
Discriminatory Brain Processes of Native and Foreign Language in Children with and without Reading Difficulties
 ,
,
Abstract
:1. Introduction
1.1. Language Development and Speech Processing in Typical Readers and in Dyslexics
1.2. Native and Foreign Language Acquisition in Dyslexia
1.3. Phonological Deficit and Dyslexia in ERP Research
1.4. The Discriminatory Brain Responses
1.5. Hypotheses and Objectives
2. Materials and Methods
2.1. Participants
2.2. Selection Criteria and Tests
2.2.1. Reading Score
2.2.2. Cognitive Nonverbal Assessment
2.2.3. The Attention and Executive Function Questionnaire
2.3. Stimuli and Procedure
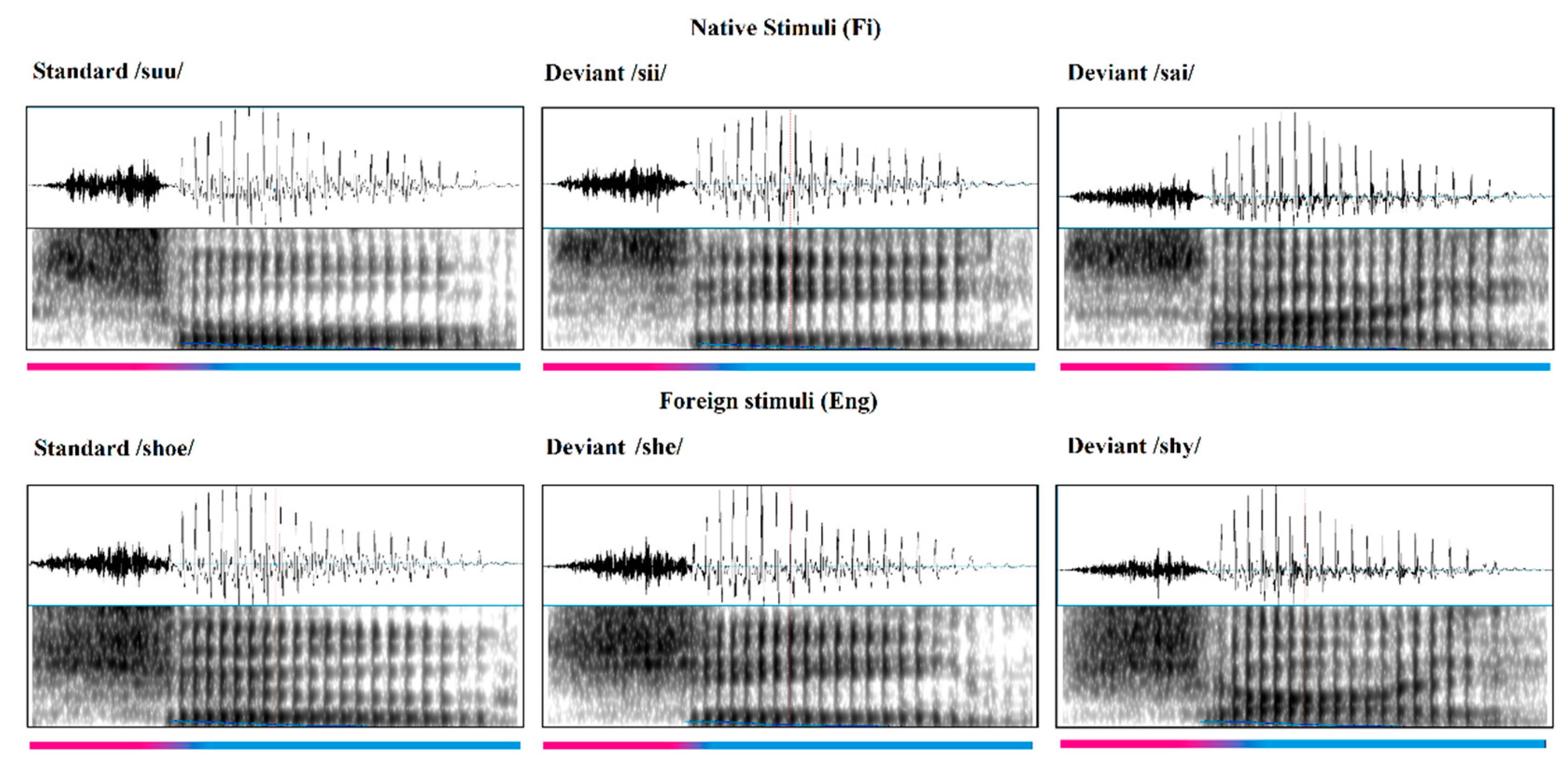
2.3.1. Stimuli
2.3.2. Euclidean Distance and Center of Gravity
2.4. EEG Recording and Pre-Processing
2.5. Statistical Analyses
3. Results
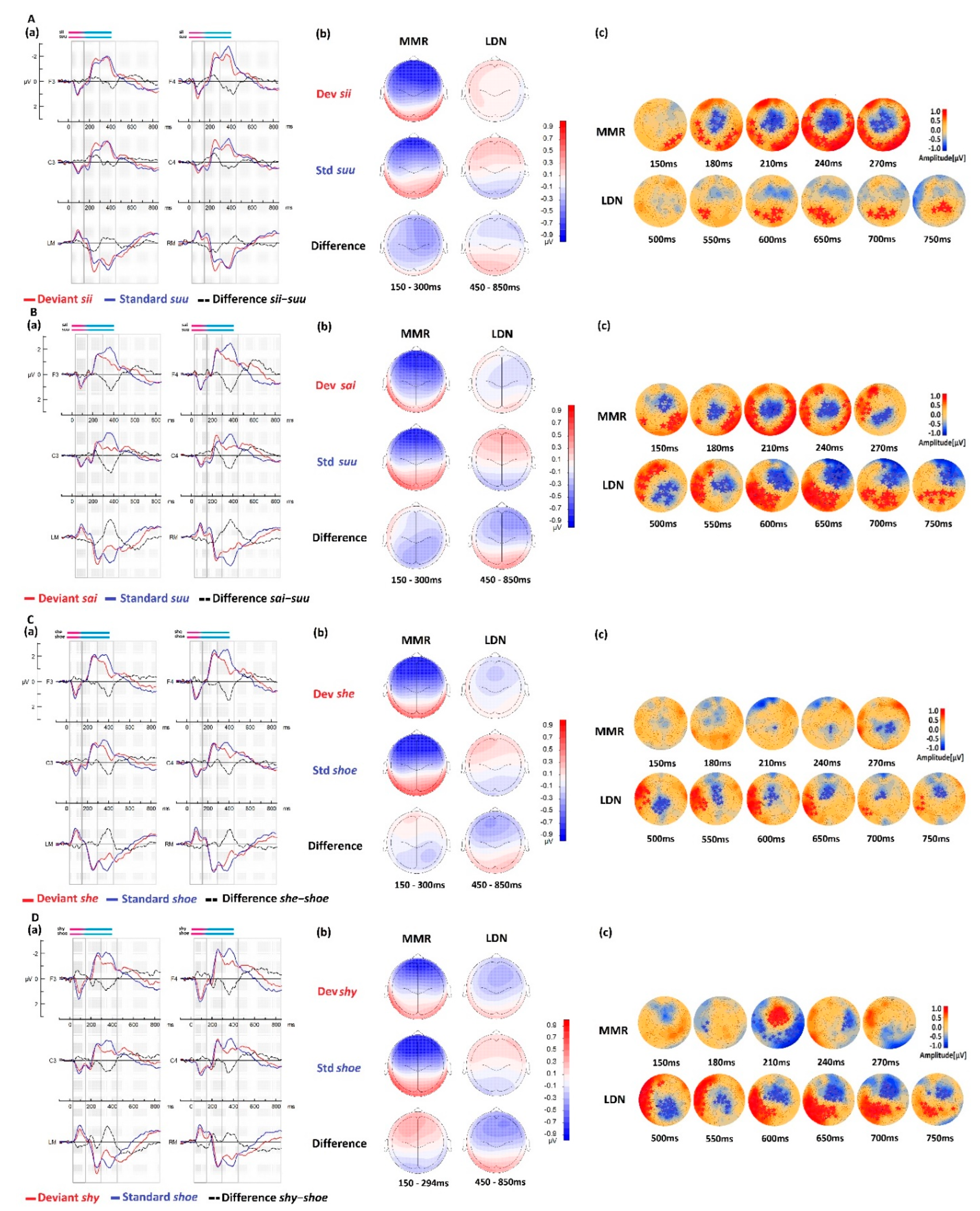
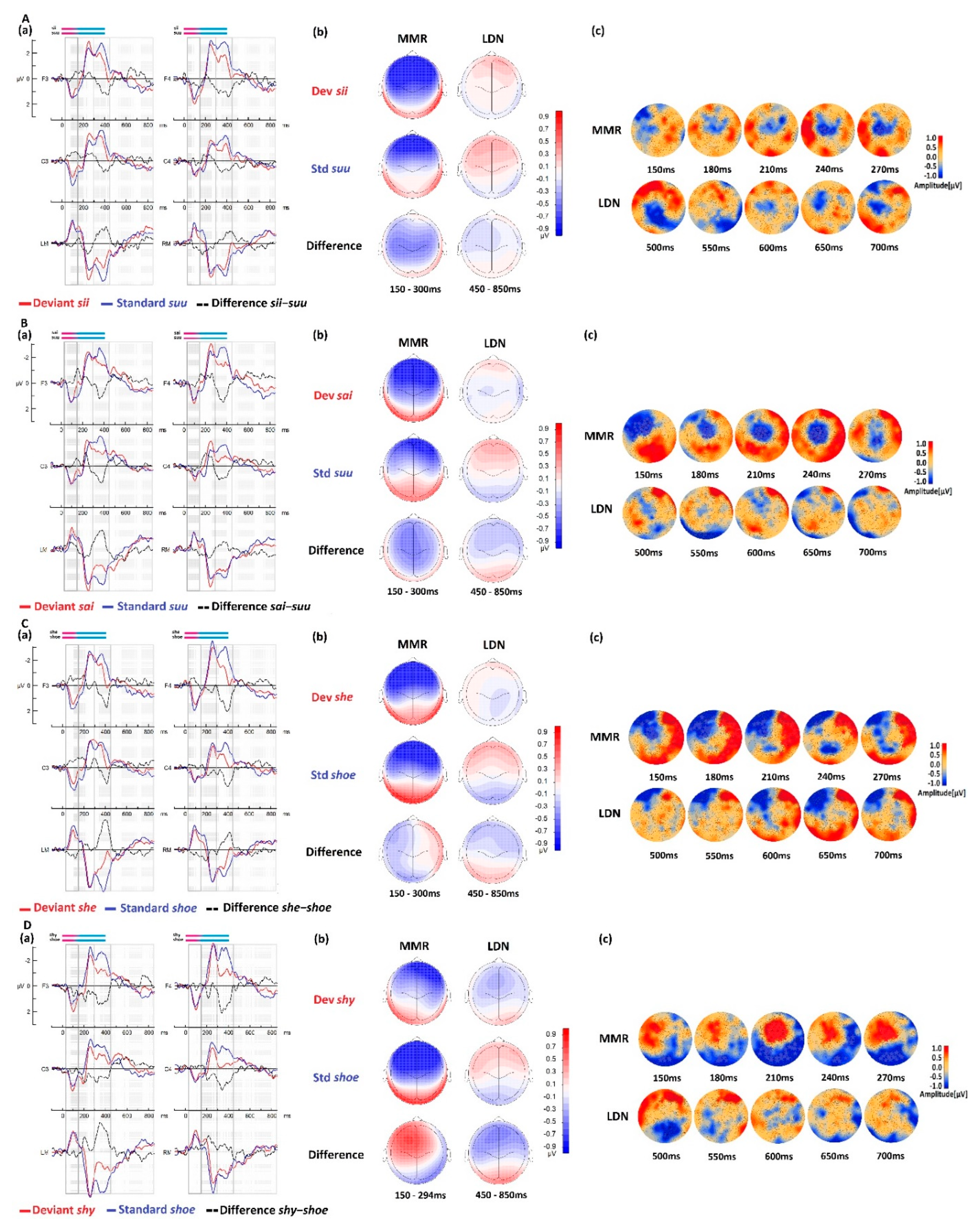
3.1. Native vs. Foreign Language Processing
3.1.1. Comparisons between Native and Foreign Language Processing in the CTR Group
3.1.2. Comparisons between Native and Foreign Language Processing in the RD Group
3.2. Group Comparison
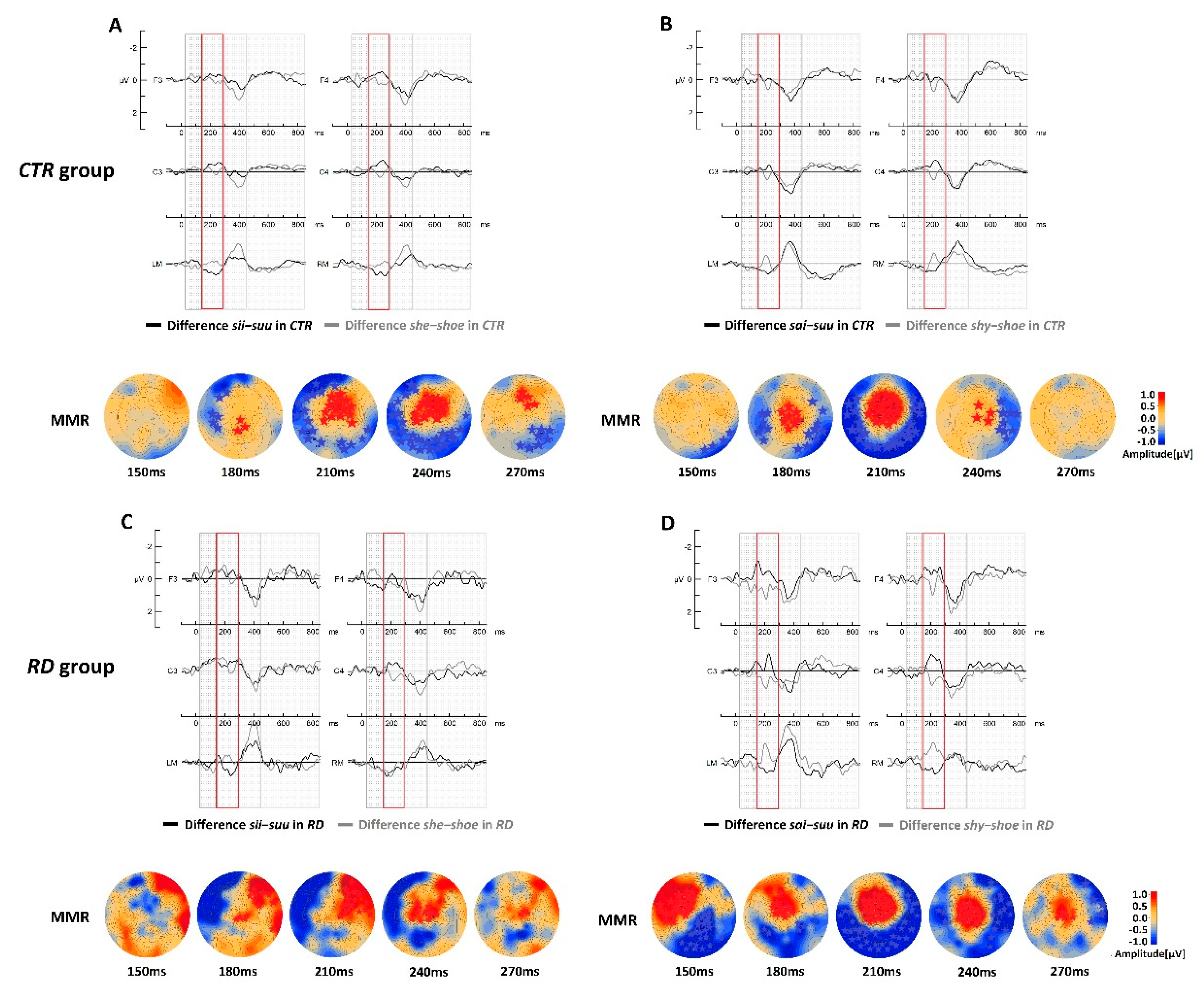
3.3. Within-Group Analyses
3.3.1. Within-CTR Group Comparisons for Native Stimuli
3.3.2. Within-CTR Group Comparisons for Foreign Stimuli
3.3.3. Within-RD Group Comparisons for Native Stimuli
3.3.4. Within-RD Group Comparisons for Foreign Stimuli
3.4. ANOVA Results
4. Discussion
4.1. Native vs. Foreign Language Processing
4.2. Group Comparison CTR vs. RD
4.3. Within Group Results
4.3.1. Native and Foreign Language Processes in Typical Readers
The Mismatch Response within the CTR Group
The Late Discriminative Response within the CTR Group
4.3.2. Native and Foreign Language Processes in Poor Readers
The Mismatch Response within the RD Group
The Late Discriminative Response within the RD Group
