

Natural Oscillatory Frequency Slowing in the Premotor Cortex of Early-Course Schizophrenia Patients: A TMS-EEG Study
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.3. Data Analysis
2.4. Statistical Analysis
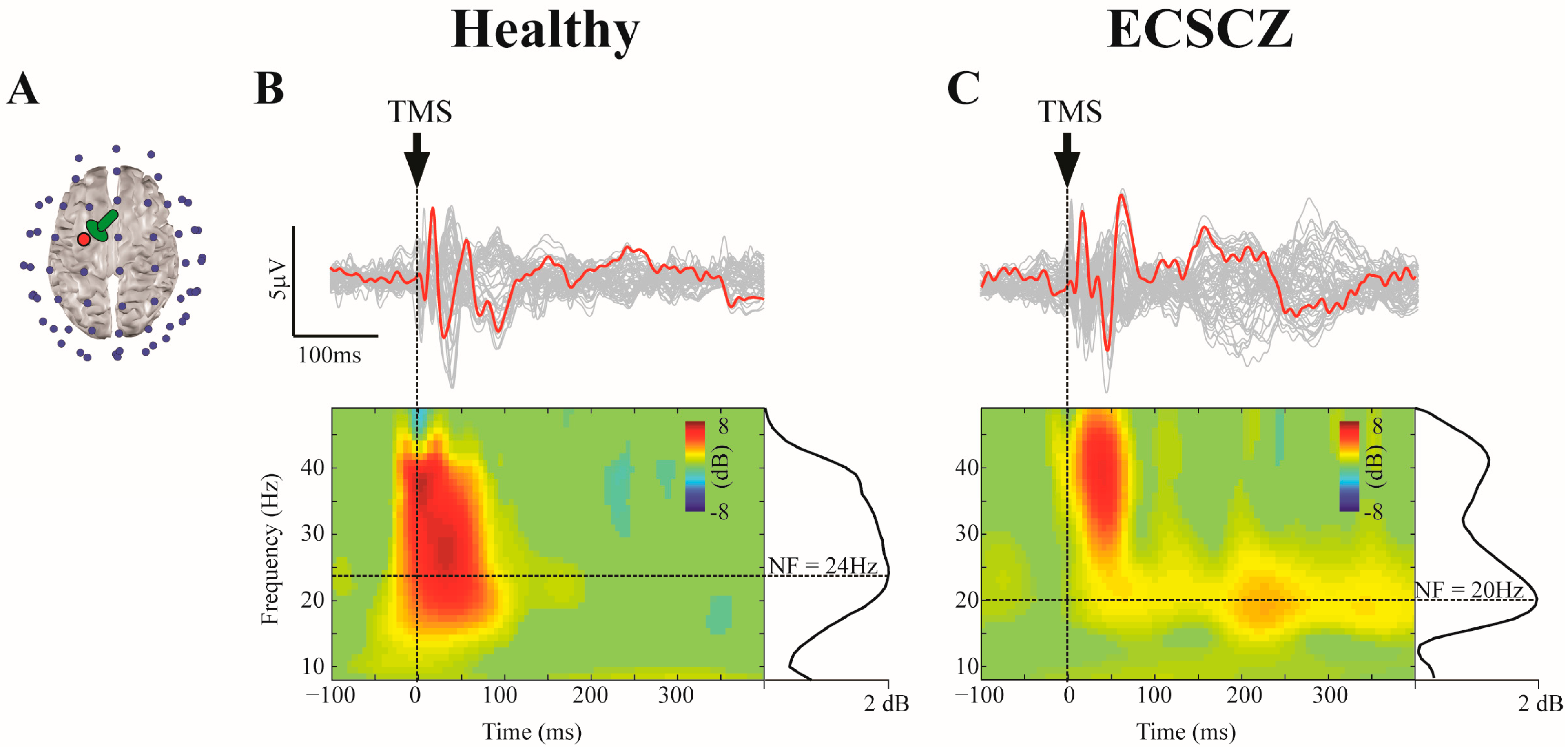
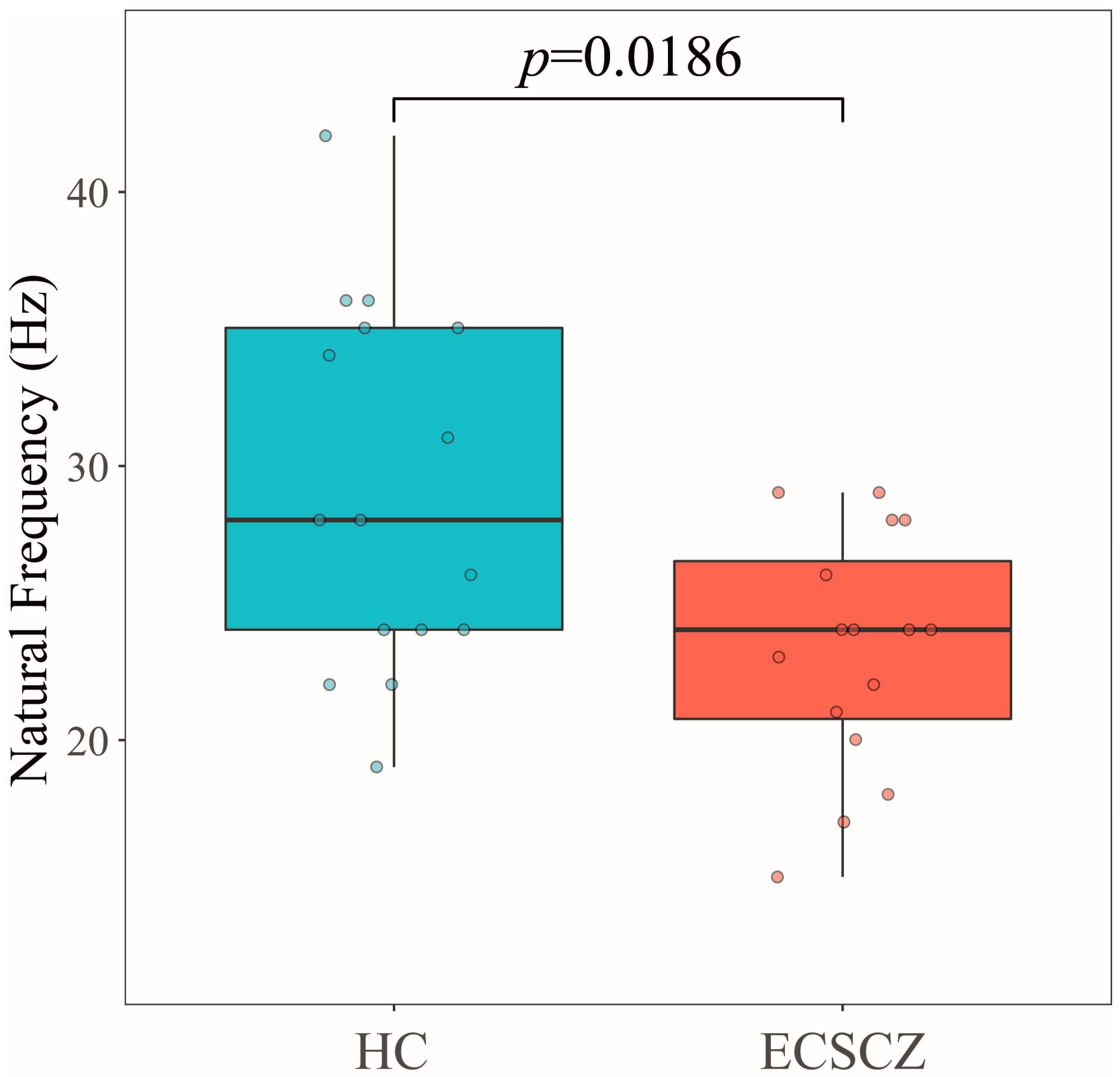
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charlson, F.J.; Ferrari, A.J.; Santomauro, D.F.; Diminic, S.; Stockings, E.; Scott, J.G.; McGrath, J.J.; Whiteford, H.A. Global Epidemiology and Burden of Schizophrenia: Findings From the Global Burden of Disease Study 2016. Schizophr. Bull 2018, 44, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Norman, R.M.G.; Manchanda, R.; Malla, A.K.; Windell, D.; Harricharan, R.; Northcott, S. Symptom and Functional Outcomes for a 5 Year Early Intervention Program for Psychoses. Schizophr. Res. 2011, 129, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Jauhar, S.; Johnstone, M.; McKenna, P.J. Schizophrenia. Lancet 2022, 399, 473–486. [Google Scholar] [CrossRef] [PubMed]
- McGorry, P.; Keshavan, M.; Goldstone, S.; Amminger, P.; Allott, K.; Berk, M.; Lavoie, S.; Pantelis, C.; Yung, A.; Wood, S.; et al. Biomarkers and Clinical Staging in Psychiatry. World Psychiatry 2014, 13, 211–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, F.L.; D’Agostino, A.; Ferrarelli, F. Neurocognitive and Neurophysiological Endophenotypes in Schizophrenia: An Overview. Biomark. Neuropsychiatry 2020, 3, 100017. [Google Scholar] [CrossRef]
- Rosson, S.; de Filippis, R.; Croatto, G.; Collantoni, E.; Pallottino, S.; Guinart, D.; Brunoni, A.R.; Dell’Osso, B.; Pigato, G.; Hyde, J.; et al. Brain Stimulation and Other Biological Non-Pharmacological Interventions in Mental Disorders: An Umbrella Review. Neurosci. Biobehav. Rev. 2022, 139, 104743. [Google Scholar] [CrossRef] [PubMed]
- Price, R.B.; Gillan, C.M.; Hanlon, C.; Ferrarelli, F.; Kim, T.; Karim, H.T.; Renard, M.; Kaskie, R.; Degutis, M.; Wears, A.; et al. Effect of Experimental Manipulation of the Orbitofrontal Cortex on Short-Term Markers of Compulsive Behavior: A Theta Burst Stimulation Study. Am. J. Psychiatry 2021, 178, 459–468. [Google Scholar] [CrossRef]
- Casali, A.G.; Casarotto, S.; Rosanova, M.; Mariotti, M.; Massimini, M. General Indices to Characterize the Electrical Response of the Cerebral Cortex to TMS. NeuroImage 2010, 49, 1459–1468. [Google Scholar] [CrossRef]
