
Selective Modulation of Hippocampal Theta Oscillations in Response to Morphine versus Natural Reward

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Surgery
2.2. Drugs
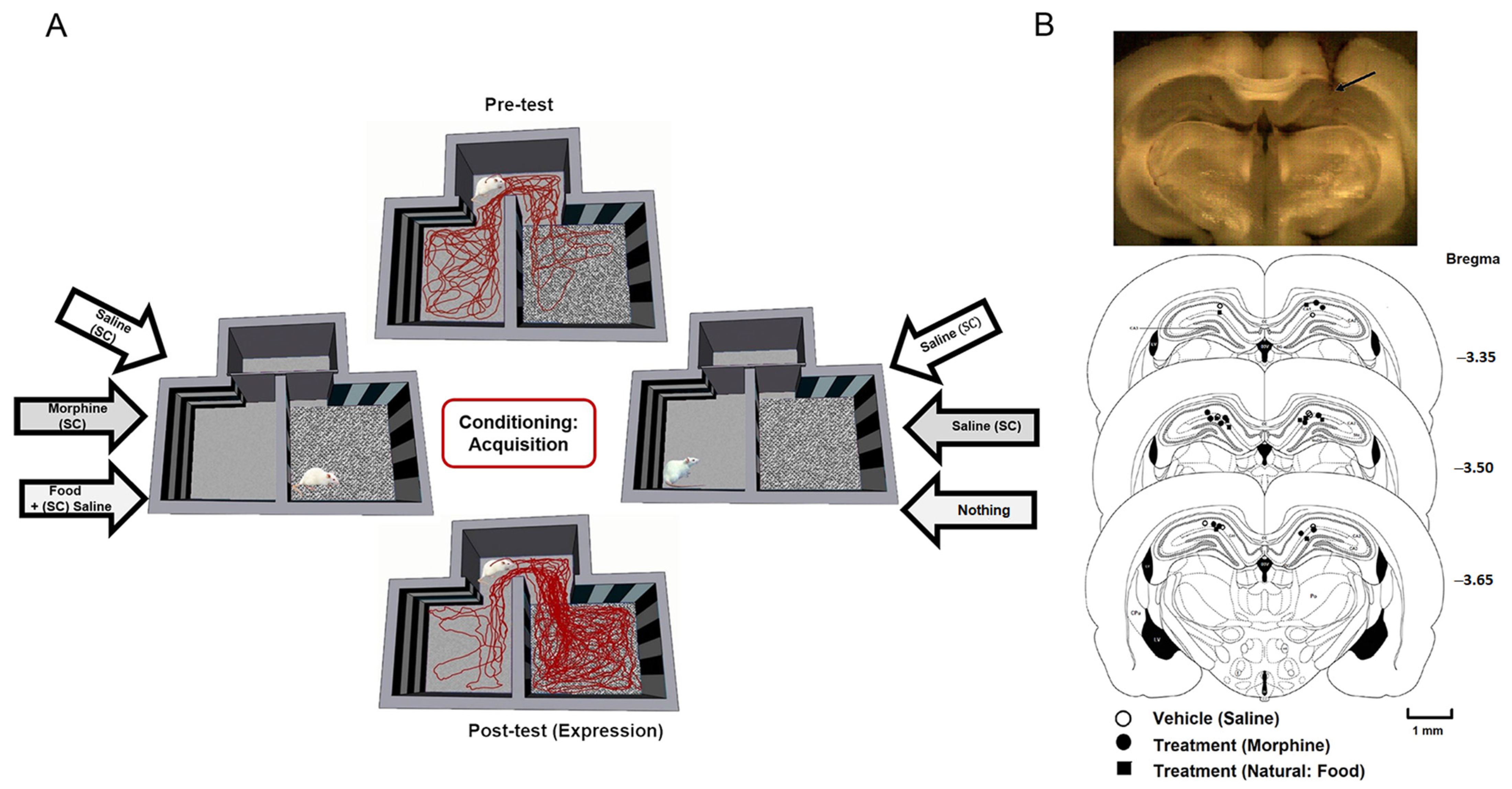
2.3. Conditioned Place Preference Paradigm
2.3.1. Conditioning Phase (Saline, Morphine, Food)
2.3.2. Post-Conditioning Phase (Post-Test)
2.4. Behavioral and Electrophysiological Recordings
2.5. Data Analysis
2.6. Statistics
3. Results
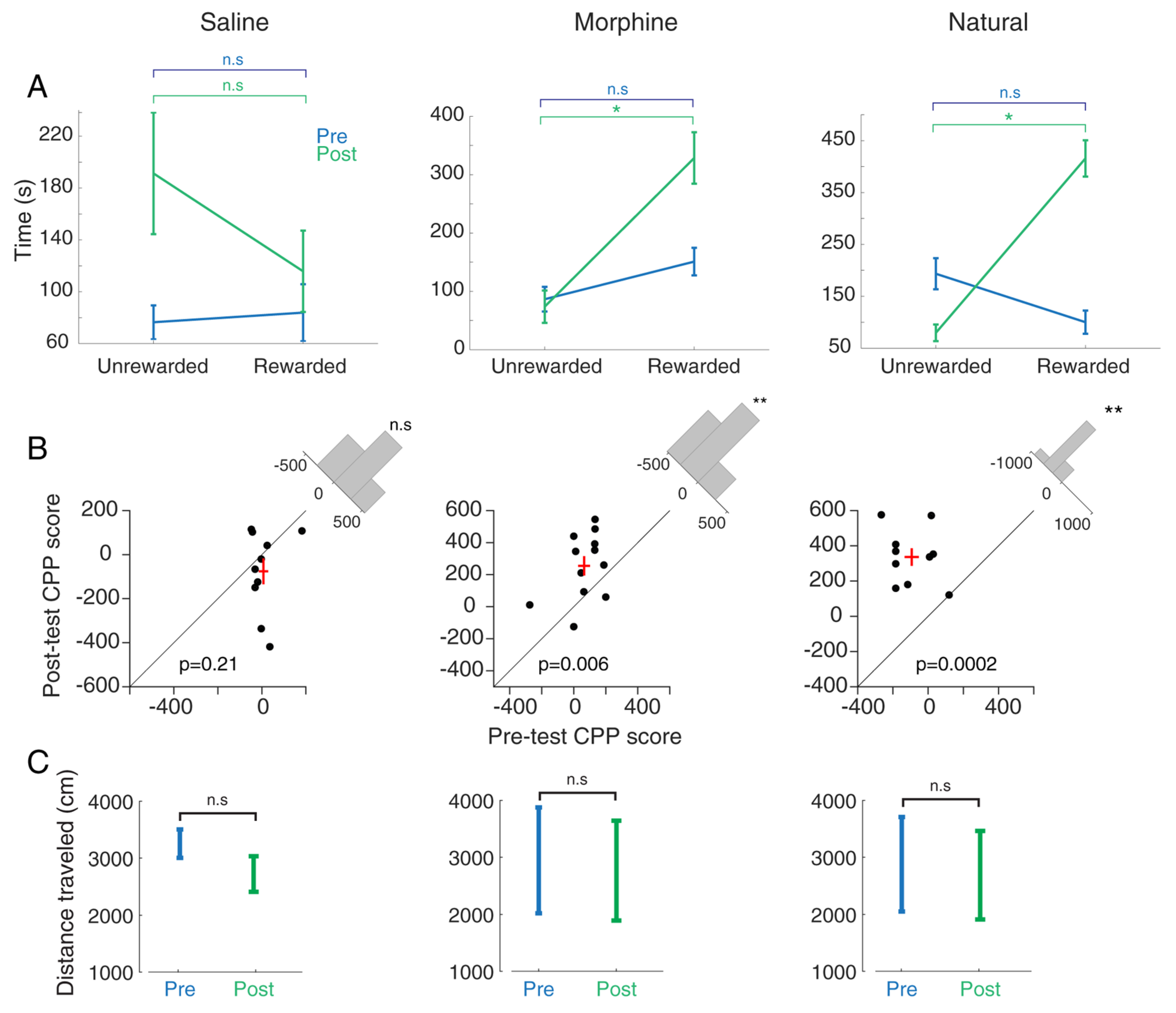
3.1. Comparing Morphine- and Food-Induced Conditioned Place Preference
3.2. Comparing Local Field Potential Activity for Morphine and Food Reward in Conditioned Place Preference Paradigm
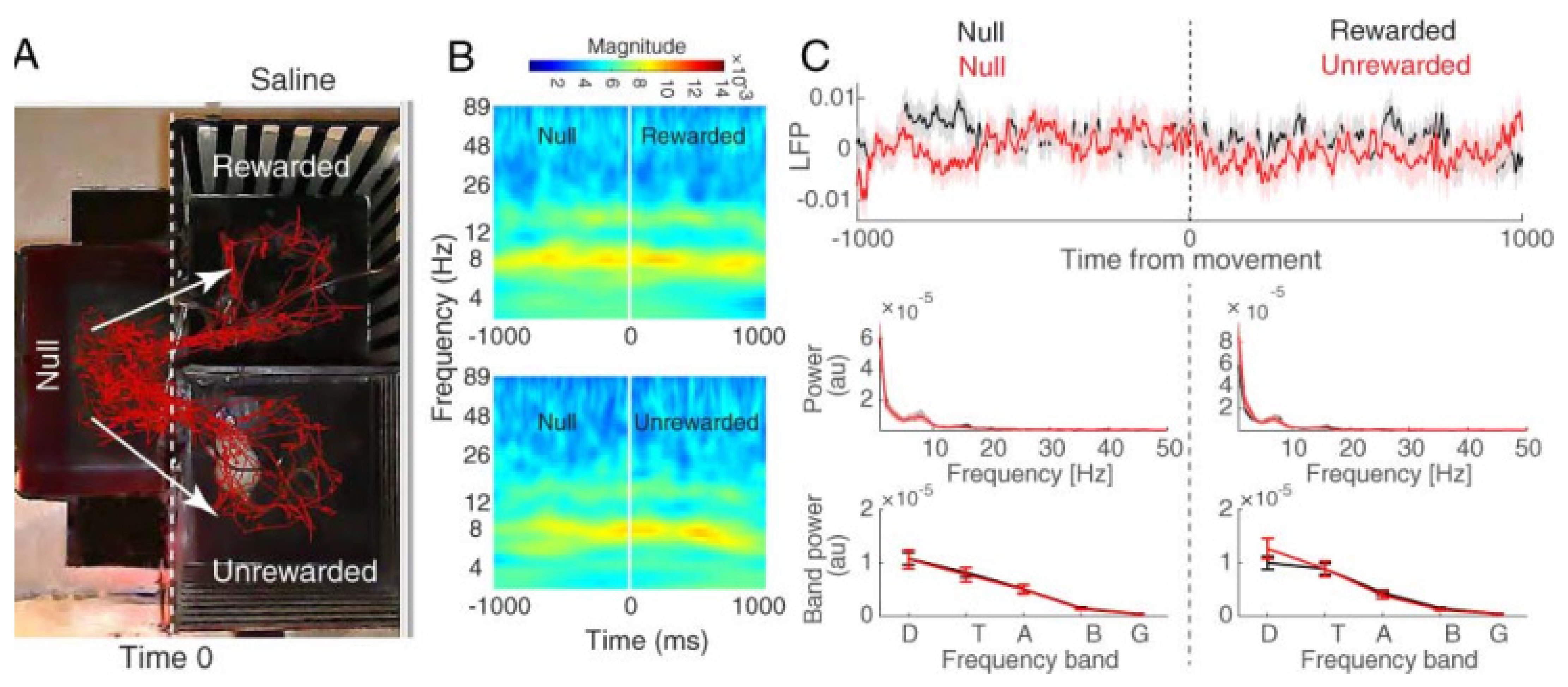
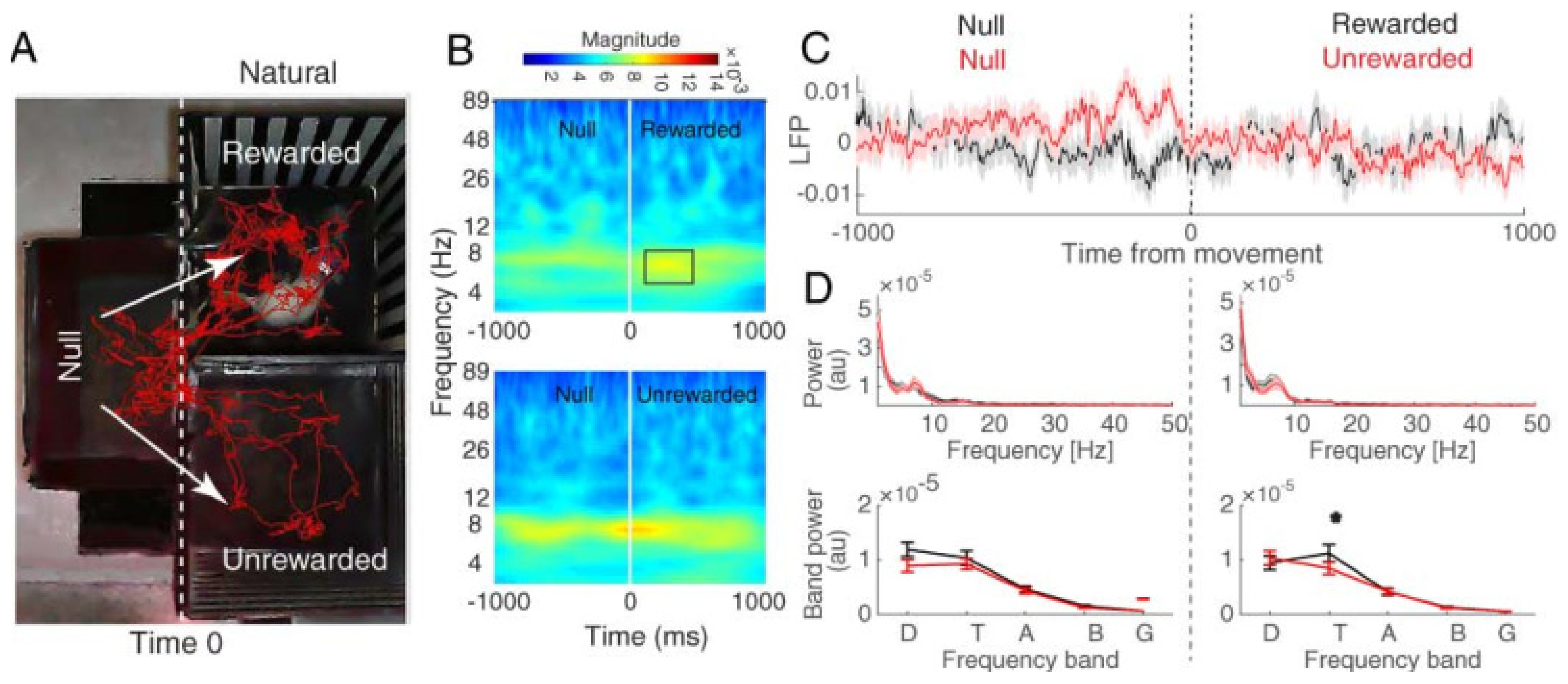
3.2.1. Hippocampal CA1 Theta Activity Relation to Approach to Rewarded and Unrewarded Compartments during the Post-Test in Saline-Treated Animals
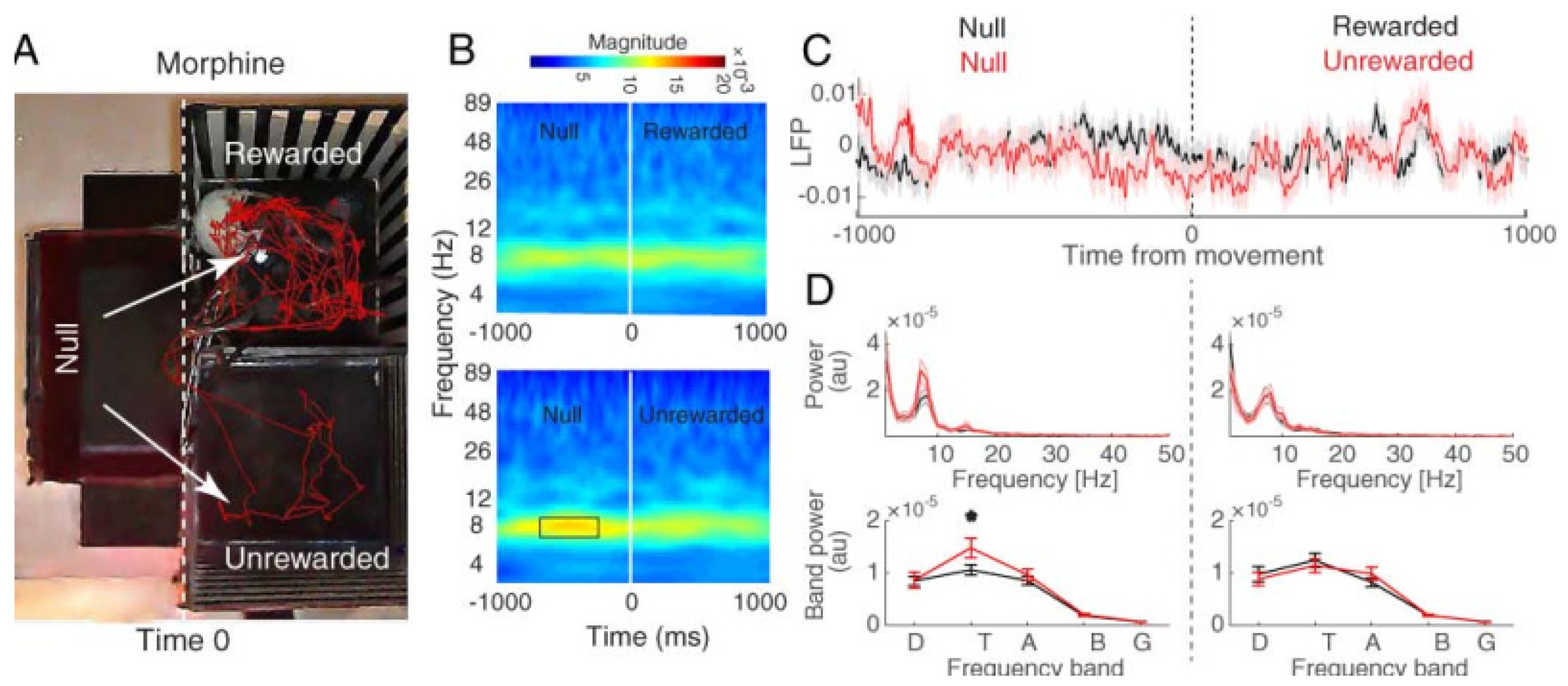
3.2.2. Hippocampal CA1 Theta Activity Relation to Approach to Rewarded and Unrewarded Compartments during the Post-Test with Morphine Reward
3.2.3. Hippocampal CA1 Theta Activity Relation to Approach to Rewarded and Unrewarded Compartments during the Post-Test with Food Reward
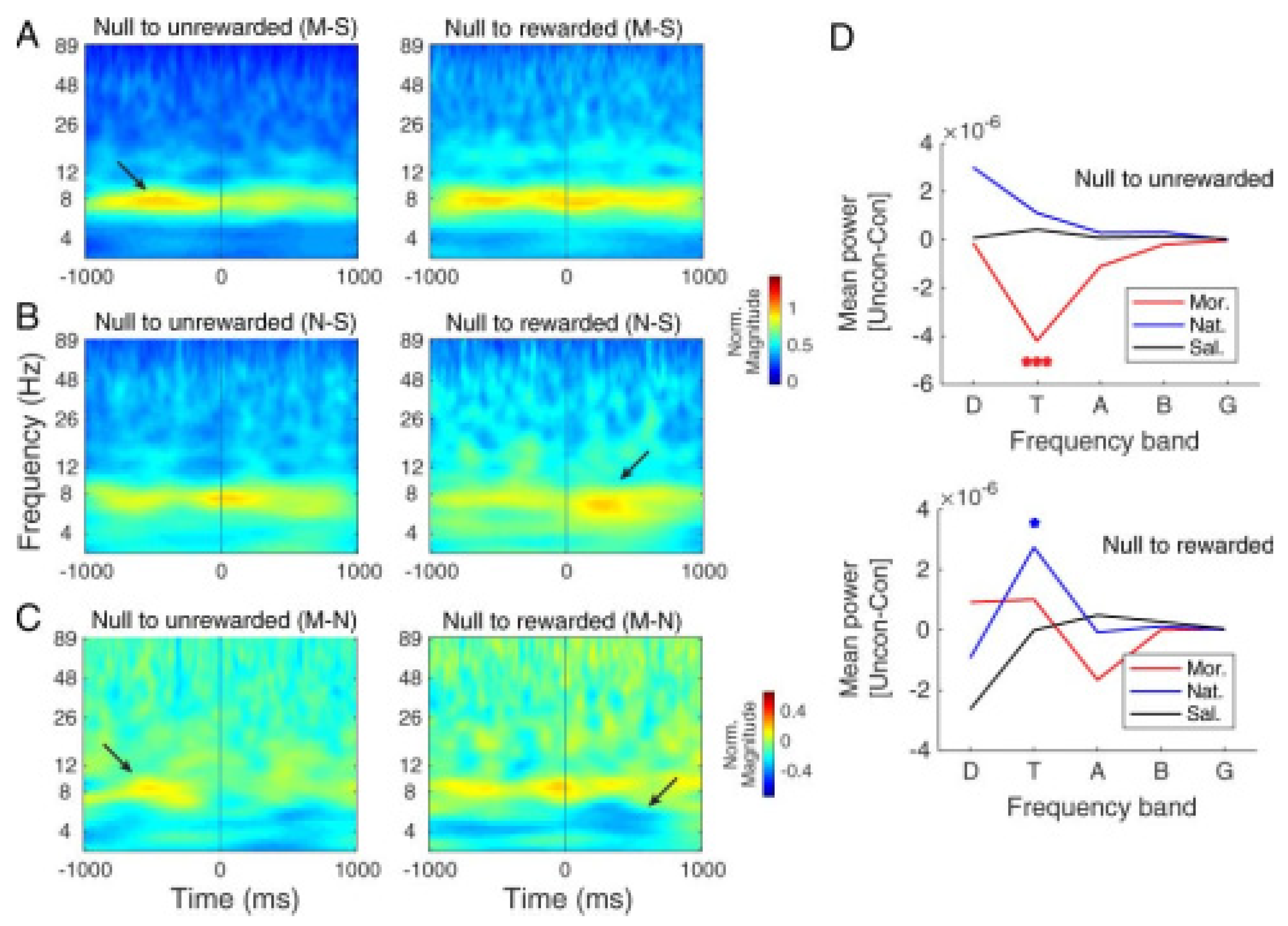
3.2.4. Comparing Hippocampal CA1 Theta Pattern between Morphine- and Natural-Induced CPP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Volkow, N.D.; Michaelides, M.; Baler, R. The neuroscience of drug reward and addiction. Physiol. Rev. 2019, 99, 2115–2140. [Google Scholar] [CrossRef] [PubMed]
- Trotzke, P.; Starcke, K.; Muller, A.; Brand, M. Cue-induced craving and symptoms of online-buying-shopping disorder interfere with performance on the Iowa Gambling Task modified with online-shopping cues. Addict. Behav. 2019, 96, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Rockloff, M.; Armstrong, T.; Hing, N.; Browne, M.; Russell, A.M.T.; Bellringer, M.; du Preez, K.P.; Lowe, G. Legacy Gambling Harms: What Happens Once the Gambling Stops? Curr. Addict. Rep. 2022, 9, 392–399. [Google Scholar] [CrossRef]
- Whatnall, M.; Skinner, J.A.; Leary, M.; Burrows, T.L. Food Addiction: A Deep Dive into ‘Loss of Control’ and ‘Craving’. Curr. Addict. Rep. 2022, 9, 318–325. [Google Scholar] [CrossRef]
- Chatzittofis, A.; Boström, A.D.E.; Savard, J.; Öberg, K.G.; Arver, S.; Jokinen, J. Neurochemical and Hormonal Contributors to Compulsive Sexual Behavior Disorder. Curr. Addict. Rep. 2022, 9, 23–31. [Google Scholar] [CrossRef]
- Olsen, C.M. Natural rewards, neuroplasticity, and non-drug addictions. Neuropharmacology 2011, 61, 1109–1122. [Google Scholar] [CrossRef]
