

Low Levels of Adenosine and GDNF Are Potential Risk Factors for Parkinson’s Disease with Sleep Disorders

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Participants
2.1.1. Ethics Approval
2.1.2. Inclusion and Exclusion Criteria
- Aged ≥ 30 years;
- Normal listening, understanding and language expression, and could cooperate with clinicians in all neuropsychological, motor and mental behavior assessments;
- Patient or a legal representative signed the informed consent form.
- Other neurological diseases shown by imaging examination (CT or MRI), including various dementias, stroke, brain infection, tumor, or severe brain injury;
- Secondary Parkinsonian syndrome (PS) or Parkinson-plus syndrome (P-PS) caused by trauma, infection, tumor, or drugs;
- Severe systemic diseases (such as liver and kidney failure, circulatory system or blood system disorders);
- Severe mental illness (such as schizophrenia, or bipolar disorder);
- Other diseases that can cause sleep disorders (e.g., chronic poisoning, cancer pain, endocrine diseases, or urinary system diseases);
- Incomplete data.
- Understood the content and purpose of this study, willingly engaged in this study, and signed the informed consent form;
- Able to complete all scale assessments under physician guidance;
- Those with a Pittsburgh Sleep Quality Index (PSQI) ≤ 7 points.
- Long-term alcoholism, smoking, drug abuse, or bad habits that seriously affect health (e.g., gambling addiction, unprotected sex, and sexually transmitted diseases);
- A history of mental illness or sedative use;
- Incomplete data.
2.2. Sample Collection and Testing
2.3. Medical Data Gathering
2.4. Statistical Analysis
3. Results
3.1. Demographic Characteristics
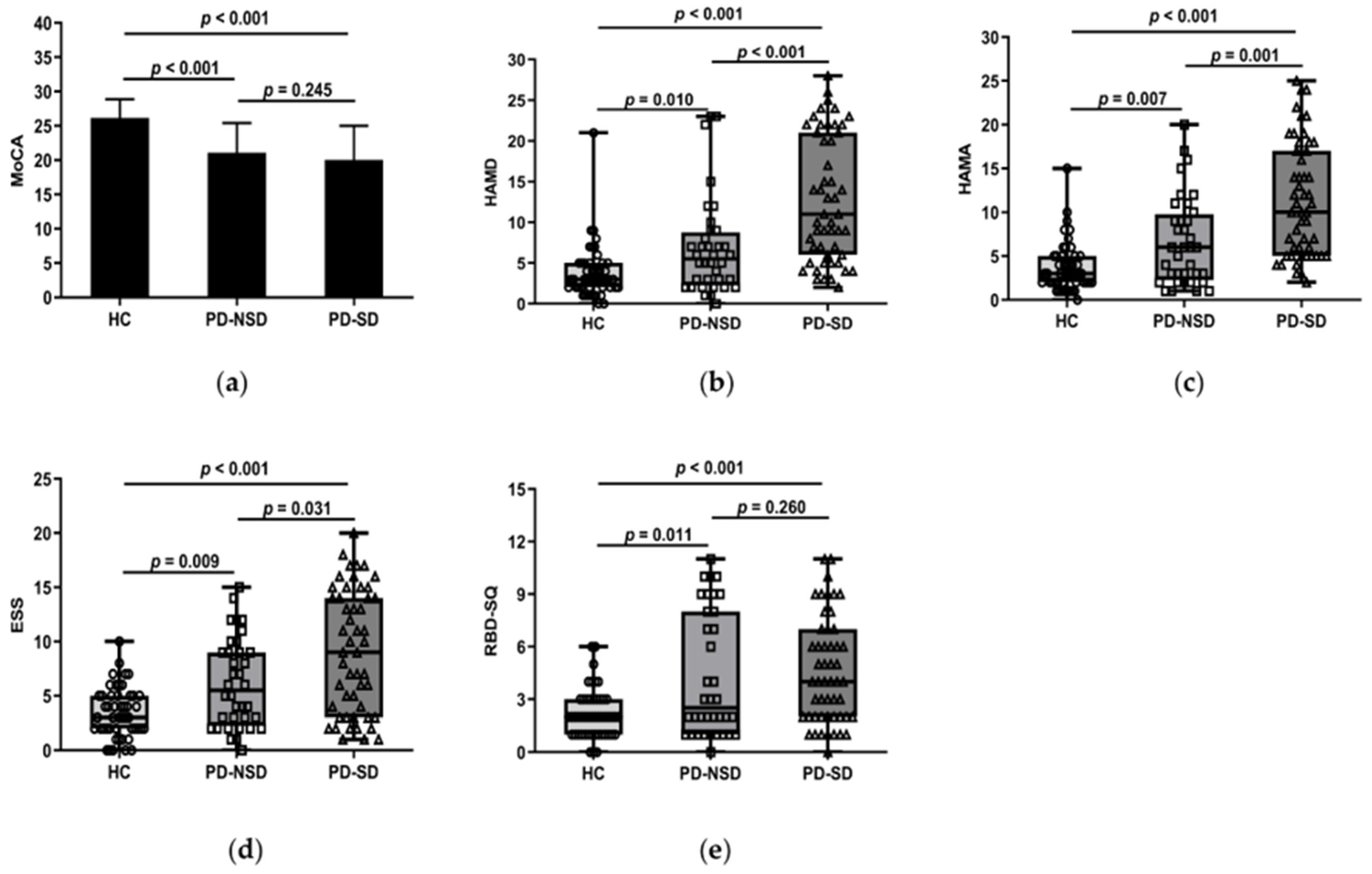
3.2. Non-Motor Symptoms and Sleep Disorders
3.2.1. Cognitive and Sleep Disorders
3.2.2. Anxiety, Depression, and Sleep Disorders
3.2.3. Constipation and Sleep Disorders
3.2.4. Comparison of PDSS, ESS, RBD-SQ, and RLS among Groups
3.3. Motor Symptoms and Sleep Disorders
3.4. Antiparkinsonian Medications between PD Groups
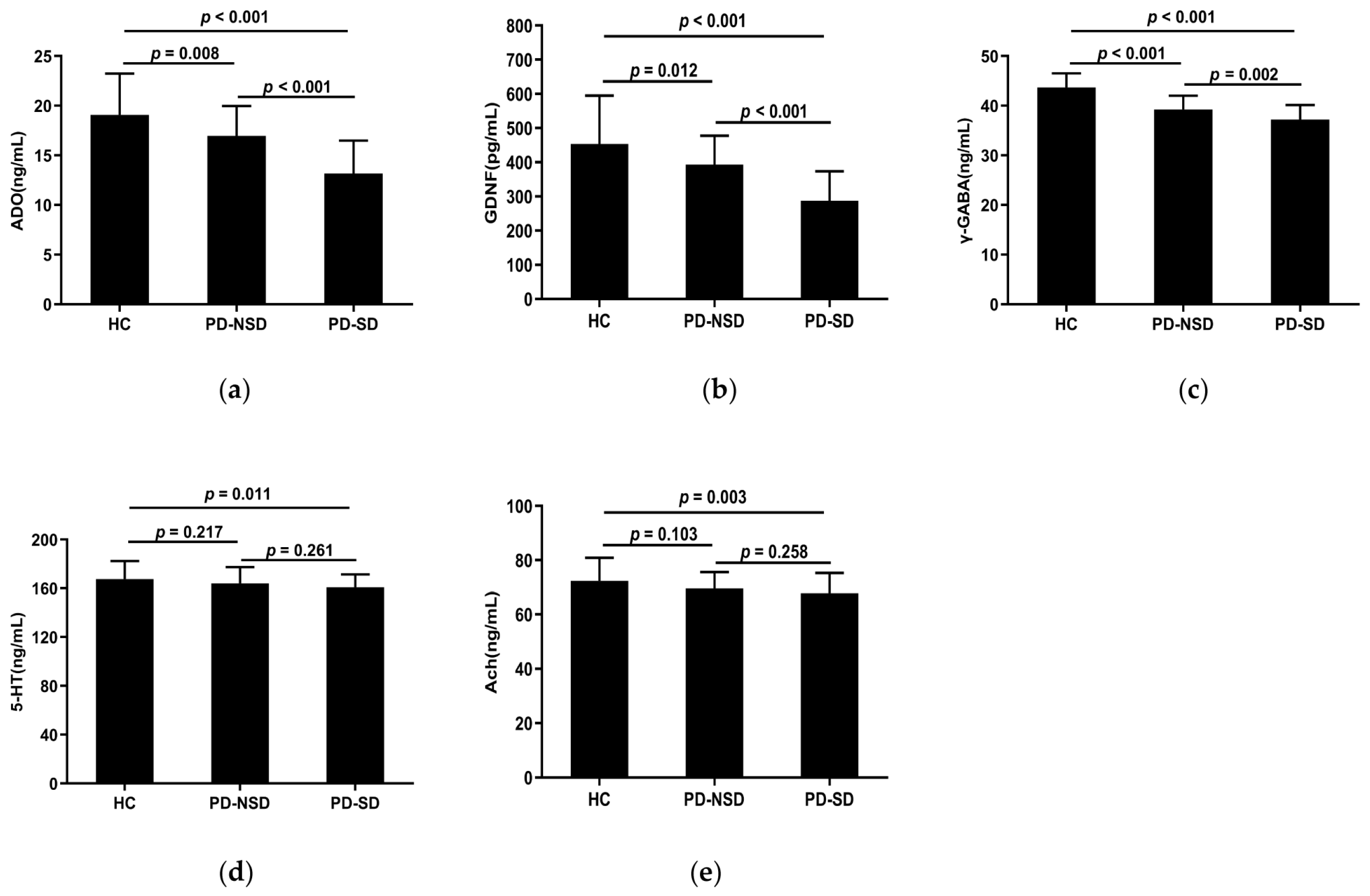
3.5. Levels of Serum Adenosine, GDNF, and Related Neurotransmitters in Each Group
3.6. Correlation Analysis of Each Index and the PSQI Scale in PD Patients
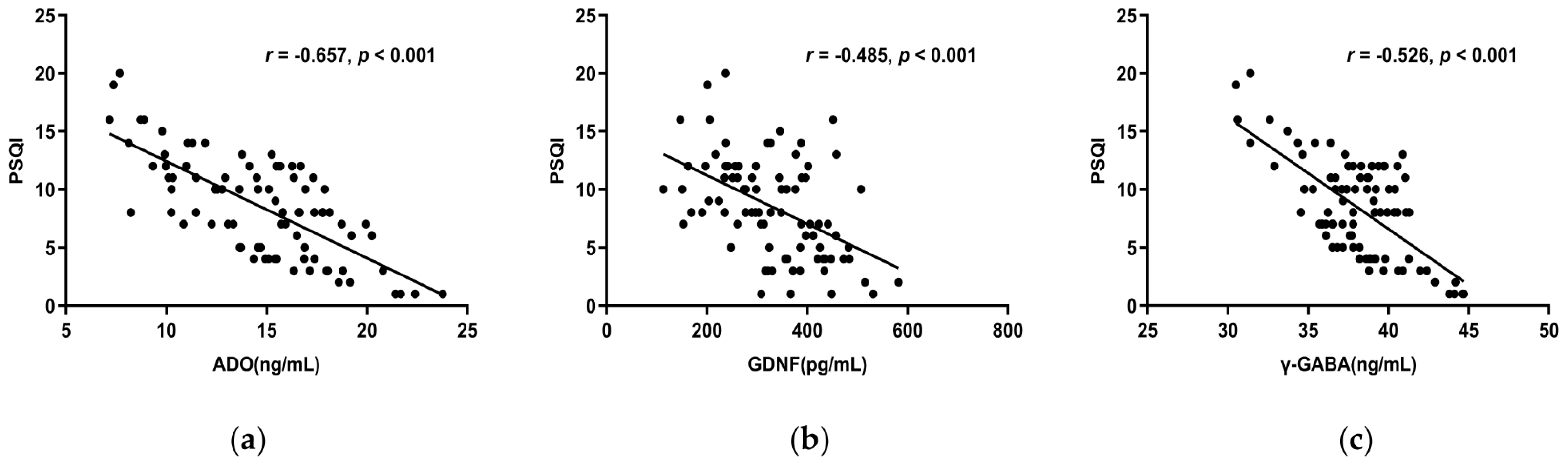
3.6.1. The Correlations between ADO, GDNF, γ-GABA, 5-HT, Ach, and the PSQI Scale
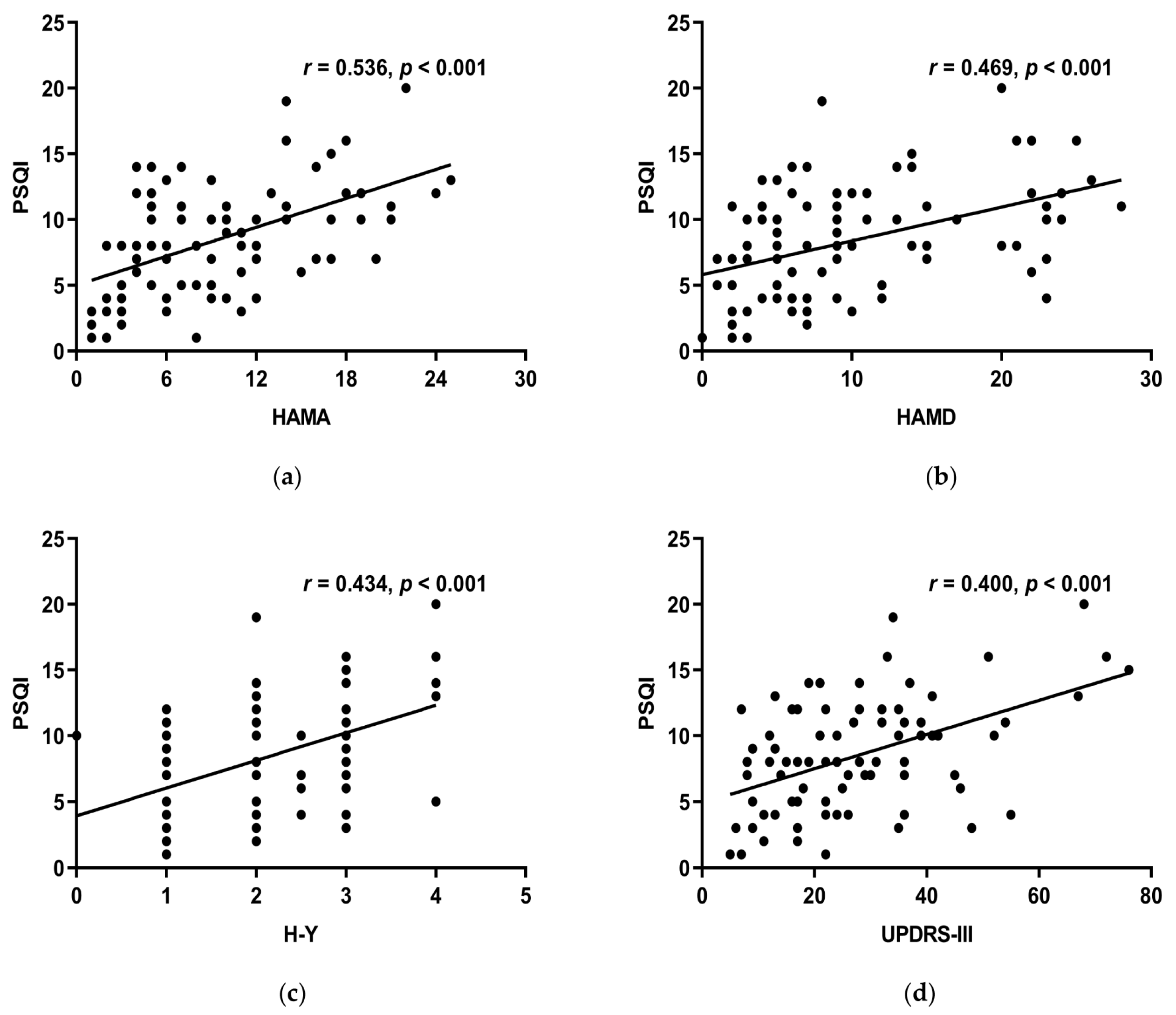
3.6.2. The Correlations between MoCA, HAMA, HAMD, UPDRS-III, H-Y, Course of the Disease, LED, RBD-SQ, and the PSQI Scale
3.7. Risk Factors of Sleep Disturbances
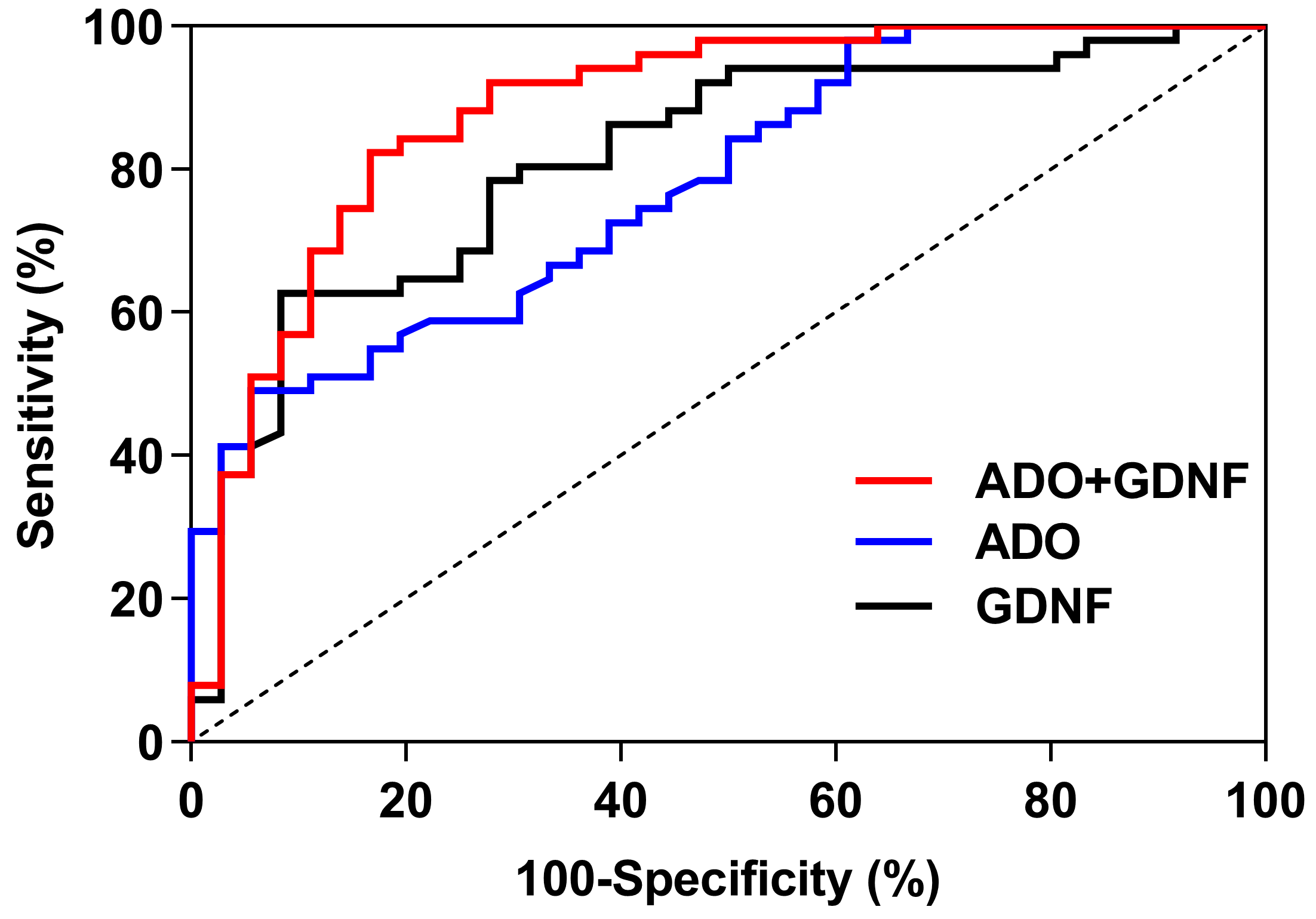
3.8. The ROC Curve Analysis of Serum ADO and GDNF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madetko, N.; Migda, B.; Alster, P.; Turski, P.; Koziorowski, D.; Friedman, A. Platelet-to-lymphocyte ratio and neutrophil-to-lymphocyte ratio may reflect differences in PD and MSA-P neuroinflammation patterns. Neurol. Neurochir. Pol. 2022, 56, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Von Stockum, S.; Nardin, A.; Schrepfer, E.; Ziviani, E. Mitochondrial dynamics and mitophagy in Parkinson’s disease: A fly point of view. Neurobiol. Dis. 2016, 90, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ingiosi, A.M.; Opp, M.R.; Krueger, J.M. Sleep and immune function: Glial contributions and consequences of aging. Curr. Opin. Neurobiol. 