

The Impact of Sleep on Face Recognition Memory: A Scoping Review

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
1.1. On Sleep and Memory
1.2. On Sleep and Face Recognition: The Current Review
2. Method
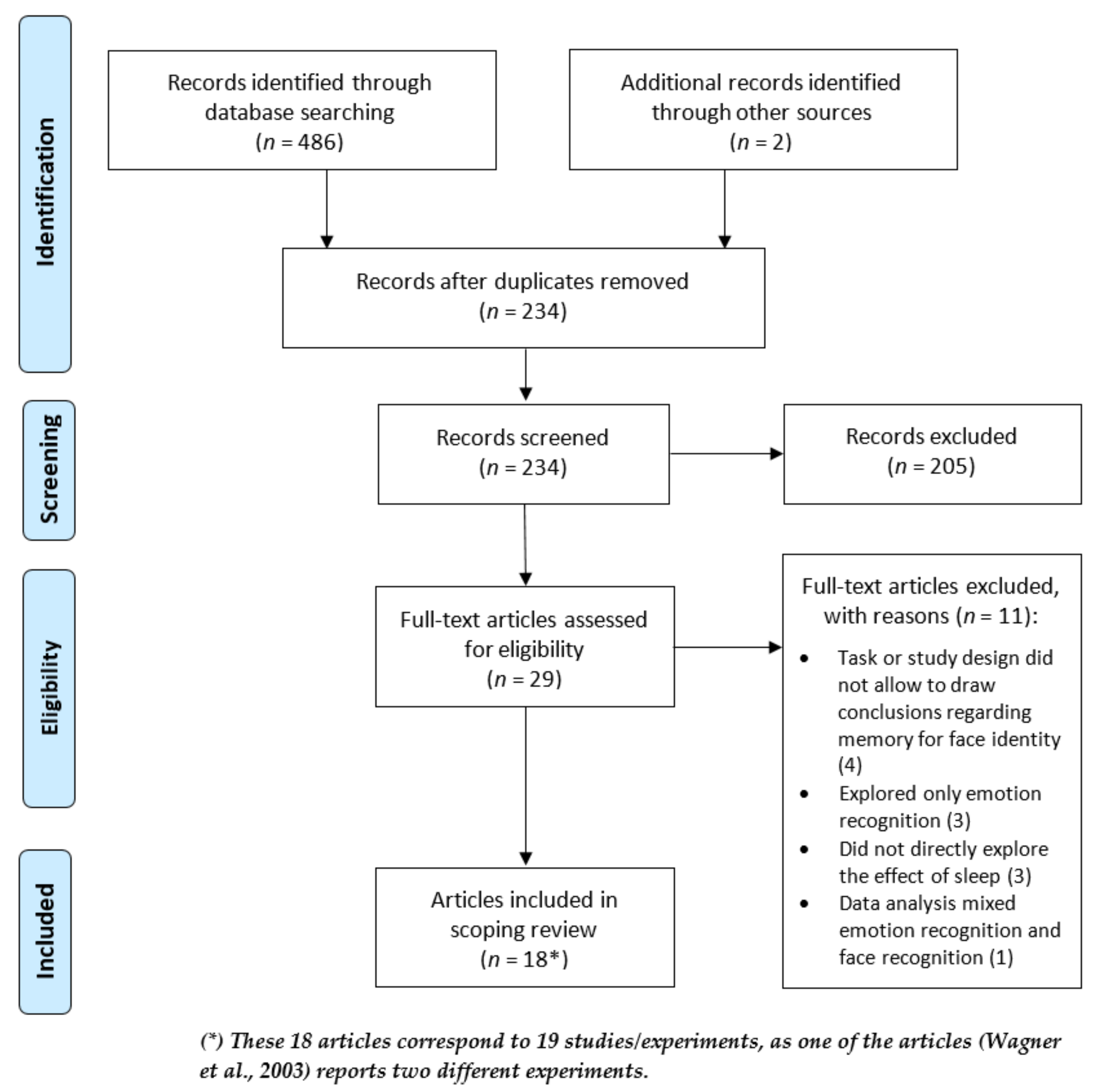
2.1. Search Strategy and Eligibility Criteria
2.2. Data Extraction and Synthesis Strategy
3. Results
3.1. Sample and Demographic Characteristics
3.2. Experimental Design and Sleep Manipulation/Evaluation
3.3. Face Recognition Task
3.4. Retention Interval and Time of Day of Learning and Testing
3.5. The Effects of Sleep on Face Recognition—Summary of Evidence
3.6. The Effects of Sleep on Face Recognition—Detailed Findings
3.6.1. Positive Effects, i.e., Beneficial Effects of Sleep on Performance
3.6.2. Negative Effects, i.e., Detrimental Effects of Sleep on Performance
3.6.3. Null Effects, i.e., Sleep Manipulation Has No Effect on Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mallon, L.; Broman, J.E.; Hetta, J. Sleep Complaints Predict Coronary Artery Disease Mortality in Males: A 12-Year Follow-up Study of a Middle-Aged Swedish Population. J. Intern. Med. 2002, 251, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.; Almeida, T.C.; Ramos, C.; Sousa, S.; Brito, J. Bidirectional Relationship between Perceived Stress and Insomnia Symptoms: The Role of Coping and Quality of Life. Sleep Biol. Rhythm. 2021, 19, 23–31. [Google Scholar] [CrossRef]
- Jarrin, D.C.; Alvaro, P.K.; Bouchard, M.-A.; Jarrin, S.D.; Drake, C.L.; Morin, C.M. Insomnia and Hypertension: A Systematic Review. Sleep Med. Rev. 2018, 41, 3–38. [Google Scholar] [CrossRef] [PubMed]
- Palagini, L.; Bruno, R.M.; Gemignani, A.; Baglioni, C.; Ghiadoni, L.; Riemann, D. Sleep Loss and Hypertension: A Systematic Review. Curr. Pharm. Des. 2013, 19, 2409–2419. [Google Scholar] [CrossRef]
- Anothaisintawee, T.; Reutrakul, S.; Van Cauter, E.; Thakkinstian, A. Sleep Disturbances Compared to Traditional Risk Factors for Diabetes Development: Systematic Review and Meta-Analysis. Sleep Med. Rev. 2016, 30, 11–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, Y.; Zhang, J.; Li, L.; Liu, X.; Wang, T.; Gao, Z. Association between Insomnia and Type 2 Diabetes Mellitus in Han Chinese Individuals in Shandong Province, China. Sleep Breath. 2019, 23, 349–354. [Google Scholar] [CrossRef]
- Irwin, M.R. Why Sleep Is Important for Health: A Psychoneuroimmunology Perspective. Annu. Rev. Psychol. 2015, 66, 143–172. [Google Scholar] [CrossRef] [Green Version]
- Vgontzas, A.N.; Fernandez-Mendoza, J.; Liao, D.; Bixler, E.O. Insomnia with Objective Short Sleep Duration: The Most Biologically Severe Phenotype of the Disorder. Sleep Med. Rev. 2013, 17, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Buysse, D.J. Insomnia, Depression and Aging. Assessing Sleep and Mood Interactions in Older Adults. Geriatrics 2004, 59, 47–51. [Google Scholar]
- Shaffery, J.; Hoffmann, R.; Armitage, R. The Neurobiology of Depression: Perspectives from Animal and Human Sleep Studies. Neuroscientist 2003, 9, 82–98. [Google Scholar] [CrossRef]