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Démonet, J.F.; Taylor, M.J.; Chaix, Y. Developmental dyslexia. Lancet 2004, 363, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Noordenbos, M.W.; Segers, E.; Serniclaes, W.; Mitterer, H.; Verhoeven, L. Neural evidence of allophonic perception in children at risk for dyslexia. Neuropsychologia 2012, 50, 2010–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramus, F. Developmental dyslexia: Specific phonological deficit or general sensorimotor dysfunction? Curr. Opin. Neurobiol. 2003, 13, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vellutino, F.R.; Fletcher, J.M.; Snowling, M.J.; Scanlon, D.M. Specific reading disability (dyslexia): What have we learned in the past four decades? J. Child Psychol. Psychiatry 2004, 45, 2–40. [Google Scholar] [CrossRef] [PubMed]
- Ylinen, S.; Junttila, K.; Laasonen, M.; Iverson, P.; Ahonen, L.; Kujala, T. Diminished brain responses to second-language words are linked with native-language literacy skills in dyslexia. Neuropsychologia 2019, 122, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.L.; Pennington, B.F. Developmental dyslexia. Annu. Rev. Clin. Psychol. 2015, 11, 283–307. [Google Scholar] [CrossRef]
- Hornickel, J.; Kraus, N. Unstable representation of sound: A biological marker of dyslexia. J. Neurosci. 2013, 33, 3500–3504. [Google Scholar] [CrossRef] [Green Version]
- Snowling, M.J. From language to reading and dyslexia. Dyslexia 2001, 7, 37–46. [Google Scholar] [CrossRef]
- Boets, B.; de Beeck, H.P.O.; Vandermosten, M.; Scott, S.K.; Gillebert, C.R.; Mantini, D.; Bulthé, J.; Sunaert, S.; Wouters, J.; Ghesquière, P. Intact but less accessible phonetic representations in adults with dyslexia. Science 2013, 342, 1251–1254. [Google Scholar] [CrossRef] [Green Version]
- Hoeft, F.; McCandliss, B.D.; Black, J.M.; Gantman, A.; Zakerani, N.; Hulme, C.; Lyytinen, H.; Whitfield-Gabrieli, S.; Glover, G.H.; Reiss, A.L.; et al. Neural systems predicting long-term outcome in dyslexia. Proc. Natl. Acad. Sci. USA 2011, 108, 361–366. [Google Scholar] [CrossRef]
- Ramus, F.; Szenkovits, G. What phonological deficit? Q. J. Exp. Psychol. 2008, 61, 129–141. [Google Scholar] [CrossRef]
