

Minocycline Attenuates Sevoflurane-Induced Postoperative Cognitive Dysfunction in Aged Mice by Suppressing Hippocampal Apoptosis and the Notch Signaling Pathway-Mediated Neuroinflammation
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Minocycline Treatment and Sevoflurane Exposure
2.3. Cell Culture and Sevoflurane Treatment
2.4. Bromodeoxyuridine (BrdU)-Labeling
2.5. Morris Water Maze (MWM) Test
2.6. Fear Conditioning Assay
2.7. Brain Immunofluorescence
2.8. Cell Immunofluorescence
2.9. TUNEL-Based Cell Apoptosis Analysis
2.10. Western Blotting
2.11. Reverse Transcription and Real-Time Quantitative PCR
2.12. Electrophysiological Analysis
2.13. Statistical Analysis
3. Results
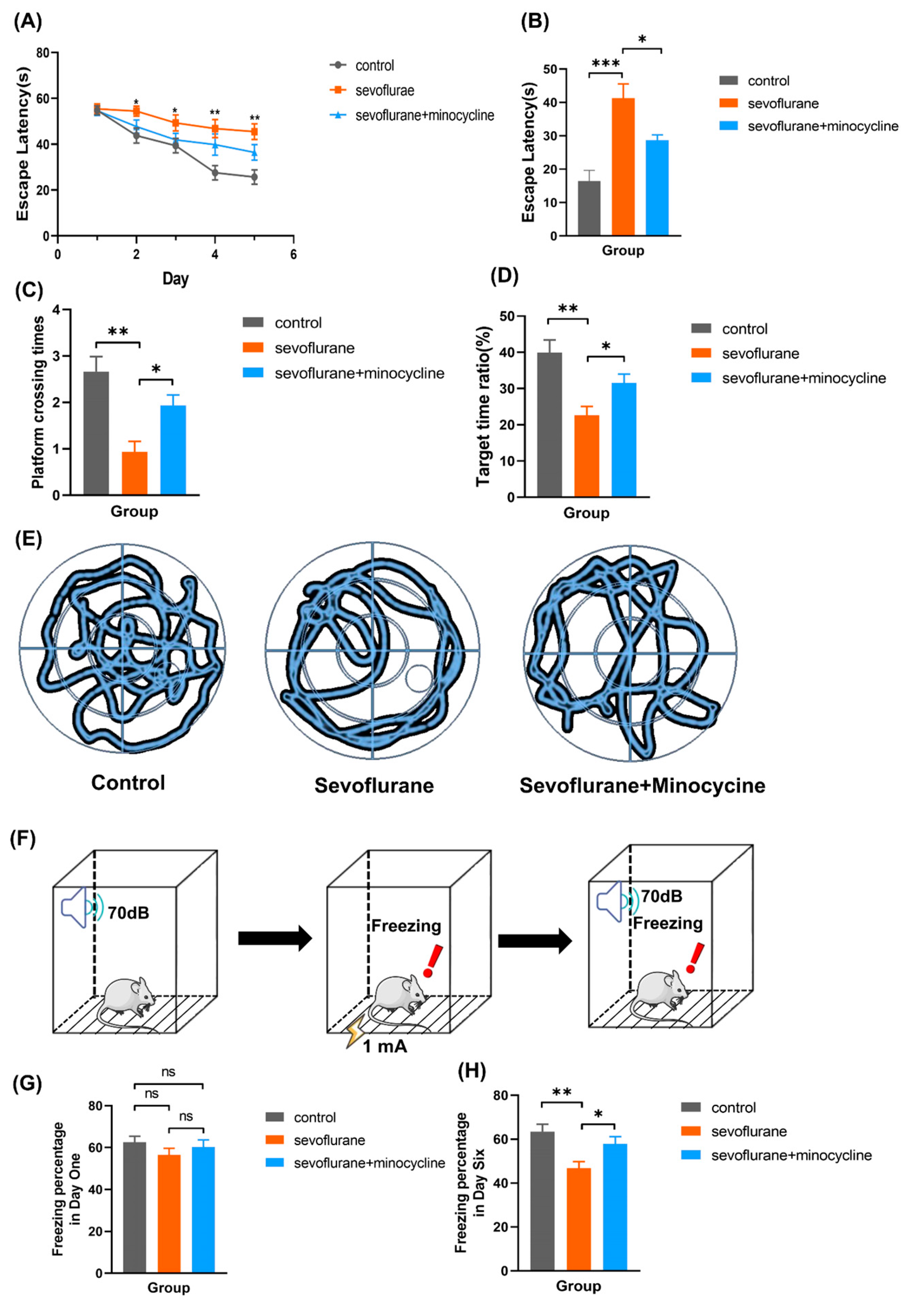
3.1. Minocycline Alleviates Sevoflurane-Induced Learning and Memory Impairments

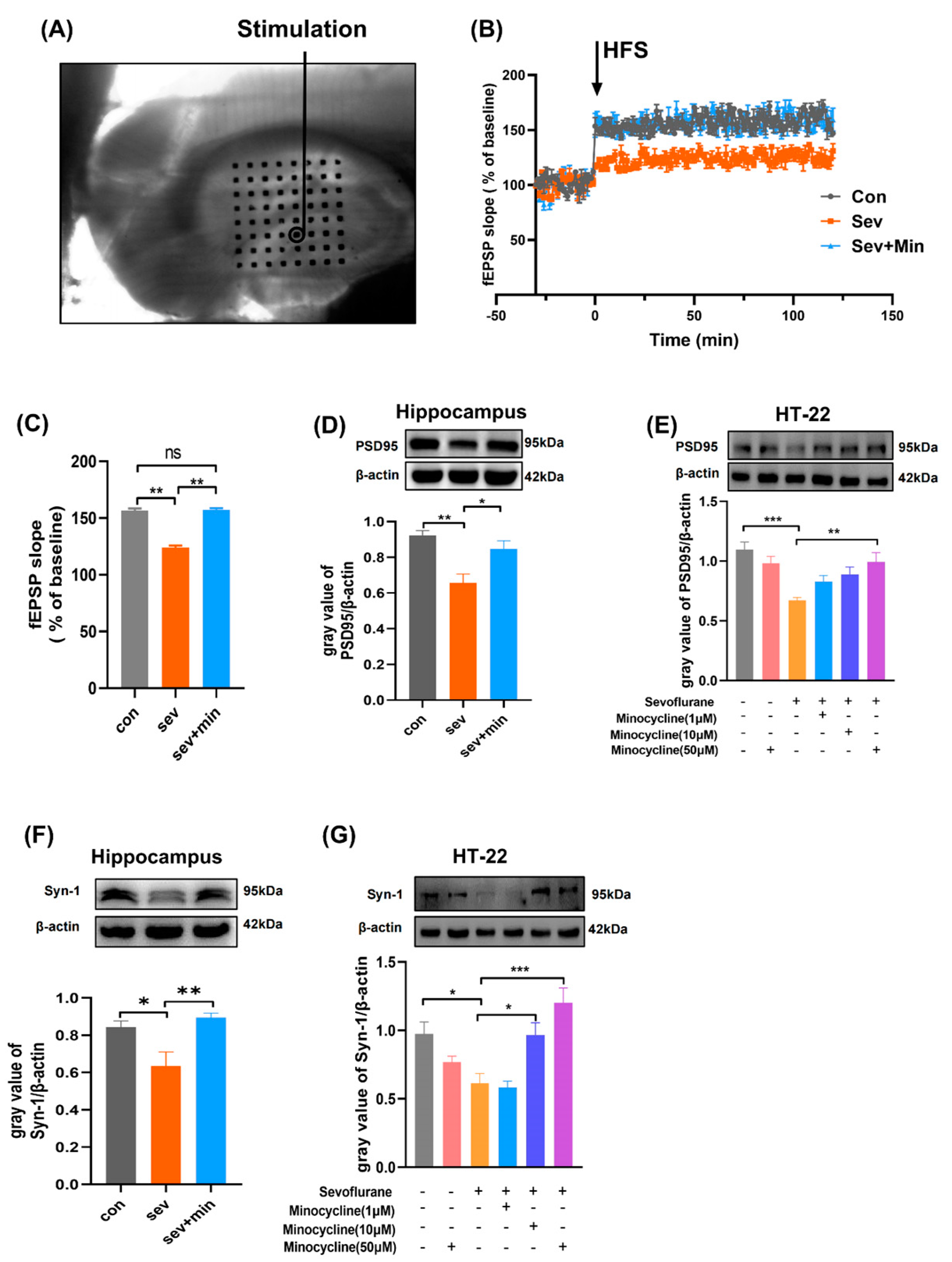
3.2. Minocycline Attenuates Sevoflurane-Induced Synaptic Plasticity Impairment
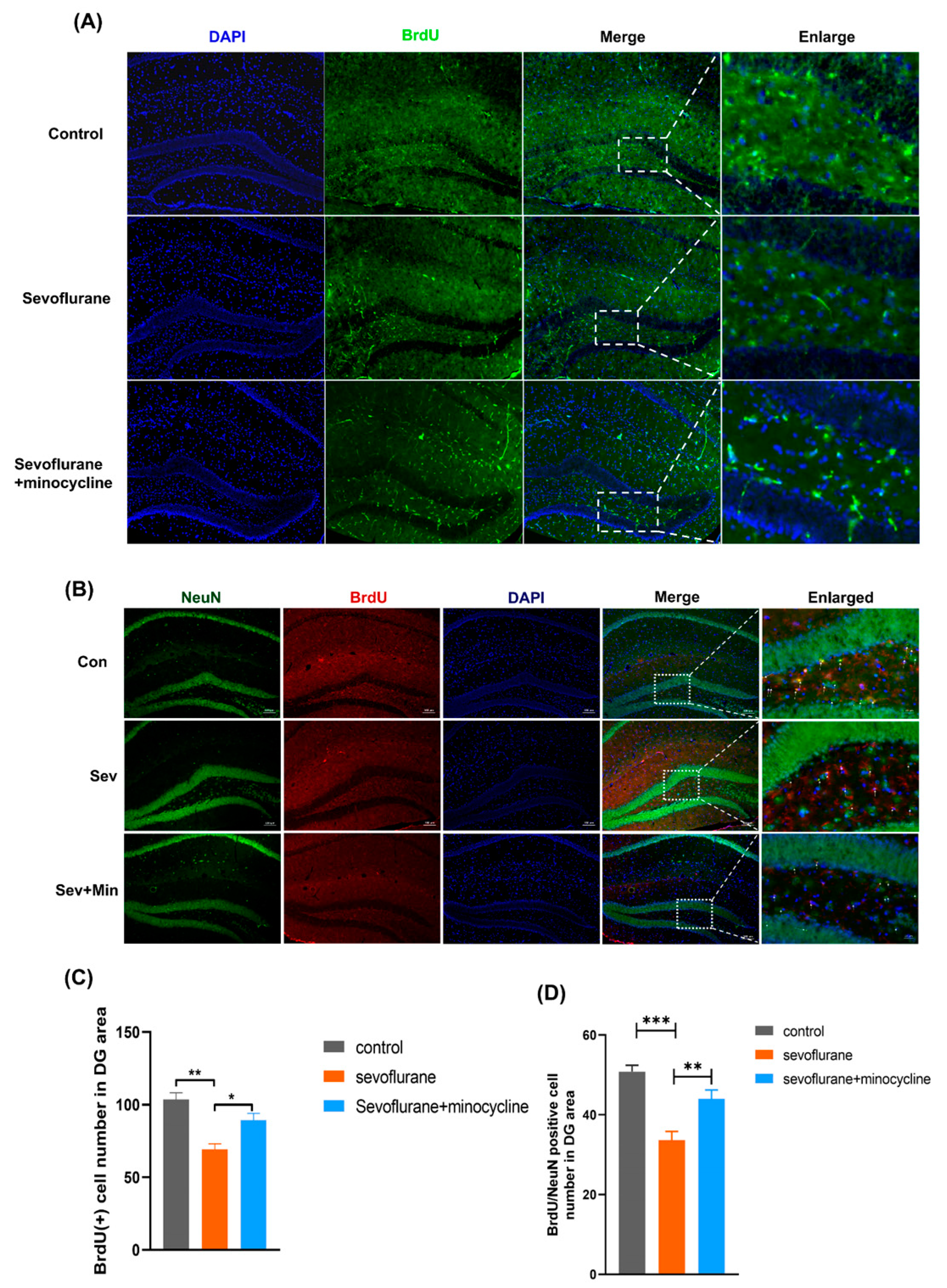
3.3. Minocycline Alleviates Sevoflurane-Induced Neurogenesis Dysfunction
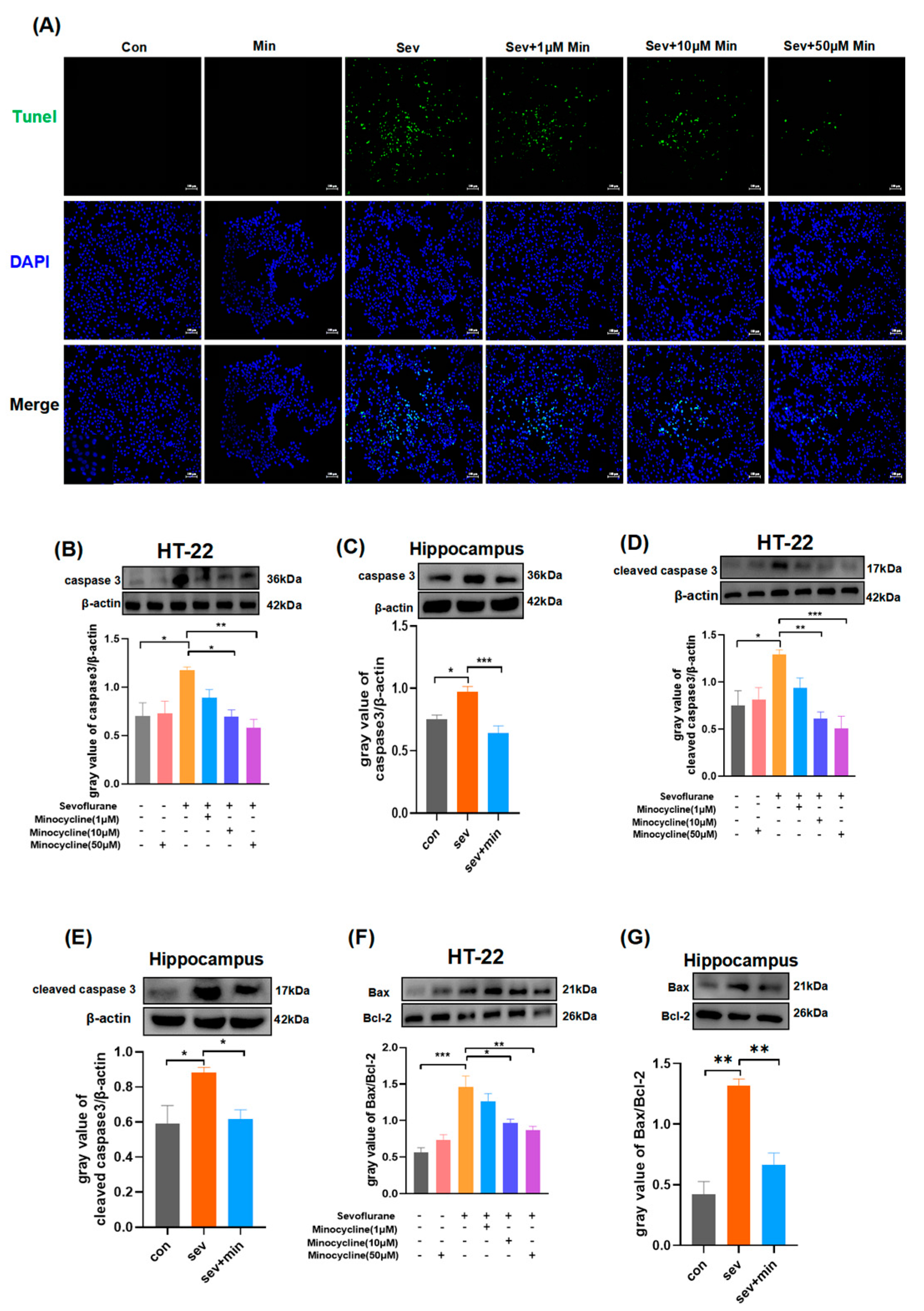
3.4. Minocycline Suppresses Hippocampal Apoptosis Induced by Sevoflurane
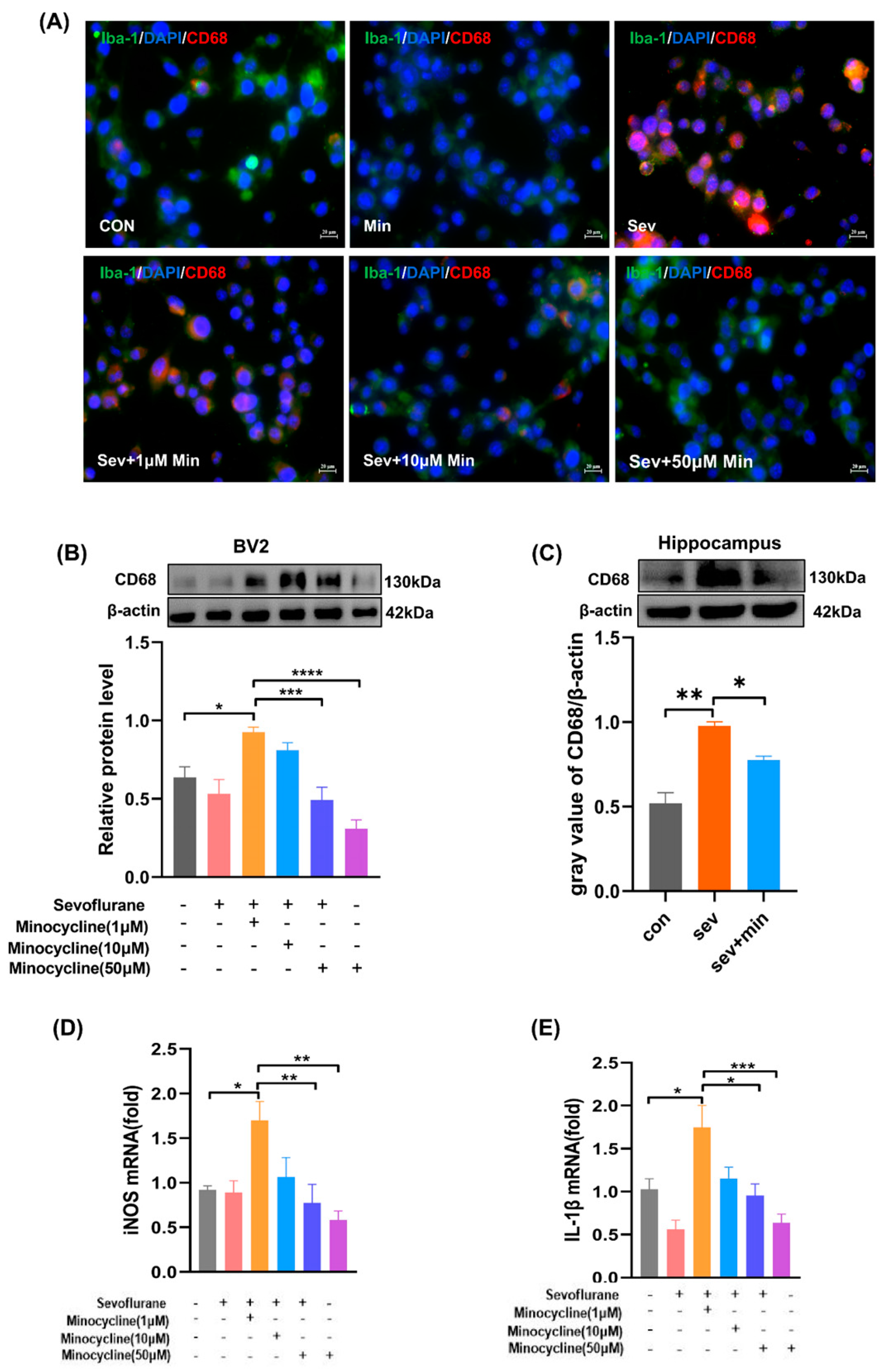
3.5. Minocycline Suppresses Sevoflurane-Associated Microglial Activation to the M1 Stage and Reduces Proinflammatory Cytokine Production
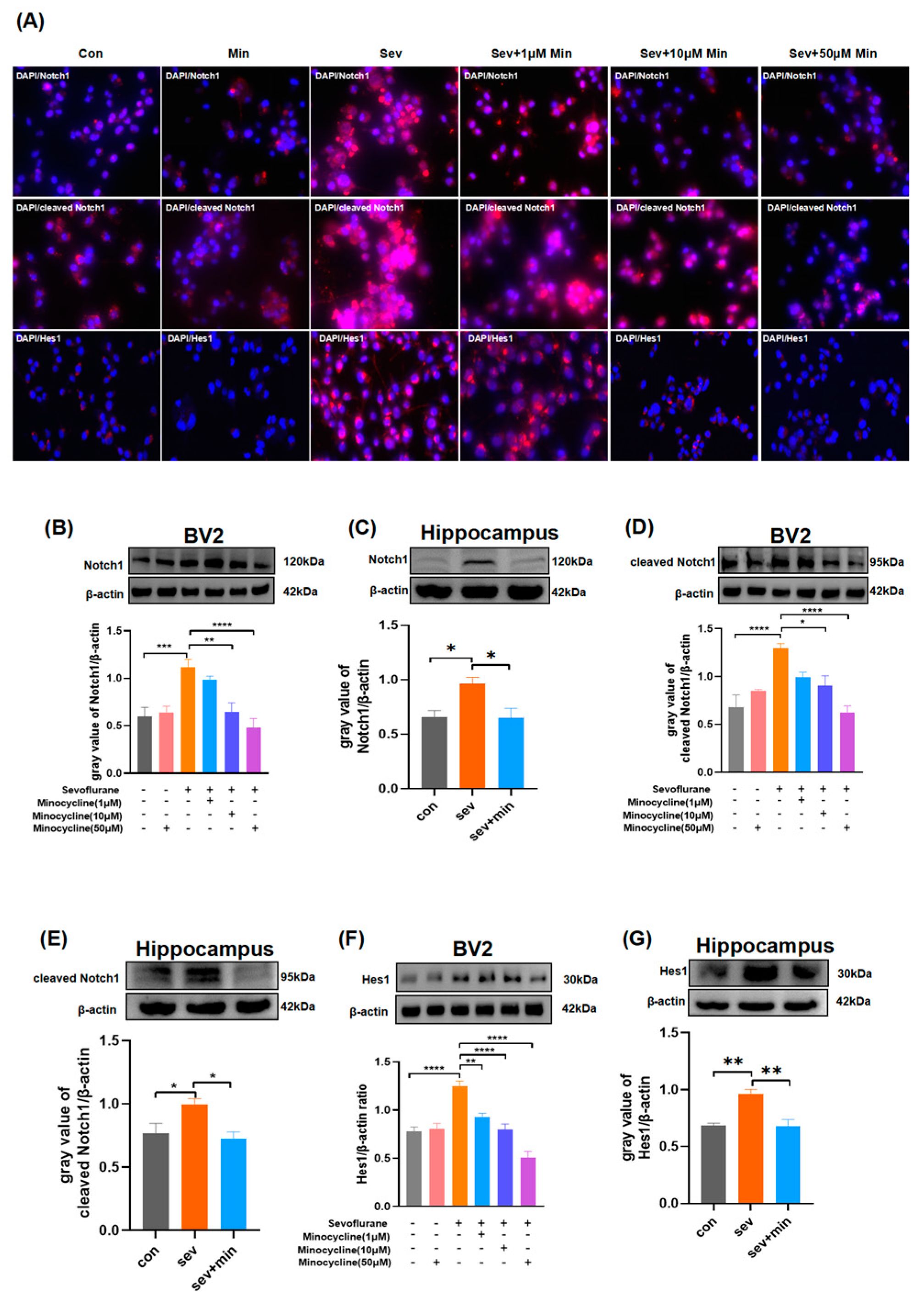
3.6. Minocycline Alleviates Sevoflurane-Induced Neuroinflammation via Notch Signaling Suppression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kotekar, N.; Shenkar, A.; Nagaraj, R. Postoperative cognitive dysfunction—Current preventive strategies. Clin. Interv. Aging 2018, 13, 2267–2273. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.; Lewis, S.R.; Pritchard, M.W.; Schofield-Robinson, O.J.; Shelton, C.L.; Alderson, P.; Smith, A.F. Intravenous versus inhalational maintenance of anaesthesia for postoperative cognitive outcomes in elderly people undergoing non-cardiac surgery. Cochrane Database Syst. Rev. 2018, 8, CD012317. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, P.; Lin, X.; Zhang, H.; Miao, J.; Zhou, Y.; Chen, G. Mitophagy impairment is involved in sevoflurane-induced cognitive dysfunction in aged rats. Aging 2020, 12, 17235–17256. [Google Scholar] [CrossRef]
