
Effective Intracerebral Connectivity in Acute Stroke: A TMS–EEG Study
 , , , and
, , , and
Abstract
:1. Introduction
Aim
2. Methods
2.1. Subjects
- -
- Age 18–90;
- -
- NIH stroke scale (NIHSS) range 6–24;
- -
- Single ischemic stroke in the middle cerebral artery territory of the left hemisphere within 10 days;
- -
- Upper arm paresis (upper arm at least NIHSS > 1).
- -
- Exclusion criteria were:
- -
- Symptom onset more distant than 10 days;
- -
- Associated neurological diseases;
- -
- Multiple ischemic strokes;
- -
- Previous ischemic or hemorrhagic stroke;
- -
- TMS contraindication, according to the recommendations of the International Federation of Clinical Neurophysiology (IFCN, [24]);
- -
- Compromised vigilance or severe hemodynamic, neurological, or respiratory conditions;
- -
- Poor middle cerebral artery insonation through transcranial Doppler;
- -
- Hemodynamic carotid stenosis (it could determine a compensatory dilatation in the distal circulation, with a consequent reduction of basal VMR);
- -
- Refusal to sign the informed consent.
2.2. TMS–EEG Experimental Setup and Protocol
2.3. Data Analysis
2.3.1. MEP and EEG Data
2.3.2. Data-Processing Pipeline for TMS–EEG Data
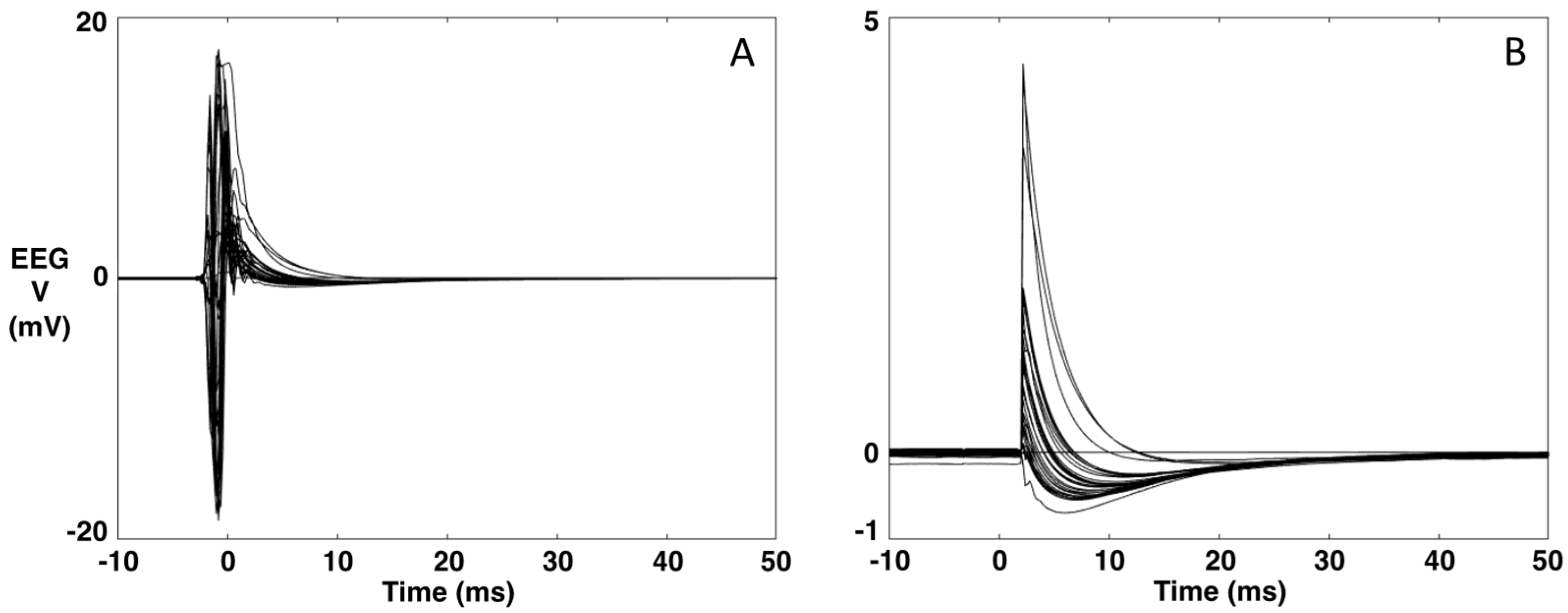
- A typical EEG recording during TMS is presented in Figure 1A,B. The data are averaged over the repeated TMS stimuli (t = 0 ms). As can be seen, the TMS artifact is several orders of magnitude higher than the brain signal. Via software, we simulate the hardware intervention of the sample-and-hold amplifiers [27] (as our TMS system is not equipped with this) by replacing the 5 ms interval around the TMS stimulus (−3 to 2 ms) with the 5 ms interval of the baseline (−8 to −3 ms) (Figure 1).Figure 1. Example of an EEG recording during TMS. (A). Original data averaged over the TMS stimulus (t = 0). (B). The data after replacing the [−3, 2] ms interval with the 5-ms interval of the baseline [−8, −3] ms.Figure 1. Example of an EEG recording during TMS. (A). Original data averaged over the TMS stimulus (t = 0). (B). The data after replacing the [−3, 2] ms interval with the 5-ms interval of the baseline [−8, −3] ms.