- White, J.K.; Bromberg-Martin, E.S.; Heilbronner, S.R.; Zhang, K.; Pai, J.; Haber, S.N.; Monosov, I.E. A neural network for information seeking. Nat. Commun. 2019, 10, 5168. [Google Scholar] [CrossRef]
- Appelhans, B.M.; Woolf, K.; Pagoto, S.L.; Schneider, K.L.; Whited, M.C.; Liebman, R. Inhibiting food reward: Delay discounting, food reward sensitivity, and palatable food intake in overweight and obese women. Obesity 2011, 19, 2175–2182. [Google Scholar] [CrossRef]
- Leigh, S.-J.; Morris, M.J. The role of reward circuitry and food addiction in the obesity epidemic: An update. Biol. Psychol. 2018, 131, 31–42. [Google Scholar] [CrossRef]
- Rogers, P.J. Food and drug addictions: Similarities and differences. Pharmacol. Biochem. Behav. 2017, 153, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Campana, B.; Brasiel, P.G.; de Aguiar, A.S.; Dutra, S.C.P.L. Obesity and food addiction: Similarities to drug addiction. Obes. Med. 2019, 16, 100136. [Google Scholar] [CrossRef]
- Nall, R.W.; Heinsbroek, J.A.; Nentwig, T.B.; Kalivas, P.W.; Bobadilla, A.C. Circuit selectivity in drug versus natural reward seeking behaviors. J. Neurochem. 2021, 157, 1450–1472. [Google Scholar] [CrossRef] [PubMed]
- Andrianarivelo, A.; Saint-Jour, E.; Pousinha, P.; Fernandez, S.P.; Petitbon, A.; de Smedt-Peyrusse, V.; Heck, N.; Ortiz, V.; Allichon, M.-C.; Kappès, V. Disrupting D1-NMDA or D2-NMDA receptor heteromerization prevents cocaine’s rewarding effects but preserves natural reward processing. Sci. Adv. 2021, 7, eabg5970. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Dixon, J.B. Food for thought: Reward mechanisms and hedonic overeating in obesity. Curr. Obes. Rep. 2017, 6, 353–361. [Google Scholar] [CrossRef]
- Khalili, M.; Rahimi-Movaghar, A.; Shadloo, B.; Mojtabai, R.; Mann, K.; Amin-Esmaeili, M. Global scientific production on illicit drug addiction: A two-decade analysis. Eur. Addict. Res. 2018, 24, 60–70. [Google Scholar] [CrossRef]
- Lowenstein, E.G.; Velazquez-Ulloa, N.A. A Fly’s eye view of natural and drug reward. Front. Physiol. 2018, 9, 407. [Google Scholar] [CrossRef]
- Onaolapo, A.Y.; Onaolapo, O.J. Food additives, food and the concept of ‘food addiction’: Is stimulation of the brain reward circuit by food sufficient to trigger addiction? Pathophysiology 2018, 25, 263–276. [Google Scholar] [CrossRef]
- Hryhorczuk, C.; Sheng, Z.; Décarie-Spain, L.; Giguère, N.; Ducrot, C.; Trudeau, L.; Routh, V.H.; Alquier, T.; Fulton, S. Oleic Acid in the Ventral Tegmental Area Inhibits Feeding, Food Reward, and Dopamine Tone. Neuropsychopharmacology 2018, 43, 607–616. [Google Scholar] [CrossRef]