2013, 23, 806–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastião, A.M.; Ribeiro, J.A. Adenosine A2 receptor-mediated excitatory actions on the nervous system. Prog. Neurobiol. 1996, 48, 167–189. [Google Scholar] [CrossRef] [PubMed]
- Feldberg, W.; Sherwood, S.L. Injections of drugs into the lateral ventricle of the cat. J. Physiol. 1954, 123, 148–167. [Google Scholar] [CrossRef]
- Portas, C.M.; Thakkar, M.; Rainnie, D.G.; Greene, R.W.; McCarley, R.W. Role of adenosine in behavioral state modulation: A microdialysis study in the freely moving cat. Neuroscience 1997, 79, 225–235. [Google Scholar] [CrossRef]
- Benington, J.H.; Heller, H.C. Restoration of brain energy metabolism as the function of sleep. Prog. Neurobiol. 1995, 45, 347–360. [Google Scholar] [CrossRef]
- Dunwiddie, T.V.; Worth, T. Sedative and anticonvulsant effects of adenosine analogs in mouse and rat. J. Pharmacol. Exp. Ther. 1982, 220, 70–76. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar]
- Snyder, S.H.; Katims, J.J.; Annau, Z.; Bruns, R.F.; Daly, J.W. Adenosine receptors and behavioral actions of methylxanthines. Proc. Natl. Acad. Sci. USA 1981, 78, 3260–3264. [Google Scholar] [CrossRef]
- Bjorness, T.E.; Dale, N.; Mettlach, G.; Sonneborn, A.; Sahin, B.; Fienberg, A.A.; Yanagisawa, M.; Bibb, J.A.; Greene, R.W. An Adenosine-Mediated Glial-Neuronal Circuit for Homeostatic Sleep. J. Neurosci. 2016, 36, 3709–3721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palchykova, S.; Winsky-Sommerer, R.; Shen, H.Y.; Boison, D.; Gerling, A.; Tobler, I. Manipulation of adenosine kinase affects sleep regulation in mice. J. Neurosci. 2010, 30, 13157–13165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Huang, Z.L.; Fredholm, B.B.; Urade, Y.; Hayaishi, O. Adenosine in the tuberomammillary nucleus inhibits the histaminergic system via A1 receptors and promotes non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2008, 105, 19992–19997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, M.; Dale, N. Activity-dependent release of adenosine: A critical re-evaluation of mechanism. Curr. Neuropharmacol. 2008, 6, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scammell, T.E.; Gerashchenko, D.Y.; Mochizuki, T.; McCarthy, M.T.; Estabrooke, I.V.; Sears, C.A.; Saper, C.B.; Urade, Y.; Hayaishi, O. An adenosine A2a agonist increases sleep and induces Fos in ventrolateral preoptic neurons. Neuroscience 2001, 107, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Greene, R.W. Adenosine and Sleep. Curr. Neuropharmacol. 2009, 7, 238–245. [Google Scholar] [CrossRef]
- Basheer, R.; Strecker, R.E.; Thakkar, M.M.; McCarley, R.W. Adenosine and sleep-wake regulation. Prog. Neurobiol. 2004, 73, 379–396. [Google Scholar] [CrossRef]
