
On the Homology of the Dominant and Non-Dominant Corticospinal Tracts: A Novel Neurophysiological Assessment

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Methods
2.1. Study Design
2.2. Healthy Volunteers’ Population
2.3. MEP Collection and Analysis
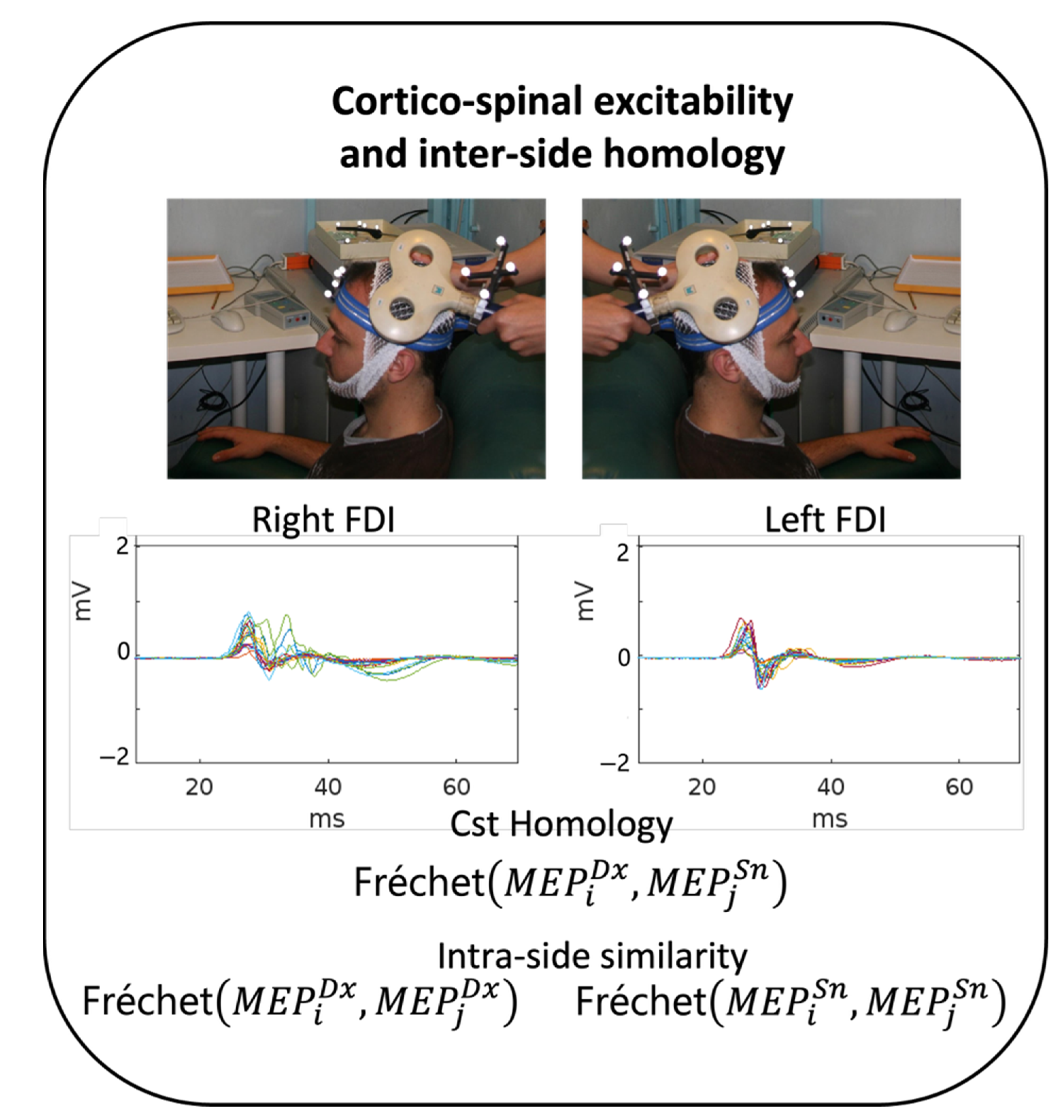
2.3.1. Stimulation and Recording Setup
2.3.2. MEP Morphology Similarity Estimate
2.3.3. MEP Amplitude Estimate
2.3.4. Statistical Analysis
2.3.5. Data Availability
3. Results
3.1. Healthy Volunteers’ Population and MEP Features
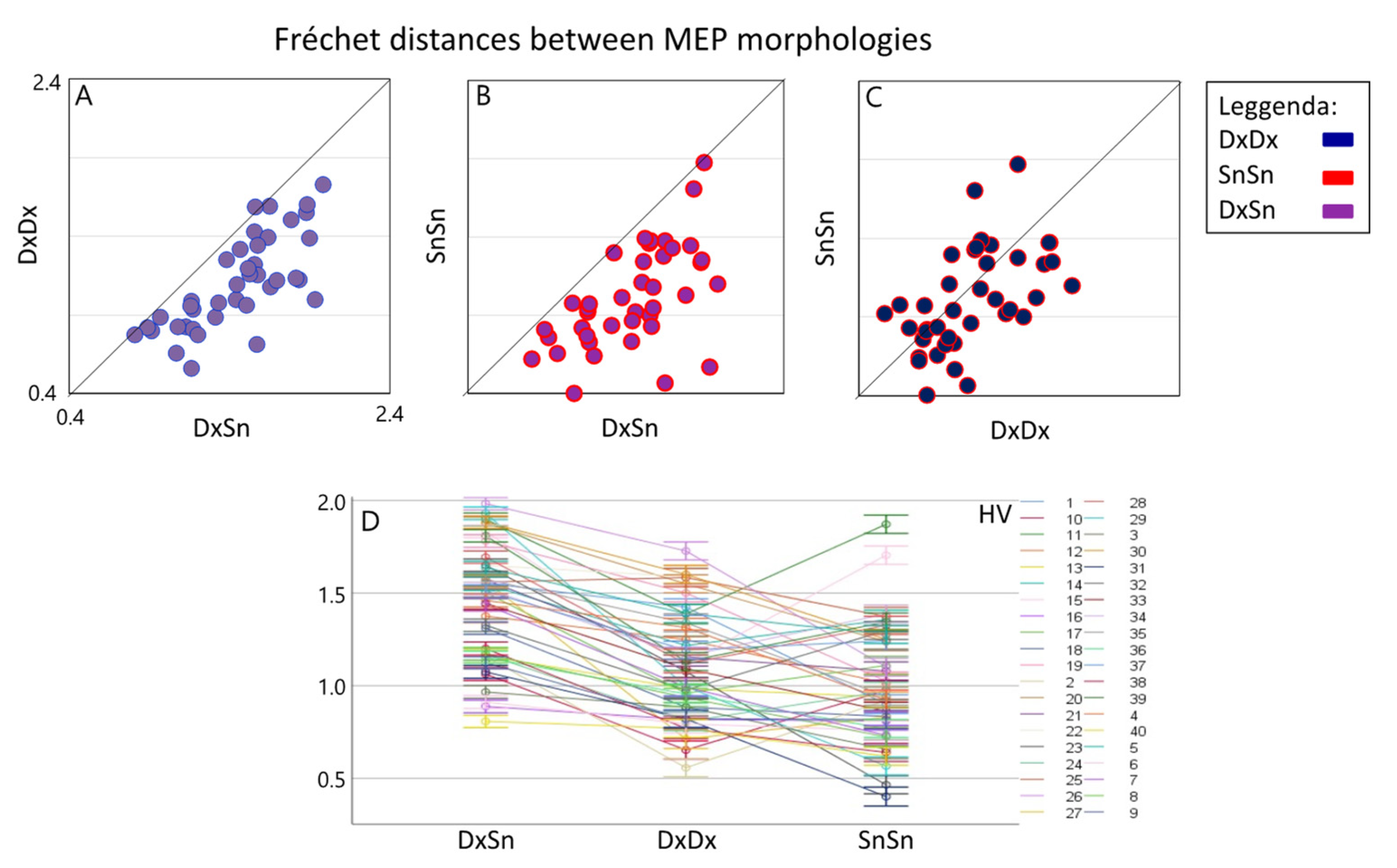
3.2. CTS Homology via MEP’s Fréchet Distance
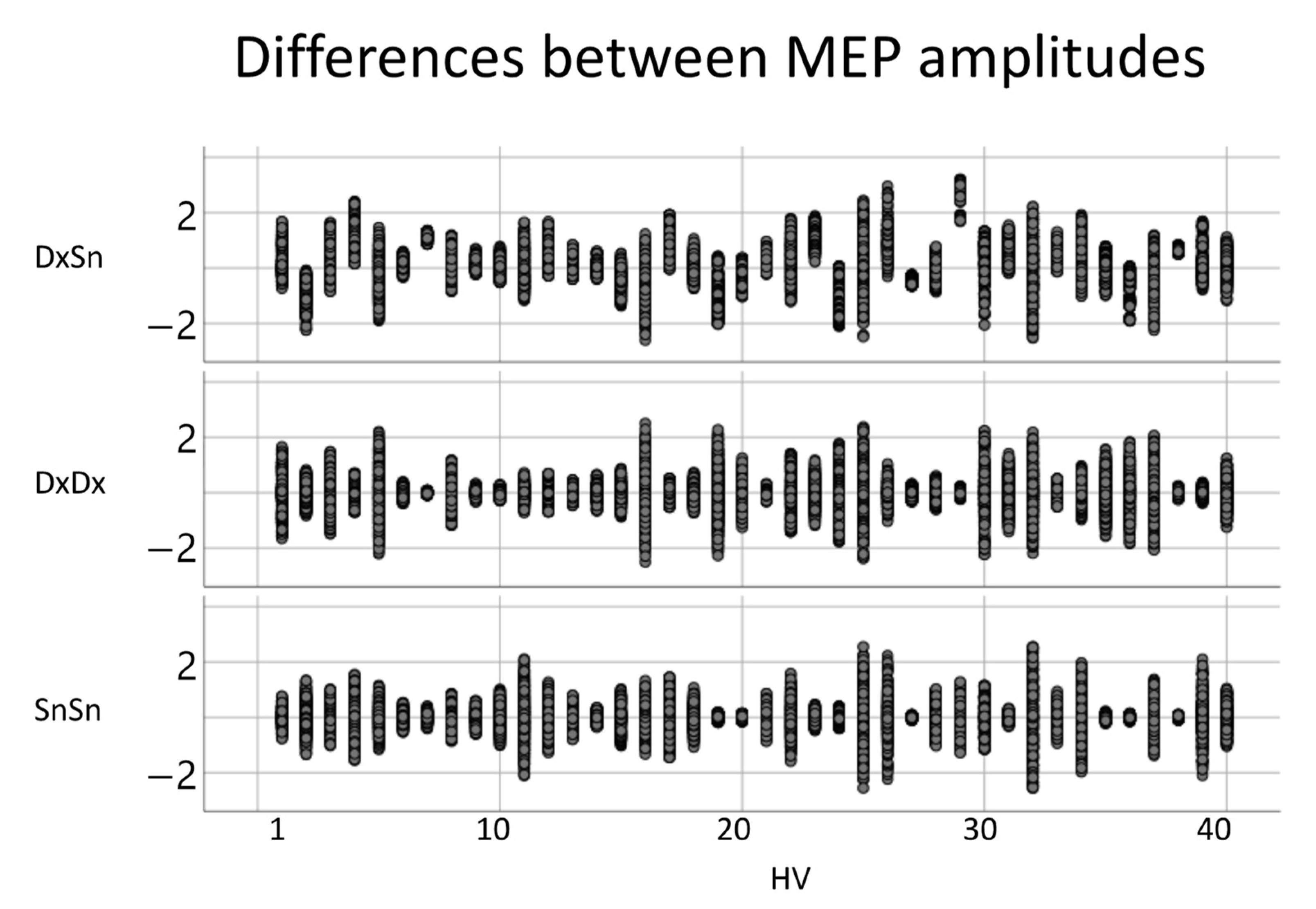
3.3. CTS Homology via MEP’s Amplitudes
4. Discussion
4.1. Higher Variability of Recruitment Pattern in the Dominant than the Non-Dominant Hemibody
4.2. CST Homology vs. Intra-Side Recruitment Pattern Similarity
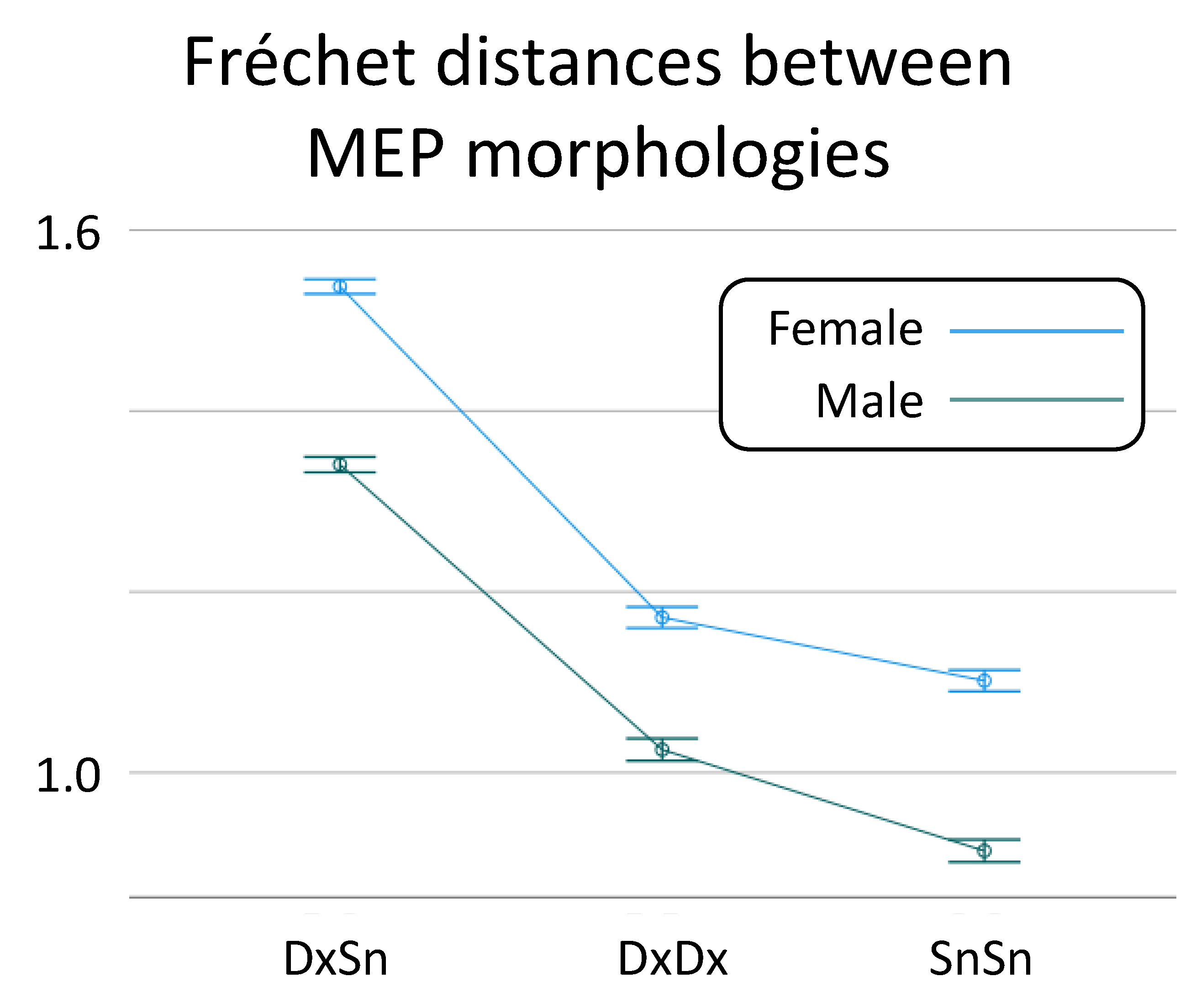
4.3. Sex Effect on CST Recruitment Variability
4.4. Future Neuroscience and Clinical Implications
4.5. MEP Shape vs. Amplitude
4.6. Limitations of the Present Work
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deco, G.; Corbetta, M. The Dynamical Balance of the Brain at Rest. Neuroscientist 2011, 17, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Cottone, C.; Tomasevic, L.; Porcaro, C.; Filligoi, G.; Tecchio, F. Physiological Aging Impacts the Hemispheric Balances of Resting State Primary Somatosensory Activities. Brain Topogr. 2013, 26, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.J.; Fields, R.D.; Johansen-Berg, H. Plasticity in Gray and White: Neuroimaging Changes in Brain Structure during Learning. Nat. Neurosci. 2012, 15, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Carson, R.G. Inter-Hemispheric Inhibition Sculpts the Output of Neural Circuits by Co-Opting the Two Cerebral Hemispheres. J. Physiol. 2020, 598, 4781–4802. [Google Scholar] [CrossRef]