- Leemburg, S.; Canonica, T.; Luft, A. Motor skill learning and reward consumption differentially affect VTA activation. Sci. Rep. 2018, 8, 687. [Google Scholar] [CrossRef]
- Soares-Cunha, C.; de Vasconcelos, N.A.P.; Coimbra, B.; Domingues, A.V.; Silva, J.M.; Loureiro-Campos, E.; Gaspar, R.; Sotiropoulos, I.; Sousa, N.; Rodrigues, A.J. Nucleus accumbens medium spiny neurons subtypes signal both reward and aversion. Mol. Psychiatry 2019, 25, 3448. [Google Scholar] [CrossRef] [Green Version]
- LeGates, T.A.; Kvarta, M.D.; Tooley, J.R.; Francis, T.C.; Lobo, M.K.; Creed, M.C.; Thompson, S.M. Reward behaviour is regulated by the strength of hippocampus-nucleus accumbens synapses. Nature 2018, 564, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Wassum, K.M.; Izquierdo, A. The basolateral amygdala in reward learning and addiction. Neurosci. Biobehav. Rev. 2015, 57, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Caballero, J.P.; Scarpa, G.B.; Remage-Healey, L.; Moorman, D.E. Differential Effects of Dorsal and Ventral Medial Prefrontal Cortex Inactivation during Natural Reward Seeking, Extinction, and Cue-Induced Reinstatement. eNeuro 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Chaaya, N.; Battle, A.R.; Johnson, L.R. An update on contextual fear memory mechanisms: Transition between Amygdala and Hippocampus. Neurosci. Biobehav. Rev. 2018, 92, 43–54. [Google Scholar] [CrossRef]
- Goyal, A.; Miller, J.; Watrous, A.J.; Lee, S.A.; Coffey, T.; Sperling, M.R.; Sharan, A.; Worrell, G.; Berry, B.; Lega, B.; et al. Electrical Stimulation in Hippocampus and Entorhinal Cortex Impairs Spatial and Temporal Memory. J. Neurosci. 2018, 38, 4471–4481. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.; Schumacher, A.; Sivagurunathan, S.; Van Der Meer, M.; Ito, R. Ventral, but not dorsal, hippocampus inactivation impairs reward memory expression and retrieval in contexts defined by proximal cues. Hippocampus 2017, 27, 822–836. [Google Scholar] [CrossRef]
- Adeniyi, P.A.; Shrestha, A.; Ogundele, O.M. Distribution of VTA Glutamate and Dopamine Terminals, and their Significance in CA1 Neural Network Activity. Neuroscience 2020, 446, 171–198. [Google Scholar] [CrossRef]
- Stuber, G.D.; Sparta, D.R.; Stamatakis, A.M.; van Leeuwen, W.A.; Hardjoprajitno, J.E.; Cho, S.; Tye, K.M.; Kempadoo, K.A.; Zhang, F.; Deisseroth, K.; et al. Excitatory transmission from the amygdala to nucleus accumbens facilitates reward seeking. Nature 2011, 475, 377–380. [Google Scholar] [CrossRef]
- Pavuluri, M.; Volpe, K.; Yuen, A. Nucleus accumbens and its role in reward and emotional circuitry: A potential hot mess in substance use and emotional disorders. AIMS Neurosci. 2017, 4, 52–70. [Google Scholar] [CrossRef]