- Rosanova, M.; Casarotto, S.; Pigorini, A.; Canali, P.; Casali, A.G.; Massimini, M. Combining Transcranial Magnetic Stimulation with Electroencephalography to Study Human Cortical Excitability and Effective Connectivity. In Neuronal Network Analysis: Concepts and Experimental Approaches; Fellin, T., Halassa, M., Eds.; Neuromethods; Humana Press: Totowa, NJ, USA, 2012; pp. 435–457. ISBN 978-1-61779-633-3. [Google Scholar]
- Ferreri, F.; Pasqualetti, P.; Määttä, S.; Ponzo, D.; Ferrarelli, F.; Tononi, G.; Mervaala, E.; Miniussi, C.; Rossini, P.M. Human Brain Connectivity during Single and Paired Pulse Transcranial Magnetic Stimulation. NeuroImage 2011, 54, 90–102. [Google Scholar] [CrossRef]
- Daskalakis, Z.J.; Farzan, F.; Barr, M.S.; Maller, J.J.; Chen, R.; Fitzgerald, P.B. Long-Interval Cortical Inhibition from the Dorsolateral Prefrontal Cortex: A TMS–EEG Study. Neuropsychopharmacol 2008, 33, 2860–2869. [Google Scholar] [CrossRef] [Green Version]
- Farzan, F.; Barr, M.S.; Wong, W.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Suppression of γ-Oscillations in the Dorsolateral Prefrontal Cortex Following Long Interval Cortical Inhibition: A TMS–EEG Study. Neuropsychopharmacol 2009, 34, 1543–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreri, F.; Ponzo, D.; Hukkanen, T.; Mervaala, E.; Könönen, M.; Pasqualetti, P.; Vecchio, F.; Rossini, P.M.; Määttä, S. Human Brain Cortical Correlates of Short-Latency Afferent Inhibition: A Combined EEG–TMS Study. J. Neurophysiol. 2012, 108, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Noda, Y.; Cash, R.F.H.; Zomorrodi, R.; Dominguez, L.G.; Farzan, F.; Rajji, T.K.; Barr, M.S.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. A Combined TMS-EEG Study of Short-Latency Afferent Inhibition in the Motor and Dorsolateral Prefrontal Cortex. J. Neurophysiol. 2016, 116, 938–948. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, S.; Rogasch, N.C.; Premoli, I.; Blumberger, D.M.; Casarotto, S.; Chen, R.; Di Lazzaro, V.; Farzan, F.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Clinical Utility and Prospective of TMS-EEG. Clin. Neurophysiol 2019, 130, 802–844. [Google Scholar] [CrossRef]
- Rosanova, M.; Casali, A.; Bellina, V.; Resta, F.; Mariotti, M.; Massimini, M. Natural Frequencies of Human Corticothalamic Circuits. J. Neurosci. 2009, 29, 7679–7685. [Google Scholar] [CrossRef] [PubMed]
