
A Comprehensive Review on Anti-Inflammatory Response of Flavonoids in Experimentally-Induced Epileptic Seizures
 ,
,  , ,
, ,
Abstract
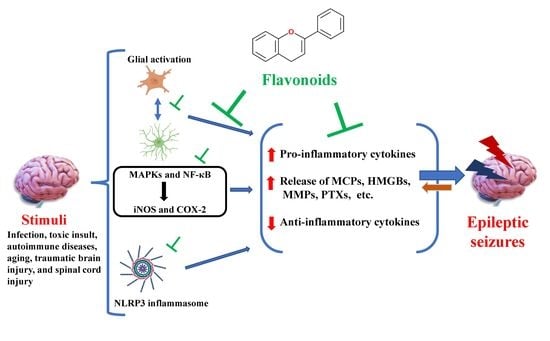
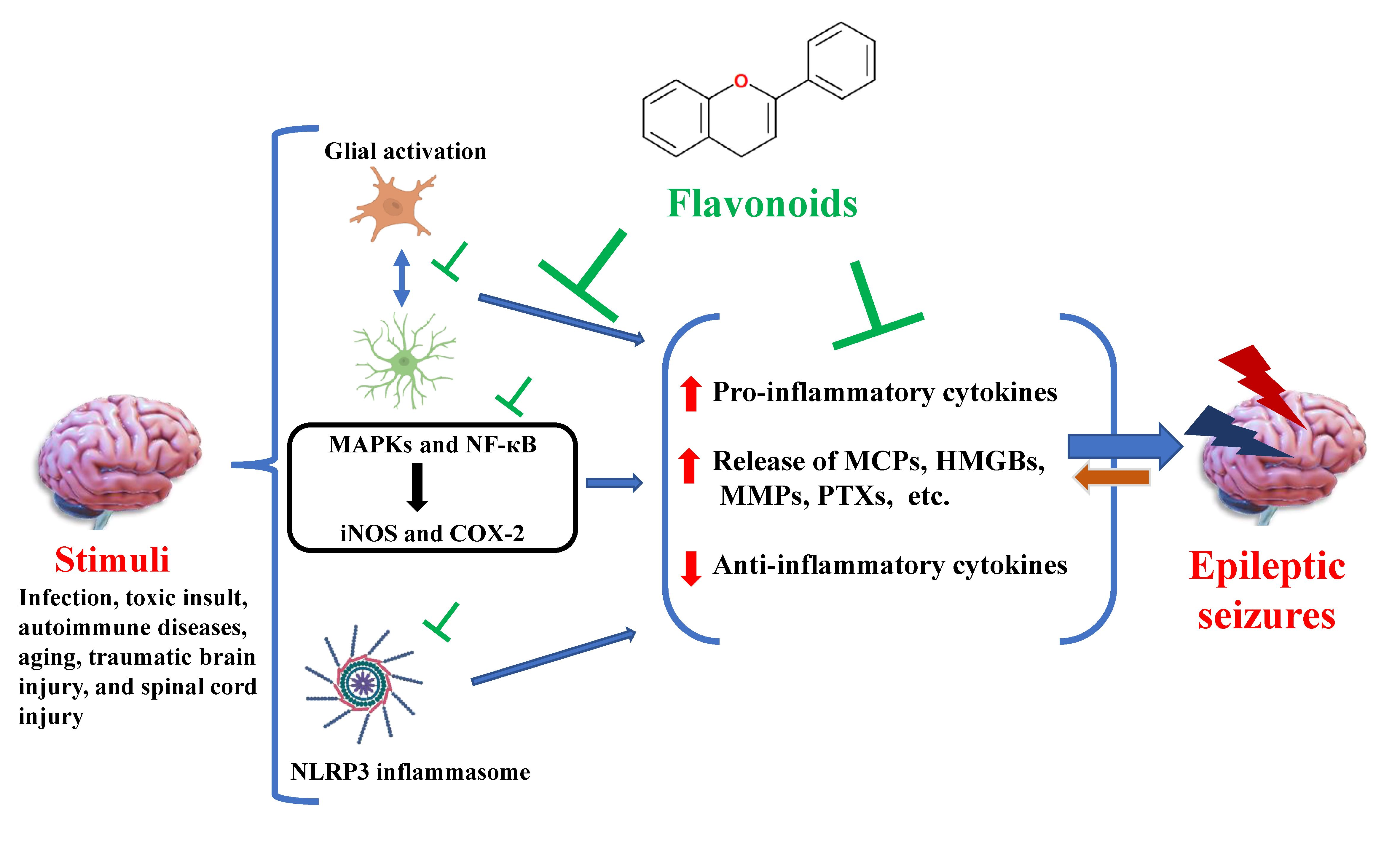
:
1. Introduction
2. Neuroinflammation in Epilepsy
3. Flavonoids
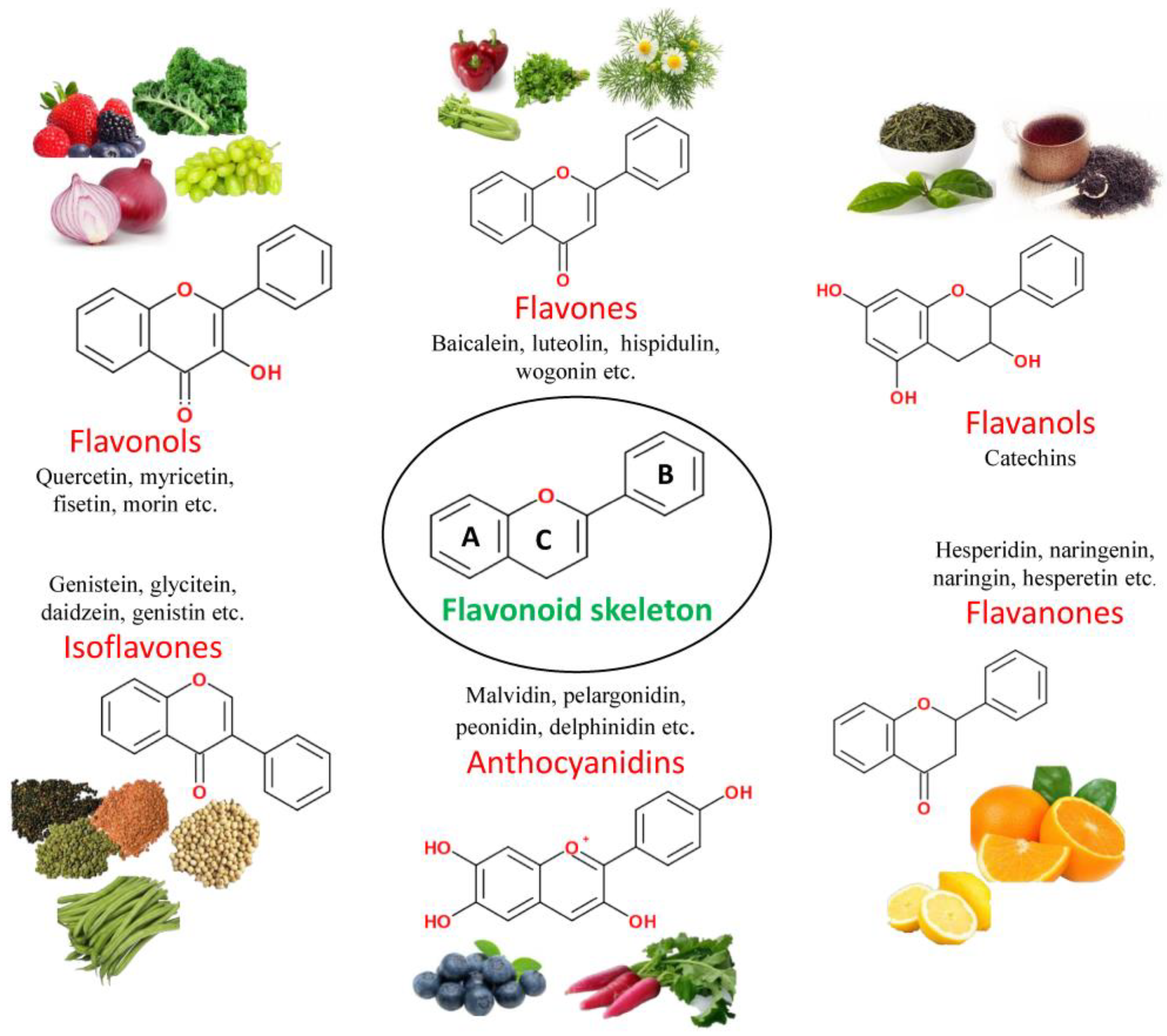
3.1. Classification of Flavonoids
3.1.1. Flavones
3.1.2. Flavonols
3.1.3. Flavanones
3.1.4. Anthocyanidins
3.1.5. Isoflavones
3.1.6. Flavanols
4. Flavonoids with Anti-Inflammatory Response in Epilepsy
4.1. Baicalein
4.2. Luteolin
4.3. Hispidulin
4.4. Schaftoside
4.5. Vitexin
4.6. Wogonin
4.7. Rhoifolin
4.8. Amentoflavone
4.9. Quercetin
4.10. Rutin
4.11. Fisetin
4.12. Kaempferol
4.13. Morin
4.14. Myricetin
4.15. Myricitrin
4.16. Galangin
4.17. Naringin
4.18. Naringenin
4.19. Hesperetin
4.20. Hesperidin
4.21. Silibinin
4.22. Genistein
4.23. Catechin
4.24. Epigallocatechin-3-Gallate
5. Mechanism of Action of Flavonoids in Epilepsy
6. Conclusion and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, R.S.K.; Leung, C.Y.W.; Ho, C.C.A.; Yung, A. Classifications of Seizures and Epilepsies, Where Are We?—A Brief Historical Review and Update. J. Formos. Med. Assoc. 2017, 116, 736–741. [Google Scholar] [CrossRef]
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE Official Report: A Practical Clinical Definition of Epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Mula, M.; Sander, J.W. Psychosocial Aspects of Epilepsy: A Wider Approach. BJPsych Open 2016, 2, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Steiger, B.K.; Jokeit, H. Why Epilepsy Challenges Social Life. Seizure 2017, 44, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Jovel, C.; Toledano, R.; Aledo-Serrano, Á.; García-Morales, I.; Gil-Nagel, A. Epidemiological Profile of Epilepsy in Low Income Populations. Seizure 2018, 56, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Shorvon, S.D. The Causes of Epilepsy: Changing Concepts of Etiology of Epilepsy over the Past 150 Years. Epilepsia 2011, 52, 1033–1044. [Google Scholar] [CrossRef]
- Kandratavicius, L.; Balista, P.; Lopes-Aguiar, C.; Ruggiero, R.; Umeoka, E.; Garcia-Cairasco, N.; Bueno-Junior, L.; Leite, J. Animal Models of Epilepsy: Use and Limitations. Neuropsychiatr. Dis. Treat. 2014, 10, 1693–1705. [Google Scholar] [CrossRef] [Green Version]
- Grone, B.P.; Baraban, S.C. Animal Models in Epilepsy Research: Legacies and New Directions. Nat. Neurosci. 2015, 18, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Aronica, E.; Bauer, S.; Bozzi, Y.; Caleo, M.; Dingledine, R.; Gorter, J.A.; Henshall, D.C.; Kaufer, D.; Koh, S.; Löscher, W.; et al. Neuroinflammatory Targets and Treatments for Epilepsy Validated in Experimental Models. Epilepsia 2017, 58, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Barker-Haliski, M.L.; Löscher, W.; White, H.S.; Galanopoulou, A.S. Neuroinflammation in Epileptogenesis: Insights and Translational Perspectives from New Models of Epilepsy. Epilepsia 2017, 58, 39–47. [Google Scholar] [CrossRef] [PubMed]
- de Vries, E.E.; van den Munckhof, B.; Braun, K.P.J.; van Royen-Kerkhof, A.; de Jager, W.; Jansen, F.E. Inflammatory Mediators in Human Epilepsy: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2016, 63, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; French, J.; Bartfai, T.; Baram, T.Z. The Role of Inflammation in Epilepsy. Nat. Rev. Neurol. 2011, 7, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingledine, R.; Varvel, N.H.; Dudek, F.E. When and How Do Seizures Kill Neurons, and Is Cell Death Relevant to Epileptogenesis? Adv. Exp. Med. Biol. 2014, 813, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Laxer, K.D.; Trinka, E.; Hirsch, L.J.; Cendes, F.; Langfitt, J.; Delanty, N.; Resnick, T.; Benbadis, S.R. The Consequences of Refractory Epilepsy and Its Treatment. Epilepsy Behav. 2014, 37, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, S.; Romano, A. Production of plant secondary metabolites by using biotechnological tools. In Secondary Metabolites–Sources and Applications; InTech: London, UK, 2018. [Google Scholar]
- Costa, M.; Sezgin-Bayindir, Z.; Losada-Barreiro, S.; Paiva-Martins, F.; Saso, L.; Bravo-Díaz, C. Polyphenols as Antioxidants for Extending Food Shelf-Life and in the Prevention of Health Diseases: Encapsulation and Interfacial Phenomena. Biomedicines 2021, 9, 1909. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Sobhani, M.; Hosein Farzaei, M.; Kiani, S.; Khodarahmi, R. Immunomodulatory; Anti-Inflammatory/Antioxidant Effects of Polyphenols: A Comparative Review on the Parental Compounds and Their Metabolites. Food Rev. Int. 2021, 37, 759–811. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant Properties of Flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- Keddy, P.G.W.; Dunlop, K.; Warford, J.; Samson, M.L.; Jones, Q.R.D.; Rupasinghe, H.P.V.; Robertson, G.S. Neuroprotective and Anti-Inflammatory Effects of the Flavonoid-Enriched Fraction AF4 in a Mouse Model of Hypoxic-Ischemic Brain Injury. PLoS ONE 2012, 7, e51324. [Google Scholar] [CrossRef] [Green Version]
- Dourado, N.S.; Souza, C.D.S.; De Almeida, M.M.A.; Bispo da Silva, A.; Dos Santos, B.L.; Silva, V.D.A.; De Assis, A.M.; da Silva, J.S.; Souza, D.O.; Costa, M.D.F.D.; et al. Neuroimmunomodulatory and Neuroprotective Effects of the Flavonoid Apigenin in in Vitro Models of Neuroinflammation Associated With Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 119. [Google Scholar] [CrossRef]
- Heimfarth, L.; da Silva Nascimento, L.; da Silva, M.D.J.A.; de Lucca Junior, W.; Lima, E.S.; Quintans-Junior, L.J.; da Veiga-Junior, V.F. Neuroprotective and Anti-Inflammatory Effect of Pectolinarigenin, a Flavonoid from Amazonian Aegiphila Integrifolia (Jacq.), against Lipopolysaccharide-Induced Inflammation in Astrocytes via NFкB and MAPK Pathways. Food Chem. Toxicol. 2021, 157, 112538. [Google Scholar] [CrossRef]