- Xiao, Z.; Lin, K.; Fellous, J.-M. Conjunctive reward–place coding properties of dorsal distal CA1 hippocampus cells. Biol. Cybern. 2020, 114, 285–301. [Google Scholar] [CrossRef]
- Assar, N.; Mahmoudi, D.; Farhoudian, A.; Farhadi, M.H.; Fatahi, Z.; Haghparast, A. D1-and D2-like dopamine receptors in the CA1 region of the hippocampus are involved in the acquisition and reinstatement of morphine-induced conditioned place preference. Behav. Brain Res. 2016, 312, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Sjulson, L.; Peyrache, A.; Cumpelik, A.; Cassataro, D.; Buzsáki, G. Cocaine place conditioning strengthens location-specific hippocampal inputs to the nucleus accumbens. Neuron 2018, 98, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Churchland, M.M.; Cunningham, J.P.; Kaufman, M.T.; Foster, J.D.; Nuyujukian, P.; Ryu, S.I.; Shenoy, K.V. Neural population dynamics during reaching. Nature 2012, 487, 51–56. [Google Scholar] [CrossRef]
- Sakimoto, Y.; Takeda, K.; Okada, K.; Hattori, M.; Sakata, S. Transient decline in rats’ hippocampal theta power relates to inhibitory stimulus-reward association. Behav. Brain Res. 2013, 246, 132–138. [Google Scholar] [CrossRef]
- Ridley, R.; Timothy, C.; Maclean, C.; Baker, H. Conditional learning and memory impairments following neurotoxic lesion of the CA1 field of the hippocampus. Neuroscience 1995, 67, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, R.; Jamali, S.; Pourhamzeh, M.; Koruji, M.; Ahadi, R.; Haghparast, A. The blockade of D1-and D2-like dopamine receptors within the dentate gyrus attenuates food deprivation stress-induced reinstatement of morphine-extinguished conditioned place preference in rats. Pharmacol. Biochem. Behav. 2020, 196, 172967. [Google Scholar] [CrossRef] [PubMed]
- Nazari-Serenjeh, F.; Jamali, S.; Rezaee, L.; Zarrabian, S.; Haghparast, A. D1-but not D2-like dopamine receptor antagonist in the CA1 region of the hippocampus reduced stress-induced reinstatement in extinguished morphine-conditioning place preference in the food-deprived rats. Behav. Pharmacol. 2020, 31, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Jamali, S.; Aliyari Shoorehdeli, M.; Daliri, M.R.; Haghparast, A. Differential Aspects of Natural and Morphine Reward-Related Behaviors in Conditioned Place Preference Paradigm. Basic Clin. Neurosci. 2022, 13, 731–744. [Google Scholar] [CrossRef]
- Jamali, S.; Zarrabian, S.; Haghparast, A. Similar role of mPFC orexin-1 receptors in the acquisition and expression of morphine-and food-induced conditioned place preference in male rats. Neuropharmacology 2021, 198, 108764. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Dezfouli, M.A.P.; Dezfouli, M.P.; Rad, H.S. A novel approach for baseline correction in 1 H-MRS signals based on ensemble empirical mode decomposition. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014; pp. 3196–3199. [Google Scholar]