- Daskalakis, Z.J.; Christensen, B.K.; Fitzgerald, P.B.; Roshan, L.; Chen, R. The Mechanisms of Interhemispheric Inhibition in the Human Motor Cortex. J. Physiol. 2002, 543, 317–326. [Google Scholar] [CrossRef]
- Rokni, U.; Steinberg, O.; Vaadia, E.; Sompolinsky, H. Cortical Representation of Bimanual Movements. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 11577–11586. [Google Scholar] [CrossRef]
- Mahan, M.Y.; Georgopoulos, A.P. Motor Directional Tuning across Brain Areas: Directional Resonance and the Role of Inhibition for Directional Accuracy. Front. Neural Circuits 2013, 7, 92. [Google Scholar] [CrossRef]
- Georgopoulos, A.P.; Carpenter, A.F. Coding of Movements in the Motor Cortex. Curr. Opin. Neurobiol. 2015, 33, 34–39. [Google Scholar] [CrossRef]
- Kolasinski, J.; Logan, J.P.; Hinson, E.L.; Makin, T.R.; Emir, U.E.; Stagg Correspondence, C.J. A Mechanistic Link from GABA to Cortical Architecture and Perception. Curr. Biol. 2017, 27, 1685–1691.e3. [Google Scholar] [CrossRef]
- Merchant, H.; Naselaris, T.; Georgopoulos, A.P. Dynamic Sculpting of Directional Tuning in the Primate Motor Cortex during Three-Dimensional Reaching. J. Neurosci. 2008, 28, 9164–9172. [Google Scholar] [CrossRef]
- Georgopoulos, A.P.; Stefanis, C.N. The Motor Cortical Circuit. In Handbook of Brain Microcircuits; Oxford University Press: Oxford, UK, 2013; pp. 39–45. [Google Scholar]