![Brainsci 13 00233 g001]()
- A channel that has been minimally corrupted by a TMS stimulus artifact is chosen through visual inspection. The data are re-referenced against this channel. In our case, the Cz channel is the channel that is least corrupted by the TMS artifact.
- Application of the SOurce-Utilized Noise Discarding (SOUND) algorithm for cleaning the channels separately for the baseline and the data after the TMS stimulus [26]. TMS–EEG data can contain various other noise sources that corrupt individual channels or cause strange voltage patterns on the scalp. The SOUND algorithm cross-validates EEG channels via consecutive inverse and forward computations. The cross-validation outputs an estimate for the noise distribution across the EEG channels, which is used to form a spatial Wiener filter that highlights the neuronal EEG signals. SOUND filters out those signal components that are not likely to originate from intracranial post-synaptic currents, e.g., electrode-polarization, line-noise, and electrode-movement artifacts [26,28,29].

- 4.
- Re-referencing the data to the mean reference. At this point, the data are processed by the SSP–SIR (Signal-Space Projection—Source-Informed Reconstruction) algorithm for automatic cleaning of residual artifacts, such as muscle artifacts [28,30]. SSP–SIR substantially improved the signal quality of artifactual TMS–EEG data, causing minimal distortion in the neuronal signal components. In the SSP–SIR approach, the artifact signal subspace containing TMS-evoked muscle artifacts is estimated from the high-pass-filtered (cutoff frequency 100 Hz) data using principal component analysis. The rationale is that EEG signals above 100 Hz mainly consist of non-neuronal signals. The estimated artifact subspace is projected out using SSP [30]. The remaining artifact-suppressed signals are used to estimate an equivalent source distribution model exploiting anatomical brain constraints. When solving the inverse problem, the artifact dimensions must be projected out also from the lead field (or gain) matrix. In the final SIR step, the obtained source estimates are projected back onto the original signal space.
- 5.
- Finally, independent component analysis using the fastICA [31] algorithm is applied to the averaged data to eliminate eye blinking and suppress random noise (first and fourth rows of Figure 3). Using data cleaned by SOUND, which is most efficient in eliminating the TMS and the motor artifacts, we used a deep experience in exploiting statistical features of ocular artifacts, thus well identified by the ICA approach [32,33].