- Ferrarelli, F.; Sarasso, S.; Guller, Y.; Riedner, B.A.; Peterson, M.J.; Bellesi, M.; Massimini, M.; Postle, B.R.; Tononi, G. Reduced Natural Oscillatory Frequency of Frontal Thalamocortical Circuits in Schizophrenia. Arch. Gen. Psychiatry 2012, 69, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ambrosio, S.; Jiménez-Jiménez, D.; Silvennoinen, K.; Zagaglia, S.; Perulli, M.; Poole, J.; Comolatti, R.; Fecchio, M.; Sisodiya, S.M.; Balestrini, S. Physiological Symmetry of Transcranial Magnetic Stimulation-Evoked EEG Spectral Features. Hum. Brain Mapp. 2022, 43, 5465–5477. [Google Scholar] [CrossRef]
- Parmigiani, S.; Mikulan, E.; Russo, S.; Sarasso, S.; Zauli, F.M.; Rubino, A.; Cattani, A.; Fecchio, M.; Giampiccolo, D.; Lanzone, J.; et al. Simultaneous Stereo-EEG and High-Density Scalp EEG Recordings to Study the Effects of Intracerebral Stimulation Parameters. Brain Stimul. 2022, 15, 664–675. [Google Scholar] [CrossRef]
- Ferrarelli, F.; Massimini, M.; Peterson, M.J.; Riedner, B.A.; Lazar, M.; Murphy, M.J.; Huber, R.; Rosanova, M.; Alexander, A.L.; Kalin, N.; et al. Reduced Evoked Gamma Oscillations in the Frontal Cortex in Schizophrenia Patients: A TMS/EEG Study. Am. J. Psychiatry 2008, 165, 996–1005. [Google Scholar] [CrossRef]
- Levit-Binnun, N.; Litvak, V.; Pratt, H.; Moses, E.; Zaroor, M.; Peled, A. Differences in TMS-Evoked Responses between Schizophrenia Patients and Healthy Controls Can Be Observed without a Dedicated EEG System. Clin. Neurophysiol 2010, 121, 332–339. [Google Scholar] [CrossRef]
- Radhu, N.; Garcia Dominguez, L.; Farzan, F.; Richter, M.A.; Semeralul, M.O.; Chen, R.; Fitzgerald, P.B.; Daskalakis, Z.J. Evidence for Inhibitory Deficits in the Prefrontal Cortex in Schizophrenia. Brain 2015, 138, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhu, N.; Dominguez, L.G.; Greenwood, T.A.; Farzan, F.; Semeralul, M.O.; Richter, M.A.; Kennedy, J.L.; Blumberger, D.M.; Chen, R.; Fitzgerald, P.B.; et al. Investigating Cortical Inhibition in First-Degree Relatives and Probands in Schizophrenia. Scientific Reports 2017, 7, 43629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzan, F.; Barr, M.S.; Levinson, A.J.; Chen, R.; Wong, W.; Fitzgerald, P.B.; Daskalakis, Z.J. Evidence for Gamma Inhibition Deficits in the Dorsolateral Prefrontal Cortex of Patients with Schizophrenia. Brain 2010, 133, 1505–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, Y.; Barr, M.S.; Zomorrodi, R.; Cash, R.F.H.; Rajji, T.K.; Farzan, F.; Chen, R.; George, T.P.; Daskalakis, Z.J.; Blumberger, D.M. Reduced Short-Latency Afferent Inhibition in Prefrontal but Not Motor Cortex and Its Association With Executive Function in Schizophrenia: A Combined TMS-EEG Study. Schizophr. Bull. 2018, 44, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smucny, J.; Dienel, S.J.; Lewis, D.A.; Carter, C.S. Mechanisms Underlying Dorsolateral Prefrontal Cortex Contributions to Cognitive Dysfunction in Schizophrenia. Neuropsychopharmacol. 