- Chai, G.; Wu, J.; Fang, R.; Liu, Y.; Wang, X.; Wang, X.; Zhang, J.; Zhou, J.; Jiang, Z.; Yi, H.; et al. Sevoflurane inhibits histone acetylation and contributes to cognitive dysfunction by enhancing the expression of ANP32A in aging mice. Behav. Brain Res. 2022, 431, 113949. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yang, Y.; Tan, H.; Boukhali, M.; Khatri, A.; Yu, Y.; Hua, F.; Liu, L.; Li, M.; Yang, G.; et al. Tau Contributes to Sevoflurane-induced Neurocognitive Impairment in Neonatal Mice. Anesthesiology 2020, 133, 595–610. [Google Scholar] [CrossRef]
- Wang, X.; Dong, Y.; Zhang, Y.; Li, T.; Xie, Z. Sevoflurane induces cognitive impairment in young mice via autophagy. PLoS ONE 2019, 14, e0216372. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.H.; Zhang, X.R.; He, L.; Zhu, J.; Ma, Q.J. Effect of sevoflurane and halothane anesthesia on cognitive function and immune function in young rats. Saudi J. Biol. Sci. 2018, 25, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Liang, F.; Huang, L.; Fang, F.; Yang, G.; Tanzi, R.E.; Zhang, Y.; Quan, Q.; Xie, Z. The anesthetic sevoflurane induces tau trafficking from neurons to microglia. Commun. Biol. 2021, 4, 560. [Google Scholar] [CrossRef]
- Li, G.; Wang, Y.; Cao, F.; Wang, D.; Zhou, L.; Jin, Y. Sevoflurane Promotes Neurodegeneration Through Inflammasome Formation in APP/PS1 Mice. Front. Neurosci. 2021, 15, 647136. [Google Scholar] [CrossRef]
- Gui, L.; Lei, X.; Zuo, Z. Decrease of glial cell-derived neurotrophic factor contributes to anesthesia- and surgery-induced learning and memory dysfunction in neonatal rats. J. Mol. Med. 2017, 95, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, Y.; Xu, M.; Dai, Q.; Meng, W.; Yang, J.; Qin, R. Activation of Notch signal pathway is associated with a poorer prognosis in acute myeloid leukemia. Med. Oncol. 2011, 28 (Suppl. S1), S483–S489. [Google Scholar] [CrossRef]