2.3.3. Stroke vs. Healthy Volunteer Comparison
3. Results
3.1. Enrolled Population
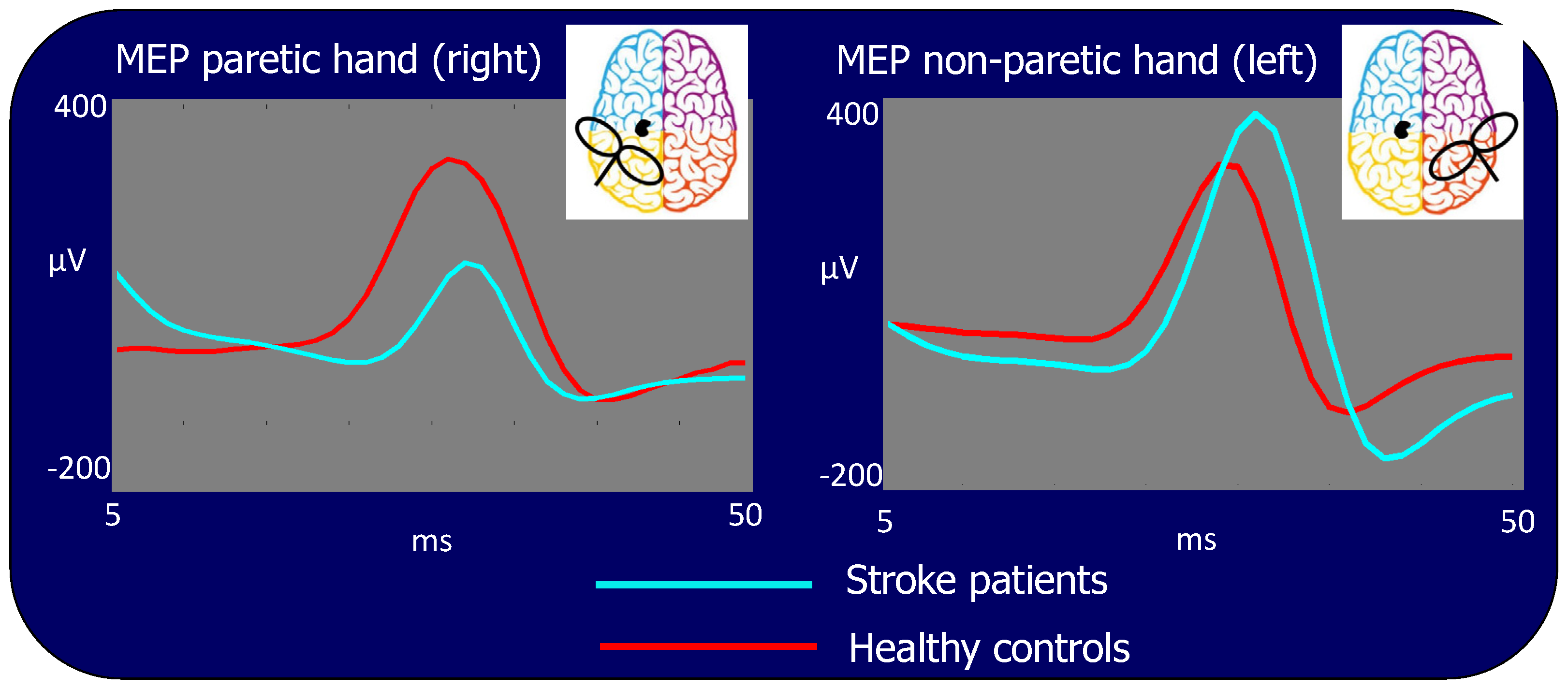
3.2. MEP Amplitude
3.3. Quality of Artefact Removal
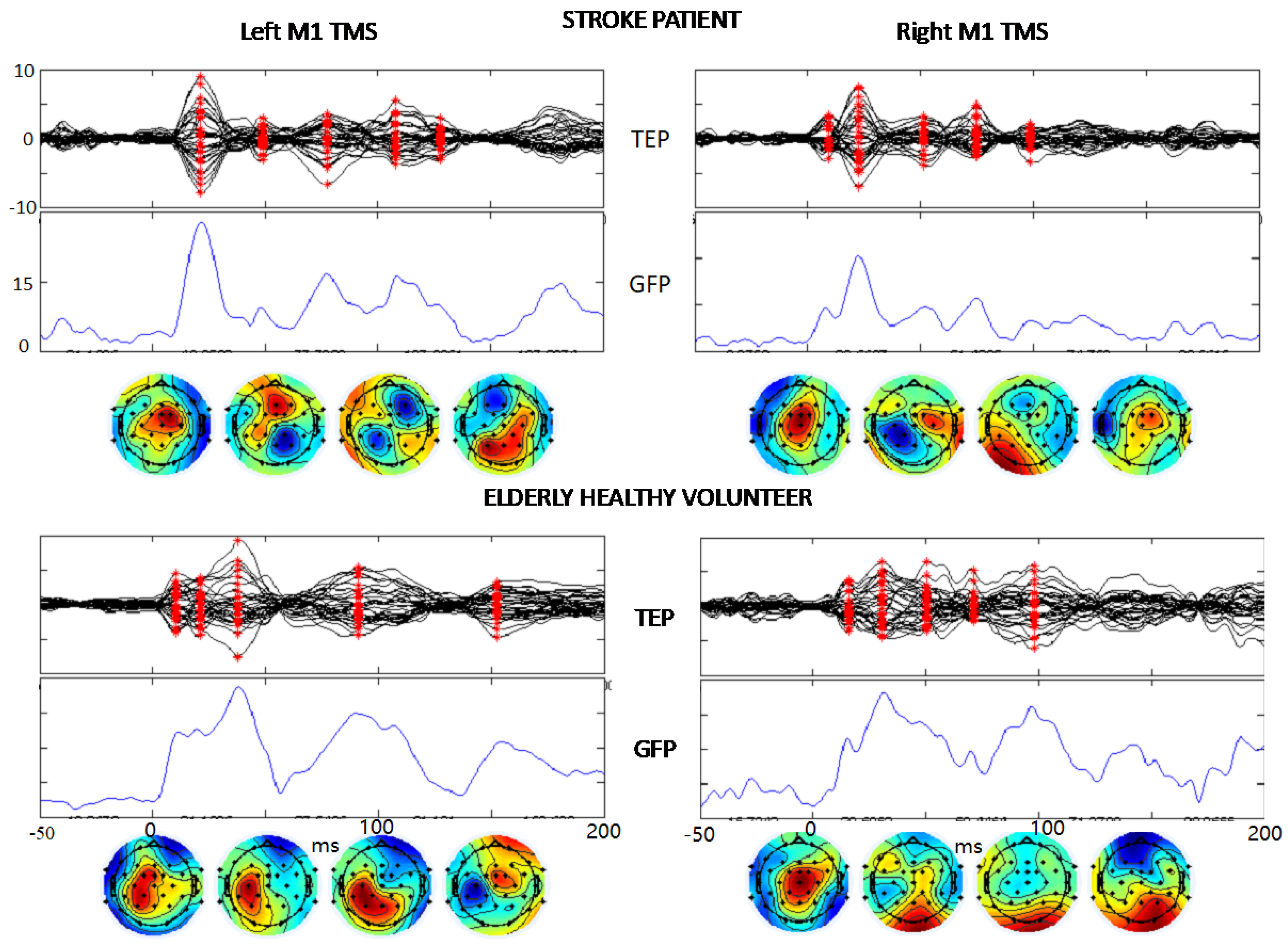
3.4. Global Intracerebral Effective M1 Connectivity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rossini, P.M.; Rossi, S. Transcranial Magnetic Stimulation: Diagnostic, Therapeutic, and Research Potential. Neurology 2007, 68, 484–488. [Google Scholar] [CrossRef]
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-Invasive Magnetic Stimulation of Human Motor Cortex. Lancet 1985, 325, 1106–1107. [Google Scholar] [CrossRef]
- Bertazzoli, G.; Esposito, R.; Mutanen, T.P.; Ferrari, C.; Ilmoniemi, R.J.; Miniussi, C.; Bortoletto, M. The Impact of Artifact Removal Approaches on TMS-EEG Signal. Neuroimage 2021, 239, 118272. [Google Scholar] [CrossRef]
- Belardinelli, P.; König, F.; Liang, C.; Premoli, I.; Desideri, D.; Müller-Dahlhaus, F.; Gordon, P.C.; Zipser, C.; Zrenner, C.; Ziemann, U. TMS-EEG Signatures of Glutamatergic Neurotransmission in Human Cortex. Sci. Rep. 2021, 11, 8159. [Google Scholar] [CrossRef]
- Ferreri, F.; Rossini, P.M. TMS and TMS-EEG Techniques in the Study of the Excitability, Connectivity, and Plasticity of the Human Motor Cortex. Rev. Neurosci. 2013, 24, 431–442. [Google Scholar] [CrossRef]
- Paus, T.; Sipila, P.K.; Strafella, A.P. Synchronization of Neuronal Activity in the Human Primary Motor Cortex by Transcranial Magnetic Stimulation: An EEG Study. J. Neurophysiol. 2001, 86, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Zappasodi, F.; Olejarczyk, E.; Marzetti, L.; Assenza, G.; Pizzella, V.; Tecchio, F. Fractal Dimension of EEG Activity Senses Neuronal Impairment in Acute Stroke. PLoS ONE 2014, 9, e100199. [Google Scholar] [CrossRef] [Green Version]
- Tecchio, F.; Zappasodi, F.; Pasqualetti, P.; Tombini, M.; Salustri, C.; Oliviero, A.; Pizzella, V.; Vernieri, F.; Rossini, P.M. Rhythmic Brain Activity at Rest from Rolandic Areas in Acute Mono-Hemispheric Stroke: A Magnetoencephalographic Study. Neuroimage 2005, 28, 72–83. [Google Scholar] [CrossRef]
- Tecchio, F.; Pasqualetti, P.; Zappasodi, F.; Tombini, M.; Lupoi, D.; Vernieri, F.; Rossini, P.M. Outcome Prediction in Acute Monohemispheric Stroke via Magnetoencephalography. J. Neurol. 2007, 254, 296–305. [Google Scholar] [CrossRef]