- Boada, R.; Pennington, B.F. Deficient implicit phonological representations in children with dyslexia. J. Exp. Child Psychol. 2006, 95, 153–193. [Google Scholar] [CrossRef]
- Litt, R.A.; Nation, K. The nature and specificity of paired associate learning deficits in children with dyslexia. J. Mem. Lang. 2014, 71, 71–88. [Google Scholar] [CrossRef]
- Soroli, E.; Szenkovits, G.; Ramus, F. Exploring dyslexics’ phonological deficit III: Foreign speech perception and production. Dyslexia 2010, 16, 318–340. [Google Scholar] [CrossRef] [PubMed]
- Łockiewicz, M.; Sarzała-Przybylska, Z.; Lipowska, M. Early predictors of learning a foreign language in pre-school–Polish as a first language, English as a foreign language. Front. Psychol. 2018, 9, 1813. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Körne, G.; Bruder, J. Clinical neurophysiology of visual and auditory processing in dyslexia: A review. Clin. Neurophysiol. 2010, 121, 1794–1809. [Google Scholar] [CrossRef]
- Best, C.C.; McRoberts, G.W. Infant perception of non-native consonants contrasts that adults assimilate in different ways. Lang. Speech 2003, 46, 183–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obleser, J.; Elbert, T.; Lahiri, A.; Eulitz, C. Cortical representation of vowels reflects acoustic dissimilarity determined by formant frequencies. Cogn. Brain Res. 2003, 15, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Werker, J.F.; Tees, R.C. Cross-language speech perception: Evidence for perceptual reorganization during the first year of life. Infant Behav. Dev. 1984, 7, 49–63. [Google Scholar] [CrossRef]
- Winkler, I.; Kujala, T.; Tiitinen, H.; Sivonen, P.; Alku, P.; Lehtokoski, A.; Czigler, I.; Csépe, V.; Ilmoniemi, R.J.; Näätänen, R. Brain responses reveal the learning of foreign language phonemes. Psychophysiology 1999, 36, 638–642. [Google Scholar] [CrossRef]
- Serniclaes, W. Allophonic theory of dyslexia: A short overview. JSM Commun. Disord. 2018. Available online: https://hal.archives-ouvertes.fr/hal-02065653/ (accessed on 29 December 2022).
