
TMS-Induced Modulation of EEG Functional Connectivity Is Affected by the E-Field Orientation
 , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
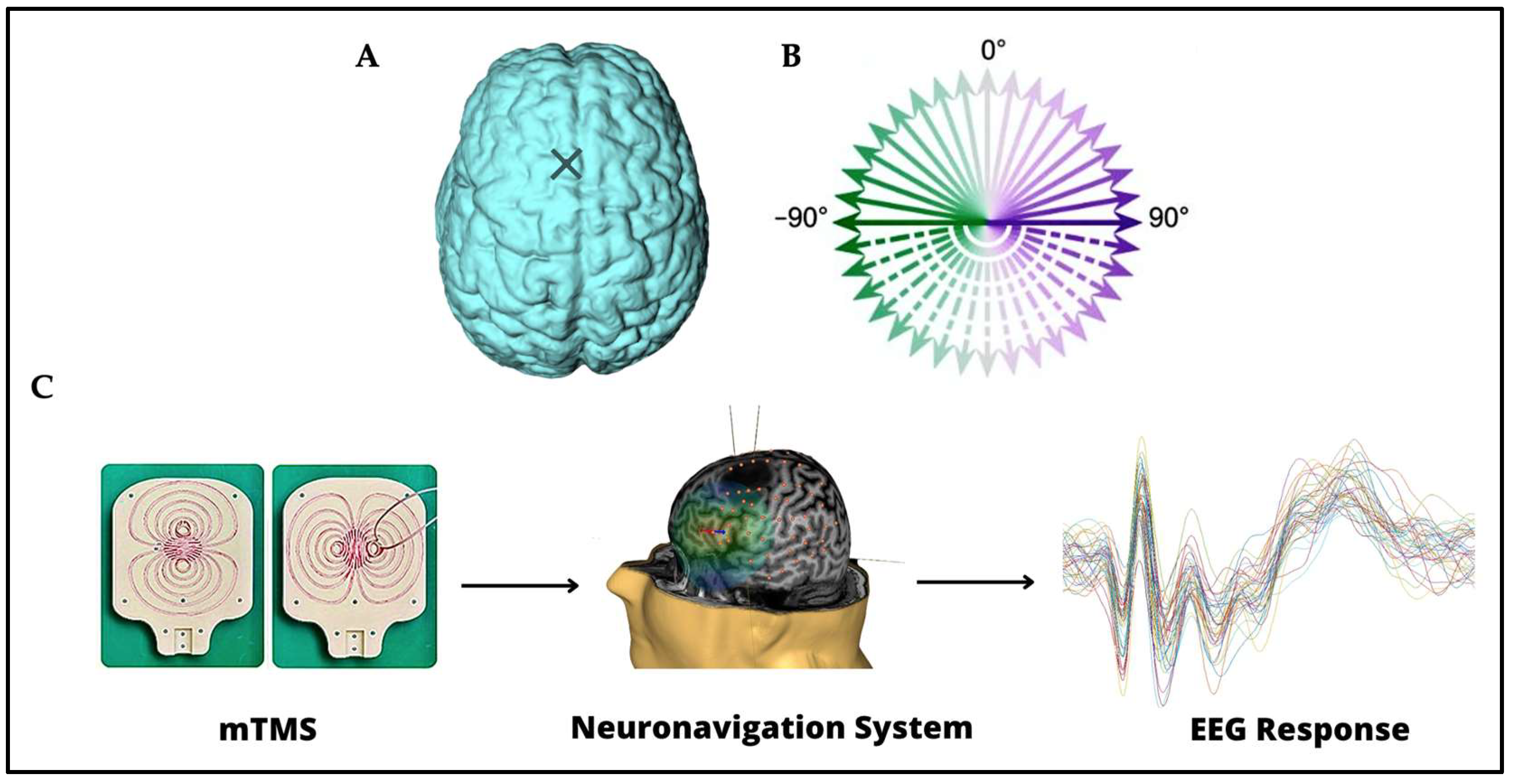
2.2. The mTMS–EEG Setup
2.3. Procedure
3. Data analysis
3.1. Preprocessing
3.2. Functional Connectivity Estimation
3.3. Statistical Assessment
4. Results
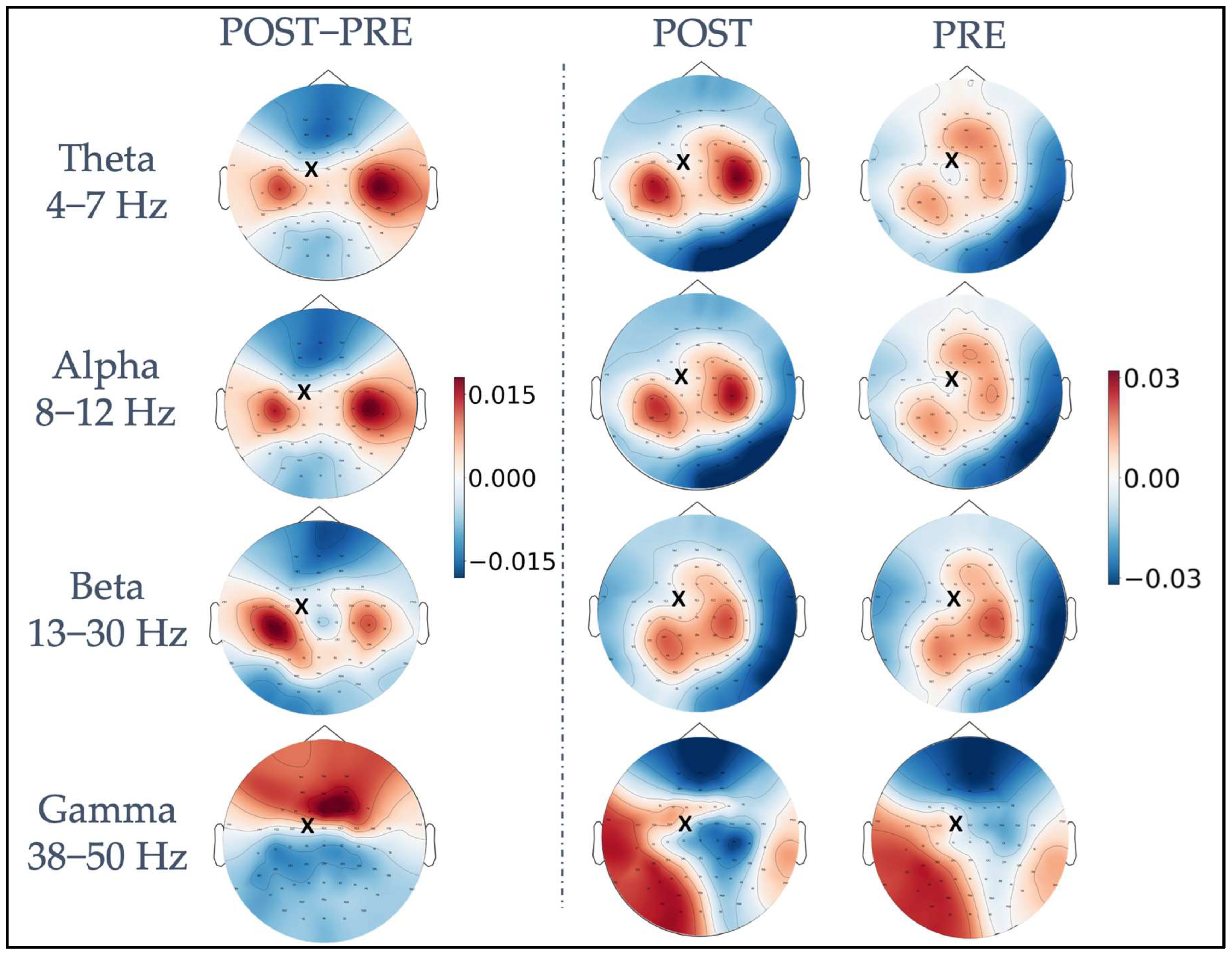
4.1. iPLV Modulation with Respect to the TMS
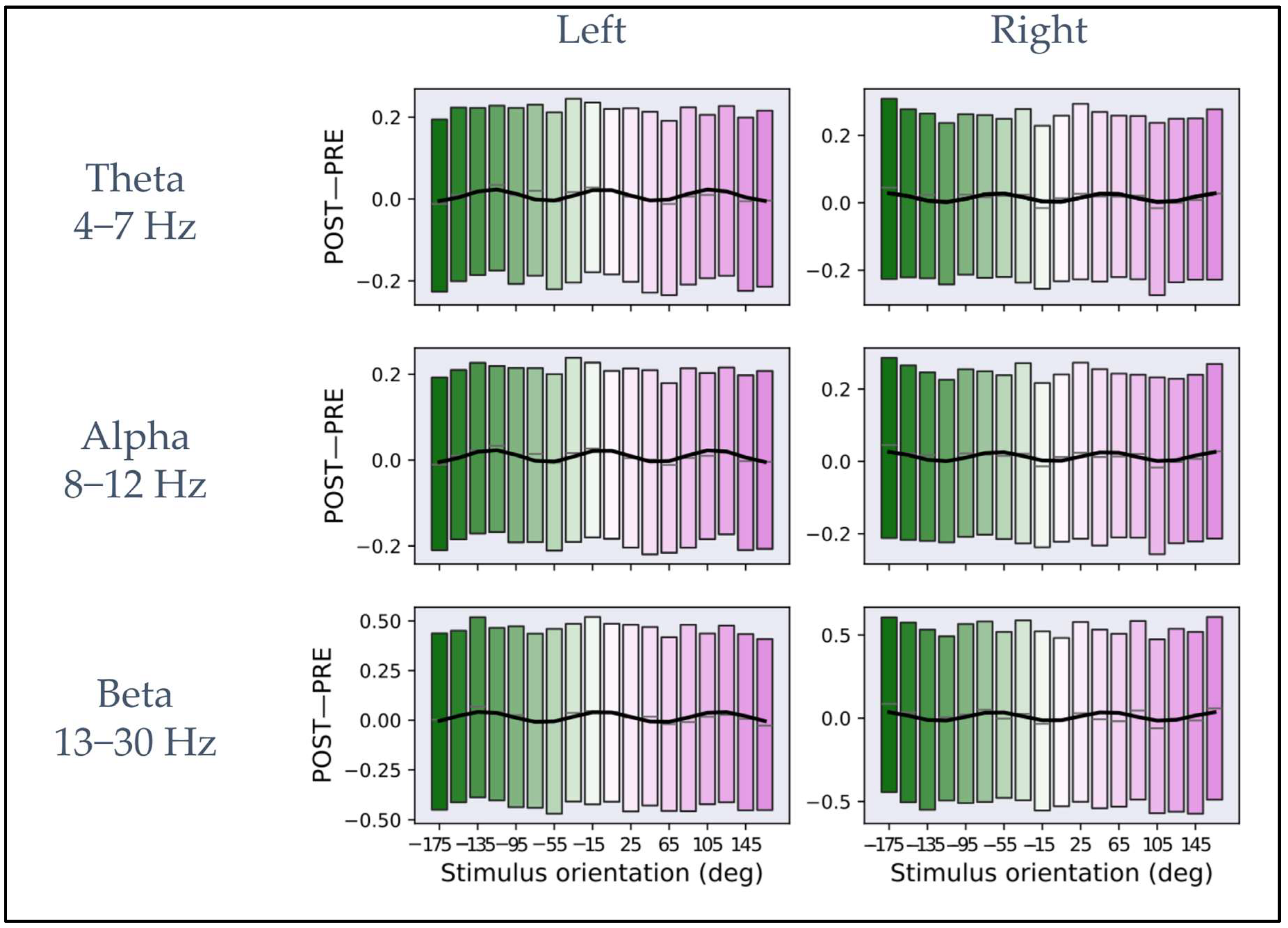
4.2. Functional Connectivity Depends on the Stimulus Orientation
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vlachos, I.; Tzirini, M.; Chatzikyriakou, E.; Markakis, I.; Rouni, M.A.; Samaras, T.; Roth, Y.; Zangen, A.; Rotenberg, A.; Kugiumtzis, D.; et al. The Relation between Induced Electric Field and TMS-Evoked Potentials: A Deep TMS-EEG Study. Appl. Sci. 2022, 12, 7437. [Google Scholar] [CrossRef]
- Lefaucheur, J.-P.; Aleman, A.; Baeken, C.; Benninger, D.H.; Brunelin, J.; Di Lazzaro, V.; Filipović, S.R.; Grefkes, C.; Hasan, A.; Hummel, F.C.; et al. Evidence-Based Guidelines on the Therapeutic Use of Repetitive Transcranial Magnetic Stimulation (RTMS): An Update (2014-2018). Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2020, 131, 474–528. [Google Scholar] [CrossRef]
- Hoogendam, J.M.; Ramakers, G.M.J.; Di Lazzaro, V. Physiology of Repetitive Transcranial Magnetic Stimulation of the Human Brain. Brain Stimulat. 2010, 3, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Hampson, M.; Hoffman, R. Transcranial Magnetic Stimulation and Connectivity Mapping: Tools for Studying the Neural Bases of Brain Disorders. Front. Syst. Neurosci. 2010, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Bolognini, N.; Ro, T. Transcranial Magnetic Stimulation: Disrupting Neural Activity to Alter and Assess Brain Function. J. Neurosci. 2010, 30, 9647–9650. [Google Scholar] [CrossRef] [Green Version]