- Zappasodi, F.; Croce, P.; Giordani, A.; Assenza, G.; Giannantoni, N.M.; Profice, P.; Granata, G.; Rossini, P.M.; Tecchio, F. Prognostic Value of EEG Microstates in Acute Stroke. Brain Topogr. 2017, 30, 698–710. [Google Scholar] [CrossRef]
- Tecchio, F.; Malosio, M.L.; Zappasodi, F.; Assenza, G.; Squitti, R. Multi-Modal Factors for Recovery Prognosis in Acute Stroke. Aging Clin. Exp. Res. 2021, 33, 1717–1719. [Google Scholar] [CrossRef]
- Zappasodi, F.; Pasqualetti, P.; Rossini, P.M.; Tecchio, F. Acute Phase Neuronal Activity for the Prognosis of Stroke Recovery. Neural Plast. 2019, 2019, 1971875. [Google Scholar] [CrossRef] [Green Version]
- Chiarelli, A.M.; Croce, P.; Assenza, G.; Merla, A.; Granata, G.; Giannantoni, N.M.; Pizzella, V.; Tecchio, F.; Zappasodi, F. Electroencephalography-Derived Prognosis of Functional Recovery in Acute Stroke Through Machine Learning Approaches. Int. J. Neural Syst. 2020, 30, 1971875. [Google Scholar] [CrossRef]
- Heald, A.; Bates, D.; Cartlidge, N.E.F.; French, J.M.; Miller, S. Longitudinal Study of Central Motor Conduction Time Following Stroke. 2. Central Motor Conduction Measured within 72 h after Stroke as a Predictor of Functional Outcome at 12 Months. Brain 1993, 116 (Pt 6), 1371–1385. [Google Scholar] [CrossRef]
- Wang, D.; Xiang, J.; He, Y.; Yuan, M.; Dong, L.; Ye, Z.; Mao, W. The Mechanism and Clinical Application of Constraint-Induced Movement Therapy in Stroke Rehabilitation. Front. Behav. Neurosci. 2022, 16, 211. [Google Scholar] [CrossRef]
- Oliveri, M.; Rossini, P.M.; Traversa, R.; Cicinelli, P.; Filippi, M.M.; Pasqualetti, P.; Tomaiuolo, F.; Caltagirone, C. Left Frontal Transcranial Magnetic Stimulation Reduces Contralesional Extinction in Patients with Unilateral Right Brain Damage. Brain 1999, 122 (Pt 9), 1731–1739. [Google Scholar] [CrossRef] [Green Version]
- Oliveri, M.; Rossini, P.M.; Filippi, M.M.; Traversa, R.; Cicinelli, P.; Palmieri, M.G.; Pasqualetti, P.; Caltagirone, C. Time-Dependent Activation of Parieto-Frontal Networks for Directing Attention to Tactile Space. A Study with Paired Transcranial Magnetic Stimulation Pulses in Right-Brain-Damaged Patients with Extinction. Brain 2000, 123 (Pt 9), 1939–1947. [Google Scholar] [CrossRef] [Green Version]
- Cicinelli, P.; Pasqualetti, P.; Zaccagnini, M.; Traversa, R.; Olivefi, M.; Rossini, P.M. Interhemispheric Asymmetries of Motor Cortex Excitability in the Postacute Stroke Stage: A Paired-Pulse Transcranial Magnetic Stimulation Study. Stroke 2003, 34, 2653–2658. [Google Scholar] [CrossRef]