- Yamagata, K.; Hakata, K.; Maeda, A.; Mochizuki, C.; Matsufuji, H.; Chino, M.; Yamori, Y. Adenosine induces expression of glial cell line-derived neurotrophic factor (GDNF) in primary rat astrocytes. Neurosci. Res. 2007, 59, 467–474. [Google Scholar] [CrossRef]
- Gerlai, R.; McNamara, A.; Choi-Lundberg, D.L.; Armanini, M.; Ross, J.; Powell-Braxton, L.; Phillips, H.S. Impaired water maze learning performance without altered dopaminergic function in mice heterozygous for the GDNF mutation. Eur. J. Neurosci. 2001, 14, 1153–1163. [Google Scholar] [CrossRef]
- Zhang, J.; Tan, H.; Jiang, W.; Zuo, Z. Amantadine alleviates postoperative cognitive dysfunction possibly by increasing glial cell line-derived neurotrophic factor in rats. Anesthesiology 2014, 121, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Kushikata, T.; Kubota, T.; Fang, J.; Krueger, J.M. Glial cell line-derived neurotrophic factor promotes sleep in rats and rabbits. Am. J. Physiol. 2001, 280, R1001–R1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef]
- Fahn, S.; Parkinson Study Group. Does levodopa slow or hasten the rate of progression of Parkinson’s disease? J. Neurol. 2005, 252, IV37–IV42. [Google Scholar] [CrossRef]
- Hoehn, M.M.; Yahr, M.D. Parkinsonism: Onset, progression, and mortality. Neurology 1967, 17, 427–442. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, C.L.; Stowe, R.; Patel, S.; Rick, C.; Gray, R.; Clarke, C.E. Systematic review of levodopa dose equivalency reporting in Parkinson’s disease. Mov. Disord. 2010, 25, 2649–2653. [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Martinez-Martin, P.; Brown, R.G.; Sethi, K.; Stocchi, F.; Odin, P.; Ondo, W.; Abe, K.; Macphee, G.; Macmahon, D.; et al. The metric properties of a novel non-motor symptoms scale for Parkinson’s disease: Results from an international pilot study. Mov. Disord. 2007, 22, 1901–1911. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A Brief Screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Torbey, E.; Pachana, N.A.; Dissanayaka, N.N. Depression rating scales in Parkinson’s disease: A critical review updating recent literature. J. Affect. Disord. 2015, 184, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M. The assessment of anxiety states by rating. Br. J. Med. Psychol. 1959, 32, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Stiasny-Kolster, K.; Mayer, G.; Schäfer, S.; Möller, J.C.; Heinzel-Gutenbrunner, M.; Oertel, W.H. The REM sleep behavior disorder screening questionnaire--a new diagnostic instrument. Mov. Disord. 2007, 22, 2386–2393. [Google Scholar] [CrossRef] [PubMed]
- Johns, M.W. A New Method for Measuring Daytime Sleepiness: The Epworth Sleepiness Scale. Sleep 1991, 14, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.P.; Picchietti, D.L.; Garcia-Borreguero, D.; Ondo, W.G.; Walters, A.S.; Winkelman, J.W.; Zucconi, M.; Ferri, R.; Trenkwalder, C.; Lee, H.B.; et al. Restless legs syndrome/Willis-Ekbom disease diagnostic criteria: Updated International Restless Legs Syndrome Study Group (IRLSSG) consensus criteria--history, rationale, description, and significance. Sleep Med. 2014, 15, 860–873. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A.; Hasler, W.L. Rome IV-Functional GI Disorders: Disorders of Gut-Brain Interaction INTRODUCTION. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Liu, X.C.; Tang, M.Q.; Hu, L.; Wang, A.Z.; Wu, H.X.; Zhao, G.F.; Gao, C.N.; Li, W.S. Reliability and validity of the Pittsburgh sleep quality index. Chin. J. Psychiatry. 1996, 29, 103–107. (In Chinese) [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Pal, S.; DiMarco, A.; Whately-Smith, C.; Bridgman, K.; Mathew, R.; Pezzela, F.R.; Forbes, A. The Parkinson’s disease sleep scale: A new instrument for assessing sleep and nocturnal disability in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2002, 73, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Horsager, J.; Andersen, K.B.; Knudsen, K.; Skjaerbaek, C.; Fedorova, T.D.; Okkels, N.; Schaeffer, E.; Bonkat, S.K.; Geday, J.; Otto, M.; et al. Brain-first versus body-first Parkinson’s disease: A multimodal imaging case-control study. Brain 2020, 143, 3077–3088. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Braak, H.; Sastre, M.; Del Tredici, K. Development of alpha-synuclein immunoreactive astrocytes in the forebrain parallels stages of intraneuronal pathology in sporadic Parkinson’s disease. Acta Neuropathol. 2007, 114, 231–241. [Google Scholar] [CrossRef]
- Liu, M.; Qin, L.; Wang, L.; Tan, J.; Zhang, H.; Tang, J.; Shen, X.; Tan, L.; Wang, C. alphasynuclein induces apoptosis of astrocytes by causing dysfunction of the endoplasmic reticulumGolgi compartment. Mol. Med. Rep. 2018, 18, 322–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Agnaf, O.M.; Salem, S.A.; Paleologou, K.E.; Curran, M.D.; Gibson, M.J.; Court, J.A.; Schlossmacher, M.G.; Allsop, D. Detection of oligomeric forms of alpha-synuclein protein in human plasma as a potential biomarker for Parkinson’s disease. FASEB J. 2006, 20, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Patel, S.; Lee, S.J. Intravesicular localization and exocytosis of alpha-synuclein and its aggregates. J. Neurosci. 2005, 25, 6016–6024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halassa, M.M. Thalamocortical dynamics of sleep: Roles of purinergic neuromodulation. Semin. Cell Dev. Biol. 2011, 22, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Blutstein, T.; Haydon, P.G. The Importance of astrocyte-derived purines in the modulation of sleep. Glia 2013, 61, 129–139. [Google Scholar] [CrossRef]
- Cunha, R.A. Different cellular sources and different roles of adenosine: A1 receptor-mediated inhibition through astrocytic-driven volume transmission and synapse-restricted A2A receptor-mediated facilitation of plasticity. Neurochem. Int. 2008, 52, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Van Dort, C.J.; Baghdoyan, H.A.; Lydic, R. Adenosine A(1) and A(2A) receptors in mouse prefrontal cortex modulate acetylcholine release and behavioral arousal. J. Neurosci. 2009, 29, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Tong, Q.; Xu, Q.; Xia, Q.; Yuan, Y.; Zhang, L.; Sun, H.; Shan, H.; Zhang, K. Correlations between plasma levels of amino acids and nonmotor symptoms in Parkinson’s disease. J. Neural Transm. 2015, 122, 411–417. [Google Scholar] [CrossRef]