- Opitz, A.; Fox, M.D.; Craddock, R.C.; Colcombe, S.; Milham, M.P. An Integrated Framework For Targeting Functional Networks via Transcranial Magnetic Stimulation. NeuroImage 2016, 127, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Casarotto, S.; Lauro, L.J.R.; Bellina, V.; Casali, A.G.; Rosanova, M.; Pigorini, A.; Defendi, S.; Mariotti, M.; Massimini, M. EEG Responses to TMS Are Sensitive to Changes in the Perturbation Parameters and Repeatable over Time. PLoS ONE 2010, 5, e10281. [Google Scholar] [CrossRef] [Green Version]
- Hannula, H.; Ilmoniemi, R.J. Basic Principles of Navigated TMS. In Navigated Transcranial Magnetic Stimulation in Neurosurgery; Springer: Cham, Switzerland, 2017; pp. 3–29. [Google Scholar] [CrossRef]
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-Invasive Magnetic Stimulation of Human Motor Cortex. Lancet Lond. Engl. 1985, 1, 1106–1107. [Google Scholar] [CrossRef]
- Lioumis, P.; Rosanova, M. The Role of Neuronavigation in TMS–EEG Studies: Current Applications and Future Perspectives. J. Neurosci. Methods 2022, 380, 109677. [Google Scholar] [CrossRef]
- Caulfield, K.A.; Fleischmann, H.H.; Cox, C.E.; Wolf, J.P.; George, M.S.; McTeague, L.M. Neuronavigation Maximizes Accuracy and Precision in TMS Positioning: Evidence from 11,230 Distance, Angle, and Electric Field Modeling Measurements. Brain Stimulat. 2022, 15, 1192–1205. [Google Scholar] [CrossRef]
- Nieminen, A.E.; Nieminen, J.O.; Stenroos, M.; Novikov, P.; Nazarova, M.; Vaalto, S.; Nikulin, V.; Ilmoniemi, R.J. Accuracy and Precision of Navigated Transcranial Magnetic Stimulation. J. Neural Eng. 2022, 19, 066037. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M. Transcranial Magnetic Stimulation and the Human Brain. Nature 2000, 406, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Krieg, S.M.; Lioumis, P.; Mäkelä, J.P.; Wilenius, J.; Karhu, J.; Hannula, H.; Savolainen, P.; Lucas, C.W.; Seidel, K.; Laakso, A.; et al. Protocol for Motor and Language Mapping by Navigated TMS in Patients and Healthy Volunteers; Workshop Report. Acta Neurochir. 2017, 159, 1187–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caulfield, K.A.; Brown, J.C. The Problem and Potential of TMS’ Infinite Parameter Space: A Targeted Review and Road Map Forward. Front. Psychiatry 2022, 13, 867091. [Google Scholar] [CrossRef]
