental Model of Chronic Epilepsy. Neurochem. Int. 2015, 89, 40–50. [Google Scholar] [CrossRef]
- Lee, H.S.; Jung, K.K.; Cho, J.Y.; Rhee, M.H.; Hong, S.; Kwon, M.; Kim, S.H.; Kang, S.Y. Neuroprotective Effect of Curcumin Is Mainly Mediated by Blockade of Microglial Cell Activation. Pharmazie 2007, 62, 937–942. [Google Scholar]
- Ortinski, P.I.; Dong, J.; Mungenast, A.; Yue, C.; Takano, H.; Watson, D.J.; Haydon, P.G.; Coulter, D.A. Selective Induction of Astrocytic Gliosis Generates Deficits in Neuronal Inhibition. Nat. Neurosci. 2010, 13, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellin, T.; Haydon, P.G. Do Astrocytes Contribute to Excitation Underlying Seizures? Trends Mol. Med. 2005, 11, 530–533. [Google Scholar] [CrossRef]
- Kapur, J. Role of Neuronal Loss in the Pathogenesis of Recurrent Spontaneous Seizures. Epilepsy Curr. 2003, 3, 166–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komoltsev, I.G.; Frankevich, S.O.; Shirobokova, N.I.; Volkova, A.A.; Onufriev, M.V.; Moiseeva, J.V.; Novikova, M.R.; Gulyaeva, N.V. Neuroinflammation and Neuronal Loss in the Hippocampus Are Associated with Immediate Posttraumatic Seizures and Corticosterone Elevation in Rats. Int. J. Mol. Sci. 2021, 22, 5883. [Google Scholar] [CrossRef] [PubMed]
- Young, N.A.; Szabó, C.Á.; Phelix, C.F.; Flaherty, D.K.; Balaram, P.; Foust-Yeoman, K.B.; Collins, C.E.; Kaas, J.H. Epileptic Baboons Have Lower Numbers of Neurons in Specific Areas of Cortex. Proc. Natl. Acad. Sci. USA 2013, 110, 19107–19112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Si, Z.; Li, X.; Lu, L.; Pan, Y.; Liu, J. FK506 Attenuated Pilocarpine-Induced Epilepsy by Reducing Inflammation in Rats. Front. Neurol. 2019, 10, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, C.K.; Herskowitz, J.H. Dendritic Spines: Mediators of Cognitive Resilience in Aging and Alzheimer’s Disease. Neuroscientist 2021, 27, 487–505. [Google Scholar] [CrossRef]
- Šišková, Z.; Justus, D.; Kaneko, H.; Friedrichs, D.; Henneberg, N.; Beutel, T.; Pitsch, J.; Schoch, S.; Becker, A.; von der Kammer, H.; et al. Dendritic Structural Degeneration Is Functionally Linked to Cellular Hyperexcitability in a Mouse Model of Alzheimer’s Disease. Neuron 2014, 84, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Teskey, G.C.; Monfils, M.-H.; Silasi, G.; Kolb, B. Neocortical Kindling Is Associated with Opposing Alterations in Dendritic Morphology in Neocortical Layer V and Striatum from Neocortical Layer III. Synapse 2006, 59, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Pérez, J.J.; Martínez-Mota, L.; Jiménez-Rubio, G.; Ortiz-López, L.; Cabrera-Muñoz, E.A.; Galindo-Sevilla, N.; Zambrano, E.; Hernández-Luis, F.; Ramírez-Rodríguez, G.B.; Flores-Ramos, M. Dehydroepiandrosterone Increases the Number and Dendrite Maturation of Doublecortin Cells in the Dentate Gyrus of Middle Age Male Wistar Rats Exposed to Chronic Mild Stress. Behav. Brain. Res 2017, 321, 137–147. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prakash, C.; Rabidas, S.S.; Tyagi, J.; Sharma, D. Dehydroepiandrosterone Attenuates Astroglial Activation, Neuronal Loss and Dendritic Degeneration in Iron-Induced Post-Traumatic Epilepsy. Brain Sci. 2023, 13, 563. https://doi.org/10.3390/brainsci13040563
Prakash C, Rabidas SS, Tyagi J, Sharma D. Dehydroepiandrosterone Attenuates Astroglial Activation, Neuronal Loss and Dendritic Degeneration in Iron-Induced Post-Traumatic Epilepsy. Brain Sciences. 2023; 13(4):563. https://doi.org/10.3390/brainsci13040563
Chicago/Turabian StylePrakash, Chandra, Shyam Sunder Rabidas, Jyoti Tyagi, and Deepak Sharma. 2023. "Dehydroepiandrosterone Attenuates Astroglial Activation, Neuronal Loss and Dendritic Degeneration in Iron-Induced Post-Traumatic Epilepsy" Brain Sciences 13, no. 4: 563. https://doi.org/10.3390/brainsci13040563