- Shimizu, T.; Hosaki, A.; Hino, T.; Sato, M.; Komori, T.; Hirai, S.; Rossini, P.M. Motor Cortical Disinhibition in the Unaffected Hemisphere after Unilateral Cortical Stroke. Brain 2002, 125, 1896–1907. [Google Scholar] [CrossRef] [Green Version]
- Bütefisch, C.M. Plasticity in the Human Cerebral Cortex: Lessons from the Normal Brain and from Stroke. Neuroscientist 2004, 10, 163–173. [Google Scholar] [CrossRef]
- di Lazzaro, V.; Profice, P.; Pilato, F.; Capone, F.; Ranieri, F.; Pasqualetti, P.; Colosimo, C.; Pravatã, E.; Cianfoni, A.; Dileone, M. Motor Cortex Plasticity Predicts Recovery in Acute Stroke. Cereb. Cortex 2010, 20, 1523–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, M.A.; Cohen, L.G. Mechanisms Underlying Functional Changes in the Primary Motor Cortex Ipsilateral to an Active Hand. J. Neurosci. 2008, 28, 5631–5640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Avanzini, G.; Bestmann, S.; Berardelli, A.; Brewer, C.; Canli, T.; Cantello, R.; et al. Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Rossini, P.M.; Barker, A.T.; Berardelli, A.; Caramia, M.D.; Caruso, G.; Cracco, R.Q.; Dimitrijević, M.R.; Hallett, M.; Katayama, Y.; Lücking, C.H.; et al. Non-Invasive Electrical and Magnetic Stimulation of the Brain, Spinal Cord and Roots: Basic Principles and Procedures for Routine Clinical Application. Report of an IFCN Committee. Electroencephalogr. Clin. Neurophysiol. 1994, 91, 79–92. [Google Scholar] [CrossRef]
- Pennisi, G.; Rapisarda, G.; Bella, R.; Calabrese, V.; Maertens De Noordhout, A.; Delwaide, P.J. Absence of Response to Early Transcranial Magnetic Stimulation in Ischemic Stroke Patients: Prognostic Value for Hand Motor Recovery. Stroke 1999, 30, 2666–2670. [Google Scholar] [CrossRef] [Green Version]
- Mutanen, T.P.; Metsomaa, J.; Liljander, S.; Ilmoniemi, R.J. Automatic and Robust Noise Suppression in EEG and MEG: The SOUND Algorithm. Neuroimage 2018, 166, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, J.; Ruohonen, J.; Näätänen, R.; Ilmoniemi, R.J. Instrumentation for the Measurement of Electric Brain Responses to Transcranial Magnetic Stimulation. Med. Biol. Eng. Comput. 1999, 37, 322–326. [Google Scholar] [CrossRef]
- Mutanen, T.P.; Kukkonen, M.; Nieminen, J.O.; Stenroos, M.; Sarvas, J.; Ilmoniemi, R.J. Recovering TMS-Evoked EEG Responses Masked by Muscle Artifacts. Neuroimage 2016, 139, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Mutanen, T.P.; Biabani, M.; Sarvas, J.; Ilmoniemi, R.J.; Rogasch, N.C. Source-Based Artifact-Rejection Techniques Available in TESA, an Open-Source TMS-EEG Toolbox. Brain Stimul. 2020, 13, 1349–1351. [Google Scholar] [CrossRef]