- Gilhus, N.E.; Deuschl, G. Neuroinflammation—A Common Thread in Neurological Disorders. Nat. Rev. Neurol. 2019, 15, 429–430. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 2019, 10, 1008. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gómez, J.A.; Kavanagh, E.; Engskog-Vlachos, P.; Engskog, M.K.R.; Herrera, A.J.; Espinosa-Oliva, A.M.; Joseph, B.; Hajji, N.; Venero, J.L.; Burguillos, M.A. Microglia: Agents of the CNS Pro-Inflammatory Response. Cells 2020, 9, 1717. [Google Scholar] [CrossRef]
- Takata, F.; Nakagawa, S.; Matsumoto, J.; Dohgu, S. Blood-Brain Barrier Dysfunction Amplifies the Development of Neuroinflammation: Understanding of Cellular Events in Brain Microvascular Endothelial Cells for Prevention and Treatment of BBB Dysfunction. Front. Cell. Neurosci. 2021, 15, 661838. [Google Scholar] [CrossRef]
- DiSabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The Devil Is in the Details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [Green Version]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory Pathways as Treatment Targets and Biomarkers in Epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Rana, A.; Musto, A.E. The Role of Inflammation in the Development of Epilepsy. J. Neuroinflamm. 2018, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- Pracucci, E.; Pillai, V.; Lamers, D.; Parra, R.; Landi, S. Neuroinflammation: A Signature or a Cause of Epilepsy? Int. J. Mol. Sci. 2021, 22, 6981. [Google Scholar] [CrossRef]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshé, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Falip, M.; Jaraba, S.; Rodríguez-Bel, L.; Castañer, S.; Mora, J.; Arroyo, P.; Miro, J.; Sala-Padró, J.; Martínez-Yélamos, S.; Casasnovas, C.; et al. Seizures and Epilepsy of Autoimmune Origin: A Long-Term Prospective Study. Seizure 2020, 81, 157–165. [Google Scholar] [CrossRef]
- Steriade, C.; Titulaer, M.J.; Vezzani, A.; Sander, J.W.; Thijs, R.D. The Association between Systemic Autoimmune Disorders and Epilepsy and Its Clinical Implications. Brain 2021, 144, 372–390. [Google Scholar] [CrossRef]
- Pitkänen, A.; Löscher, W.; Vezzani, A.; Becker, A.J.; Simonato, M.; Lukasiuk, K.; Gröhn, O.; Bankstahl, J.P.; Friedman, A.; Aronica, E.; et al. Advances in the Development of Biomarkers for Epilepsy. Lancet Neurol. 2016, 15, 843–856. [Google Scholar] [CrossRef]
- Ravizza, T.; Vezzani, A. Pharmacological Targeting of Brain Inflammation in Epilepsy: Therapeutic Perspectives from Experimental and Clinical Studies. Epilepsia Open 2018, 3, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.; Dingledine, R. Molecular Cascades That Mediate the Influence of Inflammation on Epilepsy. Epilepsia 2011, 52, 33–39. [Google Scholar] [CrossRef]
- Alyu, F.; Dikmen, M. Inflammatory Aspects of Epileptogenesis: Contribution of Molecular Inflammatory Mechanisms. Acta Neuropsychiatr. 2017, 29, 1–16. [Google Scholar] [CrossRef]
- Numis, A.L.; Foster-Barber, A.; Deng, X.; Rogers, E.E.; Barkovich, A.J.; Ferriero, D.M.; Glass, H.C. Early Changes in Pro-Inflammatory Cytokine Levels in Neonates with Encephalopathy Are Associated with Remote Epilepsy. Pediatr. Res. 2019, 86, 616–621. [Google Scholar] [CrossRef]
- Chmielewska, N.; Maciejak, P.; Osuch, B.; Kursa, M.B.; Szyndler, J. Pro-Inflammatory Cytokines, but Not Brain- and Extracellular Matrix-Derived Proteins, Are Increased in the Plasma Following Electrically Induced Kindling of Seizures. Pharmacol. Rep. 2021, 73, 506–515. [Google Scholar] [CrossRef]
- Hanke, M.L.; Kielian, T. Toll-like Receptors in Health and Disease in the Brain: Mechanisms and Therapeutic Potential. Clin. Sci. 2011, 121, 367–387. [Google Scholar] [CrossRef] [Green Version]
- Xiang, W.; Chao, Z.-Y.; Feng, D.-Y. Role of Toll-like Receptor/MYD88 Signaling in Neurodegenerative Diseases. Rev. Neurosci. 2015, 26, 407–414. [Google Scholar] [CrossRef]
- Abdelsalam, M.; Abd Elmagid, D.S.; Magdy, H.; El-Sabbagh, A.M.; Mostafa, M. The Association between Toll-like Receptor 4 (TLR4) Genotyping and the Risk of Epilepsy in Children. Egypt J. Med. Hum. Genet. 2020, 21, 61. [Google Scholar] [CrossRef]
- Hsieh, M.Y.; Lin, J.J.; Hsia, S.H.; Huang, J.L.; Yeh, K.W.; Chang, K.W.; Lee, W.I. Diminished Toll-like Receptor Response in Febrile Infection-Related Epilepsy Syndrome (FIRES). Biomed. J. 2020, 43, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Avalos, A.M.; Ploegh, H.L. Accessory Molecules for Toll-like Receptors and Their Function. Nat. Rev. Immunol. 2012, 12, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Paudel, Y.N.; Semple, B.D.; Jones, N.C.; Othman, I.; Shaikh, M.F. High Mobility Group Box 1 (HMGB1) as a Novel Frontier in Epileptogenesis: From Pathogenesis to Therapeutic Approaches. J. Neurochem. 2019, 151, 542–557. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.; Zheng, Y.; Wang, Y.; Chen, Z. HMGB1, Neuronal Excitability and Epilepsy. Acta Epileptologica 2021, 3, 13. [Google Scholar] [CrossRef]
- Spencer, J.P.; Vafeiadou, K.; Williams, R.J.; Vauzour, D. Neuroinflammation: Modulation by flavonoids and mechanisms of action. Mol. Aspects Med. 2011, 33, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Jiang, J.; Ganesh, T.; Yang, M.-S.; Lelutiu, N.; Gueorguieva, P.; Dingledine, R. Cyclooxygenase-2 in Epilepsy. Epilepsia 2014, 55, 17–25. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Zhu, W.; Pan, X.; Wang, Q.; Gao, X.; Wang, C.; Zhang, X.; Liu, Y.; Li, S.; et al. Role of Elevated Thrombospondin-1 in Kainic Acid-Induced Status Epilepticus. Neurosci. Bull. 2020, 36, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Michev, A.; Orsini, A.; Santi, V.; Bassanese, F.; Veraldi, D.; Brambilla, I.; Marseglia, G.L.; Savasta, S.; Foiadelli, T. An Overview of The Role of Tumor Necrosis Factor-Alpha in Epileptogenesis and Its Terapeutic Implications. Acta Biomed. 2021, 92, e2021418. [Google Scholar] [CrossRef]
- Chen, R.; Xue, G.; Hölscher, C. The Role of the TNFα-Mediated Astrocyte Signaling Pathway in Epilepsy. Acta Epileptologica 2021, 3, 24. [Google Scholar] [CrossRef]
