
Inter-Task Transfer of Prism Adaptation through Motor Imagery †
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedure
2.2.1. Familiarization
2.2.2. Pre-Tests
2.2.3. Exposure
2.2.4. Post-Tests
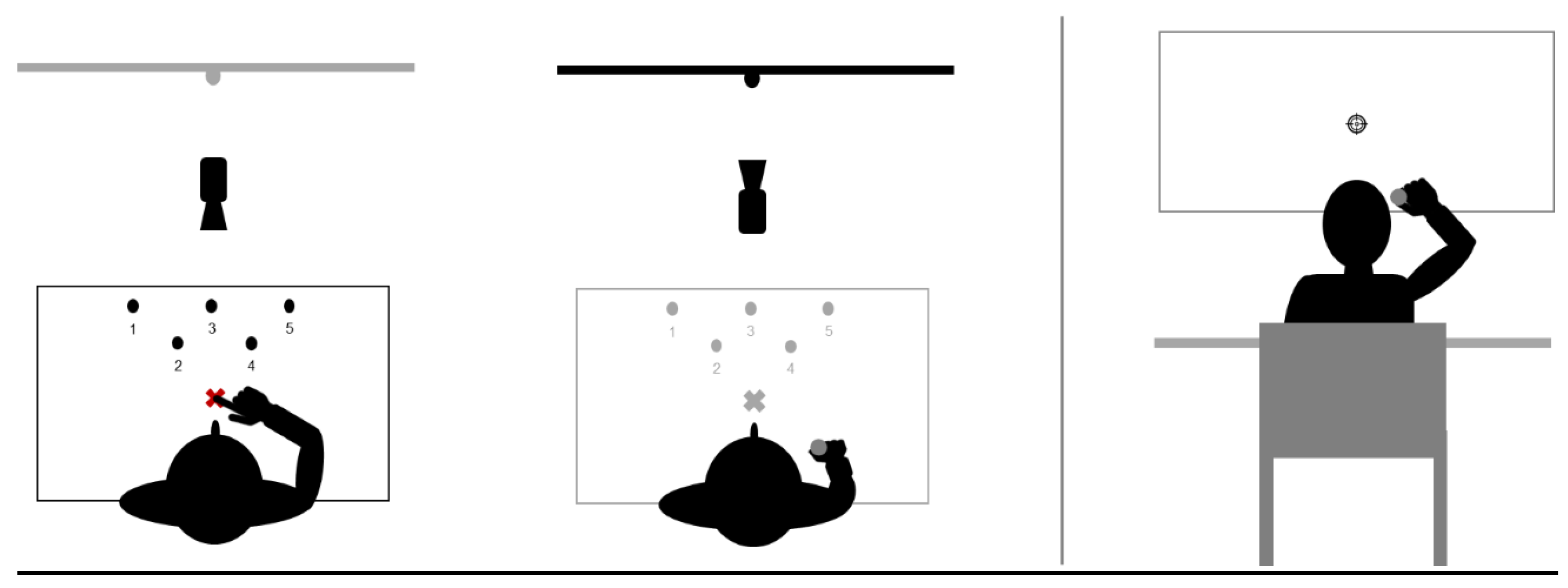
2.3. Tasks
2.3.1. Pointing Task
- Sequences of pointing movements (familiarization and exposure)
- Unique straightforward pointing movements (pre and post-tests).
2.3.2. Throwing Task
- Closed-loop throwing movements (familiarization).
- Open-loop throwing movements (pre-tests and post-tests).