2022, 47, 292–308. [Google Scholar] [CrossRef]
- Hernandez-Pavon, J.C.; Veniero, D.; Bergmann, T.O.; Belardinelli, P.; Bortoletto, M.; Casarotto, S.; Casula, E.P.; Farzan, F.; Fecchio, M.; Julkunen, P.; et al. TMS Combined with EEG: Recommendations and Open Issues for Data Collection and Analysis. Brain Stimul. 2023, 16, 567–593. [Google Scholar] [CrossRef]
- Korhonen, R.J.; Hernandez-Pavon, J.C.; Metsomaa, J.; Mäki, H.; Ilmoniemi, R.J.; Sarvas, J. Removal of Large Muscle Artifacts from Transcranial Magnetic Stimulation-Evoked EEG by Independent Component Analysis. Med. Biol. Eng. Comput. 2011, 49, 397–407. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Biabani, M.; Mutanen, T.P. Designing and Comparing Cleaning Pipelines for TMS-EEG Data: A Theoretical Overview and Practical Example. J. Neurosci. Methods 2022, 371, 109494. [Google Scholar] [CrossRef]
- Belardinelli, P.; Biabani, M.; Blumberger, D.M.; Bortoletto, M.; Casarotto, S.; David, O.; Desideri, D.; Etkin, A.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Reproducibility in TMS-EEG Studies: A Call for Data Sharing, Standard Procedures and Effective Experimental Control. Brain Stimul. 2019, 12, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Mutanen, T.; Mäki, H.; Ilmoniemi, R.J. The Effect of Stimulus Parameters on TMS–EEG Muscle Artifacts. Brain Stimul. 2013, 6, 371–376. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Thomson, R.H.; Daskalakis, Z.J.; Fitzgerald, P.B. Short-Latency Artifacts Associated with Concurrent TMS–EEG. Brain Stimul. 2013, 6, 868–876. [Google Scholar] [CrossRef]
- Newton, R.; Rouleau, A.; Nylander, A.-G.; Loze, J.-Y.; Resemann, H.K.; Steeves, S.; Crespo-Facorro, B. Diverse Definitions of the Early Course of Schizophrenia—A Targeted Literature Review. Npj Schizophr. 2018, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kübler, U. Structured Clinical Interview for DSM-IV (SCID). In Encyclopedia of Behavioral Medicine; Gellman, M.D., Turner, J.R., Eds.; Springer: New York, NY, YSA, 2013; pp. 1919–1920. ISBN 978-1-4419-1005-9. [Google Scholar]