- Dew, M.A.; Hoch, C.C.; Buysse, D.J.; Monk, T.H.; Begley, A.E.; Houck, P.R.; Hall, M.; Kupfer, D.J.; Reynolds, C.F. Healthy Older Adults’ Sleep Predicts All-Cause Mortality at 4 to 19 Years of Follow-Up. Psychosom. Med. 2003, 65, 63–73. [Google Scholar] [CrossRef]
- Kripke, D.F.; Garfinkel, L.; Wingard, D.L.; Klauber, M.R.; Marler, M.R. Mortality Associated with Sleep Duration and Insomnia. Arch. Gen. Psychiatry 2002, 59, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-J.; Dimsdale, J.E. The Effect of Psychosocial Stress on Sleep: A Review of Polysomnographic Evidence. Behav. Sleep. Med. 2007, 5, 256–278. [Google Scholar] [CrossRef] [Green Version]
- Gilley, R.R. The Role of Sleep in Cognitive Function: The Value of a Good Night’s Rest. Clin. EEG Neurosci. 2022, 15500594221090067. [Google Scholar] [CrossRef]
- Nieto, M.; Motos, B.; Navarro, B.; Jimeno, M.V.; Fernández-Aguilar, L.; Ros, L.; Ricarte, J.J.; Latorre, J.M. Relation between Nighttime Sleep Duration and Executive Functioning in a Nonclinical Sample of Preschool Children. Scand. J. Psychol. 2022, 63, 191–198. [Google Scholar] [CrossRef]
- De Bruin, E.J.; van Run, C.; Staaks, J.; Meijer, A.M. Effects of Sleep Manipulation on Cognitive Functioning of Adolescents: A Systematic Review. Sleep Med. Rev. 2017, 32, 45–57. [Google Scholar] [CrossRef]
- Cruz, T.; García, L.; Álvarez, M.A.; Manzanero, A.L. Sleep Quality and Memory Function in Healthy Ageing. Neurol. Engl. Ed. 2022, 37, 31–37. [Google Scholar] [CrossRef]
- Ferrie, J.E.; Shipley, M.J.; Akbaraly, T.N.; Marmot, M.G.; Kivimäki, M.; Singh-Manoux, A. Change in Sleep Duration and Cognitive Function: Findings from the Whitehall II Study. Sleep 2011, 34, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Kronholm, E.; Sallinen, M.; Suutama, T.; Sulkava, R.; Era, P.; Partonen, T. Self-Reported Sleep Duration and Cognitive Functioning in the General Population. J. Sleep Res. 2009, 18, 436–446. [Google Scholar] [CrossRef]
- Lin, L.-H.; Xu, W.-Q.; Wang, S.-B.; Hu, Q.; Zhang, P.; Huang, J.-H.; Ke, Y.-F.; Ding, K.-R.; Hou, C.-L.; Jia, F.-J. U-Shaped Association between Sleep Duration and Subjective Cognitive Complaints in Chinese Elderly: A Cross-Sectional Study. BMC Psychiatry 2022, 22, 147. [Google Scholar] [CrossRef]
- Mellow, M.L.; Crozier, A.J.; Dumuid, D.; Wade, A.T.; Goldsworthy, M.R.; Dorrian, J.; Smith, A.E. How Are Combinations of Physical Activity, Sedentary Behaviour and Sleep Related to Cognitive Function in Older Adults? A Systematic Review. Exp. Gerontol. 2022, 159, 111698. [Google Scholar] [CrossRef]
- Richards, A.; Inslicht, S.S.; Metzler, T.J.; Mohlenhoff, B.S.; Rao, M.N.; O’Donovan, A.; Neylan, T.C. Sleep and Cognitive Performance from Teens to Old Age: More Is Not Better. Sleep 2017, 40, zsw029. [Google Scholar] [CrossRef]
- Burke, T.M.; Scheer, F.A.J.L.; Ronda, J.M.; Czeisler, C.A.; Wright, K.P., Jr. Sleep Inertia, Sleep Homeostatic and Circadian Influences on Higher-Order Cognitive Functions. J. Sleep Res. 2015, 24, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, J.P.K.; DeVito, A.; Calamia, M. Subjectively and Objectively Measured Sleep Predict Differing Aspects of Cognitive Functioning in Adults. Arch. Clin. Neuropsychol. 2019, 34, 1127–1137. [Google Scholar] [CrossRef]