2.4. Data Collection and Processing
2.5. Statistical Analysis
3. Results
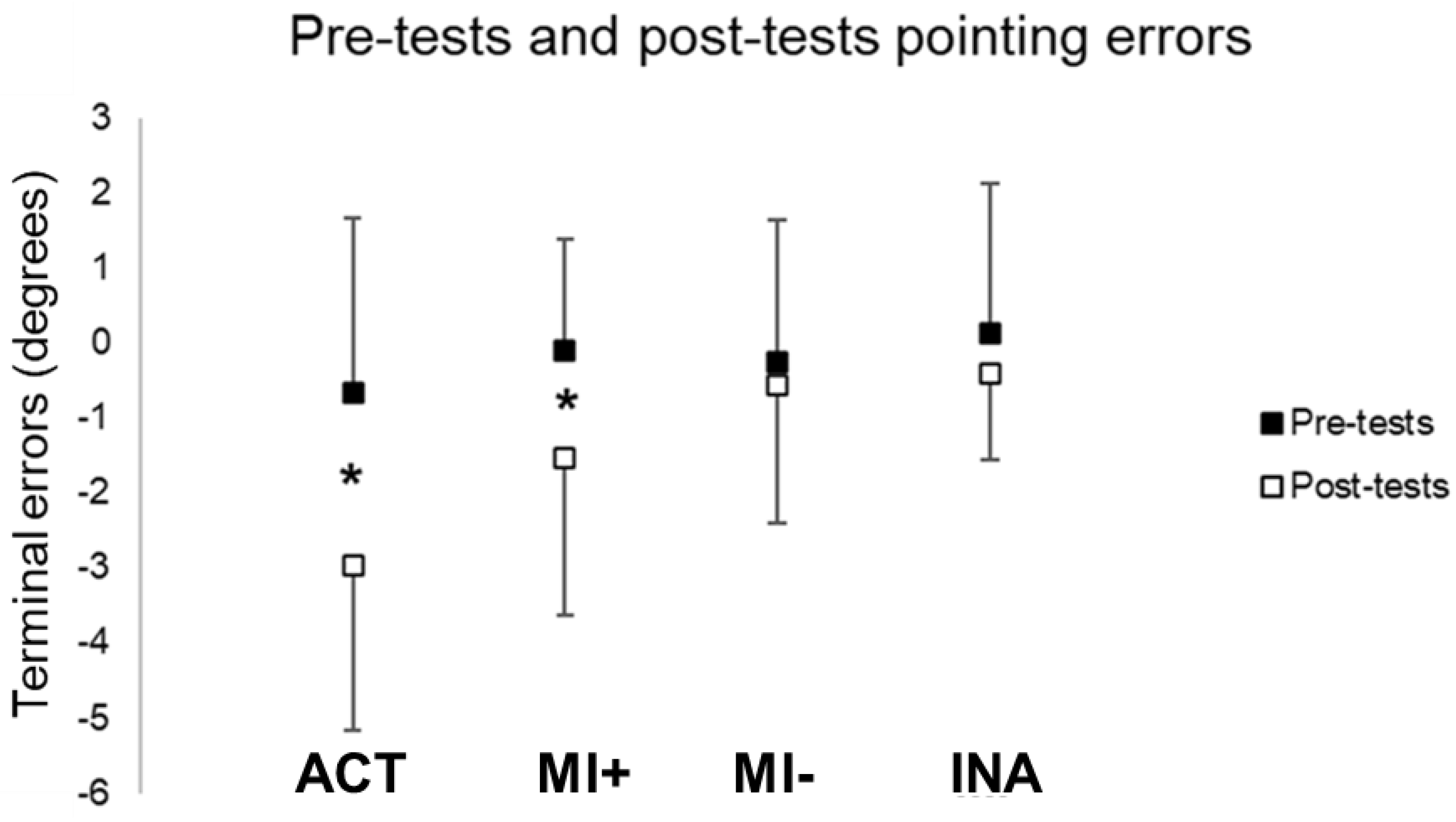
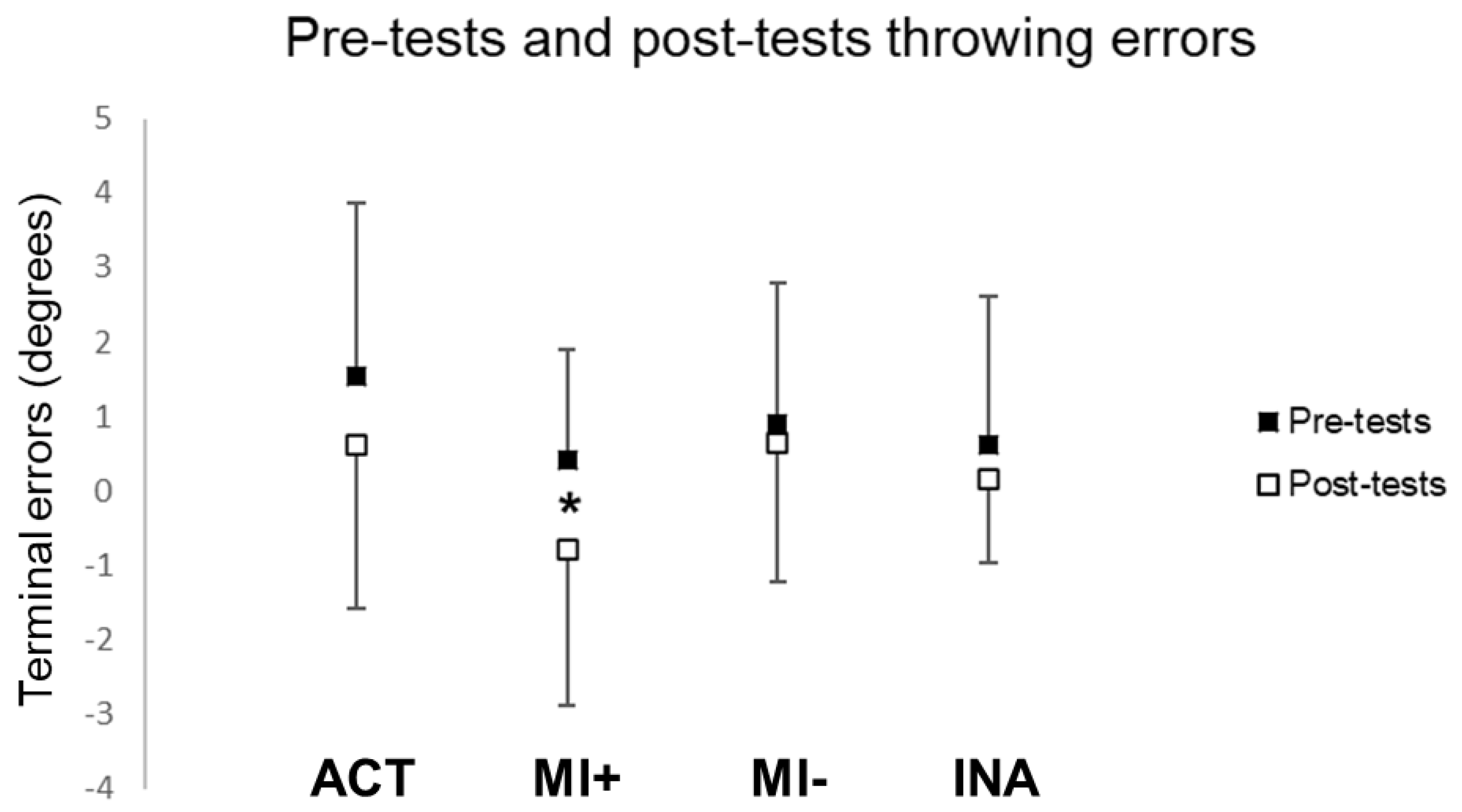
3.1. Comparisons between Pre-Tests and Post-Tests Performances
3.1.1. Pointing Performances
3.1.2. Throwing Performances
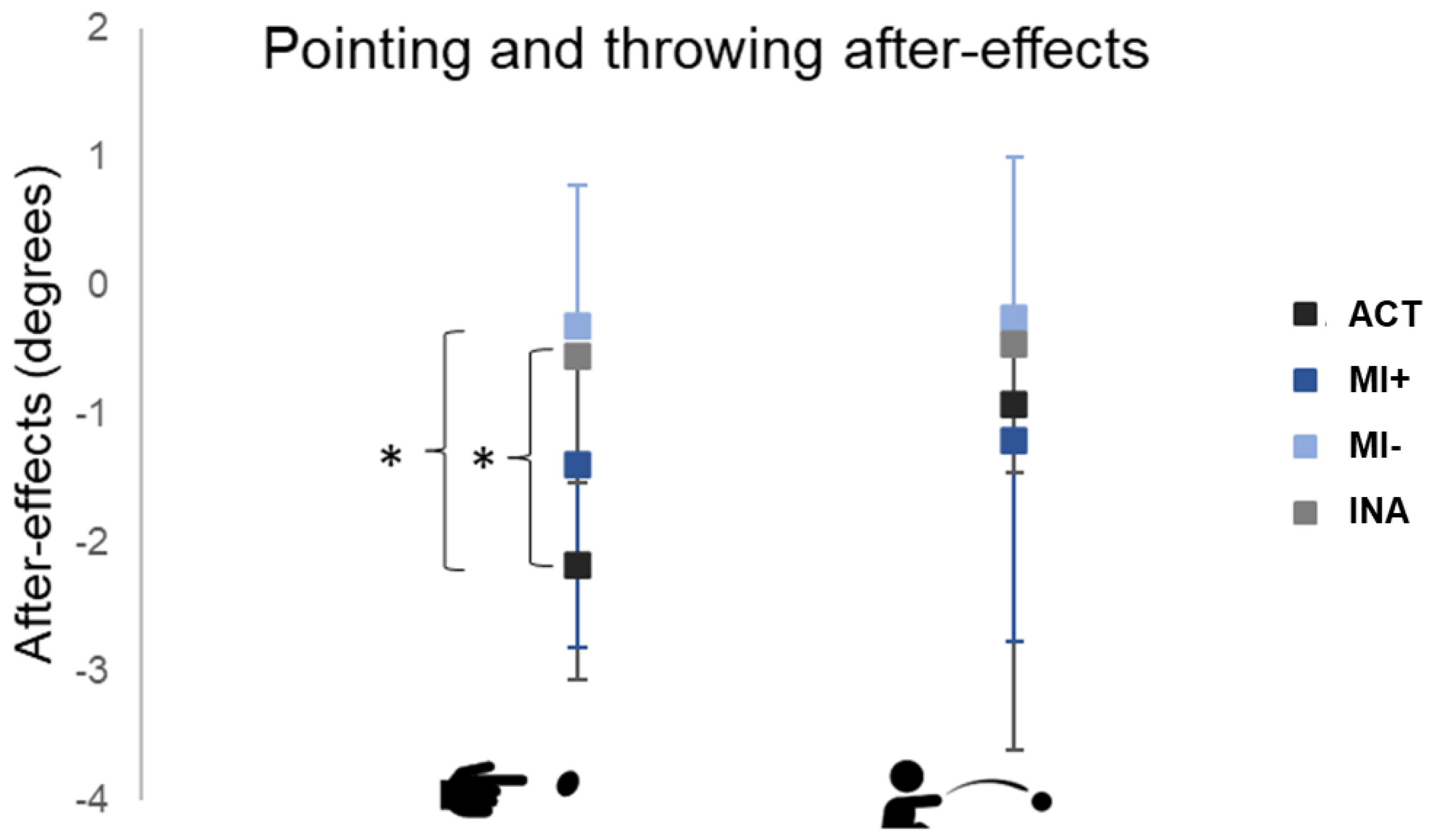
3.2. Comparisons between Groups
3.3. Task Order Influences Pointing and Throwing After-Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleury, L.; Prablanc, C.; Priot, A.-E. Do Prism and Other Adaptation Paradigms Really Measure the Same Processes? Cortex J. Devoted Study Nerv. Syst. Behav. 2019, 119, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Poggio, T.; Bizzi, E. Generalization in Vision and Motor Control. Nature 2004, 431, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Roemmich, R.T.; Bastian, A.J. Closing the Loop: From Motor Neuroscience to Neurorehabilitation. Annu. Rev. Neurosci. 2018, 41, 415–429. [Google Scholar] [CrossRef]
- Redding, G.M.; Rossetti, Y.; Wallace, B. Applications of Prism Adaptation: A Tutorial in Theory and Method. Neurosci. Biobehav. Rev. 2005, 29, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Prablanc, C.; Panico, F.; Fleury, L.; Pisella, L.; Nijboer, T.; Kitazawa, S.; Rossetti, Y. Adapting Terminology: Clarifying Prism Adaptation Vocabulary, Concepts, and Methods. Neurosci. Res. 2019, 153, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.J.; Hallett, M.; Funkenstein, H.H. Adaptation to Lateral Displacement of Vision in Patients with Lesions of the Central Nervous System. Neurology 1983, 33, 766–772. [Google Scholar] [CrossRef]
- Redding, G.M.; Wallace, B. Strategie Calibration and Spatial Alignment: A Model From Prism Adaptation. J. Mot. Behav. 2002, 34, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Petitet, P.; O’Reilly, J.X.; O’Shea, J. Towards a Neuro-Computational Account of Prism Adaptation. Neuropsychologia 2017, 115, 188–203. [Google Scholar] [CrossRef] [PubMed]