- Uusitalo, M.A.; Ilmoniemi, R.J. Signal-Space Projection Method for Separating MEG or EEG into Components. Med. Biol. Eng. Comput. 1997, 35, 135–140. [Google Scholar] [CrossRef]
- Hyvärinen, A. Fast and Robust Fixed-Point Algorithms for Independent Component Analysis. IEEE Trans. Neural. Netw. 1999, 10, 626–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbati, G.; Porcaro, C.; Zappasodi, F.; Rossini, P.M.; Tecchio, F. Optimization of an Independent Component Analysis Approach for Artifact Identification and Removal in Magnetoencephalographic Signals. Clin. Neurophysiol. 2004, 115, 1220–1232. [Google Scholar] [CrossRef] [PubMed]
- Porcaro, C.; Medaglia, M.T.; Krott, A. Removing Speech Artifacts from Electroencephalographic Recordings during Overt Picture Naming. Neuroimage 2015, 105, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.; Skrandies, W. Spatial Analysis of Evoked Potentials in Man-a Review. Prog. Neurobiol. 1984, 23, 227–250. [Google Scholar] [CrossRef]
- Bonato, C.; Miniussi, C.; Rossini, P.M. Transcranial Magnetic Stimulation and Cortical Evoked Potentials: A TMS/EEG Co-Registration Study. Clin. Neurophysiol. 2006, 117, 1699–1707. [Google Scholar] [CrossRef]
- Mochizuki, H.; Huang, Y.Z.; Rothwell, J.C. Interhemispheric Interaction between Human Dorsal Premotor and Contralateral Primary Motor Cortex. J. Physiol. 2004, 561, 331–338. [Google Scholar] [CrossRef]
- Vecchio, F.; Pappalettera, C.; Miraglia, F.; Deinite, G.; Manenti, R.; Judica, E.; Caliandro, P.; Rossini, P.M. Prognostic Role of Hemispherical Functional Connectivity in Stroke: A Study via Graph Theory Versus Coherence of Electroencephalography Rhythms. Stroke 2022. epub ahead of print. [Google Scholar] [CrossRef]
- Grefkes, C.; Fink, G.R. Connectivity-Based Approaches in Stroke and Recovery of Function. Lancet Neurol. 2014, 13, 206–216. [Google Scholar] [CrossRef]
- Duering, M.; Righart, R.; Wollenweber, F.A.; Zietemann, V.; Gesierich, B.; Dichgans, M. Acute Infarcts Cause Focal Thinning in Remote Cortex via Degeneration of Connecting Fiber Tracts. Neurology 2015, 84, 1685–1692. [Google Scholar] [CrossRef] [Green Version]
- Calautti, C.; Baron, J.C. Functional Neuroimaging Studies of Motor Recovery after Stroke in Adults: A Review. Stroke 2003, 34, 1553–1566. [Google Scholar] [CrossRef]