- Tindemans, I.; van Schoonhoven, A.; KleinJan, A.; de Bruijn, M.J.; Lukkes, M.; van Nimwegen, M.; van den Branden, A.; Bergen, I.M.; Corneth, O.B.; van IJcken, W.F.; et al. Notch signaling licenses allergic airway inflammation by promoting Th2 cell lymph node egress. J. Clin. Investig. 2020, 130, 3576–3591. [Google Scholar] [CrossRef] [Green Version]
- Lavoz, C.; Poveda, J.; Marquez-Exposito, L.; Rayego-Mateos, S.; Rodrigues-Diez, R.R.; Ortiz, A.; Egido, J.; Mezzano, S.; Ruiz-Ortega, M. Gremlin activates the Notch pathway linked to renal inflammation. Clin. Sci. 2018, 132, 1097–1115. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Imagama, S.; Ohgomori, T.; Hirano, K.; Uchimura, K.; Sakamoto, K.; Hirakawa, A.; Takeuchi, H.; Suzumura, A.; Ishiguro, N.; et al. Minocycline selectively inhibits M1 polarization of microglia. Cell Death Dis. 2013, 4, e525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, J.; Mo, H.; Chen, J.; Qian, C.; Yan, F.; Gu, C.; Hu, Q.; Wang, L.; Chen, G. Minocycline Protects Against NLRP3 Inflammasome-Induced Inflammation and P53-Associated Apoptosis in Early Brain Injury After Subarachnoid Hemorrhage. Mol. Neurobiol. 2016, 53, 2668–2678. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Guo, S.; Wu, X.; Ma, L.; Zhao, X. Minocycline alleviates sevoflurane-induced cognitive impairment in aged rats. Cell Mol. Neurobiol. 2015, 35, 585–594. [Google Scholar] [CrossRef]
- Yang, C.; Gao, J.; Wu, B.; Yan, N.; Li, H.; Ren, Y.; Kan, Y.; Liang, J.; Jiao, Y.; Yu, Y. Minocycline attenuates the development of diabetic neuropathy by inhibiting spinal cord Notch signaling in rat. Biomed. Pharmacother. 2017, 94, 380–385. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, M.; Li, Y.; Li, Y.; Hua, Y.; Fan, Y. Minocycline promotes functional recovery in ischemic stroke by modulating microglia polarization through STAT1/STAT6 pathways. Biochem. Pharmacol. 2021, 186, 114464. [Google Scholar] [CrossRef]
- Dai, X.J.; Li, N.; Yu, L.; Chen, Z.-Y.; Hua, R.; Qin, X. Activation of BV2 microglia by lipopolysaccharide triggers an inflammatory reaction in PC12 cell apoptosis through a toll-like receptor 4-dependent pathway. Cell Stress Chaperones 2015, 20, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Shen, J.; Yu, L.; Sun, J.; McQuillan, P.M.; Hu, Z.; Yan, M. Role of autophagy in sevoflurane-induced