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-Invasive Magnetic Stimulation of Human Motor Cortex. Lancet 1985, 325, 1106–1107. [Google Scholar] [CrossRef]
- Groppa, S.; Oliviero, A.; Eisen, A.; Quartarone, A.; Cohen, L.G.; Mall, V.; Kaelin-Lang, A.; Mima, T.; Rossi, S.; Thickbroom, G.W.; et al. A Practical Guide to Diagnostic Transcranial Magnetic Stimulation: Report of an IFCN Committee. Clin. Neurophysiol. 2012, 123, 858–882. [Google Scholar] [CrossRef]
- Tecchio, F.; Pasqualetti, P.; Pizzella, V.; Romani, G.; Rossini, P.M. Morphology of Somatosensory Evoked Fields: Inter-Hemispheric Similarity as a Parameter for Physiological and Pathological Neural Connectivity. Neurosci. Lett. 2000, 287, 203–206. [Google Scholar] [CrossRef]
- Tecchio, F.; Zappasodi, F.; Pasqualetti, P.; Rossini, P.M. Neural Connectivity in Hand Sensorimotor Brain Areas: An Evaluation by Evoked Field Morphology. Hum. Brain Mapp. 2005, 24, 99–108. [Google Scholar] [CrossRef]
- Fréchet, M.M. Sur Quelques Points Du Calcul Fonctionnel. Rend. Circ. Mat. Palermo 1906, 22, 1–72. [Google Scholar] [CrossRef]
- Wylie, T.; Zhu, B. Following a Curve with the Discrete Fréchet Distance. Theor. Comput. Sci. 2014, 556, 34–44. [Google Scholar] [CrossRef]
- Fournel, A.P.; Reynaud, E.; Brammer, M.J.; Simmons, A.; Ginestet, C.E. Group Analysis of Self-Organizing Maps Based on Functional MRI Using Restricted Frechet Means. NeuroImage 2013, 76, 373–385. [Google Scholar] [CrossRef]
- Wright, R.; Makropoulos, A.; Kyriakopoulou, V.; Patkee, P.A.; Koch, L.M.; Rutherford, M.A.; Hajnal, J.V.; Rueckert, D.; Aljabar, P. Construction of a Fetal Spatio-Temporal Cortical Surface Atlas from in Utero MRI: Application of Spectral Surface Matching. NeuroImage 2015, 120, 467–480. [Google Scholar] [CrossRef]