- Serniclaes, W.; Seck, M.B. Enhanced sensitivity to subphonemic segments in dyslexia: A new instance of allophonic perception. Brain Sci. 2018, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttorm, T.K.; Leppänen, P.H.; Hämäläinen, J.A.; Eklund, K.M.; Lyytinen, H.J. Newborn event-related potentials predict poorer pre-reading skills in children at risk for dyslexia. J. Learn. Disabil. 2010, 43, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Leppänen, P.H.T.; Hämäläinen, J.A.; Salminen, H.K.; Eklund, K.M.; Guttorm, T.K.; Lohvansuu, K.; Puolakanaho, A.; Lyytinen, H. Newborn brain event-related potentials revealing atypical processing of sound frequency and the subsequent association with later literacy skills in children with familial dyslexia. Cortex 2010, 46, 1362–1376. [Google Scholar] [CrossRef] [PubMed]
- Lohvansuu, K.; Hämäläinen, J.A.; Ervast, L.; Lyytinen, H.; Leppänen, P.H. Longitudinal interactions between brain and cognitive measures on reading development from 6 months to 14 years. Neuropsychologia 2018, 108, 6–12. [Google Scholar] [CrossRef]
- Molfese, D.L. Predicting dyslexia at 8 years of age using neonatal brain responses. Brain Lang. 2000, 72, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Schaadt, G.; Männel, C.; van der Meer, E.; Pannekamp, A.; Oberecker, R.; Friederici, A.D. Present and past: Can writing abilities in school children be associated with their auditory discrimination capacities in infancy? Res. Dev. Disabil. 2015, 47, 318–333. [Google Scholar] [CrossRef]
- Leppänen, P.H.T.; Richardson, U.; Pihko, E.; Eklund, K.M.; Guttorm, T.K.; Aro, M.; Lyytinen, H. Brain responses to changes in speech sound durations differ between infants with and without familial risk for dyslexia. Dev. Neuropsychol. 2002, 22, 407–422. [Google Scholar] [CrossRef]
- Leppänen, P.H.T.; Hämäläinen, J.A.; Guttorm, T.K.; Eklund, K.M.; Salminen, H.; Tanskanen, A.; Torppa, M.; Puolakanaho, A.; Richardson, U.; Pennala, R.; et al. Infant brain responses associated with reading-related skills before school and at school age. Neurophysiol. Clin./Clin. Neurophysiol. 2012, 42, 35–41. [Google Scholar] [CrossRef]
- Lyytinen, H.; Erskine, J.; Hämäläinen, J.; Torppa, M.; Ronimus, M. Dyslexia—Early identification and prevention: Highlights from the Jyväskylä longitudinal study of dyslexia. Curr. Dev. Disord. Rep. 2015, 2, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Van der Leij, A.; Van Bergen, E.; van Zuijen, T.; De Jong, P.; Maurits, N.; Maassen, B. Precursors of developmental dyslexia: An overview of the longitudinal Dutch dyslexia programme study. Dyslexia 2013, 19, 191–213. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Been, P.; Kuijpers, C.; Zwarts, F.; Maassen, B.; van der Leij, A. Mismatch response is absent in 2-month-old infants at risk for dyslexia. NeuroReport 2006, 17, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Van Zuijen, T.L.; Plakas, A.; Maassen, B.A.; Been, P.; Maurits, N.M.; Krikhaar, E.; van Driel, J.; van der Leij, A. Temporal auditory processing at 17 months of age is associated with preliterate language comprehension and later word reading fluency: An ERP study. Neurosci. Lett. 2012, 528, 31–35. [Google Scholar] [CrossRef]
- Kim, M.; Crossley, S.A.; Kim, B.K. Second language reading and writing in relation to first language, vocabulary knowledge, and learning backgrounds. Int. J. Biling. Educ. Biling. 2022, 25, 1992–2005. [Google Scholar] [CrossRef]
- Botezatu, M.R.; Guo, T.; Kroll, J.F.; Peterson, S.; Garcia, D.L. Sources of variation in second and native language speaking proficiency among college-aged second language learners. Stud. Second. Lang. Acquis. 2022, 44, 305–330. [Google Scholar] [CrossRef]
- Cisero, C.A.; Royer, J.M. The development and cross-language transfer of phonological awareness. Contemp. Educ. Psychol. 1995, 20, 275–303. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G.; Dupoux, E.; Gout, A. Electrophysiological correlates of phonological processing: A cross-linguistic study. J. Cogn. Neurosci. 2000, 12, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Durgunoğlu, A.Y.; Nagy, W.E.; Hancin-Bhatt, B.J. Cross-language transfer of phonological awareness. J. Educ. Psychol. 1993, 85, 453. [Google Scholar] [CrossRef]
- Macizo, P. Phonological coactivation in the bilinguals’ two languages: Evidence from the color naming task. Bilingualism 2016, 19, 361. [Google Scholar] [CrossRef]
- Goswami, U. Phonological representations, reading development and dyslexia: Towards a cross-linguistic theoretical framework. Dyslexia 2000, 6, 133–151. [Google Scholar] [CrossRef]
- Helland, T.; Kaasa, R. Dyslexia in English as a second language. Dyslexia 2005, 11, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Miller-Guron, L.; Lundberg, I. Dyslexia and second language reading: A second bite at the apple? Read. Writ. 2000, 12, 41–61. [Google Scholar] [CrossRef]
- Hermans, D.; Bongaerts, T.; De Bot, K.; Schreuder, R. Producing words in a foreign language: Can speakers prevent interference from their first language? Biling. Lang. Cogn. 1998, 1, 213–229. [Google Scholar] [CrossRef]
- Nation, P. The role of the first language in foreign language learning. Asian EFL J. 2003, 5, 1–8. [Google Scholar]
- Dehaene, S. Fitting two languages into one brain. Brain 1999, 122, 2207–2208. [Google Scholar] [CrossRef]
- Friederici, A.D. Language in Our Brain: The Origins of a Uniquely Human Capacity; MIT Press: Cambridge, MA, USA, 2017. [Google Scholar] [CrossRef]
- Kim, K.H.; Relkin, N.R.; Lee, K.M.; Hirsch, J. Distinct cortical areas associated with native and second languages. Nature 1997, 388, 171–174. [Google Scholar] [CrossRef]
- Perani, D.; Dehaene, S.; Grassi, F.; Cohen, L.; Cappa, S.F.; Dupoux, E.; Fazio, F.; Mehler, J. Brain processing of native and foreign languages. NeuroReport 1996, 7, 2439–2444. [Google Scholar] [CrossRef]
- Nora, A.; Karvonen, L.; Renvall, H.; Parviainen, T.; Kim, J.Y.; Service, E.; Salmelin, R. Children show right-lateralized effects of spoken word-form learning. PLoS ONE 2017, 12, e0171034. [Google Scholar] [CrossRef] [Green Version]