neurotoxicity in neonatal rat hippocampal cells. Brain Res. Bull. 2018, 140, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xue, B.; You, J.; Zhang, B.; Chai, G. Suberoylanilide hydroxamic acid reversed cognitive and synaptic plasticity impairments induced by sevoflurane exposure in adult mice. Neuroreport 2019, 30, 274–279. [Google Scholar] [CrossRef]
- Zhao, X.P.; Zhong, F.; Luo, R.Y.; Zhang, Y.L.; Luo, C.; Li, H.; Dai, R.P. Early-life sevoflurane exposure impairs fear memory by suppressing extracellular signal-regulated kinase signaling in the bed nucleus of stria terminalis GABAergic neurons. Neuropharmacology 2021, 191, 108584. [Google Scholar] [CrossRef] [PubMed]
- Ju, L.S.; Jia, M.; Sun, J.; Sun, X.R.; Zhang, H.; Ji, M.H.; Yang, J.J.; Wang, Z.Y. Hypermethylation of Hippocampal Synaptic Plasticity-Related genes is Involved in Neonatal Sevoflurane Exposure-Induced Cognitive Impairments in Rats. Neurotox. Res. 2016, 29, 243–255. [Google Scholar] [CrossRef]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Zhu, J.; Yang, Q.; Wang, Y.; Zhang, Z.; Chen, C. The Role of Histone Acetylation in the Sevoflurane-induced Inhibition of Neurogenesis in the Hippocampi of Young Mice. Neuroscience 2020, 432, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Guo, Y.; Wang, G.; Sun, G.; Sun, W.; Li, J.; Li, X.; Wu, J.; Zhang, M. Inhibition of Adult Hippocampal Neurogenesis Plays a Role in Sevoflurane-Induced Cognitive Impairment in Aged Mice Through Brain-Derived Neurotrophic Factor/Tyrosine Receptor Kinase B and Neurotrophin-3/Tropomyosin Receptor Kinase C Pathways. Front. Aging Neurosci. 2022, 14, 782932. [Google Scholar] [CrossRef] [PubMed]
- Stefanis, L.; Burke, R.E.; Greene, L.A. Apoptosis in neurodegenerative disorders. Curr. Opin. Neurol. 1997, 10, 299–305. [Google Scholar] [CrossRef]
- Kang, W.; Lu, D.; Yang, X.; Ma, W.; Chen, X.; Chen, K.; Xu, X.; Zhou, X.; Zhou, L.; Feng, X. Sevoflurane Induces Hippocampal Neuronal Apoptosis by Altering the Level of Neuropeptide Y in Neonatal Rats. Neurochem. Res. 2020, 45, 1986–1996. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Xu, M.; Chen, G. Autophagy Is Involved in the Sevoflurane Anesthesia-Induced Cognitive Dysfunction of Aged Rats. PLoS ONE 2016, 11, e0153505. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Mesa, N.; Zarzuelo, A.; Gálvez, J. Minocycline: Far beyond an antibiotic. Br. J. Pharmacol. 2013, 169, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Qian, D.; Li, L.; Rong, Y.; Liu, W.; Wang, Q.; Zhou, Z.; Gu, C.; Huang, Y.; Zhao, X.; Chen, J.; et al. Blocking Notch signal pathway suppresses the activation of neurotoxic A1 astrocytes after spinal cord injury. Cell Cycle 2019, 18, 3010–3029. [Google Scholar] [CrossRef]
- Wu, L.; Li, Y.; Yu, M.; Yang, F.; Tu, M.; Xu, H. Notch Signaling Regulates Microglial Activation and Inflammatory Reactions in a Rat Model of Temporal Lobe Epilepsy. Neurochem. Res. 2018, 43, 1269–1282. [Google Scholar] [CrossRef]
- Bedford, P.D. Adverse cerebral effects of anaesthesia on old people. Lancet 1955, 269, 259–263. [Google Scholar] [CrossRef]
- Boone, M.D.; Sites, B.; von Recklinghausen, F.M.; Mueller, A.; Taenzer, A.H.; Shaefi, S. Economic Burden of Postoperative Neurocognitive Disorders Among US Medicare Patients. JAMA Netw. Open 2020, 3, e208931. [Google Scholar] [CrossRef]
- Evered, L.A.; Silbert, B.S.; Scott, D.A.; Maruff, P.; Ames, D. Prevalence of Dementia 7.5 Years after Coronary Artery Bypass Graft Surgery. Anesthesiology 2016, 125, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Relander, K.; Hietanen, M.; Rantanen, K.; Rämö, J.; Vento, A.; Saastamoinen, K.P.; Roine, R.O.; Soinne, L. Postoperative cognitive change after cardiac surgery predicts long-term cognitive outcome. Brain Behav. 2020, 10, e01750. [Google Scholar] [CrossRef]
- Kitsis, P.; Zisimou, T.; Gkiatas, I.; Kostas-Agnantis, I.; Gelalis, I.; Korompilias, A.; Pakos, E. Postoperative Delirium and Postoperative Cognitive Dysfunction in Patients with Elective Hip or Knee Arthroplasty: A Narrative Review of the Literature. Life 2022, 12, 314. [Google Scholar] [CrossRef]