- Weber, F.; Chung, S.; Beier, K.T.; Xu, M.; Luo, L.; Dan, Y. Control of REM sleep by ventral medulla GABAergic neurons. Nature 2015, 526, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.J.; Lydic, R.; Baghdoyan, H.A. Sleep and GABA levels in the oral part of rat pontine reticular formation are decreased by local and systemic administration of morphine. Neuroscience 2007, 144, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Fell, M.J.; Flik, G.; Dijkman, U.; Folgering, J.H.; Perry, K.W.; Johnson, B.J.; Westerink, B.H.; Svensson, K.A. Glutamatergic regulation of brain histamine neurons: In vivo microdialysis and electrophysiology studies in the rat. Neuropharmacology 2015, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, K.D.; Irwin, I.; Valverde, J.; Brennan, T.J.; Langston, J.W.; Hefti, F. GDNF Induces a Dystonia-like State in Neonatal Rats and Stimulates Dopamine and Serotonin Synthesis. Neuron 1996, 16, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Borreguero, D.; Larrosa, O.; Bravo, M. Parkinson’s disease and sleep. Sleep Med. Rev. 2003, 7, 115–129. [Google Scholar] [CrossRef]
- Monaca, C.; Laloux, C.; Jacquesson, J.M.; Gele, P.; Marechal, X.; Bordet, R.; Destee, A.; Derambure, P. Vigilance states in a parkinsonian model, the MPTP mouse. Eur. J. Neurosci. 2004, 20, 2474–2478. [Google Scholar] [CrossRef]
- Decker, M.J.; Keating, G.; Hue, G.E.; Freeman, A.; Rye, D.D. Mesolimbic dopamine’s modulation of REM sleep. J. Sleep Res. 2002, 51, 51–52. [Google Scholar]
- Rye, D.B.; Bliwise, D.L.; Dihenia, B.; Gurecki, P. Daytime sleepiness in Parkinson’s disease. J. Sleep Res. 2000, 9, 63–69. [Google Scholar] [CrossRef]
- Jouvet, M. The role of monoamines and acetylcholine-containing neurons in the regulation of the sleep-waking cycle. Ergeb. Physiol. 1972, 64, 166–307. [Google Scholar] [CrossRef]
- Hipólide, D.C.; Moreira, K.M.; Barlow, K.B.; Wilson, A.A.; Nobrega, J.N.; Tufik, S. Distinct effects of sleep deprivation on binding to norepinephrine and serotonin transporters in rat brain. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 297–303. [Google Scholar] [CrossRef]
- Thannickal, T.C.; Lai, Y.Y.; Siegel, J.M. Hypocretin (orexin) cell loss in Parkinson’s disease. Brain 2007, 130, 1586–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fronczek, R.; Overeem, S.; Lee, S.Y.; Hegeman, I.M.; van Pelt, J.; van Duinen, S.G.; Lammers, G.J.; Swaab, D.F. Hypocretin (orexin) loss in Parkinson’s disease. Brain 2007, 130, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Drouot, X.; Moutereau, S.; Nguyen, J.P.; Lefaucheur, J.P.; Creange, A.; Remy, P.; Goldenberg, F.; d’Ortho, M.P. Low levels of ventricular CSF orexin/hypocretin in advanced PD. Neurology 2003, 61, 540–543. [Google Scholar] [CrossRef]
- Wienecke, M.; Werth, E.; Poryazova, R.; Baumann-Vogel, H.; Bassetti, C.L.; Weller, M.; Waldvogel, D.; Storch, A.; Baumann, C.R. Progressive dopamine and hypocretin deficiencies in Parkinson’s disease: Is there an impact on sleep and wakefulness? J. Sleep Res. 2012, 21, 710–717. [Google Scholar] [CrossRef]