- Nazarova, M.; Kulikova, S.; Piradov, M.A.; Limonova, A.S.; Dobrynina, L.A.; Konovalov, R.N.; Novikov, P.A.; Sehm, B.; Villringer, A.; Saltykova, A.; et al. Multimodal Assessment of the Motor System in Patients With Chronic Ischemic Stroke. Stroke 2020, 52, 241–249. [Google Scholar] [CrossRef]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Cancelli, A.; Cottone, C.; Giordani, A.; Asta, G.; Lupoi, D.; Pizzella, V.; Tecchio, F. MRI-Guided Regional Personalized Electrical Stimulation in Multisession and Home Treatments. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Suckley, J.J.; Waters, T.J.; Tran, M.; Stapley, P.J.; Shemmell, J.; Walsh, J.A.; McAndrew, D.J. Randomising Stimulus Intensity Improves the Variability and Reliability of the Assessment of Corticospinal Excitability. J. Neurosci. Methods 2020, 342, 108813. [Google Scholar] [CrossRef] [PubMed]
- Eiter, T.; Mannila, H. Computing Discrete Frechet Distance. CiteSeerX 1994. [Google Scholar]
- Tecchio, F.; Bertoli, M.; Gianni, E.; L’Abbate, T.; Paulon, L.; Zappasodi, F. To Be Is to Become. Fractal Neurodynamics of the Body-Brain Control System. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Faisal, A.A.; Selen, L.P.J.; Wolpert, D.M. Noise in the Nervous System. Nat. Rev. Neurosci. 2008, 9, 292–303. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal Olscillations in Cortical Networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Marder, E.; Goaillard, J.M. Variability, Compensation and Homeostasis in Neuron and Network Function. Nat. Rev. Neurosci. 2006, 7, 563–574. [Google Scholar] [CrossRef]