- Dąbrowski, J.; Czajka, A.; Zielińska-Turek, J.; Jaroszyński, J.; Furtak-Niczyporuk, M.; Mela, A.; Poniatowski, Ł.A.; Drop, B.; Dorobek, M.; Barcikowska-Kotowicz, M.; et al. Brain Functional Reserve in the Context of Neuroplasticity after Stroke. Neural Plast. 2019, 2019, 9708905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traversa, R.; Cicinelli, P.; Bassi, A.; Rossini, P.M.; Bernardi, G. Mapping of Motor Cortical Reorganization after Stroke. A Brain Stimulation Study with Focal Magnetic Pulses. Stroke 1997, 28, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Liepert, J.; Storch, P.; Fritsch, A.; Weiller, C. Motor Cortex Disinhibition in Acute Stroke. Clin. Neurophysiol. 2000, 111, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Mahan, M.Y.; Georgopoulos, A.P. Motor Directional Tuning across Brain Areas: Directional Resonance and the Role of Inhibition for Directional Accuracy. Front. Neural Circuits 2013, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, A.P.; Carpenter, A.F. Coding of Movements in the Motor Cortex. Curr. Opin. Neurobiol. 2015, 33, 34–39. [Google Scholar] [CrossRef]
- Kolasinski, J.; Logan, J.P.; Hinson, E.L.; Makin, T.R.; Emir, U.E.; Stagg Correspondence, C.J. A Mechanistic Link from GABA to Cortical Architecture and Perception. Curr. Biol. 2017, 27, 1685–1691.e3. [Google Scholar] [CrossRef] [Green Version]
- Merchant, H.; Naselaris, T.; Georgopoulos, A.P. Dynamic Sculpting of Directional Tuning in the Primate Motor Cortex during Three-Dimensional Reaching. J. Neurosci. 2008, 28, 9164–9172. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, A.P.; Stefanis, C.N. The Motor Cortical Circuit. In Handbook of Brain Microcircuits; Oxford University Press: Oxford, UK, 2013; pp. 39–45. [Google Scholar]
- Carson, R.G. Inter-Hemispheric Inhibition Sculpts the Output of Neural Circuits by Co-Opting the Two Cerebral Hemispheres. J. Physiol. 2020, 598, 4781–4802. [Google Scholar] [CrossRef]
- Daskalakis, Z.J.; Christensen, B.K.; Fitzgerald, P.B.; Roshan, L.; Chen, R. The Mechanisms of Interhemispheric Inhibition in the Human Motor Cortex. J. Physiol. 2002, 543, 317–326. [Google Scholar] [CrossRef]
- Rokni, U.; Steinberg, O.; Vaadia, E.; Sompolinsky, H. Cortical Representation of Bimanual Movements. J. Neurosci. 2003, 23, 11577–11586. [Google Scholar] [CrossRef]