- Mills, K.R.; Boniface, S.J.; Schubert, M. Magnetic Brain Stimulation with a Double Coil: The Importance of Coil Orientation. Electroencephalogr. Clin. Neurophysiol. Potentials Sect. 1992, 85, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Herwig, U.; Satrapi, P.; Schönfeldt-Lecuona, C. Using the International 10-20 EEG System for Positioning of Transcranial Magnetic Stimulation. Brain Topogr. 2003, 16, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Rossini, P.M. TMS and TMS-EEG Techniques in the Study of the Excitability, Connectivity, and Plasticity of the Human Motor Cortex. Rev. Neurosci. 2013, 24, 431–442. [Google Scholar] [CrossRef]
- Ilmoniemi, R.J.; Kičić, D. Methodology for Combined TMS and EEG. Brain Topogr. 2010, 22, 233–248. [Google Scholar] [CrossRef] [Green Version]
- Rosanova, M.; Casali, A.; Bellina, V.; Resta, F.; Mariotti, M.; Massimini, M. Natural Frequencies of Human Corticothalamic Circuits. J. Neurosci. 2009, 29, 7679–7685. [Google Scholar] [CrossRef]
- Casarotto, S.; Fecchio, M.; Rosanova, M.; Varone, G.; D’Ambrosio, S.; Sarasso, S.; Pigorini, A.; Russo, S.; Comanducci, A.; Ilmoniemi, R.J.; et al. The Rt-TEP Tool: Real-Time Visualization of TMS-Evoked Potentials to Maximize Cortical Activation and Minimize Artifacts. J. Neurosci. Methods 2022, 370, 109486. [Google Scholar] [CrossRef]
- Casarotto, S.; Määttä, S.; Herukka, S.-K.; Pigorini, A.; Napolitani, M.; Gosseries, O.; Niskanen, E.; Könönen, M.; Mervaala, E.; Rosanova, M.; et al. Transcranial Magnetic Stimulation-Evoked EEG/Cortical Potentials in Physiological and Pathological Aging. Neuroreport 2011, 22, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, S.; Rogasch, N.C.; Premoli, I.; Blumberger, D.M.; Casarotto, S.; Chen, R.; Di Lazzaro, V.; Farzan, F.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Clinical Utility and Prospective of TMS-EEG. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2019, 130, 802–844. [Google Scholar] [CrossRef] [PubMed]