- Deacon, S.H.; Kirby, J.R. Morphological awareness: Just “more phonological”? The roles of morphological and phonological awareness in reading development. Appl. Psycholinguist. 2004, 25, 223. [Google Scholar] [CrossRef]
- Hogan, T.P.; Catts, H.W.; Little, T.D. The relationship between phonological awareness and reading. Lang. Speech Hear. Serv. Sch. 2005, 36, 285–293. [Google Scholar] [CrossRef]
- Schulte-Körne, G.; Deimel, W.; Bartling, J.; Remschmidt, H. The role of phonological awareness, speech perception, and auditory temporal processing for dyslexia. Eur. Child Adolesc. Psychiatry 1999, 8, S28–S34. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.C.; Goswami, U. Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychol. Bull. 2005, 131, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomert, L.; Mitterer, H. The fragile nature of the speech-perception deficit in dyslexia: Natural vs. synthetic speech. Brain Lang. 2004, 89, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Elbro, C.; Jensen, M.N. Quality of phonological representations, verbal learning, and phoneme awareness in dyslexic and normal readers. Scand. J. Psychol. 2005, 46, 375–384. [Google Scholar] [CrossRef]
- Kita, Y.; Yamamoto, H.; Oba, K.; Terasawa, Y.; Moriguchi, Y.; Uchiyama, H.; Seki, A.; Koeda, T.; Inagaki, M. Altered brain activity for phonological manipulation in dyslexic Japanese children. Brain 2013, 136, 3696–3708. [Google Scholar] [CrossRef] [PubMed]
- McBride-Chang, C. Models of speech perception and phonological processing in reading. Child Dev. 1996, 67, 1836–1856. [Google Scholar] [CrossRef]
- Ramus, F.; Ahissar, M. Developmental dyslexia: The difficulties of interpreting poor performance, and the importance of normal performance. Cogn. Neuropsychol. 2012, 29, 104–122. [Google Scholar] [CrossRef] [PubMed]
- Adlard, A.; Hazan, V. Speech perception in children with specific reading difficulties (dyslexia). Q. J. Exp. Psychol. Sect. A 1998, 51, 153–177. [Google Scholar] [CrossRef]
- Boets, B.; Vandermosten, M.; Poelmans, H.; Luts, H.; Wouters, J.; Ghesquiere, P. Preschool impairments in auditory processing and speech perception uniquely predict future reading problems. Res. Dev. Disabil. 2011, 32, 560–570. [Google Scholar] [CrossRef]
- Hickok, G.; Small, S.L. (Eds.) Neurobiology of Language; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Näätänen, R. Mismatch negativity (MMN): Perspectives for application. Int. J. Psychophysiol. 2000, 37, 3–10. [Google Scholar] [CrossRef]
- Näätänen, R.; Sussman, E.S.; Salisbury, D.; Shafer, V.L. Mismatch negativity (MMN) as an index of cognitive dysfunction. Brain Topogr. 2014, 27, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kujala, T.; Näätänen, R. The mismatch negativity in evaluating central auditory dysfunction in dyslexia. Neurosci. Biobehav. Rev. 2001, 25, 535–543. [Google Scholar] [CrossRef]
- Leppänen, P.H.T.; Tóth, D.; Honbolygó, F.; Lohvansuu, K.; Hämäläinen, J.A.; Demonet, J.F.; Schulte-Körne, G.; Csépe, V. Reproducibility of brain responses: High for speech perception, low for reading difficulties. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pakarinen, S.; Teinonen, T.; Shestakova, A.; Kwon, M.S.; Kujala, T.; Hämäläinen, H.; Näätänen, R.; Huotilainen, M. Fast parametric evaluation of central speech-sound processing with mismatch negativity (MMN). Int. J. Psychophysiol. 2013, 87, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Partanen, E.; Torppa, R.; Pykäläinen, J.; Kujala, T.; Huotilainen, M. Children’s brain responses to sound changes in pseudo words in a multifeature paradigm. Clin. Neurophysiol. 2013, 124, 1132–1138. [Google Scholar] [CrossRef]
- Čeponienė, R.; Torki, M.; Alku, P.; Koyama, A.; Townsend, J. Event-related potentials reflect spectral differences in speech and non-speech stimuli in children and adults. Clin. Neurophysiol. 2008, 119, 1560–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaralı, M.; Yağcıoğlu, S. Effect of consonant duration on formation of consonant-vowel syllable evoked auditory cortical potentials. J. Int. Adv. Otol. 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, C.; Roux, S.; Bonnet-Brilhault, F.; Ferré, S.; Gomot, M. Brain responses to change in phonological structures of varying complexity in children and adults. Psychophysiology 2020, 57, e13621. [Google Scholar] [CrossRef]
- Virtala, P.; Putkinen, V.; Kailaheimo-Lönnqvist, L.; Thiede, A.; Partanen, E.; Kujala, T. Infancy and early childhood maturation of neural auditory change detection and its associations to familial dyslexia risk. Clin. Neurophysiol. 2022, 137, 159–176. [Google Scholar] [CrossRef]
- Näätänen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef]
- Bishop, D.V.M. Using mismatch negativity to study central auditory processing in developmental language and literacy impairments: Where are we, and where should we be going? Psychol. Bull. 2007, 133, 651. [Google Scholar] [CrossRef] [PubMed]
- Volkmer, S.; Schulte-Koerne, G. Cortical responses to tone and phoneme mismatch as a predictor of dyslexia? A systematic review. Schizophr. Res. 2018, 191, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.I.; Kilner, J.M.; Stephan, K.E.; Friston, K.J. The mismatch negativity: A review of underlying mechanisms. Clin. Neurophysiol. 2009, 120, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Winkler, I. Interpreting the mismatch negativity. J. Psychophysiol. 2007, 21, 147–163. [Google Scholar] [CrossRef]