- Ishikawa, N.; Kobayashi, Y.; Fujii, Y.; Kobayashi, M. Increased Interleukin-6 and High-Sensitivity C-Reactive Protein Levels in Pediatric Epilepsy Patients with Frequent, Refractory Generalized Motor Seizures. Seizure 2015, 25, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Pelkey, K.A.; Barksdale, E.; Craig, M.T.; Yuan, X.; Sukumaran, M.; Vargish, G.A.; Mitchell, R.M.; Wyeth, M.S.; Petralia, R.S.; Chittajallu, R.; et al. Pentraxins Coordinate Excitatory Synapse Maturation and Circuit Integration of Parvalbumin Interneurons. Neuron 2015, 85, 1257–1272. [Google Scholar] [CrossRef] [Green Version]
- Gómez de San José, N.; Massa, F.; Halbgebauer, S.; Oeckl, P.; Steinacker, P.; Otto, M. Neuronal Pentraxins as Biomarkers of Synaptic Activity: From Physiological Functions to Pathological Changes in Neurodegeneration. J. Neural. Transm. 2022, 129, 207–230. [Google Scholar] [CrossRef]
- Pijet, B.; Stefaniuk, M.; Kostrzewska-Ksiezyk, A.; Tsilibary, P.E.; Tzinia, A.; Kaczmarek, L. Elevation of MMP-9 Levels Promotes Epileptogenesis After Traumatic Brain Injury. Mol. Neurobiol. 2018, 55, 9294–9306. [Google Scholar] [CrossRef] [Green Version]
- Bronisz, E.; Kurkowska-Jastrzębska, I. Matrix Metalloproteinase 9 in Epilepsy: The Role of Neuroinflammation in Seizure Development. Mediat. Inflamm. 2016, 2016, 7369020. [Google Scholar] [CrossRef] [Green Version]
- Dwir, D.; Giangreco, B.; Xin, L.; Tenenbaum, L.; Cabungcal, J.H.; Steullet, P.; Goupil, A.; Cleusix, M.; Jenni, R.; Chtarto, A.; et al. MMP9/RAGE Pathway Overactivation Mediates Redox Dysregulation and Neuroinflammation, Leading to Inhibitory/Excitatory Imbalance: A Reverse Translation Study in Schizophrenia Patients. Mol. Psychiatry 2020, 25, 2889–2904. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 Inflammasome: Molecular Activation and Regulation to Therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Paik, S.; Kim, J.K.; Silwal, P.; Sasakawa, C.; Jo, E.K. An Update on the Regulatory Mechanisms of NLRP3 Inflammasome Activation. Cell. Mol. Immunol. 2021, 18, 1141–1160. [Google Scholar] [CrossRef]
- Yue, J.; Wei, Y.J.; Yang, X.L.; Liu, S.Y.; Yang, H.; Zhang, C.Q. NLRP3 Inflammasome and Endoplasmic Reticulum Stress in the Epileptogenic Zone in Temporal Lobe Epilepsy: Molecular Insights into Their Interdependence. Neuropathol. Appl. Neurobiol. 2020, 46, 770–785. [Google Scholar] [CrossRef]
- Cristina de Brito Toscano, E.; Leandro Marciano Vieira, É.; Boni Rocha Dias, B.; Vidigal Caliari, M.; Paula Gonçalves, A.; Varela Giannetti, A.; Maurício Siqueira, J.; Kimie Suemoto, C.; Elaine Paraizo Leite, R.; Nitrini, R.; et al. NLRP3 and NLRP1 Inflammasomes Are Up-Regulated in Patients with Mesial Temporal Lobe Epilepsy and May Contribute to Overexpression of Caspase-1 and IL-β in Sclerotic Hippocampi. Brain Res. 2021, 1752, 147230. [Google Scholar] [CrossRef]
- Pohlentz, M.S.; Müller, P.; Cases-Cunillera, S.; Opitz, T.; Surges, R.; Hamed, M.; Vatter, H.; Schoch, S.; Becker, A.J.; Pitsch, J. Characterisation of NLRP3 Pathway-Related Neuroinflammation in Temporal Lobe Epilepsy. PLoS ONE 2022, 17, e0271995. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as Natural Phenolic Compounds and Their Role in Therapeutics: An Overview. Future J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Abou Baker, D.H. An Ethnopharmacological Review on the Therapeutical Properties of Flavonoids and Their Mechanisms of Actions: A Comprehensive Review Based on up to Date Knowledge. Toxicol. Rep. 2022, 9, 445–469. [Google Scholar] [CrossRef]
- Billowria, K.; Ali, R.; Rangra, K.; Kumar, R.; Chawla, P.A.; Rouchan, A.; Rangra, N.K. Bioactive Flavonoids: A Comprehensive Review on Pharmacokinetics and Analytical Aspects. Crit. Rev. Anal. Chem. 2022. [Google Scholar] [CrossRef]
- Alex, A.M.; Arehally Marappa, M.; Joghee, S.; Chidambaram, S.B. Therapeutic Benefits of Flavonoids against Neuroinflammation: A Systematic Review. Inflammopharmacology 2022, 30, 111–136. [Google Scholar] [CrossRef]
- Brodowska, K.M. Natural Flavonoids: Classification, Potential Role, and Application of Flavonoid Analogues. Eur. J. Biol. Res. 2017, 7, 108–123. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Kay, C.D.; Pereira-Caro, G.; Ludwig, I.A.; Clifford, M.N.; Crozier, A. Anthocyanins and Flavanones Are More Bioavailable than Previously Perceived: A Review of Recent Evidence. Annu. Rev. Food Sci. Technol. 2017, 8, 155–180. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of Anthocyanins in Common Foods in the United States and Estimation of Normal Consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef]
- Iwashina, T. The Structure and Distribution of the Flavonoids in Plants. J. Plant Res. 2000, 113, 287–299. [Google Scholar] [CrossRef]
- Popa, D.-S.; Rusu, M.E. Isoflavones: Vegetable sources, biological activity, and analytical methods for their assessment. In Superfood and Functional Food—The Development of Superfoods and Their Roles as Medicine; InTech: London, UK, 2017; pp. 133–153. [Google Scholar]
- Ku, Y.-S.; Ng, M.-S.; Cheng, S.-S.; Lo, A.W.-Y.; Xiao, Z.; Shin, T.-S.; Chung, G.; Lam, H.-M. Understanding the Composition, Biosynthesis, Accumulation and Transport of Flavonoids in Crops for the Promotion of Crops as Healthy Sources of Flavonoids for Human Consumption. Nutrients 2020, 12, 1717. [Google Scholar] [CrossRef]
- Hosseinzade, A.; Sadeghi, O.; Biregani, A.N.; Soukhtehzari, S.; Brandt, G.S.; Esmaillzadeh, A. Immunomodulatory Effects of Flavonoids: Possible Induction of T CD4+ Regulatory Cells through Suppression of MTOR Pathway Signaling Activity. Front. Immunol. 2019, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Wang, Z.R.; Zheng, J.J.; Ding, J.Q.; Zhong, J.G.; Zhang, T.Y.; Li, W.; Zhang, M. Baicalein Improves Cognitive Deficits and Hippocampus Impairments in Temporal Lobe Epilepsy Rats. Brain Res. 2019, 1714, 111–118. [Google Scholar] [CrossRef]
- Fu, P.; Yuan, Q.; Sun, Y.; Wu, X.; Du, Z.; Li, Z.; Yu, J.; Lv, K.; Hu, J. Baicalein Ameliorates Epilepsy Symptoms in a Pilocarpine-Induced Rat Model by Regulation of IGF1R. Neurochem. Res. 2020, 45, 3021–3033. [Google Scholar] [CrossRef]