- Koponen, L.M.; Nieminen, J.O.; Ilmoniemi, R.J. Multi-Locus Transcranial Magnetic Stimulation—Theory and Implementation. Brain Stimulat. 2018, 11, 849–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, V.H.; Nieminen, J.O.; Tugin, S.; Koponen, L.M.; Baffa, O.; Ilmoniemi, R.J. TMS with Fast and Accurate Electronic Control: Measuring the Orientation Sensitivity of Corticomotor Pathways. Brain Stimulat. 2022, 15, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, J.O.; Sinisalo, H.; Souza, V.H.; Malmi, M.; Yuryev, M.; Tervo, A.E.; Stenroos, M.; Milardovich, D.; Korhonen, J.T.; Koponen, L.M.; et al. Multi-Locus Transcranial Magnetic Stimulation System for Electronically Targeted Brain Stimulation. Brain Stimulat. 2022, 15, 116–124. [Google Scholar] [CrossRef]
- Tervo, A.E.; Metsomaa, J.; Nieminen, J.O.; Sarvas, J.; Ilmoniemi, R.J. Automated Search of Stimulation Targets with Closed-Loop Transcranial Magnetic Stimulation. NeuroImage 2020, 220, 117082. [Google Scholar] [CrossRef]
- Tervo, A.E.; Nieminen, J.O.; Lioumis, P.; Metsomaa, J.; Souza, V.H.; Sinisalo, H.; Stenroos, M.; Sarvas, J.; Ilmoniemi, R.J. Closed-Loop Optimization of Transcranial Magnetic Stimulation with Electroencephalography Feedback. Brain Stimulat. 2022, 15, 523–531. [Google Scholar] [CrossRef]
- Casula, E.P.; Leodori, G.; Ibáñez, J.; Benussi, A.; Rawji, V.; Tremblay, S.; Latorre, A.; Rothwell, J.C.; Rocchi, L. The Effect of Coil Orientation on the Stimulation of the Pre–Supplementary Motor Area: A Combined TMS and EEG Study. Brain Sci. 2022, 12, 1358. [Google Scholar] [CrossRef]
- Janssen, A.M.; Oostendorp, T.F.; Stegeman, D.F. The Coil Orientation Dependency of the Electric Field Induced by TMS for M1 and Other Brain Areas. J. NeuroEngineering Rehabil. 2015, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Fries, P. A Mechanism for Cognitive Dynamics: Neuronal Communication through Neuronal Coherence. Trends Cogn. Sci. 2005, 9, 474–480. [Google Scholar] [CrossRef]
- Cabral, J.; Hugues, E.; Sporns, O.; Deco, G. Role of Local Network Oscillations in Resting-State Functional Connectivity. NeuroImage 2011, 57, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Momi, D.; Ozdemir, R.A.; Tadayon, E.; Boucher, P.; Shafi, M.M.; Pascual-Leone, A.; Santarnecchi, E. Network-Level Macroscale Structural Connectivity Predicts Propagation of Transcranial Magnetic Stimulation. NeuroImage 2021, 229, 117698. [Google Scholar] [CrossRef] [PubMed]