- Roth, T.; Costa e Silva, J.A.; Chase, M.H. Sleep and Cognitive (Memory) Function: Research and Clinical Perspectives. Sleep Med. 2001, 2, 379–387. [Google Scholar] [CrossRef]
- Goldstein, A.N.; Walker, M.P. The Role of Sleep in Emotional Brain Function. Annu. Rev. Clin. Psychol. 2014, 10, 679–708. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.; Platten, C.R. Sleep Deprivation Lowers Inhibition and Enhances Impulsivity to Negative Stimuli. Behav. Brain Res. 2011, 217, 463–466. [Google Scholar] [CrossRef]
- Motomura, Y.; Kitamura, S.; Oba, K.; Terasawa, Y.; Enomoto, M.; Katayose, Y.; Hida, A.; Moriguchi, Y.; Higuchi, S.; Mishima, K. Sleep Debt Elicits Negative Emotional Reaction through Diminished Amygdala-Anterior Cingulate Functional Connectivity. PLoS ONE 2013, 8, e56578. [Google Scholar] [CrossRef]
- Prather, A.A.; Bogdan, R.; Ahmad, R.; Hariri, P. Impact of Sleep Quality on Amygdala Reactivity, Negative Affect, and Perceived Stress. Psychosom. Med. 2013, 75, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.-S.; Gujar, N.; Hu, P.; Jolesz, F.A.; Walker, M.P. The Human Emotional Brain without Sleep--a Prefrontal Amygdala Disconnect. Curr. Biol. CB 2007, 17, R877–R878. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, J.F.A.; Popp, R.; Haas, J.; Zulley, J.; Geisler, P.; Alpers, G.W.; Osterheider, M.; Eisenbarth, H. Shortened Night Sleep Impairs Facial Responsiveness to Emotional Stimuli. Biol. Psychol. 2013, 93, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Van der Helm, E.; Gujar, N.; Walker, M.P. Sleep Deprivation Impairs the Accurate Recognition of Human Emotions. Sleep 2010, 33, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Guadagni, V.; Burles, F.; Ferrara, M.; Iaria, G. The Effects of Sleep Deprivation on Emotional Empathy. J. Sleep Res. 2014, 23, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.P. The Role of Sleep in Cognition and Emotion. Ann. N. Y. Acad. Sci. 2009, 1156, 168–197. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.G.; Dallenbach, K.M. Obliviscence during Sleep and Waking. Am. J. Psychol. 1924, 35, 605–612. [Google Scholar] [CrossRef]
- Menz, M.M.; Rihm, J.S.; Salari, N.; Born, J.; Kalisch, R.; Pape, H.C.; Marshall, L.; Büchel, C. The Role of Sleep and Sleep Deprivation in Consolidating Fear Memories. NeuroImage 2013, 75, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Seeck-Hirschner, M.; Baier, P.C.; Weinhold, S.L.; Dittmar, M.; Heiermann, S.; Aldenhoff, J.B.; Göder, R. Declarative Memory Performance Is Associated with the Number of Sleep Spindles in Elderly Women. Am. J. Geriatr. Psychiatry 2012, 20, 782–788. [Google Scholar] [CrossRef]