- Honey, C.J.; Sporns, O.; Cammoun, L.; Gigandet, X.; Thiran, J.P.; Meuli, R.; Hagmann, P. Predicting Human Resting-State Functional Connectivity from Structural Connectivity. Proc. Natl. Acad. Sci. USA 2009, 106, 2035–2040. [Google Scholar] [CrossRef]
- Van Vreeswijk, C.; Sompolinsky, H. Chaos in Neuronal Networks with Balanced Excitatory and Inhibitory Activity. Science 1996, 274, 1724–1726. [Google Scholar] [CrossRef]
- Averbeck, B.B.; Latham, P.E.; Pouget, A. Neural Correlations, Population Coding and Computation. Nat. Rev. Neurosci. 2006, 7, 358–366. [Google Scholar] [CrossRef]
- Volz, L.J.; Hamada, M.; Rothwell, J.C.; Grefkes, C. What Makes the Muscle Twitch: Motor System Connectivity and TMS-Induced Activity. Cereb. Cortex 2015, 25, 2346–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellicciari, M.C.; Miniussi, C.; Ferrari, C.; Koch, G.; Bortoletto, M. Ongoing Cumulative Effects of Single TMS Pulses on Corticospinal Excitability: An Intra- and Inter-Block Investigation. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2016, 127, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Sohn, Y.H.; Hallett, M. Hemispheric Asymmetry of Surround Inhibition in the Human Motor System. Clin. Neurophysiol. 2009, 120, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Amunts, K.; Schlaug, G.; Schleicher, A.; Steinmetz, H.; Dabringhaus, A.; Roland, P.E.; Zilles, K. Asymmetry in the Human Motor Cortex and Handedness. NeuroImage 1996, 4, 216–222. [Google Scholar] [CrossRef]
- Graziadio, S.; Basu, A.; Tomasevic, L.; Zappasodi, F.; Tecchio, F.; Eyre, J.A. Developmental Tuning and Decay in Senescence of Oscillations Linking the Corticospinal System. J. Neurosci. 2010, 30, 3663–3674. [Google Scholar] [CrossRef]