- Marzetti, L.; Basti, A.; Chella, F.; D’Andrea, A.; Syrjälä, J.; Pizzella, V. Brain Functional Connectivity Through Phase Coupling of Neuronal Oscillations: A Perspective From Magnetoencephalography. Front. Neurosci. 2019, 13, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, S.; Sarasso, S.; Puglisi, G.E.; Dal Palù, D.; Pigorini, A.; Casarotto, S.; D’Ambrosio, S.; Astolfi, A.; Massimini, M.; Rosanova, M.; et al. TAAC-TMS Adaptable Auditory Control: A Universal Tool to Mask TMS Clicks. J. Neurosci. Methods 2022, 370, 109491. [Google Scholar] [CrossRef]
- Mutanen, T.; Metsomaa, J.; Liljander, S.; Ilmoniemi, R. Automatic and Robust Noise Suppression in EEG and MEG: The SOUND Algorithm. NeuroImage 2017, 166, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Mutanen, T.P.; Metsomaa, J.; Makkonen, M.; Varone, G.; Marzetti, L.; Ilmoniemi, R.J. Source-Based Artifact-Rejection Techniques for TMS-EEG. J. Neurosci. Methods 2022, 382, 109693. [Google Scholar] [CrossRef]
- Palva, S.; Palva, J.M. Discovering Oscillatory Interaction Networks with M/EEG: Challenges and Breakthroughs. Trends Cogn. Sci. 2012, 16, 219–230. [Google Scholar] [CrossRef]
- Nachev, P.; Wydell, H.; O’neill, K.; Husain, M.; Kennard, C. The Role of the Pre-Supplementary Motor Area in the Control of Action. NeuroImage 2007, 36 (Suppl. 2), T155–T163. [Google Scholar] [CrossRef] [Green Version]
- Wolpe, N.; Hezemans, F.H.; Rae, C.L.; Zhang, J.; Rowe, J.B. The Pre-Supplementary Motor Area Achieves Inhibitory Control by Modulating Response Thresholds. Cortex J. Devoted Study Nerv. Syst. Behav. 2022, 152, 98–108. [Google Scholar] [CrossRef]
- Ohbayashi, M. The Roles of the Cortical Motor Areas in Sequential Movements. Front. Behav. Neurosci. 2021, 15, 640659. [Google Scholar] [CrossRef]