- Held, R.; Freedman, S.J. Plasticity in Human Sensorimotor Control. Science 1963, 142, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgeois, A.; Schmid, A.; Turri, F.; Schnider, A.; Ptak, R. Visual but Not Auditory-Verbal Feedback Induces Aftereffects Following Adaptation to Virtual Prisms. Front. Neurosci. 2021, 15, 658353. [Google Scholar] [CrossRef]
- Michel, C.; Pisella, L.; Prablanc, C.; Rode, G.; Rossetti, Y. Enhancing Visuomotor Adaptation by Reducing Error Signals: Single-Step (Aware) versus Multiple-Step (Unaware) Exposure to Wedge Prisms. J. Cogn. Neurosci. 2007, 19, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Gaveau, V.; Prablanc, C.; Laurent, D.; Rossetti, Y.; Priot, A.-E. Visuomotor Adaptation Needs a Validation of Prediction Error by Feedback Error. Front. Hum. Neurosci. 2014, 8, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanajima, R.; Shadmehr, R.; Ohminami, S.; Tsutsumi, R.; Shirota, Y.; Shimizu, T.; Tanaka, N.; Terao, Y.; Tsuji, S.; Ugawa, Y.; et al. Modulation of Error-Sensitivity during a Prism Adaptation Task in People with Cerebellar Degeneration. J. Neurophysiol. 2015, 114, 2460–2471. [Google Scholar] [CrossRef] [Green Version]
- Hatada, Y.; Miall, R.C.; Rossetti, Y. Two Waves of a Long-Lasting Aftereffect of Prism Adaptation Measured over 7 Days. Exp. Brain Res. 2006, 169, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Panico, F.; Sagliano, L.; Nozzolillo, C.; Trojano, L.; Rossetti, Y. Cerebellar Contribution to Spatial Realignment: A TDCS Study during Multiple-Step Prism Adaptation. Neuropsychologia 2018, 112, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Held, R.; Mikaelian, H. Motor-Sensory Feedback versus Need in Adaptation to Rearrangement. Percept. Mot. Skills 1964, 18, 685–688. [Google Scholar] [CrossRef]