- Das, A.; Gilbert, C.D. Topography of Contextual Modulations Mediated by Short-Range Interactions in Primary Visual Cortex. Nature 1999, 399, 655–661. [Google Scholar] [CrossRef]
- Ekeberg, Ö. A Combined Neuronal and Mechanical Model of Fish Swimming. Biol. Cybern. 1993, 69, 363–374. [Google Scholar] [CrossRef]
- Dell’Acqua, M.L.; Landi, D.; Zito, G.; Zappasodi, F.; Lupoi, D.; Rossini, P.M.; Filippi, M.M.; Tecchio, F. Thalamocortical Sensorimotor Circuit in Multiple Sclerosis: An Integrated Structural and Electrophysiological Assessment. Hum. Brain Mapp. 2010, 31, 1588–1600. [Google Scholar] [CrossRef]
- Tecchio, F.; Cecconi, F.; Colamartino, E.; Padalino, M.; Valci, L.; Reinert, M. The Morphology of Somatosensory Evoked Potentials During Middle Cerebral Artery Aneurysm Clipping (MoSAC): A Pilot Study. Clin. EEG Neurosci. 2020, 51, 130–136. [Google Scholar] [CrossRef]
- Köhling, R.; Reinel, J.; Vahrenhold, J.; Hinrichs, K.; Speckmann, E.J. Spatio-Temporal Patterns of Neuronal Activity: Analysis of Optical Imaging Data Using Geometric Shape Matching. J. Neurosci. Methods 2002, 114, 17–23. [Google Scholar] [CrossRef]
- Pitcher, J.B.; Ogston, K.M.; Miles, T.S. Age and Sex Differences in Human Motor Cortex Input-Output Characteristics. J. Physiol. 2003, 546, 605–613. [Google Scholar] [CrossRef]
- de Gennaro, L.; Bertini, M.; Pauri, F.; Cristiani, R.; Curcio, G.; Ferrara, M.; Rossini, P.M. Callosal Effects of Transcranial Magnetic Stimulation (TMS): The Influence of Gender and Stimulus Parameters. Neurosci. Res. 2004, 48, 129–137. [Google Scholar] [CrossRef]