- Dezfouli, M.P.; Daliri, M.R. Single-Trial Decoding from Local Field Potential Using Bag of Word Representation. Brain Topogr. 2020, 33, 10–21. [Google Scholar] [CrossRef]
- Danielson, N.B.; Zaremba, J.D.; Kaifosh, P.; Bowler, J.; Ladow, M.; Losonczy, A. Sublayer-specific coding dynamics during spatial navigation and learning in hippocampal area CA1. Neuron 2016, 91, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Gasbarri, A.; Verney, C.; Innocenzi, R.; Campana, E.; Pacitti, C. Mesolimbic dopaminergic neurons innervating the hippocampal formation in the rat: A combined retrograde tracing and immunohistochemical study. Brain Res. 1994, 668, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Bayer, H.M.; Glimcher, P.W. Midbrain dopamine neurons encode a quantitative reward prediction error signal. Neuron 2005, 47, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Le Merre, P.; Esmaeili, V.; Charrière, E.; Galan, K.; Salin, P.-A.; Petersen, C.C.; Crochet, S. Reward-based learning drives rapid sensory signals in medial prefrontal cortex and dorsal hippocampus necessary for goal-directed behavior. Neuron 2018, 97, 83–91. [Google Scholar] [CrossRef]
- Korotkova, T.; Ponomarenko, A.; Monaghan, C.K.; Poulter, S.L.; Cacucci, F.; Wills, T.; Hasselmo, M.E.; Lever, C. Reconciling the different faces of hippocampal theta: The role of theta oscillations in cognitive, emotional and innate behaviors. Neurosci. Biobehav. Rev. 2018, 85, 65–80. [Google Scholar] [CrossRef]
- Hummos, A.; Nair, S.S. An integrative model of the intrinsic hippocampal theta rhythm. PLoS ONE 2017, 12, e0182648. [Google Scholar] [CrossRef]
- Gauthier, J.L.; Tank, D.W. A dedicated population for reward coding in the hippocampus. Neuron 2018, 99, 179–193.e177. [Google Scholar] [CrossRef]
- Sosa, M.; Joo, H.R.; Frank, L.M. Dorsal and Ventral Hippocampal Sharp-Wave Ripples Activate Distinct Nucleus Accumbens Networks. Neuron 2020, 105, 725–741.e8. [Google Scholar] [CrossRef]
- Lansink, C.S.; Meijer, G.T.; Lankelma, J.V.; Vinck, M.A.; Jackson, J.C.; Pennartz, C.M. Reward expectancy strengthens CA1 theta and beta band synchronization and hippocampal-ventral striatal coupling. J. Neurosci. 2016, 36, 10598–10610. [Google Scholar] [CrossRef]
- German, P.W.; Fields, H.L. Rat nucleus accumbens neurons persistently encode locations associated with morphine reward. J. Neurophysiol. 2007, 97, 2094–2106. [Google Scholar] [CrossRef]