- Mao, X.; Cao, Y.; Li, X.; Yin, J.; Wang, Z.; Zhang, Y.; Mao, C.; Fan, K.; Zhou, H.; Cai, J.; et al. Baicalein Ameliorates Cognitive Deficits in Epilepsy-like Tremor Rat. Neurol. Sci. 2014, 35, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jia, Z.; Xiao, Z.; Zhao, J.; Lu, Y.; Chu, L.; Shao, H.; Pei, L.; Zhang, S.; Chen, Y. Baicalin Rescues Cognitive Dysfunction, Mitigates Neurodegeneration, and Exerts Anti-Epileptic Effects through Activating Tlr4/Myd88/Caspase-3 Pathway in Rats. Drug Des. Dev. Ther. 2021, 15, 3163–3180. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Lu, C.W.; Wang, S.J. Luteolin Protects the Hippocampus against Neuron Impairments Induced by Kainic Acid in Rats. Neurotoxicology 2016, 55, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Birman, H.; Dar, K.A.; Kapucu, A.; Acar, S.; Üzüm, G. Effects of Luteolin on Liver, Kidney and Brain in Pentylentetrazol-Induced Seizures: Involvement of Metalloproteinases and NOS Activities. Balkan Med. J. 2012, 2012, 188–196. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wang, S.J.; Huang, S.K. Protective Effect of Hispidulin on Kainic Acid-Induced Seizures and Neurotoxicity in Rats. Eur. J. Pharmacol. 2015, 755, 6–15. [Google Scholar] [CrossRef]
- Dang, J.; Paudel, Y.N.; Yang, X.; Ren, Q.; Zhang, S.; Ji, X.; Liu, K.; Jin, M. Schaftoside Suppresses Pentylenetetrazol-Induced Seizures in Zebrafish via Suppressing Apoptosis, Modulating Inflammation, and Oxidative Stress. ACS Chem. Neurosci. 2021, 12, 2542–2552. [Google Scholar] [CrossRef]
- Luo, W.D.; Min, J.W.; Huang, W.X.; Wang, X.; Peng, Y.Y.; Han, S.; Yin, J.; Liu, W.H.; He, X.H.; Peng, B.W. Vitexin Reduces Epilepsy after Hypoxic Ischemia in the Neonatal Brain via Inhibition of NKCC1. J. Neuroinflamm. 2018, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, J.; Wang, N.; Mishra, A.; Li, H.; Liu, H.; Fan, Y.; Liu, N.; Wu, Z. Wogonin Preventive Impact on Hippocampal Neurodegeneration, Inflammation and Cognitive Defects in Temporal Lobe Epilepsy. Saudi J. Biol. Sci. 2020, 27, 2149–2156. [Google Scholar] [CrossRef]
- Qi, H.; Liu, L. Rhoifolin Attenuates Damage to Hippocampal Neuronal Culture Model of Acquired Epilepsy in Vitro by Regulating NF-ΚB/INOS/COX-2 Axis. Qual. Assur. Saf. Crops Foods 2022, 14, 116–123. [Google Scholar] [CrossRef]
- Rong, S.; Wan, D.; Fan, Y.; Liu, S.; Sun, K.; Huo, J.; Zhang, P.; Li, X.; Xie, X.; Wang, F.; et al. Amentoflavone Affects Epileptogenesis and Exerts Neuroprotective Effects by Inhibiting NLRP3 Inflammasome. Front. Pharmacol. 2019, 10, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Sun, T.; Niu, J.G.; He, Z.Q.; Liu, Y.; Wang, F. Amentoflavone Protects Hippocampal Neurons: Anti-Inflammatory, Antioxidative, and Antiapoptotic Effects. Neural. Regen. Res. 2015, 10, 1125. [Google Scholar] [CrossRef]
- Mkhize, N.V.P.; Qulu, L.; Mabandla, M.V. The Effect of Quercetin on Pro- and Anti-Inflammatory Cytokines in a Prenatally Stressed Rat Model of Febrile Seizures. J. Exp. Neurosci. 2017, 11, 1179069517704668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Zheng, Z.; Fan, S.; Wen, X.; Han, X.; Wang, S.; Wang, Y.; Zhang, Z.; Shan, Q.; Li, M.; et al. Ameliorating Effect of Quercetin on Epilepsy by Inhibition of Inflammation in Glial Cells. Exp. Ther. Med. 2020, 20, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, M.; Ghasemi-Kasman, M.; Ghasemi, S.; Akbari, A.; Moalem-Banhangi, M.; Zare, L.; Ahmadian, S.R. Fabrication and Evaluation of Novel Quercetin-Conjugated Fe3O4–β-Cyclodextrin Nanoparticles for Potential Use in Epilepsy Disorder. Int. J. Nanomed. 2019, 14, 6481–6495. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wei, H.; Li, P.; Zhao, H.; Li, R.; Yang, F. Quercetin Administration Following Hypoxia-Induced Neonatal Brain Damage Attenuates Later-Life Seizure Susceptibility and Anxiety-Related Behavior: Modulating Inflammatory Response. Front. Pediatr. 2022, 10, 791815. [Google Scholar] [CrossRef]
- Ahmed, H.; Khan, M.A.; Ali Zaidi, S.A.; Muhammad, S. In Silico and In Vivo: Evaluating the Therapeutic Potential of Kaempferol, Quercetin, and Catechin to Treat Chronic Epilepsy in a Rat Model. Front. Bioeng. Biotechnol. 2021, 9, 754952. [Google Scholar] [CrossRef]
- Chang, A.; Chang, Y.; Wang, S.J. Rutin Prevents Seizures in Kainic Acid-Treated Rats: Evidence of Glutamate Levels, Inflammation and Neuronal Loss Modulation. Food Funct. 2022, 13, 10401–10414. [Google Scholar] [CrossRef]
- Khatoon, S.; Agarwal, N.B.; Samim, M.; Alam, O. Neuroprotective Effect of Fisetin Through Suppression of IL-1R/TLR Axis and Apoptosis in Pentylenetetrazole-Induced Kindling in Mice. Front. Neurol. 2021, 12, 689069. [Google Scholar] [CrossRef]
- Abd El-Aal, S.A.; El-Abhar, H.S.; Abulfadl, Y.S. Morin Offsets PTZ-Induced Neuronal Degeneration and Cognitive Decrements in Rats: The Modulation of TNF-α/TNFR-1/RIPK1,3/MLKL/PGAM5/Drp-1, IL-6/JAK2/STAT3/GFAP and Keap-1/Nrf-2/HO-1 Trajectories. Eur. J. Pharmacol. 2022, 931, 175213. [Google Scholar] [CrossRef]
- Lee, J.M.; Hong, J.; Moon, G.J.; Jung, U.J.; Won, S.-Y.; Kim, S.R. Morin Prevents Granule Cell Dispersion and Neurotoxicity via Suppression of MTORC1 in a Kainic Acid-Induced Seizure Model. Exp. Neurobiol. 2018, 27, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Meng, F.; Tu, L.; Sun, L. Myricetin Attenuates the Severity of Seizures and Neuroapoptosis in Pentylenetetrazole Kindled Mice by Regulating the of BDNF-TrkB Signaling Pathway and Modulating Matrix Metalloproteinase-9 and GABAA. Exp. Ther. Med. 2019, 17, 3083–3091. [Google Scholar] [CrossRef] [Green Version]
- Keikhaei, F.; Mirshekar, M.A.; Shahraki, M.R.; Dashipour, A. Antiepileptogenic Effect of Myricitrin on Spatial Memory and Learning in a Kainate-Induced Model of Temporal Lobe Epilepsy. Learn. Motiv. 2020, 69, 101610. [Google Scholar] [CrossRef]
- de Zorzi, V.N.; Haupenthal, F.; Cardoso, A.S.; Cassol, G.; Facundo, V.A.; Bálico, L.J.; Lima, D.K.S.; Santos, A.R.S.; Furian, A.F.; Oliveira, M.S.; et al. Galangin Prevents Increased Susceptibility to Pentylenetetrazol-Stimulated Seizures by Prostaglandin E2. Neuroscience 2019, 413, 154–168. [Google Scholar] [CrossRef]
- Golechha, M.; Sarangal, V.; Bhatia, J.; Chaudhry, U.; Saluja, D.; Arya, D.S. Naringin Ameliorates Pentylenetetrazol-Induced Seizures and Associated Oxidative Stress, Inflammation, and Cognitive Impairment in Rats: Possible Mechanisms of Neuroprotection. Epilepsy Behav. 2014, 41, 98–102. [Google Scholar] [CrossRef]