- Kuo, M.F.; Paulus, W.; Nitsche, M.A. Sex Differences in Cortical Neuroplasticity in Humans. Neuroreport 2006, 17, 1703–1707. [Google Scholar] [CrossRef]
- Yperman, J.; Becker, T.; Valkenborg, D.; Hellings, N.; Cambron, M.; Dive, D.; Laureys, G.; Popescu, V.; Van Wijmeersch, B.; Peeters, L.M. Deciphering the Morphology of Motor Evoked Potentials. Front. Neuroinform. 2020, 14, 28. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RMT (%) | MEP | |||||
|---|---|---|---|---|---|---|
| Dx | Sn | Delta | Lat Dx | Lat Sn | Amp Dx | Amp Sn |
| 48.1 | 48.1 | −0.1 | 26.3 | 26.4 | 1.7 | 1.3 |
| 9.1 | 8.1 | 3.3 | 0.9 | 1.2 | 0.6 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagliara, M.R.; Cecconi, F.; Pasqualetti, P.; Bertoli, M.; Armonaite, K.; Gianni, E.; Grifoni, J.; L’Abbate, T.; Marinozzi, F.; Conti, L.; et al. On the Homology of the Dominant and Non-Dominant Corticospinal Tracts: A Novel Neurophysiological Assessment. Brain Sci. 2023, 13, 278. https://doi.org/10.3390/brainsci13020278
Pagliara MR, Cecconi F, Pasqualetti P, Bertoli M, Armonaite K, Gianni E, Grifoni J, L’Abbate T, Marinozzi F, Conti L, et al. On the Homology of the Dominant and Non-Dominant Corticospinal Tracts: A Novel Neurophysiological Assessment. Brain Sciences. 2023; 13(2):278. https://doi.org/10.3390/brainsci13020278
Chicago/Turabian StylePagliara, Maria Rita, Federico Cecconi, Patrizio Pasqualetti, Massimo Bertoli, Karolina Armonaite, Eugenia Gianni, Joy Grifoni, Teresa L’Abbate, Franco Marinozzi, Livio Conti, and et al. 2023. "On the Homology of the Dominant and Non-Dominant Corticospinal Tracts: A Novel Neurophysiological Assessment" Brain Sciences 13, no. 2: 278. https://doi.org/10.3390/brainsci13020278