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmöller, J.; Carpenter, L.L.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; et al. Safety and Recommendations for TMS Use in Healthy Subjects and Patient Populations, with Updates on Training, Ethical and Regulatory Issues: Expert Guidelines. Clin. Neurophysiol. 2021, 132, 269–306. [Google Scholar] [CrossRef] [PubMed]
- Lacadie, C.M.; Fulbright, R.K.; Constable, R.T.; Papademetris, X. More Accurate Talairach Coordinates for NeuroImaging Using Nonlinear Registration. Neuroimage 2008, 42, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casarotto, S.; Fecchio, M.; Rosanova, M.; Varone, G.; D’Ambrosio, S.; Sarasso, S.; Pigorini, A.; Russo, S.; Comanducci, A.; Ilmoniemi, R.J.; et al. The Rt-TEP Tool: Real-Time Visualization of TMS-Evoked Potentials to Maximize Cortical Activation and Minimize Artifacts. J. Neurosci. Methods 2022, 370, 109486. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.; Sarasso, S.; Puglisi, G.E.; Dal Palù, D.; Pigorini, A.; Casarotto, S.; D’Ambrosio, S.; Astolfi, A.; Massimini, M.; Rosanova, M.; et al. TAAC—TMS Adaptable Auditory Control: A Universal Tool to Mask TMS Clicks. J. Neurosci. Methods 2022, 370, 109491. [Google Scholar] [CrossRef]
- Casarotto, S.; Määttä, S.; Herukka, S.-K.; Pigorini, A.; Napolitani, M.; Gosseries, O.; Niskanen, E.; Könönen, M.; Mervaala, E.; Rosanova, M.; et al. Transcranial Magnetic Stimulation-Evoked EEG/Cortical Potentials in Physiological and Pathological Aging. Neuroreport 2011, 22, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Rosanova, M.; Fecchio, M.; Casarotto, S.; Sarasso, S.; Casali, A.G.; Pigorini, A.; Comanducci, A.; Seregni, F.; Devalle, G.; Citerio, G.; et al. Sleep-like Cortical OFF-Periods Disrupt Causality and Complexity in the Brain of Unresponsive Wakefulness Syndrome Patients. Nat. Commun. 2018, 9, 4427. [Google Scholar] [CrossRef] [Green Version]
- Gosseries, O.; Fecchio, M.; Wolff, A.; Sanz, L.R.D.; Sombrun, C.; Vanhaudenhuyse, A.; Laureys, S. Behavioural and Brain Responses in Cognitive Trance: A TMS-EEG Case Study. Clin. Neurophysiol. 2020, 131, 586–588. [Google Scholar] [CrossRef]