- Fakira, A.K.; Massaly, N.; Cohensedgh, O.; Berman, A.; Morón, J.A. Morphine-associated contextual cues induce structural plasticity in hippocampal CA1 pyramidal neurons. Neuropsychopharmacology 2016, 41, 2668–2678. [Google Scholar] [CrossRef] [PubMed]
- Morin, J.-P.; Rodríguez-Durán, L.F.; Guzmán-Ramos, K.; Perez-Cruz, C.; Ferreira, G.; Diaz-Cintra, S.; Pacheco-López, G. Palatable hyper-caloric foods impact on neuronal plasticity. Front. Behav. Neurosci. 2017, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- DiLeone, R.J.; Taylor, J.R.; Picciotto, M.R. The drive to eat: Comparisons and distinctions between mechanisms of food reward and drug addiction. Nat. Neurosci. 2012, 15, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Veit, R.; Sauer, H.; Enck, P.; Friederich, H.-C.; Unholzer, T.; Bauer, U.-M.; Linder, K.; Heni, M.; Fritsche, A. Dopamine depletion reduces food-related reward activity independent of BMI. Neuropsychopharmacology 2016, 41, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Boekhoudt, L.; Roelofs, T.; de Jong, J.; de Leeuw, A.; Luijendijk, M.; Wolterink-Donselaar, I.; van der Plasse, G.; Adan, R. Does activation of midbrain dopamine neurons promote or reduce feeding? Int. J. Obes. 2017, 41, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.D.; Black, S.W.; Fisher, S.P.; Palmerston, J.B.; Morairty, S.R.; Hoener, M.C.; Kilduff, T.S. Trace amine-associated receptor 1 regulates wakefulness and EEG spectral composition. Neuropsychopharmacology 2017, 42, 1305–1314. [Google Scholar] [CrossRef]
- Orzeł-Gryglewska, J.; Matulewicz, P.; Jurkowlaniec, E. Brainstem system of hippocampal theta induction: The role of the ventral tegmental area. Synapse 2015, 69, 553–575. [Google Scholar] [CrossRef]
- Laplante, F.; Sibley, D.R.; Quirion, R. Reduction in acetylcholine release in the hippocampus of dopamine D5 receptor-deficient mice. Neuropsychopharmacology 2004, 29, 1620–1627. [Google Scholar] [CrossRef]
- Buzsáki, G. Theta oscillations in the hippocampus. Neuron 2002, 33, 325–340. [Google Scholar] [CrossRef]
- Gu, Z.; Alexander, G.M.; Dudek, S.M.; Yakel, J.L. Hippocampus and Entorhinal Cortex Recruit Cholinergic and NMDA Receptors Separately to Generate Hippocampal Theta Oscillations. Cell Rep. 2017, 21, 3585–3595. [Google Scholar] [CrossRef] [Green Version]
- Huxter, J.R.; Zinyuk, L.E.; Roloff, E.; Clarke, V.R.; Dolman, N.P.; More, J.C.; Jane, D.E.; Collingridge, G.L.; Muller, R.U. Inhibition of kainate receptors reduces the frequency of hippocampal theta oscillations. J. Neurosci. 2007, 27, 2212–2223. [Google Scholar] [CrossRef] [PubMed]