- Jeong, K.H.; Jung, U.J.; Kim, S.R. Naringin Attenuates Autophagic Stress and Neuroinflammation in Kainic Acid-Treated Hippocampus In Vivo. Evid.-Based Complement. Altern. Med. 2015, 2015, 354326. [Google Scholar] [CrossRef] [PubMed]
- Golechha, M.; Chaudhry, U.; Bhatia, J.; Saluja, D.; Arya, D.S. Naringin Protects against Kainic Acid-Induced Status Epilepticus in Rats: Evidence for an Antioxidant, Anti-Inflammatory and Neuroprotective Intervention. Biol. Pharm. Bull. 2011, 34, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Jeong, K.H.; Shin, W.-H.; Bae, Y.-S.; Jung, U.J.; Kim, S.R. Naringenin Ameliorates Kainic Acid-Induced Morphological Alterations in the Dentate Gyrus in a Mouse Model of Temporal Lobe Epilepsy. Neuroreport 2016, 27, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Jung, U.J.; Kim, D.W.; Kim, S.; Moon, G.J.; Hong, J.; Jeon, M.T.; Shin, M.; Chang, J.H.; Kim, S.R. Beneficial Effects of Hesperetin in a Mouse Model of Temporal Lobe Epilepsy. J. Med. Food 2018, 21, 1306–1309. [Google Scholar] [CrossRef]
- Atabaki, R.; Roohbakhsh, A.; Moghimi, A.; Mehri, S. Protective Effects of Maternal Administration of Curcumin and Hesperidin in the Rat Offspring Following Repeated Febrile Seizure: Role of Inflammation and TLR4. Int. Immunopharmacol. 2020, 86, 106720. [Google Scholar] [CrossRef]
- Sharma, P.; Kumari, S.; Sharma, J.; Purohit, R.; Singh, D. Hesperidin Interacts With CREB-BDNF Signaling Pathway to Suppress Pentylenetetrazole-Induced Convulsions in Zebrafish. Front. Pharmacol. 2021, 11, 607797. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, Y.S.; Yang, F.; Wu, B.; Yu, M.H.; Tu, M.Q.; Xu, H.B. Silibinin Inhibits Inflammation and Apoptosis in a Rat Model of Temporal Lobe Epilepsy. Int. J. Clin. Exp. Med. 2018, 11, 1891–1899. [Google Scholar]
- Kim, S.; Jung, U.J.; Oh, Y.-S.; Jeon, M.-T.; Kim, H.-J.; Shin, W.-H.; Hong, J.; Kim, S.R. Beneficial Effects of Silibinin Against Kainic Acidinduced Neurotoxicity in the Hippocampus in Vivo. Exp. Neurobiol. 2017, 26, 266–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.-P.; Feng, W.; Zhang, X.; Zhou, Y.-W. Genistein Protects Epilepsy-Induced Brain Injury Through Regulating The JAK2/STAT3 and Keap1/Nrf2 Signaling Pathways in The Developing Rats. Eur. J. Pharmacol. 2021, 912, 174620. [Google Scholar] [CrossRef]
- Qu, Z.; Jia, L.; Xie, T.; Zhen, J.; Si, P.; Cui, Z.; Xue, Y.; Sun, C.; Wang, W. (–)-Epigallocatechin-3-Gallate Protects Against Lithium-Pilocarpine-Induced Epilepsy by Inhibiting the Toll-Like Receptor 4 (TLR4)/Nuclear Factor-ΚB (NF-ΚB) Signaling Pathway. Med. Sci. Monit. 2019, 25, 1749–1758. [Google Scholar] [CrossRef]
- Cano, A.; Ettcheto, M.; Espina, M.; Auladell, C.; Calpena, A.C.; Folch, J.; Barenys, M.; Sánchez-López, E.; Camins, A.; García, M.L. Epigallocatechin-3-Gallate Loaded PEGylated-PLGA Nanoparticles: A New Anti-Seizure Strategy for Temporal Lobe Epilepsy. Nanomedicine 2018, 14, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, Q.Q.; Jia, J.N.; Sun, Q.Y.; Zhou, H.H.; Jin, W.L.; Mao, X.Y. Baicalein Exerts Neuroprotective Effects in FeCl3-Induced Posttraumatic Epileptic Seizures via Suppressing Ferroptosis. Front. Pharmacol. 2019, 10, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, K.; Patel, D.K. Medicinal Importance, Pharmacological Activities, and Analytical Aspects of Hispidulin: A Concise Report. J. Tradit. Complement. Med. 2017, 7, 360–366. [Google Scholar] [CrossRef]
- Nepal, M.; Choi, H.J.; Choi, B.-Y.; Yang, M.-S.; Chae, J.-I.; Li, L.; Soh, Y. Hispidulin Attenuates Bone Resorption and Osteoclastogenesis via the RANKL-Induced NF-ΚB and NFATc1 Pathways. Eur. J. Pharmacol. 2013, 715, 96–104. [Google Scholar] [CrossRef]
- Niu, X.; Chen, J.; Wang, P.; Zhou, H.; Li, S.; Zhang, M. The Effects of Hispidulin on Bupivacaine-Induced Neurotoxicity: Role of AMPK Signaling Pathway. Cell Biochem. Biophys. 2014, 70, 241–249. [Google Scholar] [CrossRef]
- Ashaq, A.; Maqbool, M.F.; Maryam, A.; Khan, M.; Shakir, H.A.; Irfan, M.; Qazi, J.I.; Li, Y.; Ma, T. Hispidulin: A Novel Natural Compound with Therapeutic Potential against Human Cancers. Phytother. Res. 2021, 35, 771–789. [Google Scholar] [CrossRef]
- Kavvadias, D.; Sand, P.; Youdim, K.A.; Qaiser, M.Z.; Rice-Evans, C.; Baur, R.; Sigel, E.; Rausch, W.-D.; Riederer, P.; Schreier, P. The Flavone Hispidulin, a Benzodiazepine Receptor Ligand with Positive Allosteric Properties, Traverses the Blood-Brain Barrier and Exhibits Anticonvulsive Effects. Br. J. Pharmacol. 2004, 142, 811–820. [Google Scholar] [CrossRef]
- de Melo, G.O.; Muzitano, M.F.; Legora-Machado, A.; Almeida, T.A.; de Oliveira, D.B.; Kaiser, C.R.; Koatz, V.L.G.; Costa, S.S. C-Glycosylflavones from the Aerial Parts of Eleusine Indica Inhibit LPS-Induced Mouse Lung Inflammation. Planta Med. 2005, 71, 362–363. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, G.; Song, M.; Wang, J.; Shen, C.; Chen, Z.; Huang, X.; Gao, Y.; Zhu, C.; Lin, C.; et al. Activation of Farnesoid X Receptor by Schaftoside Ameliorates Acetaminophen-Induced Hepatotoxicity by Modulating Oxidative Stress and Inflammation. Antioxid. Redox Signal. 2020, 33, 87–116. [Google Scholar] [CrossRef]
- He, M.; Min, J.-W.; Kong, W.-L.; He, X.-H.; Li, J.-X.; Peng, B.-W. A Review on the Pharmacological Effects of Vitexin and Isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.K.F.; Pereira, S.K.S.; Junior, R.D.S.S.; Santos, F.P.D.S.; Nascimento, A.D.S.; Feitosa, C.M.; de Sousa Figuerêdo, J.; do Nascimento Cavalcante, A.; da C. Araújo, E.C.; Rai, M. A Brief Review on the Neuroprotective Mechanisms of Vitexin. Biomed. Res. Int. 2018, 2018, 4785089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, D.L.; Ngau, T.H.; Nguyen, N.H.; Tran, G.B.; Nguyen, C.T. Potential Therapeutic and Pharmacological Effects of Wogonin: An Updated Review. Mol. Biol. Rep. 2020, 47, 9779–9789. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Salazar, L.A.; Shaheen, S.; Abdulmajid Ayatollahi, S.; Kobarfard, F.; Imran, M.; Imran, A.; Custódio, L.; Dolores López, M.; et al. The Therapeutic Potential of Wogonin Observed in Preclinical Studies. Evid.-Based Complement. Altern. Med. 2021, 2021, 9935451. [Google Scholar] [CrossRef]