- Obeso, I.; Robles, N.; Marrón, E.M.; Redolar-Ripoll, D. Dissociating the Role of the Pre-SMA in Response Inhibition and Switching: A Combined Online and Offline TMS Approach. Front. Hum. Neurosci. 2013, 7, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachev, P.; Kennard, C.; Husain, M. Functional Role of the Supplementary and Pre-Supplementary Motor Areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Lee, J.-M.; Jo, H.J.; Kim, S.H.; Lee, J.H.; Kim, S.T.; Seo, S.W.; Cox, R.W.; Na, D.L.; Kim, S.I.; et al. Defining Functional SMA and Pre-SMA Subregions in Human MFC Using Resting State FMRI: Functional Connectivity-Based Parcellation Method. NeuroImage 2010, 49, 2375–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.W.; Lu, M.-S.; Chen, C.-Y.; Muggleton, N.G.; Hsu, T.-Y.; Juan, C.-H. Roles of the Pre-SMA and RIFG in Conditional Stopping Revealed by Transcranial Magnetic Stimulation. Behav. Brain Res. 2016, 296, 459–467. [Google Scholar] [CrossRef]
- Obeso, I.; Cho, S.S.; Antonelli, F.; Houle, S.; Jahanshahi, M.; Ko, J.H.; Strafella, A.P. Stimulation of the Pre-SMA Influences Cerebral Blood Flow in Frontal Areas Involved with Inhibitory Control of Action. Brain Stimulat. 2013, 6, 769–776. [Google Scholar] [CrossRef]
- Albertini, D.; Gerbella, M.; Lanzilotto, M.; Livi, A.; Maranesi, M.; Ferroni, C.G.; Bonini, L. Connectional Gradients Underlie Functional Transitions in Monkey Pre-Supplementary Motor Area. Prog. Neurobiol. 2020, 184, 101699. [Google Scholar] [CrossRef]
- Messel, M.S.; Raud, L.; Hoff, P.K.; Stubberud, J.; Huster, R.J. Frontal-Midline Theta Reflects Different Mechanisms Associated with Proactive and Reactive Control of Inhibition. NeuroImage 2021, 241, 118400. [Google Scholar] [CrossRef]
- Deiber, M.-P.; Sallard, E.; Ludwig, C.; Ghezzi, C.; Barral, J.; Ibanez, V. EEG Alpha Activity Reflects Motor Preparation Rather than the Mode of Action Selection. Front. Integr. Neurosci. 2012, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Souza, V.H.; Vieira, T.M.; Peres, A.S.C.; Garcia, M.A.C.; Vargas, C.D.; Baffa, O. Effect of TMS Coil Orientation on the Spatial Distribution of Motor Evoked Potentials in an Intrinsic Hand Muscle. Biomed. Eng. Biomed. Tech. 2018, 63, 635–645. [Google Scholar] [CrossRef]
- Tugin, S.; Souza, V.H.; Nazarova, M.A.; Novikov, P.A.; Tervo, A.E.; Nieminen, J.O.; Lioumis, P.; Ziemann, U.; Nikulin, V.V.; Ilmoniemi, R.J. Effect of Stimulus Orientation and Intensity on Short-Interval Intracortical Inhibition (SICI) and Facilitation (SICF): A Multi-Channel Transcranial Magnetic Stimulation Study. PLoS ONE 2021, 16, e0257554. [Google Scholar] [CrossRef]