- Lohvansuu, K.; Hämäläinen, J.A.; Tanskanen, A.; Bartling, J.; Bruder, J.; Honbolygó, F.; Schulte-Körne, G.; Démonet, J.F.; Csépe, V.; Leppänen, P.H.T. Separating mismatch negativity (MMN) response from auditory obligatory brain responses in school-aged children. Psychophysiology 2013, 50, 640–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hämäläinen, J.A.; Lohvansuu, K.; Ervast, L.; Leppänen, P.H. Event-related potentials to tones show differences between children with multiple risk factors for dyslexia and control children before the onset of formal reading instruction. Int. J. Psychophysiol. 2015, 95, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Hämäläinen, J.; Landi, N.; Loberg, O.; Lohvansuu, K.; Pugh, K.; Leppänen, P.H. Brain event-related potentials to phoneme contrasts and their correlation to reading skills in school-age children. Int. J. Behav. Dev. 2018, 42, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Lohvansuu, K.; Hämäläinen, J.A.; Tanskanen, A.; Ervast, L.; Heikkinen, E.; Lyytinen, H.; Leppänen, P.H. Enhancement of brain event-related potentials to speech sounds is associated with compensated reading skills in dyslexic children with familial risk for dyslexia. Int. J. Psychophysiol. 2014, 94, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.C.; Murphy, C.F.B.; Schochat, E. Auditory processing in children with dyslexia: Electrophysiological and behavior evaluation. CoDAS 2013, 25, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Zevin, J.D.; Datta, H.; Maurer, U.; Rosania, K.A.; McCandliss, B.D. Native language experience influences the topography of the mismatch negativity to speech. Front. Hum. Neurosci. 2010, 4, 212. [Google Scholar] [CrossRef] [Green Version]
- Cheour, M.; Korpilahti, P.; Martynova, O.; Lang, A.H. Mismatch negativity and late discriminative negativity in investigating speech perception and learning in children and infants. Audiol. Neurotol. 2001, 6, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Korpilahti, P.; Krause, C.M.; Holopainen, I.; Lang, A.H. Early and late mismatch negativity elicited by words and speech-like stimuli in children. Brain Lang. 2001, 76, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Maurer, J.; Collet, L.; Pelster, H.; Truy, E.; Gallégo, S. Auditory late cortical response and speech recognition in digisonic cochlear implant users. Laryngoscope 2002, 112, 2220–2224. [Google Scholar] [CrossRef] [PubMed]
- Giraud, A.L.; Ramus, F. Neurogenetics and auditory processing in developmental dyslexia. Curr. Opin. Neurobiol. 2013, 23, 37–42. [Google Scholar] [CrossRef]
- Neuhoff, N.; Bruder, J.; Bartling, J.; Warnke, A.; Remschmidt, H.; Müller-Myhsok, B.; Schulte-Körne, G. Evidence for the late MMN as a neurophysiological endophenotype for dyslexia. PLoS ONE 2012, 7, e34909. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Körne, G.; Deimel, W.; Bartling, J.; Remschmidt, H. Speech perception deficit in dyslexic adults as measured by mismatch negativity (MMN). Int. J. Psychophysiol. 2001, 40, 77–87. [Google Scholar] [CrossRef]
- Čeponienė, R.; Lepistö, T.; Soininen, M.; Aronen, E.; Alku, P.; Näätänen, R. Event-related potentials associated with sound discrimination versus novelty detection in children. Psychophysiology 2004, 41, 130–141. [Google Scholar] [CrossRef]
- Halliday, L.F.; Barry, J.G.; Hardiman, M.J.; Bishop, D.V. Late, not early mismatch responses to changes in frequency are reduced or deviant in children with dyslexia: An event-related potential study. J. Neurodev. Disord. 2014, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Búa, B.; Díaz, F.; Ferraces, M.J. The contribution of AERPs (MMN and LDN) to studying temporal vs. linguistic processing deficits in children with reading difficulties. Int. J. Psychophysiol. 2006, 59, 159–167. [Google Scholar] [CrossRef]
- Hommet, C.; Vidal, J.; Roux, S.; Blanc, R.; Barthez, M.A.; De Becque, B.; Barthelemy, C.; Bruneau, N.; Gomot, M. Topography of syllable change-detection electrophysiological indices in children and adults with reading disabilities. Neuropsychologia 2009, 47, 761–770. [Google Scholar] [CrossRef]
- Jakoby, H.; Goldstein, A.; Faust, M. Electrophysiological correlates of speech perception mechanisms and individual differences in second language attainment. Psychophysiology 2011, 48, 1517–1531. [Google Scholar] [CrossRef] [PubMed]
- Shestakova, A.; Huotilainen, M.; Čeponien, R.; Cheour, M. Event-related potentials associated with second language learning in children. Clin. Neurophysiol. 2003, 114, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Körne, G.; Deimel, W.; Bartling, J.; Remschmidt, H. Auditory processing and dyslexia: Evidence for a specific speech processing deficit. NeuroReport 1998, 9, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Kiili, C.; Leu, D.J.; Utriainen, J.; Coiro, J.; Kanniainen, L.; Tolvanen, A.; Lohvansuu, K.; Leppänen, P.H. Reading to learn from online information: Modeling the factor structure. J. Lit. Res. 2018, 50, 304–334. [Google Scholar] [CrossRef]
- Lindeman, J. ALLU—Ala-Asteen Lukutesti [ALLU—Reading Test for Primary School]; The Center for Learning Research, University of Turku: Turku, Finland, 1998. [Google Scholar]
- Nevala, J.; Lyytinen, H. Sanaketjutesti [Differentiate Word Test]; Niilo Mäki Instituutti Jyväskylän Yliopiston Lapsitutkimuskeskus: Jyväskylä, Finland, 2000. [Google Scholar]
- Eklund, K.; Torppa, M.; Aro, M.; Leppänen, P.H.; Lyytinen, H. Literacy skill development of children with familial risk for dyslexia through grades 2, 3, and 8. J. Educ. Psychol. 2015, 107, 126. [Google Scholar] [CrossRef]
- Kanniainen, L.; Kiili, C.; Tolvanen, A.; Aro, M.; Leppänen, P.H. Literacy skills and online research and comprehension: Struggling readers face difficulties online. Read. Writ. 2019, 32, 2201–2222. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.C.; Court, J.H. Raven’s Progressive Matrices and Vocabulary Scales (vol. 759); Oxford Pyschologists Press: Oxford, UK, 1998. [Google Scholar]
- Klenberg, L.; Jämsä, S.; Häyrinen, T.; Lahti-Nuuttila, P.E.K.K.A.; Korkman, M. The Attention and Executive Function Rating Inventory (ATTEX): Psychometric properties and clinical utility in diagnosing ADHD subtypes. Scand. J. Psychol. 2010, 51, 439–448. [Google Scholar] [CrossRef]
- Boersma, P.; Weenink, D. Praat: Doing Phonetics by Computer [Computer Program]. Version 6.2.04. 2021. Available online: http://www.praat.org/ (accessed on 18 December 2021).