- Refaat, J.; Yehia, S.Y.; Ramadan, A.; Kamel, M.S. Rhoifolin: A Review of Sources and Biological Activities. Int. J. Pharm. 2015, 2, 102–109. [Google Scholar]
- Al-Shalabi, E.; Abusulieh, S.; Hammad, A.M.; Sunoqrot, S. Rhoifolin Loaded in PLGA Nanoparticles Alleviates Oxidative Stress and Inflammation in Vitro and in Vivo. Biomater. Sci. 2022, 10, 5504–5519. [Google Scholar] [CrossRef]
- Peng, S.; Hu, C.; Liu, X.; Lei, L.; He, G.; Xiong, C.; Wu, W. Rhoifolin Regulates Oxidative Stress and Proinflammatory Cytokine Levels in Freund’s Adjuvant-Induced Rheumatoid Arthritis via Inhibition of NF-кB. Braz. J. Med. Biol. Res. 2020, 53, e9489. [Google Scholar] [CrossRef]
- Zheng, B.; Zheng, Y.; Zhang, N.; Zhang, Y.; Zheng, B. Rhoifolin from Plumula Nelumbinis Exhibits Anti-Cancer Effects in Pancreatic Cancer via AKT/JNK Signaling Pathways. Sci. Rep. 2022, 12, 5654. [Google Scholar] [CrossRef]
- Xiong, X.; Tang, N.; Lai, X.; Zhang, J.; Wen, W.; Li, X.; Li, A.; Wu, Y.; Liu, Z. Insights Into Amentoflavone: A Natural Multifunctional Biflavonoid. Front. Pharmacol. 2021, 12, 768708. [Google Scholar] [CrossRef] [PubMed]
- Saroni Arwa, P.; Zeraik, M.L.; Farias Ximenes, V.; da Fonseca, L.M.; da Silva Bolzani, V.; Siqueira Silva, D.H. Redox-Active Biflavonoids from Garcinia Brasiliensis as Inhibitors of Neutrophil Oxidative Burst and Human Erythrocyte Membrane Damage. J. Ethnopharmacol. 2015, 174, 410–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, H.M.; Almowallad, F.M.; Esmat, A.; Shehata, I.A.; Abdel-Sattar, E.A. Anti-Inflammatory Activity of Flavonoids from Chrozophora Tinctoria. Phytochem. Lett. 2015, 13, 74–80. [Google Scholar] [CrossRef]
- Chen, C.H.; Huang, Y.C.; Lee, Y.H.; Tan, Z.L.; Tsai, C.J.; Chuang, Y.C.; Tu, H.F.; Liu, T.C.; Hsu, F.T. Anticancer Efficacy and Mechanism of Amentoflavone for Sensitizing Oral Squamous Cell Carcinoma to Cisplatin. Anticancer Res. 2020, 40, 6723–6732. [Google Scholar] [CrossRef] [PubMed]
- Park, N.-H.; Lee, C.-W.; Bae, J.; Na, Y.J. Protective Effects of Amentoflavone on Lamin A-Dependent UVB-Induced Nuclear Aberration in Normal Human Fibroblasts. Bioorg. Med. Chem. Lett. 2011, 21, 6482–6484. [Google Scholar] [CrossRef]
- Zhao, N.; Sun, C.; Zheng, M.; Liu, S.; Shi, R. Amentoflavone Suppresses Amyloid Β1–42 Neurotoxicity in Alzheimer’s Disease through the Inhibition of Pyroptosis. Life Sci. 2019, 239, 117043. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; el Mahdy, N.M.; Klllç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Akyuz, E.; Paudel, Y.N.; Polat, A.K.; Dundar, H.E.; Angelopoulou, E. Enlightening the Neuroprotective Effect of Quercetin in Epilepsy: From Mechanism to Therapeutic Opportunities. Epilepsy Behav. 2021, 115, 107701. [Google Scholar] [CrossRef]
- Prakash, C.; Tyagi, J.; Rabidas, S.S.; Kumar, V.; Sharma, D. Therapeutic Potential of Quercetin and Its Derivatives in Epilepsy: Evidence from Preclinical Studies. Neuromolecular Med. 2022. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed]
- Diniz, T.C.; Silva, J.C.; Lima-Saraiva, S.R.G.D.; Ribeiro, F.P.R.D.A.; Pacheco, A.G.M.; de Freitas, R.M.; Quintans-Júnior, L.J.; Quintans, J.D.S.S.; Mendes, R.L.; Almeida, J.R.G.D.S. The Role of Flavonoids on Oxidative Stress in Epilepsy. Oxid. Med. Cell. Longev. 2015, 2015, 171756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A Dietary Antioxidant for Health Promotion. Antioxid. Redox Signal. 2013, 19, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Demchuk, O.M. New Perspectives for Fisetin. Front. Chem. 2019, 7, 697. [Google Scholar] [CrossRef]
- Das, J.; Singh, R.; Sharma, D. Antiepileptic Effect of Fisetin in Iron-Induced Experimental Model of Traumatic Epilepsy in Rats in the Light of Electrophysiological, Biochemical, and Behavioral Observations. Nutr. Neurosci. 2017, 20, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Raygude, K.S.; Kandhare, A.D.; Ghosh, P.; Bodhankar, S.L. Anticonvulsant Effect of Fisetin by Modulation of Endogenous Biomarkers. Biomed. Prev. Nutr. 2012, 2, 215–222. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Nie, Y.; Luo, Y.; Lin, F.; Zheng, Y.; Cheng, G.; Wu, H.; Zhang, K.; Su, W.; Shen, J.; et al. Protective Effects of Naringin against Paraquat-Induced Acute Lung Injury and Pulmonary Fibrosis in Mice. Food Chem. Toxicol. 2013, 58, 133–140. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef]
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent Progress Regarding Kaempferol for the Treatment of Various Diseases (Review). Exp. Ther. Med. 2019, 18, 2759–2776. [Google Scholar] [CrossRef] [Green Version]
- Silva dos Santos, J.; Gonçalves Cirino, J.P.; de Oliveira Carvalho, P.; Ortega, M.M. The Pharmacological Action of Kaempferol in Central Nervous System Diseases: A Review. Front. Pharmacol. 2021, 11, 565700. [Google Scholar] [CrossRef] [PubMed]
- Caselli, A.; Cirri, P.; Santi, A.; Paoli, P. Morin: A Promising Natural Drug. Curr. Med. Chem. 2016, 23, 774–791. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Wang, X.; Yan, H.-C. Morin Hydrate: A Comprehensive Review on Novel Natural Dietary Bioactive Compound with Versatile Biological and Pharmacological Potential. Biomed. Pharmacother. 2021, 138, 111511. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Lee, Y.; Chun, H.J.; Kim, A.H.; Kim, J.Y.; Lee, J.Y.; Ishigami, A.; Lee, J. Neuroprotective and Anti-Inflammatory Effects of Morin in a Murine Model of Parkinson’s Disease. J. Neurosci. Res. 2016, 94, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Shyma, R.L.; Mini, S. Neuroprotective Effect of Morin via TrkB/Akt Pathway against Diabetes Mediated Oxidative Stress and Apoptosis in Neuronal Cells. Toxicol. Mech. Methods 2022, 32, 695–704. [Google Scholar] [CrossRef]
- Kandhare, A.; Mukherjee, A.; Bodhankar, S. Anti-Epileptic Effect of Morin against Experimental Pentylenetetrazol-Induced Seizures via Modulating Brain Monoamines and Oxidative Stress. Asian Pac. J. Trop. Biomed. 2018, 8, 352. [Google Scholar] [CrossRef]