- Canali, P.; Sarasso, S.; Rosanova, M.; Casarotto, S.; Sferrazza-Papa, G.; Gosseries, O.; Fecchio, M.; Massimini, M.; Mariotti, M.; Cavallaro, R.; et al. Shared Reduction of Oscillatory Natural Frequencies in Bipolar Disorder, Major Depressive Disorder and Schizophrenia. J. Affect Disord. 2015, 184, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Casarotto, S.; Romero Lauro, L.J.; Bellina, V.; Casali, A.G.; Rosanova, M.; Pigorini, A.; Defendi, S.; Mariotti, M.; Massimini, M. EEG Responses to TMS Are Sensitive to Changes in the Perturbation Parameters and Repeatable over Time. PLoS ONE 2010, 5, e10281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocchi, L.; Di Santo, A.; Brown, K.; Ibáñez, J.; Casula, E.; Rawji, V.; Di Lazzaro, V.; Koch, G.; Rothwell, J. Disentangling EEG Responses to TMS Due to Cortical and Peripheral Activations. Brain Stimul. 2021, 14, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Baird, B.; Gosseries, O.; Nieminen, J.O.; Boly, M.; Postle, B.R.; Tononi, G.; Lee, S.-W. Connectivity Differences between Consciousness and Unconsciousness in Non-Rapid Eye Movement Sleep: A TMS–EEG Study. Sci Rep 2019, 9, 5175. [Google Scholar] [CrossRef] [Green Version]
- Farnes, N.; Juel, B.E.; Nilsen, A.S.; Romundstad, L.G.; Storm, J.F. Increased Signal Diversity/Complexity of Spontaneous EEG, but Not Evoked EEG Responses, in Ketamine-Induced Psychedelic State in Humans. PLoS ONE 2020, 15, e0242056. [Google Scholar] [CrossRef]
- Lehmann, D.; Skrandies, W. Reference-Free Identification of Components of Checkerboard-Evoked Multichannel Potential Fields. Electroencephalogr. Clin. Neurophysiol. 1980, 48, 609–621. [Google Scholar] [CrossRef]
- Parmigiani, S.; Ross, J.M.; Cline, C.; Minasi, C.; Gogulski, J.; Keller, C.J. Reliability and Validity of TMS-EEG Biomarkers. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2022. [Google Scholar] [CrossRef]
- Donati, F.L.; Kaskie, R.; Reis, C.C.; D’Agostino, A.; Casali, A.G.; Ferrarelli, F. Reduced TMS-Evoked Fast Oscillations in the Motor Cortex Predict the Severity of Positive Symptoms in First-Episode Psychosis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 111, 110387. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P.J.; Singer, W. Abnormal Neural Oscillations and Synchrony in Schizophrenia. Nat. Rev. Neurosci. 2010, 11, 100–113. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Oscillations and Neuronal Dynamics in Schizophrenia: The Search for Basic Symptoms and Translational Opportunities. Biol. Psychiatry 2015, 77, 1001–1009. [Google Scholar] [CrossRef]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. Parvalbumin Neurons and Gamma Rhythms Enhance Cortical Circuit Performance. Nature 2009, 459, 698–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, K. The Functional Consequences of Cortical Circuit Abnormalities on Gamma Oscillations in Schizophrenia: Insights from Computational Modeling. Front. Hum. Neurosci. 