- Galinkin, J.L.; Janiszewski, D.; Young, C.J.; Klafta, J.M.; Klock, P.A.; Coalson, D.W.; Apfelbaum, J.L.; Zacny, J.P. Subjective, psychomotor, cognitive, and analgesic effects of subanesthetic concentrations of sevoflurane and nitrous oxide. Anesthesiology 1997, 87, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Ou, S.; Han, Y.; Xu, J.; Zhou, S. Plasma amyloid beta level changes in aged mice with cognitive dysfunction following sevoflurane exposure. Exp. Gerontol. 2020, 129, 110737. [Google Scholar] [CrossRef] [PubMed]
- Shimohama, S. Apoptosis in Alzheimer’s disease—An update. Apoptosis 2000, 5, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Lev, N.; Melamed, E.; Offen, D. Apoptosis and Parkinson’s disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 245–250. [Google Scholar] [CrossRef]
- Sathasivam, S.; Ince, P.G.; Shaw, P.J. Apoptosis in amyotrophic lateral sclerosis: A review of the evidence. Neuropathol. Appl. Neurobiol. 2001, 27, 257–274. [Google Scholar] [CrossRef]
- Chen, G.; Gong, M.; Yan, M.; Zhang, X. Sevoflurane induces endoplasmic reticulum stress mediated apoptosis in hippocampal neurons of aging rats. PLoS ONE 2013, 8, e57870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Dong, Y.; Xu, Z.; Wang, H.; Miao, C.; Soriano, S.G.; Sun, D.; Baxter, M.G.; Zhang, Y.; Xie, Z. Selective anesthesia-induced neuroinflammation in developing mouse brain and cognitive impairment. Anesthesiology 2013, 118, 502–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, P.; Zhao, J.; Li, N.; Lu, L.; Li, L.; Zhang, X.; Yang, B.; Zhang, L.; Li, D. Sevoflurane exaggerates cognitive decline in a rat model of chronic intermittent hypoxia by aggravating microglia-mediated neuroinflammation via downregulation of PPAR-γ in the hippocampus. Behav. Brain Res. 2018, 347, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Siebel, C.; Lendahl, U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wei, X.; Mei, Y.; Wang, D.; Wang, J.; Zhang, Y.; Li, X.; Gu, Y.; Peng, G.; Sun, B. Modulating adult neurogenesis affects synaptic plasticity and cognitive functions in mouse models of Alzheimer’s disease. Stem Cell Rep. 2021, 16, 3005–3019. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Miao, Z.; Ye, Y.; Zhao, P.; Fan, L.; Bao, Z.; Tu, Y.; Li, C.; Chao, H.; Xu, X.; et al. Curcumin alleviates neuroinflammation, enhances hippocampal neurogenesis, and improves spatial memory after traumatic brain injury. Brain Res. Bull. 2020, 162, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Naderi, Y.; Sabetkasaei, M.; Parvardeh, S.; Zanjani, T.M. Neuroprotective effect of minocycline on cognitive impairments induced by transient cerebral ischemia/reperfusion through its anti-inflammatory and anti-oxidant properties in male rat. Brain Res. Bull. 2017, 131, 207–213. [Google Scholar] [CrossRef]
- Takazawa, T.; Horiuchi, T.; Orihara, M.; Nagumo, K.; Tomioka, A.; Ideno, Y.; Hayashi, K.; Yashima, H.; Araki, T.; Hatayama, K.; et al. Prevention of Postoperative Cognitive Dysfunction by Minocycline in Elderly Patients after Total Knee Arthroplasty: A Randomized, Double-blind, Placebo-controlled Clinical Trial. Anesthesiology 2023, 138, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Oberman, K.; Hovens, I.; de Haan, J.; Falcao-Salles, J.; van Leeuwen, B.; Schoemaker, R. Acute pre-operative ibuprofen improves cognition in a rat model for postoperative cognitive dysfunction. J. Neuroinflamm. 2021, 18, 156. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wu, X.; Guo, S.; Ma, L.; Huang, W.; Zhao, X. Minocycline attenuates sevoflurane-induced cell injury via activation of Nrf2. Int. J. Mol. Med. 2017, 39, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Z.; Li, J.; Miao, L.; Huang, Z.; Huang, W.; Liu, Y.; Li, Y. Inhibition of RhoA Activity Does Not Rescue Synaptic Development Abnormalities and Long-Term Cognitive Impairment After Sevoflurane Exposure. Neurochem. Res. 2021, 46, 468–481. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Satomoto, M.; Adachi, Y.U.; Kinoshita, H.; Makita, K. Inhibiting NADPH oxidase protects against long-term memory impairment induced by neonatal sevoflurane exposure in mice. Br. J. Anaesth. 2016, 117, 80–86. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Catalog Number | Dilution | Specificity |
|---|---|---|---|---|
| Iba-1 | Thermofisher scientific | PA5-27436 | 1:150~1:200 | Mouse |
| CD68 | CST | 97778S | 1:150~1:200 | Rabbit |
| BrdU | Abcam | Ab8955 | 1:150~1:200 | Mouse |
| NeuN | Abcam | Ab177487 | 1:150~1:200 | Rabbit |
| Notch1 | Proteintech | 20687-1-AP | 1:150~1:200 | Rabbit |