- Siahposht-Khachaki, A.; Fatahi, Z.; Yans, A.; Khodagholi, F.; Haghparast, A. Involvement of AMPA/Kainate Glutamate Receptor in the Extinction and Reinstatement of Morphine-Induced Conditioned Place Preference: A Behavioral and Molecular Study. Cell. Mol. Neurobiol. 2017, 37, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, D.M.; Dai, Y.Y.; Glantz, E.H.; Jadhav, S.P. Disruption of dorsal hippocampal–prefrontal interactions using chemogenetic inactivation impairs spatial learning. Neurobiol. Learn. Mem. 2018, 155, 351–360. [Google Scholar] [CrossRef]
- Desikan, S.; Koser, D.E.; Neitz, A.; Monyer, H. Target selectivity of septal cholinergic neurons in the medial and lateral entorhinal cortex. Proc. Natl. Acad. Sci. USA 2018, 115, E2644–E2652. [Google Scholar] [CrossRef] [PubMed]
- Rezayof, A.; Zatali, H.; Haeri-Rohani, A.; Zarrindast, M.-R. Dorsal hippocampal muscarinic and nicotinic receptors are involved in mediating morphine reward. Behav. Brain Res. 2006, 166, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Grasing, K. A threshold model for opposing actions of acetylcholine on reward behavior: Molecular mechanisms and implications for treatment of substance abuse disorders. Behav. Brain Res. 2016, 312, 148–162. [Google Scholar] [CrossRef]
- Grasing, K.W.; Xu, H.; Idowu, J.Y. The muscarinic agonist pilocarpine modifies cocaine-reinforced and food-reinforced responding in rats: Comparison with the cholinesterase inhibitor tacrine. Behav. Pharmacol. 2019, 30, 478–489. [Google Scholar] [CrossRef]
- Leung, L.S.; Shen, B. GABAB receptor blockade enhances theta and gamma rhythms in the hippocampus of behaving rats. Hippocampus 2007, 17, 281–291. [Google Scholar] [CrossRef]
- Zarrindast, M.-R.; Massoudi, R.; Sepehri, H.; Rezayof, A. Involvement of GABAB receptors of the dorsal hippocampus on the acquisition and expression of morphine-induced place preference in rats. Physiol. Behav. 2006, 87, 31–38. [Google Scholar] [CrossRef]
- Rezayof, A.; Razavi, S.; Haeri-Rohani, A.; Rassouli, Y.; Zarrindast, M.-R. GABAA receptors of hippocampal CA1 regions are involved in the acquisition and expression of morphine-induced place preference. Eur. Neuropsychopharmacol. 2007, 17, 24–31. [Google Scholar] [CrossRef]
- Liang, J.; Li, Y.; Ping, X.; Yu, P.; Zuo, Y.; Wu, L.; Han, J.-S.; Cui, C. The possible involvement of endogenous ligands for mu-, delta-and kappa-opioid receptors in modulating morphine-induced CPP expression in rats. Peptides 2006, 27, 3307–3314. [Google Scholar] [CrossRef] [PubMed]