- Kalaitzakis, M.E.; Gentleman, S.M.; Pearce, R.K. Disturbed sleep in Parkinson’s disease: Anatomical and pathological correlates. Neuropathol. Appl. Neurobiol. 2013, 39, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Bubser, M.; Fadel, J.R.; Jackson, L.L.; Meador-Woodruff, J.H.; Jing, D.; Deutch, A.Y. Dopaminergic regulation of orexin neurons. Eur. J. Neurosci. 2005, 21, 2993–3001. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, L.L.; Park, D.; Zhu, L.; Palmer, M.R.; Broadhurst, R.Y.; Arrigoni, E. Regulation of Lateral Hypothalamic Orexin Activity by Local GABAergic Neurons. J. Neurosci. 2018, 38, 1588–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Yamanaka, A. Optogenetic activation of serotonergic terminals facilitates GABAergic inhibitory input to orexin/hypocretin neurons. Sci. Rep. 2016, 6, 36039. [Google Scholar] [CrossRef]
- Ogawa, T.; Kajiyama, Y.; Ishido, H.; Chiba, S.; Revankar, G.S.; Nakano, T.; Taniguchi, S.; Kanbayashi, T.; Ikenaka, K.; Mochizuki, H. Decreased cerebrospinal fluid orexin levels not associated with clinical sleep disturbance in Parkinson’s disease: A retrospective study. PLoS ONE 2022, 17, e0279747. [Google Scholar] [CrossRef]
- Chang, C.W.; Fan, J.Y.; Chang, B.L.; Wu, Y.R. Anxiety and Levodopa Equivalent Daily Dose Are Potential Predictors of Sleep Quality in Patients With Parkinson Disease in Taiwan. Front. Neurol. 2019, 10, 340. [Google Scholar] [CrossRef]
- Parkinson Study Group. Pramipexole vs Levodopa as Initial Treatment for Parkinson Disease: A Randomized Controlled Trial. JAMA 2000, 284, 1931–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comella, C.L. Daytime sleepiness, agonist therapy, and driving in Parkinson disease. JAMA 2002, 287, 509–511. [Google Scholar] [CrossRef] [PubMed]
- Ratti, P.L.; Nègre-Pagès, L.; Pérez-Lloret, S.; Manni, R.; Damier, P.; Tison, F.; Destée, A.; Rascol, O. Subjective sleep dysfunction and insomnia symptoms in Parkinson’s disease: Insights from a cross-sectional evaluation of the French CoPark cohort. Parkinsonism Relat. Disord. 2015, 21, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Kay, D.B.; Tanner, J.J.; Bowers, D. Sleep disturbances and depression severity in patients with Parkinson’s disease. Brain Behav. 2018, 8, e00967. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Information | HC (n = 49) | PD-NSD (n = 36) | PD-SD (n = 51) | Statistic | p |
|---|---|---|---|---|---|
| Age (years) | 65.57 ± 8.85 | 66.86 ± 8.9 | 67.57 ± 8.75 | 0.652 a | 0.523 |
| Gender (%) | 0.279 b | 0.870 | |||
| Male | 25 (51.0) | 17 (47.2) | 27 (52.9) | ||
| Female | 24 (49.0) | 19 (52.8) | 24 (47.1) | ||
| Education (years) | 7.00 (6.00, 9.00) | 7.50 (6.00, 9.00) | 7.00 (6.00, 9.00) | 0.029 c | 0.985 |
| BMI (kg/m2) | 24.96 ± 3.19 | 24.07 ± 2.76 | 24.21 ± 3.16 | 1.119 a | 0.330 |
| Smoking history (%) | 6 (12.2) | 6 (16.7) | 10 (19.6) | 1.008 b | 0.604 |
| Alcohol consumption (%) | 9 (18.4) | 7 (19.4) | 10 (19.6) | 0.028 b | 0.986 |
| MoCA (scores) | 26.18 ± 2.71 | 21.08 ± 4.32 * | 20.04 ± 4.97 * | 31.021 a | <0.001 |
| HAMD (scores) | 3.00 (2.00, 5.00) | 5.50 (2.25, 8.75) * | 11.00 (6.00, 21.00) *,# | 47.003 c | <0.001 |