| Cleaved Notch1 | Affinity Bioscience | AF5307 | 1:150~1:200 | Rabbit |
| Hes1 | Affinity Bioscience | DF7569 | 1:150~1:200 | Rabbit |
| Antibody | Source | Catalog Number | Dilution | Specificity |
|---|---|---|---|---|
| Caspase3 | Affinity Biosciences | AF6311 | 1:500 | Rabbit |
| Cleaved caspase3 | Affinity Biosciences | AF7022 | 1:500 | Rabbit |
| Bcl-2 | Affinity Biosciences | AF6139 | 1:500 | Rabbit |
| Bax | Affinity Biosciences | AF0120 | 1:500 | Rabbit |
| PSD95 | Proteintech | 20665-1-AP | 1:500 | Rabbit |
| Syn-1 | Proteintech | 20258-1-AP | 1:500 | Rabbit |
| Notch1 | Proteintech | 20687-1-AP | 1:500 | Rabbit |
| Cleaved Notch1 | Affinity Biosciences | AF5307 | 1:500 | Rabbit |
| Hes1 | Affinity Biosciences | DF7569 | 1:500 | Rabbit |
| β-actin | Affinity Biosciences | AF7018 | 1:1000 | Rabbit |
| Primer | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| iNOS | CCCTTCAATGGTTTACATGG | ACATGATCTCCGTGACAGCC |
| IL-1β | ACTCATTGTGGCTGTGGAGA | TTGTTCATCTCGGAGCCTGT |
| GAPDH | AACGACCCCTTCATTGACCT | TGGAAGATGGTGATGGGCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, J.; Han, S.; Ye, C.; Zhu, H.; Wu, J.; Nie, Y.; Chai, G.; Zhao, P.; Zhang, D. Minocycline Attenuates Sevoflurane-Induced Postoperative Cognitive Dysfunction in Aged Mice by Suppressing Hippocampal Apoptosis and the Notch Signaling Pathway-Mediated Neuroinflammation. Brain Sci. 2023, 13, 512. https://doi.org/10.3390/brainsci13030512
Liang J, Han S, Ye C, Zhu H, Wu J, Nie Y, Chai G, Zhao P, Zhang D. Minocycline Attenuates Sevoflurane-Induced Postoperative Cognitive Dysfunction in Aged Mice by Suppressing Hippocampal Apoptosis and the Notch Signaling Pathway-Mediated Neuroinflammation. Brain Sciences. 2023; 13(3):512. https://doi.org/10.3390/brainsci13030512
Chicago/Turabian StyleLiang, Junjie, Shanshan Han, Chao Ye, Haimeng Zhu, Jiajun Wu, Yunjuan Nie, Gaoshang Chai, Peng Zhao, and Dengxin Zhang. 2023. "Minocycline Attenuates Sevoflurane-Induced Postoperative Cognitive Dysfunction in Aged Mice by Suppressing Hippocampal Apoptosis and the Notch Signaling Pathway-Mediated Neuroinflammation" Brain Sciences 13, no. 3: 512. https://doi.org/10.3390/brainsci13030512