- Wilf, M.; Cerra Cheraka, M.; Jeanneret, M.; Ott, R.; Perrin, H.; Crottaz-Herbette, S.; Serino, A. Combined Virtual Reality and Haptic Robotics Induce Space and Movement Invariant Sensorimotor Adaptation. Neuropsychologia 2021, 150, 107692. [Google Scholar] [CrossRef]
- Cressman, E.K.; Henriques, D.Y.P. Reach Adaptation and Proprioceptive Recalibration Following Exposure to Misaligned Sensory Input. J. Neurophysiol. 2010, 103, 1888–1895. [Google Scholar] [CrossRef] [Green Version]
- Guillot, A.; Collet, C. Duration of Mentally Simulated Movement: A Review. J. Mot. Behav. 2005, 37, 10–20. [Google Scholar] [CrossRef]
- Guillot, A.; Di Rienzo, F.; Macintyre, T.; Moran, A.; Collet, C. Imagining Is Not Doing but Involves Specific Motor Commands: A Review of Experimental Data Related to Motor Inhibition. Front. Hum. Neurosci. 2012, 6, 247. [Google Scholar] [CrossRef]
- Jeannerod, M. Neural Simulation of Action: A Unifying Mechanism for Motor Cognition. NeuroImage 2001, 14, S103–S109. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Collet, C.; Nguyen, V.A.; Malouin, F.; Richards, C.; Doyon, J. Functional Neuroanatomical Networks Associated with Expertise in Motor Imagery. NeuroImage 2008, 41, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, F.; Debarnot, U.; Daligault, S.; Saruco, E.; Delpuech, C.; Doyon, J.; Collet, C.; Guillot, A. Online and Offline Performance Gains Following Motor Imagery Practice: A Comprehensive Review of Behavioral and Neuroimaging Studies. Front. Hum. Neurosci. 2016, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Ruffino, C.; Papaxanthis, C.; Lebon, F. Neural Plasticity during Motor Learning with Motor Imagery Practice: Review and Perspectives. Neuroscience 2017, 341, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.; Gaveau, J.; Pozzo, T.; Papaxanthis, C. Prism Adaptation by Mental Practice. Cortex J. Devoted Study Nerv. Syst. Behav. 2013, 49, 2249–2259. [Google Scholar] [CrossRef]