| HAMA (scores) | 3.00 (2.00, 5.00) | 6.00 (2.25, 9.75) * | 10.00 (5.00, 17.00) *,# | 45.217 c | <0.001 |
| RBD-SQ (scores) | 2.00 (1.00, 3.00) | 2.50 (1.00, 8.00) * | 4.00 (2.00, 7.00) * | 16.746 c | <0.001 |
| ESS (scores) | 3.00 (2.00, 5.00) | 5.50 (2.25, 9.00) * | 9.00 (3.00, 14.00) *,# | 27.448 c | <0.001 |
| RLS, case (%) | 3 (6.1) | 6 (16.7) | 13 (25.5) * | 6.922 b | 0.031 |
| Constipation, case (%) | 14 (28.6) | 21 (58.3) * | 31(60.8) * | 12.266 b | 0.002 |
| UPDRS-III (scores) | NA | 22.67 ± 12.97 | 30.98 ± 16.26 | 2.547 d | 0.013 |
| NMSS (scores) | NA | 21.00 (15.25, 64.00) | 41.00 (20.00, 74.00) | −1.599 e | 0.110 |
| Phenotype (%) | 0.746 b | 0.689 | |||
| TD | NA | 20 (55.6) | 24 (47.1) | ||
| PIGD | NA | 11 (30.6) | 20 (39.2) | ||
| Indeterminate | NA | 5 (13.9) | 7 (13.7) | ||
| Disease duration (months) | NA | 54.00 (26.75, 72.00) | 66.00 (48.00, 82.00) | −1.452 e | 0.146 |
| PDSS (scores) | NA | 129.00 (102.00, 139.00) | 79.00 (72.00, 81.00) | −7.110 e | <0.001 |
| H-Y (on-stage) | NA | 2.00 (1.00, 2.50) | 2.50 (2.00, 3.00) | −2.716 e | <0.001 |
| Drug | PD-NSD (n = 36) | PD-SD (n = 51) | Z/χ2 | p |
|---|---|---|---|---|
| LED (mg) | 412.50 (337.50, 596.88) | 425.00 (337.50, 575.00) | −0.108 | 0.914 |
| Levodopa/benserazide (%) | 32 (88.9) | 48 (94.1) | 0.233 | 0.629 |
| Dopamine agonist (%) | 24 (66.7) | 38 (74.5) | 0.634 | 0.426 |
| MAO-B inhibitors (%) | 12 (33.3) | 20 (39.2) | 0.314 | 0.575 |
| Amantadine hydrochloride (%) | 4 (11.1) | 12 (23.5) | 2.168 | 0.141 |
| Test Indexes | HC (n = 49) | PD-NSD (n = 36) | PD-SD (n = 51) | F | p |
|---|---|---|---|---|---|
| ADO (ng/mL) | 19.07 ± 4.16 | 16.96 ± 3.01 * | 13.18 ± 3.31 *,# | 34.792 | <0.001 |
| GDNF (pg/mL) | 453.56 ± 141.26 | 392.81 ± 85.08 * | 287.55 ± 85.92 *,# | 29.619 | <0.001 |
| 5-HT (ng/mL) | 167.61 ± 14.88 | 164.08 ± 13.33 | 160.9 ± 10.52 * | 3.351 | 0.038 |
| γ-GABA (ng/mL) | 43.68 ± 2.84 | 39.22 ± 2.79 * | 37.21 ± 2.94 *,# | 66.045 | <0.001 |
| Ach (ng/mL) | 72.36 ± 8.53 | 69.64 ± 5.99 | 67.78 ± 7.50 * | 4.657 | 0.011 |
| Indicators | PSQI Total Score | Component 1 | Component 2 | Component 3 | Component 4 | Component 5 | Component 6 | Component 7 | |
|---|---|---|---|---|---|---|---|---|---|
| GDNF | r | −0.485 ** | −0.375 ** | −0.340 ** | −0.381 ** | −0.428 ** | −0.164 | −0.315 ** | −0.175 |
| p | <0.001 | <0.001 | 0.001 | <0.001 | <0.001 | 0.128 | 0.003 | 0.105 | |
| 5-HT | r | −0.029 | 0.110 | 0.074 | −0.154 | −0.078 | −0.053 | 0.114 | −0.039 |
| p | 0.793 | 0.308 | 0.497 | 0.154 | 0.470 | 0.625 | 0.292 | 0.720 | |
| ADO | r | −0.657 ** | −0.479 ** | −0.492 ** | −0.494 ** | −0.438 ** | −0.108 | −0.502 ** | −0.484 ** |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.318 | <0.001 | <0.001 | |
| γ-GABA | r | −0.526 ** | −0.510 ** | −0.351 ** | −0.325 ** | −0.277 ** | −0.131 | −0.468 ** | −0.583 ** |
| p | <0.001 | <0.001 | 0.001 | 0.002 | 0.009 | 0.226 | <0.001 | <0.001 | |
| Ach | r | −0.191 | −0.280 ** | −0.138 | −0.053 | −0.050 | −0.209 | −0.123 | −0.149 |
| p | 0.076 | 0.009 | 0.204 | 0.628 | 0.643 | 0.052 | 0.256 | 0.169 | |