- Park, K.S.; Chong, Y.; Kim, M.K. Myricetin: Biological Activity Related to Human Health. Appl. Biol. Chem. 2016, 59, 259–269. [Google Scholar] [CrossRef]
- Semwal, D.; Semwal, R.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xiang, H.; Huang, C.; Lu, J. Pharmacological Actions of Myricetin in the Nervous System: A Comprehensive Review of Preclinical Studies in Animals and Cell Models. Front. Pharmacol. 2021, 12, 797298. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Qin, M.; Ye, J.; Pan, R.; Meng, X.; Wang, M.; Luo, Y.; Li, Z.; Wang, H.; Sun, X. Inhibitory Effects of Myricitrin on Oxidative Stress-Induced Endothelial Damage and Early Atherosclerosis in ApoE−/− Mice. Toxicol. Appl. Pharmacol. 2013, 271, 114–126. [Google Scholar] [CrossRef]
- Domitrović, R.; Rashed, K.; Cvijanović, O.; Vladimir-Knežević, S.; Škoda, M.; Višnić, A. Myricitrin Exhibits Antioxidant, Anti-Inflammatory and Antifibrotic Activity in Carbon Tetrachloride-Intoxicated Mice. Chem. Biol. Interact. 2015, 230, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Heo, M.Y.; Sohn, S.J.; Au, W.W. Anti-Genotoxicity of Galangin as a Cancer Chemopreventive Agent Candidate. Mutat. Res. Rev. Mutat. Res. 2001, 488, 135–150. [Google Scholar] [CrossRef]
- Aloud, A.A.; Chinnadurai, V.; Govindasamy, C.; Alsaif, M.A.; Al-Numair, K.S. Galangin, a Dietary Flavonoid, Ameliorates Hyperglycaemia and Lipid Abnormalities in Rats with Streptozotocin-Induced Hyperglycaemia. Pharm. Biol. 2018, 56, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Singh Tuli, H.; Sak, K.; Adhikary, S.; Kaur, G.; Aggarwal, D.; Kaur, J.; Kumar, M.; Chaturvedi Parashar, N.; Parashar, G.; Sharma, U.; et al. Galangin: A Metabolite That Suppresses Anti-Neoplastic Activities through Modulation of Oncogenic Targets. Exp. Biol. Med. 2022, 247, 345–359. [Google Scholar] [CrossRef]
- Lei, Y.; Chen, J.; Zhang, W.; Fu, W.; Wu, G.; Wei, H.; Wang, Q.; Ruan, J. In Vivo Investigation on the Potential of Galangin, Kaempferol and Myricetin for Protection of d-Galactose-Induced Cognitive Impairment. Food Chem. 2012, 135, 2702–2707. [Google Scholar] [CrossRef]
- Chen, Q.X.; Zhou, L.; Long, T.; Qin, D.L.; Wang, Y.L.; Ye, Y.; Zhou, X.G.; Wu, J.M.; Wu, A.G. Galangin Exhibits Neuroprotective Effects in 6-OHDA-Induced Models of Parkinson’s Disease via the Nrf2/Keap1 Pathway. Pharmaceuticals 2022, 15, 1014. [Google Scholar] [CrossRef]
- Rani, N.; Bharti, S.; Krishnamurthy, B.; Bhatia, J.; Sharma, C.; Amjad Kamal, M.; Ojha, S.; Singh Arya, D. Pharmacological Properties and Therapeutic Potential of Naringenin: A Citrus Flavonoid of Pharmaceutical Promise. Curr. Pharm. Des. 2016, 22, 4341–4359. [Google Scholar] [CrossRef] [PubMed]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of Their Molecular Mechanisms and Experimental Models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Cruz-Martins, N.; Butnariu, M.; Sarac, I.; Bagiu, I.-C.; Ezzat, S.M.; Wang, J.; Koay, A.; Sheridan, H.; Adetunji, C.O.; et al. Hesperetin’s Health Potential: Moving from Preclinical to Clinical Evidence and Bioavailability Issues, to Upcoming Strategies to Overcome Current Limitations. Crit. Rev. Food Sci. Nutr. 2022, 62, 4449–4464. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, S.; Ghasemi-Kasman, M.; Ebrahimpour, A.; Ahmadian, S.R.; Pouramir, M. Anticonvulsant Effects of Hesperetin in Animal Model of Pentylenetetrazole-Induced-Seizures. J. Babol. Univ. Med. Sci. 2018, 20, 19–26. [Google Scholar] [CrossRef]
- Hajialyani, M.; Hosein Farzaei, M.; Echeverría, J.; Nabavi, S.; Uriarte, E.; Sobarzo-Sánchez, E. Hesperidin as a Neuroprotective Agent: A Review of Animal and Clinical Evidence. Molecules 2019, 24, 648. [Google Scholar] [CrossRef] [PubMed]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Neuropharmacological Properties and Pharmacokinetics of the Citrus Flavonoids Hesperidin and Hesperetin—A Mini-Review. Life Sci. 2014, 113, 1–6. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Imran, M.; Rauf, A.; Nadeem, M.; Gondal, T.A.; Ahmad, B.; Atif, M.; Mubarak, M.S.; Sytar, O.; et al. Genistein: An Integrative Overview of Its Mode of Action, Pharmacological Properties, and Health Benefits. Oxid. Med. Cell. Longev. 2021, 2021, 3268136. [Google Scholar] [CrossRef]
- Noll, C.; Lameth, J.; Paul, J.-L.; Janel, N. Effect of Catechin/Epicatechin Dietary Intake on Endothelial Dysfunction Biomarkers and Proinflammatory Cytokines in Aorta of Hyperhomocysteinemic Mice. Eur. J. Nutr. 2013, 52, 1243–1250. [Google Scholar] [CrossRef]
- Vazquez Prieto, M.A.; Bettaieb, A.; Rodriguez Lanzi, C.; Soto, V.C.; Perdicaro, D.J.; Galmarini, C.R.; Haj, F.G.; Miatello, R.M.; Oteiza, P.I. Catechin and Quercetin Attenuate Adipose Inflammation in Fructose-Fed Rats and 3T3-L1 Adipocytes. Mol. Nutr. Food Res. 2015, 59, 622–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhail, M.; parveen, A.; Husain, A.; Rehan, M. Exploring Inhibitory Mechanisms of Green Tea Catechins as Inhibitors of a Cancer Therapeutic Target, Nuclear Factor-ΚB (NF-ΚB). Biosci. Biotechnol. Res. Asia 2019, 16, 715–723. [Google Scholar] [CrossRef]
- Goodlett, C.R.; Stringer, M.; LaCombe, J.; Patel, R.; Wallace, J.M.; Roper, R.J. Evaluation of the Therapeutic Potential of Epigallocatechin-3-Gallate (EGCG) via Oral Gavage in Young Adult Down Syndrome Mice. Sci. Rep. 2020, 10, 10426. [Google Scholar] [CrossRef]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.-D. Epigallocatechin-3-Gallate (EGCG): Chemical and Biomedical Perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef] [Green Version]
- Legeay, S.; Rodier, M.; Fillon, L.; Faure, S.; Clere, N. Epigallocatechin Gallate: A Review of Its Beneficial Properties to Prevent Metabolic Syndrome. Nutrients 2015, 7, 5443–5468. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Flavonoid | Model Used | Doses | Effects | References |
|---|---|---|---|---|
| Baicalein | Pilo-induced epilepsy in rats | 40 mg/kg, i.p. | Reduces pro-inflammatory cytokines levels (TNF-α and IL-1β) | [82] |
| Pilo-induced epilepsy in rats | 20, 40, and 80 mg/kg, orally | Downregulates the expression of cytokines (IL-1β, IL-6, and TNFα), and IGF1R | [83] | |