- Kawato, M. Internal Models for Motor Control and Trajectory Planning. Curr. Opin. Neurobiol. 1999, 9, 718–727. [Google Scholar] [CrossRef]
- Miall, R.C.; Wolpert, D.M. Forward Models for Physiological Motor Control. Neural Netw. 1996, 9, 1265–1279. [Google Scholar] [CrossRef]
- Kilteni, K.; Andersson, B.J.; Houborg, C.; Ehrsson, H.H. Motor Imagery Involves Predicting the Sensory Consequences of the Imagined Movement. Nat. Commun. 2018, 9, 1617. [Google Scholar] [CrossRef] [Green Version]
- Rannaud Monany, D.; Barbiero, M.; Lebon, F.; Babič, J.; Blohm, G.; Nozaki, D.; White, O. Motor Imagery Helps Updating Internal Models during Microgravity Exposure. J. Neurophysiol. 2022, 127, 434–443. [Google Scholar] [CrossRef]
- White, O.; Gaveau, J.; Bringoux, L.; Crevecoeur, F. The Gravitational Imprint on Sensorimotor Planning and Control. J. Neurophysiol. 2020, 124, 4–19. [Google Scholar] [CrossRef]
- Fleury, L.; Pastor, D.; Revol, P.; Delporte, L.; Rossetti, Y. Inter-Task Transfer of Prism Adaptation Depends on Exposed Task Mastery. Sci. Rep. 2020, 10, 5687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, F.L. Perceptual and Cognitive Spatial Learning. J. Exp. Psychol. Hum. Percept. Perform. 1993, 19, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Redding, G.M.; Wallace, B. Generalization of Prism Adaptation. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 1006–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girardi, M.; McIntosh, R.D.; Michel, C.; Vallar, G.; Rossetti, Y. Sensorimotor Effects on Central Space Representation: Prism Adaptation Influences Haptic and Visual Representations in Normal Subjects. Neuropsychologia 2004, 42, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.; Vernet, P.; Courtine, G.; Ballay, Y.; Pozzo, T. Asymmetrical After-Effects of Prism Adaptation during Goal Oriented Locomotion. Exp. Brain Res. 2008, 185, 259–268. [Google Scholar] [CrossRef]