- Rauchs, G.; Bertran, F.; Guillery-Girard, B.; Desgranges, B.; Kerrouche, N.; Denise, P.; Foret, J.; Eustache, F. Consolidation of Strictly Episodic Memories Mainly Requires Rapid Eye Movement Sleep. Sleep 2004, 27, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Diekelmann, S.; Wilhelm, I.; Born, J. The Whats and Whens of Sleep-Dependent Memory Consolidation. Sleep Med. Rev. 2009, 13, 309–321. [Google Scholar] [CrossRef]
- Ashworth, A.; Hill, C.M.; Karmiloff-Smith, A.; Dimitriou, D. Sleep Enhances Memory Consolidation in Children. J. Sleep Res. 2014, 23, 304–310. [Google Scholar] [CrossRef] [Green Version]
- Hoedlmoser, K. Sleep and Memory in Children. Curr. Sleep Med. Rep. 2020, 6, 280–289. [Google Scholar] [CrossRef]
- Palmer, C.A.; Alfano, C.A. Sleep and Emotion Regulation: An Organizing, Integrative Review. Sleep Med. Rev. 2017, 31, 6–16. [Google Scholar] [CrossRef]
- Tempesta, D.; De Gennaro, L.; Natale, V.; Ferrara, M. Emotional Memory Processing Is Influenced by Sleep Quality. Sleep Med. 2015, 16, 862–870. [Google Scholar] [CrossRef]
- Walker, M.P.; Stickgold, R. Sleep, Memory, and Plasticity. Annu. Rev. Psychol. 2006, 57, 139–166. [Google Scholar] [CrossRef] [Green Version]
- Ellenbogen, J.M.; Hu, P.T.; Payne, J.D.; Titone, D.; Walker, M.P. Human Relational Memory Requires Time and Sleep. Proc. Natl. Acad. Sci. USA 2007, 104, 7723–7728. [Google Scholar] [CrossRef] [Green Version]
- Wagner, U.; Gais, S.; Haider, H.; Verleger, R.; Born, J. Sleep Inspires Insight. Nature 2004, 427, 352–355. [Google Scholar] [CrossRef]
- Fortier-Brochu, É.; Beaulieu-Bonneau, S.; Ivers, H.; Morin, C.M. Insomnia and Daytime Cognitive Performance: A Meta-Analysis. Sleep Med. Rev. 2012, 16, 83–94. [Google Scholar] [CrossRef]
- Ackermann, S.; Rasch, B. Differential Effects of Non-REM and REM Sleep on Memory Consolidation? Curr. Neurol. Neurosci. Rep. 2014, 14, 430. [Google Scholar] [CrossRef]
- Smith, C. Sleep States and Memory Processes in Humans: Procedural versus Declarative Memory Systems. Sleep Med. Rev. 2001, 5, 491–506. [Google Scholar] [CrossRef] [Green Version]
- Boyce, R.; Williams, S.; Adamantidis, A. REM Sleep and Memory. Curr. Opin. Neurobiol. 2017, 44, 167–177. [Google Scholar] [CrossRef]
- Siegel, J.M. The REM Sleep-Memory Consolidation Hypothesis. Science 2001, 294, 1058–1063. [Google Scholar] [CrossRef]
- Vertes, R.P.; Eastman, K.E. The Case against Memory Consolidation in REM Sleep. Behav. Brain Sci. 2000, 23, 867–876. [Google Scholar] [CrossRef]
- Smith, C.; Macneill, C. Impaired Motor Memory for a Pursuit Rotor Task Following Stage 2 Sleep Loss in College Students. J. Sleep Res. 1994, 3, 206–213. [Google Scholar] [CrossRef]
- Wagner, U.; Gais, S.; Born, J. Emotional Memory Formation Is Enhanced across Sleep Intervals with High Amounts of Rapid Eye Movement Sleep. Learn. Mem. 2001, 8, 112–119. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, K.J.; Cote, K.A. Contributions of Post-Learning REM and NREM Sleep to Memory Retrieval. Sleep Med. Rev. 2021, 59, 101453. [Google Scholar] [CrossRef]
- White, D.; Kemp, R.I.; Jenkins, R.; Matheson, M.; Burton, A.M. Passport Officers’ Errors in Face Matching. PLoS ONE 2014, 9, e103510. [Google Scholar] [CrossRef]