2009, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Burgos, G.; Cho, R.Y.; Lewis, D.A. Alterations in Cortical Network Oscillations and Parvalbumin Neurons in Schizophrenia. Biol. Psychiatry 2015, 77, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Kaar, S.J.; Angelescu, I.; Marques, T.R.; Howes, O.D. Pre-Frontal Parvalbumin Interneurons in Schizophrenia: A Meta-Analysis of Post-Mortem Studies. J. Neural. Transm 2019, 126, 1637–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premoli, I.; Bergmann, T.O.; Fecchio, M.; Rosanova, M.; Biondi, A.; Belardinelli, P.; Ziemann, U. The Impact of GABAergic Drugs on TMS-Induced Brain Oscillations in Human Motor Cortex. NeuroImage 2017, 163, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wobrock, T.; Schneider, M.; Kadovic, D.; Schneider-Axmann, T.; Ecker, U.K.H.; Retz, W.; Rösler, M.; Falkai, P. Reduced Cortical Inhibition in First-Episode Schizophrenia. Schizophr. Res. 2008, 105, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Nitsche, M.A.; Rein, B.; Schneider-Axmann, T.; Guse, B.; Gruber, O.; Falkai, P.; Wobrock, T. Dysfunctional Long-Term Potentiation-like Plasticity in Schizophrenia Revealed by Transcranial Direct Current Stimulation. Behav. Brain Res. 2011, 224, 15–22. [Google Scholar] [CrossRef]
- Julkunen, P.; Kimiskidis, V.K.; Belardinelli, P. Bridging the Gap: TMS-EEG from Lab to Clinic. J. Neurosci. Methods 2022, 369, 109482. [Google Scholar] [CrossRef]
- Alnæs, D.; Kaufmann, T.; van der Meer, D.; Córdova-Palomera, A.; Rokicki, J.; Moberget, T.; Bettella, F.; Agartz, I.; Barch, D.M.; Bertolino, A.; et al. Brain Heterogeneity in Schizophrenia and Its Association With Polygenic Risk. JAMA Psychiatry 2019, 76, 739–748. [Google Scholar] [CrossRef]
- Chand, G.B.; Dwyer, D.B.; Erus, G.; Sotiras, A.; Varol, E.; Srinivasan, D.; Doshi, J.; Pomponio, R.; Pigoni, A.; Dazzan, P.; et al. Two Distinct Neuroanatomical Subtypes of Schizophrenia Revealed Using Machine Learning. Brain 2020, 143, 1027–1038. [Google Scholar] [CrossRef]
- Canali, P.; Casarotto, S.; Rosanova, M.; Sferrazza-Papa, G.; Casali, A.G.; Gosseries, O.; Massimini, M.; Smeraldi, E.; Colombo, C.; Benedetti, F. Abnormal Brain Oscillations Persist after Recovery from Bipolar Depression. Eur. Psychiatry 2017, 41, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandran, C.; Shinn, A.K.; Öngür, D.; Perlis, R.H.; Cohen, B. Frequency of Non-Right-Handedness in Bipolar Disorder and Schizophrenia. Psychiatry Res. 2017, 253, 267–269. [Google Scholar] [CrossRef] [PubMed]