- Morton, S.M.; Lang, C.E.; Bastian, A.J. Inter- and Intra-Limb Generalization of Adaptation during Catching. Exp. Brain Res. 2001, 141, 438–445. [Google Scholar] [CrossRef]
- Fernandez-Ruiz, J.; Diaz, R.; Hall-Haro, C.; Vergara, P.; Mischner, J.; Nuñez, L.; Drucker-Colin, R.; Ochoa, A.; Alonso, M.E. Normal Prism Adaptation but Reduced After-Effect in Basal Ganglia Disorders Using a Throwing Task. Eur. J. Neurosci. 2003, 18, 689–694. [Google Scholar] [CrossRef]
- Jacquin-Courtois, S.; Rode, G.; Pisella, L.; Boisson, D.; Rossetti, Y. Wheel-Chair Driving Improvement Following Visuo-Manual Prism Adaptation. Cortex J. Devoted Study Nerv. Syst. Behav. 2008, 44, 90–96. [Google Scholar] [CrossRef]
- Hall, C.R.; Martin, K.A. Measuring Movement Imagery Abilities: A Revision of the Movement Imagery Questionnaire. J. Ment. Imag. 1997, 21, 143–154. [Google Scholar]
- Rossetti, Y.; Tadary, B.; Prablanc, C. Optimal Contributions of Head and Eye Positions to Spatial Accuracy in Man Tested by Visually Directed Pointing. Exp. Brain Res. 1994, 97, 487–496. [Google Scholar] [CrossRef]
- Michel, C. Beyond the Sensorimotor Plasticity: Cognitive Expansion of Prism Adaptation in Healthy Individuals. Front. Psychol. 2015, 6, 1979. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.U.; Kraemer, W.J.; Häkkinen, K.; Humphries, B.J.; Murphy, A.J. Kinematics, Kinetics, and Muscle Activation during Explosive Upper Body Movements. J. Appl. Biomech. 