- Lennes, M. Finnish sound structure. Phonetics, phonotactics and prosody. Puhe ja kieli 2010, 30, 121–124. [Google Scholar]
- Leech, G.; Rayson, P.; Wilson, A. Companion Website for: Word Frequencies in Written and Spoken English: Based on the British National Corpus. 2016. Available online: http://ucrel.lancs.ac.uk/bncfreq/ (accessed on 29 December 2022).
- Huovilainen, T.M. Psycholinguistic Descriptives. In The Language Bank of Finland. 2018. Available online: http://urn.fi/urn:nbn:fi:lb-2018081601 (accessed on 29 December 2022).
- Suomi, K.; Toivanen, J.; Ylitalo, R. Finnish sound structure. In Studia Humaniora Ouluensia; University of Oulu: Oulu, Finland, 2008. [Google Scholar]
- Bidelman, G.M.; Moreno, S.; Alain, C. Tracing the emergence of categorical speech perception in the human auditory system. Neuroimage 2013, 79, 201–212. [Google Scholar] [CrossRef]
- Hay, J.; Warren, P.; Drager, K. Factors influencing speech perception in the context of a merger-in-progress. J. Phon. 2006, 34, 458–484. [Google Scholar] [CrossRef]
- Jiang, J.; Auer, E.T.; Alwan, A.; Keating, P.A.; Bernstein, L.E. Similarity structure in visual speech perception and optical phonetic signals. Percept. Psychophys. 2007, 69, 1070–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakulenko, M.O. Unified Parametrization of Phonetic Features and Numerical Calculation of Phonetic Distances between Speech Sounds. J. Quant. Linguist. 2022, 1–19. [Google Scholar] [CrossRef]
- Wedel, A.; Nelson, N.; Sharp, R. The phonetic specificity of contrastive hyperarticulation in natural speech. J. Mem. Lang. 2018, 100, 61–88. [Google Scholar] [CrossRef]
- Vakulenko, M. Calculation of Phonetic Distances between Speech Sounds. J. Quant. Linguist. 2021, 28, 223–236. [Google Scholar] [CrossRef]
- Chodroff, E.; Wilson, C. Acoustic–phonetic and auditory mechanisms of adaptation in the perception of sibilant fricatives. Atten. Percept. Psychophys. 2020, 82, 2027–2048. [Google Scholar] [CrossRef] [PubMed]
- Yu, A. On the nature of the perception-production link: Individual variability in English sibilant-vowel coarticulation. Lab. Phonol. 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.J.; Sejnowski, T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef]
- Ferree, T.C. Spherical splines and average referencing in scalp electroencephalography. Brain Topogr. 2006, 19, 43–52. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Näätänen, R.; Pakarinen, S.; Rinne, T.; Takegata, R. The mismatch negativity (MMN): Towards the optimal paradigm. Clin. Neurophysiol. 2004, 115, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Abutalebi, J. Neural aspects of second language representation and language control. Acta Psychol. 2008, 128, 466–478. [Google Scholar] [CrossRef]
- Leppänen, P.H.T.; Lyytinen, H. Auditory event-related potentials in the study of developmental language-related disorders. Audiol. Neurotol. 1997, 2, 308–340. [Google Scholar] [CrossRef] [PubMed]
- Tzourio, N.; El Massioui, F.; Crivello, F.; Joliot, M.; Renault, B.; Mazoyer, B. Functional anatomy of human auditory attention studied with PET. Neuroimage 1997, 5, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Froyen, D.; Willems, G.; Blomert, L. Evidence for a specific cross-modal association deficit in dyslexia: An electrophysiological study of letter–speech sound processing. Dev. Sci. 2011, 14, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Peltola, M.S.; Kujala, T.; Tuomainen, J.; Ek, M.; Aaltonen, O.; Näätänen, R. Native and foreign vowel discrimination as indexed by the mismatch negativity (MMN) response. Neurosci. Lett. 2003, 352, 25–28. [Google Scholar] [CrossRef]
- Virtala, P.; Partanen, E.; Tervaniemi, M.; Kujala, T. Neural discrimination of speech sound changes in a variable context occurs irrespective of attention and explicit awareness. Biol. Psychol. 2018, 132, 217–227. [Google Scholar] [CrossRef]
- Liu, H.M.; Chen, Y.; Tsao, F.M. Developmental changes in mismatch responses to Mandarin consonants and lexical tones from early to middle childhood. PLoS ONE 2014, 9, e95587. [Google Scholar] [CrossRef] [PubMed]
- Čeponienė, R.; Cummings, A.; Wulfeck, B.; Ballantyne, A.; Townsend, J. Spectral vs. temporal auditory processing in specific language impairment: A developmental ERP study. Brain Lang. 2009, 110, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Vidal, J.; Bonnet-Brilhault, F.; Roux, S.; Bruneau, N. Auditory evoked potentials to tones and syllables in adults: Evidence of specific influence on N250 wave. Neurosci. Lett. 2005, 378, 145–149. [Google Scholar] [CrossRef]
- Ohde, R.N.; German, S.R. Formant onsets and formant transitions as developmental cues to vowel perception. J. Acoust. Soc. Am. 2011, 130, 1628–1642. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.R.; Chang, Y.C.; Iskarous, K. Vowel coarticulation: Landmark statistics measure vowel aggression. J. Acoust. Soc. Am. 2015, 138, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Kalaiah, M.K.; Bhat, J.S. Effect of vowel context on the recognition of initial consonants in kannada. J. Audiol. Otol. 2017, 21, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Edwards, J.; Beckman, M.E. Contrast and covert contrast: The phonetic development of voiceless sibilant fricatives in English and Japanese toddlers. J. Phon. 2009, 37, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Liker, M.; Gibbon, F.E. Differences in EPG contact dynamics between voiced and voiceless lingual fricatives. J. Int. Phon. Assoc. 2013, 43, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Cheour, M.; Leppänen, P.H.; Kraus, N. Mismatch negativity (MMN) as a tool for investigating auditory discrimination and sensory memory in infants and children. Clin. Neurophysiol. 2000, 111, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Näätänen, R.; Kreegipuu, K. The mismatch negativity (MMN). In The Oxford Handbook of Event-Related Potential Components; Luck, S.J., Kappenman, E.S., Eds.; Oxford University Press: Oxford, UK, 2012; pp. 143–157. [Google Scholar]