| Indicators | PSQI Total Score | Component 1 | Component 2 | Component 3 | Component 4 | Component 5 | Component 6 | Component 7 | |
|---|---|---|---|---|---|---|---|---|---|
| MoCA | r | −0.164 | −0.216 * | 0.050 | −0.095 | −0.127 | −0.242 * | −0.045 | −0.229 * |
| p | 0.130 | 0.044 | 0.644 | 0.380 | 0.240 | 0.024 | 0.679 | 0.033 | |
| HAMA | r | 0.536 ** | 0.418 ** | 0.264 * | 0.460 ** | 0.356 ** | 0.168 | 0.336 ** | 0.468 ** |
| p | <0.001 | <0.001 | 0.014 | <0.001 | 0.001 | 0.121 | 0.001 | <0.001 | |
| HAMD | r | 0.469 ** | 0.396 ** | 0.289 ** | 0.370** | 0.335 ** | 0.193 | 0.269 * | 0.255 * |
| p | <0.001 | <0.001 | 0.007 | <0.001 | 0.002 | 0.073 | 0.012 | 0.017 | |
| UPDRS-III | r | 0.400 ** | 0.334 ** | 0.432 ** | 0.246 * | 0.270 * | 0.027 | 0.280 ** | 0.231 * |
| p | <0.001 | 0.002 | <0.001 | 0.022 | 0.011 | 0.802 | 0.009 | 0.031 | |
| H-Y stage | r | 0.434 ** | 0.316 ** | 0.115 | 0.261 * | 0.354 ** | 0.326 ** | 0.248 * | 0.452 ** |
| p | <0.001 | 0.003 | 0.288 | 0.015 | 0.001 | 0.002 | 0.020 | <0.001 | |
| The course of disease | r | 0.064 | −0.003 | 0.014 | 0.083 | 0.108 | −0.137 | 0.084 | 0.014 |
| p | 0.557 | 0.976 | 0.899 | 0.442 | 0.321 | 0.207 | 0.439 | 0.894 | |
| LED | r | 0.205 | 0.273 * | 0.059 | 0.108 | 0.044 | 0.101 | 0.202 | 0.322 ** |
| p | 0.056 | 0.011 | 0.590 | 0.319 | 0.684 | 0.351 | 0.061 | 0.002 | |
| RBD-SQ | r | 0.178 | 0.070 | −0.100 | 0.060 | 0.208 | 0.224 * | 0.099 | 0.339 ** |
| p | 0.100 | 0.521 | 0.359 | 0.581 | 0.053 | 0.037 | 0.361 | 0.001 | |
| Factors | B | SE | Waldχ2 | p | OR (95% CI) |
|---|---|---|---|---|---|
| GDNF | −0.017 | 0.005 | 12.404 | <0.001 | 0.983 (0.974~0.993) |
| ADO | −0.577 | 0.211 | 7.464 | 0.006 | 0.562 (0.371~0.85) |
| γ-GABA | 0.441 | 0.246 | 3.202 | 0.074 | 1.554 (0.959~2.519) |
| HAMD | 0.128 | 0.059 | 4.709 | 0.030 | 1.137 (1.013~1.277) |
| HAMA | 0.001 | 0.072 | 0.001 | 0.984 | 1.001 (0.87~1.153) |
| UPDRS-III | 0.001 | 0.025 | 0.002 | 0.963 | 1.001 (0.953~1.052) |
| H-Y stage | −0.096 | 0.393 | 0.059 | 0.807 | 0.909 (0.421~1.962) |
| Constant | −3.211 | 7.317 | 0.193 | 0.661 | - |
| Parameter | GDNF | ADO | ADO + GDNF |
|---|---|---|---|
| AUC 95% CI | 0.815 0.723~0.906 | 0.780 0.685~0.875 | 0.883 0.808~0.959 |
| SE | 0.047 | 0.048 | 0.039 |
| p | <0.001 | <0.001 | <0.001 |
| The threshold value | 305.205 | 13.000 | 0.570 |
| Sensitivity (%) | 62.7 | 49.0 | 82.4 |
| Specificity (%) | 91.7 | 94.4 | 83.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Gao, Z.; Chen, G.; Geng, D.; Gao, D. Low Levels of Adenosine and GDNF Are Potential Risk Factors for Parkinson’s Disease with Sleep Disorders. Brain Sci. 2023, 13, 200. https://doi.org/10.3390/brainsci13020200
Wang L, Gao Z, Chen G, Geng D, Gao D. Low Levels of Adenosine and GDNF Are Potential Risk Factors for Parkinson’s Disease with Sleep Disorders. Brain Sciences. 2023; 13(2):200. https://doi.org/10.3390/brainsci13020200
Chicago/Turabian StyleWang, Li, Zheng Gao, Gang Chen, Deqin Geng, and Dianshuai Gao. 2023. "Low Levels of Adenosine and GDNF Are Potential Risk Factors for Parkinson’s Disease with Sleep Disorders" Brain Sciences 13, no. 2: 200. https://doi.org/10.3390/brainsci13020200