| Tremor rats | 10, 20, and 40 mg/kg, i.p. | Suppresses release of cytokines (TNF-α, IL-1β, IL-6, and IL-10), and p-JNK and p-p38 levels, while increasing pERK level | [84] | |
| PTZ-kindled rats | 50 and 100 mg/kg, orally | Decreases levels of IL-1β and IL-6 and inhibits TLR4/MYD88/Caspase-3 pathway | [85] | |
| Luteolin | KA-injected rats | 10 and 50 mg/kg, i.p. | Suppresses microglial activation in hippocampus | [86] |
| PTZ-induced seizures in rats | 10 mg/kg, i.p. | Reduces iNOS and MMP-2 activity, and increases eNOS activity | [87] | |
| Hispidulin | KA-induced seizures in rats | 10 and 50 mg/kg, i.p. | Suppresses microglialosis, pro-inflammatory cytokines production (IL-1β, IL-6, and TNF-α), c-Fos expression and MAPK activation | [88] |
| Schaftoside | PTZ-induced seizures in zebrafish | 100, 200, 400 μM | Decreases IL-1β, IL-6, NF-кB, and c-fos expression | [89] |
| Vitexin | Neonatal hypoxic ischemia-induced seizures in rats | 45 mg/kg, i.p. | Decreases neutrophil infiltration and IL-1β, IL-6, and TNF-α expression | [90] |
| Wogonin | KA-induced TLE in rats | 100 mg/kg, orally | Decreases IL-1β, TNF-α, and NF-kB expression | [91] |
| Rhoifolin | Hippocampal neuronal cell culture (HT-22 cell line) | 5, 10, and 20 µM | Reduces IL-1β, IL-6 and TNF-α levels, and inhibits NF-кB/iNOS/COX-2 pathway | [92] |
| Amentoflavone | PTZ-kindled mice | 25 mg/kg, orally | Decreases IL-1β, IL 18, and TNF-α expression, and inhibits NLRP3 inflammasome activation | [93] |
| Pilo-kindled mice | 25 mg/kg, orally | Decreases NO, PGE2, IL-1β and IL-6 production, inhibits NF-кB p65 activation | [94] | |
| Quercetin | Febrile seizures in prenatally stressed rats | 10 mg/kg, i.p. | Decreases levels of IL-1β, IL-6, and TNF-α | [95] |
| KA-induced seizures in mice | 100 mg/kg, i.p. | Reduces microglial activation, and levels of TNF-α, IL-1β, and activates NF-кB | [96] | |
| PTZ-kindled mice | 25 and 50 mg/kg, i.p. | Reduces astrocytes activation | [97] | |
| Neonatal hypoxic ischemia-induced seizures in rats | 25, 50, and 100 mg/kg, i.p. | Reduces IL-1β, IL-6, TNF-α, MCP-1 and iNOS levels and TLR4/NF-кB signaling in hippocampus | [98] | |
| PTZ-kindled rats | 100 mg/kg, orally | Suppresses TNF-α, IL-6, IL-1β and NF-kB expression, and increases IL1Ra, IL-4, and IL-10 expression | [99] | |
| Rutin | KA-kindled rats | 50 and 100 mg/kg, orally | Suppresses astrocytes activation, downregulates IL-1β, IL-6, TNF-α, HMGB1, IL-1R1, and TLR-4 expression, and upregulates IL-10 expression | [100] |
| Fisetin | PTZ-kindled mice | 5, 10, and 20 mg/kg, orally | Decreases HMGB1, TLR-4, IL-1R1, IL-1β, IL-6, and TNF-a levels, and NF-kB and COX-2 expression | [101] |
| Kaempferol | PTZ-kindled rats | 100 mg/kg, orally | Downregulates TNF-α, IL-6, IL-1β and NF-kB expression and upregulates IL1Ra, IL-4, and IL-10 expression | [99] |
| Morin | PTZ-kindled rats | 10 mg/kg, i.p. | Suppresses TNF-α expression, mitigates astrocyte activation, and IL-6/p-JAK-2/p-STAT3 signaling | [102] |
| KA-kindled mice | 20, 40, and 80 mg/kg, orally | Decreases microglial activation and IL-1β, TNF-α, and iNOS levels and inhibits mTORC1 pathway | [103] | |
| Myricetin | PTZ-kindled mice | 100 and 200 mg/kg, orally | Downregulate MMP-9 expression | [104] |
| Myricitrin | KA-induced TLE in rats | 5 mg/kg, i.p. | Decreases TNF-α concentration | [105] |
| Galangin | PTZ-kindled mice | 30 mg/kg, i.p. | Decreases microglial and astrocytic activation | [106] |
| Naringin | PTZ-kindled rats | 20, 40, and 80 mg/kg, i.p. | Reduces TNF-α levels | [107] |
| KA-kindled mice | 80 mg/kg, i.p. | Decreases TNF-α expression in activated microglial cells | [108] | |
| KA-induced status epilepticus in rats | 20, 40, and 80 mg/kg, i.p. | Decreases TNF-α expression | [109] | |
| Naringenin | KA-kindled mice | 50 and100 mg/kg, i.p. | Reduces IL-1β, and TNF-α levels in microglial cells and inhibits mTORC1 pathway | [110] |
| Hesperetin | KA-induced TLE in mice | 5, 10, and 20 mg/kg, orally | Reduces TNFα, IL-1β, and iNOS levels | [111] |
| Hesperidin | Febrile seizure in rat pups | 100 mg/kg, orally (Maternal administration) | Decreases TNF-α, IL 10, and TLR4 expression | [112] |
| PTZ-induced seizures in Zebrafish larvae | 1, 5, and 10 μM (Preincubated) | Reduces c-fos and IL-10 expression | [113] | |
| Silibinin | Lithium-Pilo-induced TLE in rats | 50 and 100 mg/kg, orally | Inhibits TNF-α, IL-1β, IL-6, and HIF-1α expression | [114] |
| KA-kindled mice | 50, 100, 200 mg/kg, i.p. | Inhibits TNF-α and IL-1β expressions and mTORC1 pathway | [115] | |
| Genistein | PTZ-kindled rats | 5 and 15 mg/kg, i.p. | Reduces astrocytes and microglial activation, and TNF-α, IL-1β, p-JAK2, p-STAT3 expression | [116] |
| Catechin | PTZ-kindled rats | 100 mg/kg, orally | Suppresses TNF-α, IL-6, and IL-1β levels, and NF-kB 4 expression, and upregulates IL1Ra, IL-4, and IL-10 expression | [99] |
| Epigallocatechin-3-gallate | Lithium-Pilo-induced TLE in rats | 25 mg/kg, i.p. | Decreases TLR4, NF-kB, and IL-1β expression | [117] |
| KA-induced kindling TLE in mice | 30 mg/kg, i.p. | Inhibits astrocytes and microglial activation | [118] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabidas, S.S.; Prakash, C.; Tyagi, J.; Suryavanshi, J.; Kumar, P.; Bhattacharya, J.; Sharma, D. A Comprehensive Review on Anti-Inflammatory Response of Flavonoids in Experimentally-Induced Epileptic Seizures. Brain Sci. 2023, 13, 102. https://doi.org/10.3390/brainsci13010102
Rabidas SS, Prakash C, Tyagi J, Suryavanshi J, Kumar P, Bhattacharya J, Sharma D. A Comprehensive Review on Anti-Inflammatory Response of Flavonoids in Experimentally-Induced Epileptic Seizures. Brain Sciences. 2023; 13(1):102. https://doi.org/10.3390/brainsci13010102
Chicago/Turabian StyleRabidas, Shyam Sunder, Chandra Prakash, Jyoti Tyagi, Jyoti Suryavanshi, Pavan Kumar, Jaydeep Bhattacharya, and Deepak Sharma. 2023. "A Comprehensive Review on Anti-Inflammatory Response of Flavonoids in Experimentally-Induced Epileptic Seizures" Brain Sciences 13, no. 1: 102. https://doi.org/10.3390/brainsci13010102