- Dodd, K.C.; Nair, V.A.; Prabhakaran, V. Role of the Contralesional vs. Ipsilesional Hemisphere in Stroke Recovery. Front. Hum. Neurosci. 2017, 11, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murase, N.; Duque, J.; Mazzocchio, R.; Cohen, L.G. Influence of Interhemispheric Interactions on Motor Function in Chronic Stroke. Ann. Neurol. 2004, 55, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Rogasch, N.C.; Hoy, K.E.; Fitzgerald, P.B. Measuring Brain Stimulation Induced Changes in Cortical Properties Using TMS-EEG. Brain Stimul. 2015, 8, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, L.; di Santo, A.; Brown, K.; Ibáñez, J.; Casula, E.; Rawji, V.; di Lazzaro, V.; Koch, G.; Rothwell, J. Disentangling EEG Responses to TMS Due to Cortical and Peripheral Activations. Brain Stimul. 2021, 14, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Rostami, M.; Zomorrodi, R.; Rostami, R.; Hosseinzadeh, G.A. Impact of Methodological Variability on EEG Responses Evoked by Transcranial Magnetic Stimulation: A Meta-Analysis. Clin. Neurophysiol. 2022, 142, 154–180. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wave | Topography | Functional Connectivity | Synaptic Substrate |
|---|---|---|---|
| N7 | F3 | ipsi-lateral motor associative, PM | NMDA |
| P13 | Fp2 F4 F8 C4 T4 T6 | contra-lateral homolog M1 | GABAA |
| N18 | P3 | ipsi-lateral PPC | |
| P30 | ExtendedC | contra-lateral thalamo-cortical nodes | |
| N44 | ExtendedC&I | ipsi- and contral-lateral M1 | |
| P60 | T5 | GABAB | |
| N100 | ExtendedI | ipsi-lateral M1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tecchio, F.; Giambattistelli, F.; Porcaro, C.; Cottone, C.; Mutanen, T.P.; Pizzella, V.; Marzetti, L.; Ilmoniemi, R.J.; Vernieri, F.; Rossini, P.M. Effective Intracerebral Connectivity in Acute Stroke: A TMS–EEG Study. Brain Sci. 2023, 13, 233. https://doi.org/10.3390/brainsci13020233
Tecchio F, Giambattistelli F, Porcaro C, Cottone C, Mutanen TP, Pizzella V, Marzetti L, Ilmoniemi RJ, Vernieri F, Rossini PM. Effective Intracerebral Connectivity in Acute Stroke: A TMS–EEG Study. Brain Sciences. 2023; 13(2):233. https://doi.org/10.3390/brainsci13020233
Chicago/Turabian StyleTecchio, Franca, Federica Giambattistelli, Camillo Porcaro, Carlo Cottone, Tuomas P. Mutanen, Vittorio Pizzella, Laura Marzetti, Risto J. Ilmoniemi, Fabrizio Vernieri, and Paolo Maria Rossini. 2023. "Effective Intracerebral Connectivity in Acute Stroke: A TMS–EEG Study" Brain Sciences 13, no. 2: 233. https://doi.org/10.3390/brainsci13020233