- Savela, J.; Kujala, T.; Tuomainen, J.; Ek, M.; Aaltonen, O.; Näätänen, R. The mismatch negativity and reaction time as indices of the perceptual distance between the corresponding vowels of two related languages. Cogn. Brain Res. 2003, 16, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, N.; Schröger, E. On the development of auditory distraction: A review. PsyCh J. 2014, 3, 72–91. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, X.; Gao, Y.; Wang, Z.; Wang, X. Meaning enhances discrimination of merged phonemes: A mismatch negativity study. Brain Res. 2019, 1724, 146433. [Google Scholar] [CrossRef]
- Horváth, J.; Roeber, U.; Schröger, E. The utility of brief, spectrally rich, dynamic sounds in the passive oddball paradigm. Neurosci. Lett. 2009, 461, 262–265. [Google Scholar] [CrossRef]
- Jacobsen, T.; Horváth, J.; Schröger, E.; Lattner, S.; Widmann, A.; Winkler, I. Pre-attentive auditory processing of lexicality. Brain Lang. 2004, 88, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, T.; Bäß, P.; Roye, A.; Winkler, I.; Schröger, E.; Horváth, J. Word class and word frequency in the MMN looking glass. Brain Lang. 2021, 218, 104964. [Google Scholar] [CrossRef] [PubMed]
- Bitz, U.; Gust, K.; Spitzer, M.; Kiefer, M. Phonological deficit in school children is reflected in the mismatch negativity. NeuroReport 2007, 18, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S. Phonological recoding deficit in working memory of dyslexic teenagers. J. Res. Read. 2000, 23, 28–40. [Google Scholar] [CrossRef]
- Swan, D.; Goswami, U. Phonological awareness deficits in developmental dyslexia and the phonological representations hypothesis. J. Exp. Child Psychol. 1997, 66, 18–41. [Google Scholar] [CrossRef]
- Tijms, J. Verbal memory and phonological processing in dyslexia. J. Res. Read. 2004, 27, 300–310. [Google Scholar] [CrossRef]
- Datta, H.; Hestvik, A.; Vidal, N.; Tessel, C.; Hisagi, M.; Wróbleski, M.; Shafer, V.L. Automaticity of speech processing in early bilingual adults and children. Biling. Lang. Cogn. 2020, 23, 429–445. [Google Scholar] [CrossRef] [Green Version]
- Azaiez, N.; Loberg, O.; Hamalainen, J.; Leppänen, P.H. Brain source correlates of speech perception and reading processes in children with and without reading difficulties. Front. Neurosci. 2022, 16, 921977. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ALLU | Word Chain | PW Reading | RAVEN | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | t-Value | p-Value | Cohen’s d | t-Value | p-Value | Cohen’s d | t-Value | p-Value | Cohen’s d | t-Value | p-Value | Cohen’s d | |
| CTR vs. RD | 111 | <0.001 | 1.824 | <0.001 | 2.017 | <0.001 | 1.39 | 0.192 | 0.26 | ||||
| M | SD | M | SD | M | SD | M | SD | ||||||
| CTR | 4.36 | 52.39 | 8.80 | 0.94 | 45.39 | 12.66 | 3.23 | 32.96 | 3.40 | 3.14 | 22.95 | 3.37 | |
| RD | 1.00 | 36.69 | 7.88 | 0.99 | 22.23 | 5.89 | 1.25 | 27.11 | 6.19 | 3.95 | 22.11 | 2.59 | |
| MMR (150–300 ms) | LDN (450–850 ms) | |
|---|---|---|
| CTR group | ||
| she-shoevs.sii-suu | Cluster [171–279 ms], Positive (max. 231 ms), p < 0.001 Cluster [170–294 ms], Negative (max. 253 ms), p < 0.001 | ns |
| shy-shoevs.sai-suu | Cluster [166–252 ms], Positive (max. 208 ms), p < 0.001 Cluster [150–254 ms] Negative (max. 206 ms) p < 0.001 | ns |
| RD group | ||
| she-shoevs.sii-suu | Cluster [152–276 ms], Positive (max. 195 ms), p < 0.005 | ns |
| shy-shoevs.sai-suu | Cluster [150–277 ms], Positive (max. 205 ms), p < 0.001 Cluster [150–281 ms], Negative (max. 239 ms), p < 0.002 | ns |
| Condition (Deviant–Standard) | ||
|---|---|---|
| MMR (150–300 ms) | LDN (450–850 ms) | |
| sii-suu | Cluster [154–300 ms], Negative (max. 246 ms), p < 0.001 Cluster [150–300 ms], Positive (max. 258 ms), p < 0.001 | Cluster [532–800 ms], Positive (max. 661 ms), p < 0.02 |
| sai-suu | Cluster [150–300 ms], Negative (max. 203 ms), p < 0.001 Cluster [150–300 ms], Positive (max. 217 ms), p < 0.001 | Cluster [450–849 ms], Negative (max. 586 ms), p < 0.001 Cluster [450–850 ms], Positive (max. 640 ms), p < 0.001 |
| she-shoe | Cluster [207–295 ms], Negative (max. 294 ms), p <0.02 | Cluster [450–849 ms], Negative (max. 503 ms), p < 0.005 |
| shy-shoe | Cluster [183–234 ms], Negative (max. 209 ms), p < 0.001 Cluster [175–275 ms], Positive (max. 214 ms), p < 0.001 Cluster [231–295 ms], Positive (max. 257 ms), p < 0.03 Cluster [263–297 ms], Negative (max. 290 ms), p < 0.03 | Cluster [450–850 ms], Positive (max. 575 ms), p < 0.001 Cluster [450–849 ms], Negative (max. 671 ms), p < 0.001 |
| Condition (Deviant–Standard) | ||
|---|---|---|
| MMR (150−300 ms) | LDN (450−850 ms) | |
| sii-suu | ns | ns |
| sai-suu | Cluster [150–300 ms], Negative (max. 232 ms), p < 0.001 Cluster [150–289 ms], Positive (max. 156 ms), p < 0.003 | ns |
| she-shoe | Cluster [150–300 ms], Positive(max 177 ms), p < 0.001 Cluster [150–259 ms], Negative(max. 199 m), p < 0.005 | ns |
| shy-shoe | Cluster [150–295 ms], Negative (max. 209 ms), p < 0.001 Cluster [182–294 ms], Positive (max.294 ms), p < 0.002 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azaiez, N.; Loberg, O.; Lohvansuu, K.; Ylinen, S.; Hämäläinen, J.A.; Leppänen, P.H.T. Discriminatory Brain Processes of Native and Foreign Language in Children with and without Reading Difficulties. Brain Sci. 2023, 13, 76. https://doi.org/10.3390/brainsci13010076
Azaiez N, Loberg O, Lohvansuu K, Ylinen S, Hämäläinen JA, Leppänen PHT. Discriminatory Brain Processes of Native and Foreign Language in Children with and without Reading Difficulties. Brain Sciences. 2023; 13(1):76. https://doi.org/10.3390/brainsci13010076
Chicago/Turabian StyleAzaiez, Najla, Otto Loberg, Kaisa Lohvansuu, Sari Ylinen, Jarmo A. Hämäläinen, and Paavo H. T. Leppänen. 2023. "Discriminatory Brain Processes of Native and Foreign Language in Children with and without Reading Difficulties" Brain Sciences 13, no. 1: 76. https://doi.org/10.3390/brainsci13010076