- van Veelen, N.M.J.; Vink, M.; Ramsey, N.F.; Sommer, I.E.C.; van Buuren, M.; Hoogendam, J.M.; Kahn, R.S. Reduced Language Lateralization in First-Episode Medication-Naive Schizophrenia. Schizophr. Res. 2011, 127, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Zhu, Y.; Lu, Z.; Liu, N.; Huang, N.; Zhang, Z.; Tan, L.; Li, C.; Yu, X. Altered Volume and Lateralization of Language-Related Regions in First-Episode Schizophrenia. Schizophr. Res. 2013, 148, 168–174. [Google Scholar] [CrossRef]
- Ferrarelli, F.; Phillips, M.L. Examining and Modulating Neural Circuits in Psychiatric Disorders With Transcranial Magnetic Stimulation and Electroencephalography: Present Practices and Future Developments. AJP 2021, 178, 400–413. [Google Scholar] [CrossRef]
- Gogulski, J.; Ross, J.M.; Talbot, A.; Cline, C.C.; Donati, F.L.; Munot, S.; Kim, N.; Gibbs, C.; Bastin, N.; Yang, J.; et al. Personalized RTMS for Depression: A Review. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2022. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HC (N = 16) | ECSCZ (N = 16) | p Val | |
|---|---|---|---|
| Age | 23.43 ± 3.88 | 23.87 ± 4.29 | 0.7657 |
| Gender | 10 M; 6 F | 11 M; 5 F | 0.7097 |
| SAPS | - | 8.81 ± 6.99 | - |
| SANS | - | 32 ± 11.3 | - |
| CPZ eq | - | 331.16 ± 178.15 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, F.L.; Mayeli, A.; Sharma, K.; Janssen, S.A.; Lagoy, A.D.; Casali, A.G.; Ferrarelli, F. Natural Oscillatory Frequency Slowing in the Premotor Cortex of Early-Course Schizophrenia Patients: A TMS-EEG Study. Brain Sci. 2023, 13, 534. https://doi.org/10.3390/brainsci13040534
Donati FL, Mayeli A, Sharma K, Janssen SA, Lagoy AD, Casali AG, Ferrarelli F. Natural Oscillatory Frequency Slowing in the Premotor Cortex of Early-Course Schizophrenia Patients: A TMS-EEG Study. Brain Sciences. 2023; 13(4):534. https://doi.org/10.3390/brainsci13040534
Chicago/Turabian StyleDonati, Francesco L., Ahmad Mayeli, Kamakashi Sharma, Sabine A. Janssen, Alice D. Lagoy, Adenauer G. Casali, and Fabio Ferrarelli. 2023. "Natural Oscillatory Frequency Slowing in the Premotor Cortex of Early-Course Schizophrenia Patients: A TMS-EEG Study" Brain Sciences 13, no. 4: 534. https://doi.org/10.3390/brainsci13040534