- Brasil-Neto, J.P.; Cohen, L.G.; Panizza, M.; Nilsson, J.; Roth, B.J.; Hallett, M. Optimal Focal Transcranial Magnetic Activation of the Human Motor Cortex: Effects of Coil Orientation, Shape of the Induced Current Pulse, and Stimulus Intensity. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 1992, 9, 132–136. [Google Scholar] [CrossRef]
- Luppino, G.; Matelli, M.; Camarda, R.; Rizzolatti, G. Corticocortical Connections of Area F3 (SMA-Proper) and Area F6 (Pre-SMA) in the Macaque Monkey. J. Comp. Neurol. 1993, 338, 114–140. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Frequency Bands | a | b | c | d |
|---|---|---|---|---|
| Theta Left | 0.009 | −0.014 | 1.100 | −0.001 |
| Theta Right | 0.015 | 0.013 | 1.089 | −0.002 |
| Alpha Left | 0.009 | −0.014 | 1.083 | 0.003 |
| Alpha Right | 0.013 | 0.013 | 1.086 | −0.002 |
| Beta Left | 0.016 | −0.019 | 1.022 | 0.019 |
| Beta Right | 0.010 | 0.025 | 1.072 | −0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pieramico, G.; Guidotti, R.; Nieminen, A.E.; D’Andrea, A.; Basti, A.; Souza, V.H.; Nieminen, J.O.; Lioumis, P.; Ilmoniemi, R.J.; Romani, G.L.; et al. TMS-Induced Modulation of EEG Functional Connectivity Is Affected by the E-Field Orientation. Brain Sci. 2023, 13, 418. https://doi.org/10.3390/brainsci13030418
Pieramico G, Guidotti R, Nieminen AE, D’Andrea A, Basti A, Souza VH, Nieminen JO, Lioumis P, Ilmoniemi RJ, Romani GL, et al. TMS-Induced Modulation of EEG Functional Connectivity Is Affected by the E-Field Orientation. Brain Sciences. 2023; 13(3):418. https://doi.org/10.3390/brainsci13030418
Chicago/Turabian StylePieramico, Giulia, Roberto Guidotti, Aino E. Nieminen, Antea D’Andrea, Alessio Basti, Victor H. Souza, Jaakko O. Nieminen, Pantelis Lioumis, Risto J. Ilmoniemi, Gian Luca Romani, and et al. 2023. "TMS-Induced Modulation of EEG Functional Connectivity Is Affected by the E-Field Orientation" Brain Sciences 13, no. 3: 418. https://doi.org/10.3390/brainsci13030418