- Gibula-Tarlowska, E.; Kedzierska, E.; Piechura, K.; Silberring, J.; Kotlinska, J. The influence of a new derivate of kisspeptin-10–Kissorphin (KSO) on the rewarding effects of morphine in the conditioned place preference (CPP) test in male rats. Behav. Brain Res. 2019, 372, 112043. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Saanijoki, T.; Tuominen, L.; Hirvonen, J.; Tuulari, J.J.; Nuutila, P.; Kalliokoski, K. μ-opioid receptor system mediates reward processing in humans. Nat. Commun. 2018, 9, 1500. [Google Scholar] [CrossRef] [PubMed]
- Ziauddeen, H.; Chamberlain, S.R.; Nathan, P.; Koch, A.; Maltby, K.; Bush, M.; Tao, W.; Napolitano, A.; Skeggs, A.L.; Brooke, A. Effects of the mu-opioid receptor antagonist GSK1521498 on hedonic and consummatory eating behaviour: A proof of mechanism study in binge-eating obese subjects. Mol. Psychiatry 2013, 18, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Łupina, M.; Talarek, S.; Kotlińska, J.; Gibuła-Tarłowska, E.; Listos, P.; Listos, J. The role of linagliptin, a selective dipeptidyl peptidase-4 inhibitor, in the morphine rewarding effects in rats. Neurochem. Int. 2020, 133, 104616. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 neurons in the nucleus of the solitary tract project directly to the ventral tegmental area and nucleus accumbens to control for food intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Orsini, C.; Bonito-Oliva, A.; Conversi, D.; Cabib, S. Susceptibility to conditioned place preference induced by addictive drugs in mice of the C57BL/6 and DBA/2 inbred strains. Psychopharmacology 2005, 181, 327–336. [Google Scholar] [CrossRef]
- Vogel, H.; Kraemer, M.; Rabasa, C.; Askevik, K.; Adan, R.A.; Dickson, S.L. Genetic predisposition to obesity affects behavioural traits including food reward and anxiety-like behaviour in rats. Behav. Brain Res. 2017, 328, 95–104. [Google Scholar] [CrossRef]
- Dickson, P.E.; McNaughton, K.A.; Hou, L.; Anderson, L.C.; Long, K.H.; Chesler, E.J. Sex and strain influence attribution of incentive salience to reward cues in mice. Behav. Brain Res. 2015, 292, 305–315. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamali, S.; Dezfouli, M.P.; Kalbasi, A.; Daliri, M.R.; Haghparast, A. Selective Modulation of Hippocampal Theta Oscillations in Response to Morphine versus Natural Reward. Brain Sci. 2023, 13, 322. https://doi.org/10.3390/brainsci13020322
Jamali S, Dezfouli MP, Kalbasi A, Daliri MR, Haghparast A. Selective Modulation of Hippocampal Theta Oscillations in Response to Morphine versus Natural Reward. Brain Sciences. 2023; 13(2):322. https://doi.org/10.3390/brainsci13020322
Chicago/Turabian StyleJamali, Shole, Mohsen Parto Dezfouli, AmirAli Kalbasi, Mohammad Reza Daliri, and Abbas Haghparast. 2023. "Selective Modulation of Hippocampal Theta Oscillations in Response to Morphine versus Natural Reward" Brain Sciences 13, no. 2: 322. https://doi.org/10.3390/brainsci13020322