1996, 12, 31–43. [Google Scholar] [CrossRef]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error Correction, Sensory Prediction, and Adaptation in Motor Control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Meulen, M.; Allali, G.; Rieger, S.W.; Assal, F.; Vuilleumier, P. The Influence of Individual Motor Imagery Ability on Cerebral Recruitment during Gait Imagery. Hum. Brain Mapp. 2014, 35, 455–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, R.; Miall, R.C.; Galea, J.M. No Consistent Effect of Cerebellar Transcranial Direct Current Stimulation on Visuomotor Adaptation. J. Neurophysiol. 2017, 118, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Redding, G.M.; Wallace, B. Effects of Pointing Rate and Availability of Visual Feedback on Visual and Proprioceptive Components of Prism Adaptation. J. Mot. Behav. 1992, 24, 226–237. [Google Scholar] [CrossRef]
- Redding, G.M.; Wallace, B. Components of Prism Adaptation in Terminal and Concurrent Exposure: Organization of the Eye-Hand Coordination Loop. Percept. Psychophys. 1988, 44, 59–68. [Google Scholar] [CrossRef]
- Di Rienzo, F.; Collet, C.; Hoyek, N.; Guillot, A. Impact of Neurologic Deficits on Motor Imagery: A Systematic Review of Clinical Evaluations. Neuropsychol. Rev. 2014, 24, 116–147. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleury, L.; Dreyer, L.; El Makkaoui, R.; Leroy, E.; Rossetti, Y.; Collet, C. Inter-Task Transfer of Prism Adaptation through Motor Imagery. Brain Sci. 2023, 13, 114. https://doi.org/10.3390/brainsci13010114
Fleury L, Dreyer L, El Makkaoui R, Leroy E, Rossetti Y, Collet C. Inter-Task Transfer of Prism Adaptation through Motor Imagery. Brain Sciences. 2023; 13(1):114. https://doi.org/10.3390/brainsci13010114
Chicago/Turabian StyleFleury, Lisa, Léa Dreyer, Rola El Makkaoui, Elise Leroy, Yves Rossetti, and Christian Collet. 2023. "Inter-Task Transfer of Prism Adaptation through Motor Imagery" Brain Sciences 13, no. 1: 114. https://doi.org/10.3390/brainsci13010114