- Bruce, V.; Young, A. Understanding Face Recognition. Br. J. Psychol. Lond. Engl. 1953 1986, 77 Pt 3, 305–327. [Google Scholar] [CrossRef]
- Haxby, J.V.; Hoffman, E.A.; Gobbini, M.I. The Distributed Human Neural System for Face Perception. Trends Cogn. Sci. 2000, 4, 223–233. [Google Scholar] [CrossRef]
- Li, J.; He, D.; Zhou, L.; Zhao, X.; Zhao, T.; Zhang, W.; He, X. The Effects of Facial Attractiveness and Familiarity on Facial Expression Recognition. Front. Psychol. 2019, 10, 2496. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Macedo, A.F.; Albuquerque, P.B.; Arantes, J. Always on My Mind? Recognition of Attractive Faces May Not Depend on Attention. Front. Psychol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulmann, S.; Pell, M.D. Facial Expression Decoding as a Function of Emotional Meaning Status: ERP Evidence. Neuroreport 2009, 20, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.M.; Iglesias, J.; Olivares, E.I.; Young, A.W. Differential Effects of Object-Based Attention on Evoked Potentials to Fearful and Disgusted Faces. Neuropsychologia 2008, 46, 1468–1479. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.M.; Young, A.W. Inferring Social Attributes from Different Face Regions: Evidence for Holistic Processing. Q. J. Exp. Psychol. 2011, 64, 751–766. [Google Scholar] [CrossRef]
- Sutherland, C.A.M.; Oldmeadow, J.A.; Santos, I.M.; Towler, J.; Michael Burt, D.; Young, A.W. Social Inferences from Faces: Ambient Images Generate a Three-Dimensional Model. Cognition 2013, 127, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Wilmer, J.B.; Germine, L.; Chabris, C.F.; Chatterjee, G.; Williams, M.; Loken, E.; Nakayama, K.; Duchaine, B. Human Face Recognition Ability Is Specific and Highly Heritable. Proc. Natl. Acad. Sci. USA 2010, 107, 5238–5241. [Google Scholar] [CrossRef] [Green Version]
- Kanwisher, N.; McDermott, J.; Chun, M.M. The Fusiform Face Area: A Module in Human Extrastriate Cortex Specialized for Face Perception. J. Neurosci. 1997, 17, 4302–4311. [Google Scholar] [CrossRef] [Green Version]
- Gosling, A.; Eimer, M. An Event-Related Brain Potential Study of Explicit Face Recognition. Neuropsychologia 2011, 49, 2736–2745. [Google Scholar] [CrossRef]
- Avidan, G.; Behrmann, M. Implicit Familiarity Processing in Congenital Prosopagnosia. J. Neuropsychol. 2008, 2, 141–164. [Google Scholar] [CrossRef] [Green Version]
- Diekelmann, S.; Born, J. The Memory Function of Sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef]
- Stickgold, R. Sleep-Dependent Memory Consolidation. Nature 2005, 437, 1272–1278. [Google Scholar] [CrossRef] [Green Version]
- Beattie, L. How Does Sleep Affect the Perception of Facial Emotion? Sleep 2018, 41, zsy030. [Google Scholar] [CrossRef]
- Beattie, L.; Walsh, D.; McLaren, J.; Biello, S.M.; White, D. Perceptual Impairment in Face Identification with Poor Sleep. R. Soc. Open Sci. 2016, 3, 160321. [Google Scholar] [CrossRef] [Green Version]
- Crönlein, T.; Langguth, B.; Eichhammer, P.; Busch, V. Impaired Recognition of Facially Expressed Emotions in Different Groups of Patients with Sleep Disorders. PLoS ONE 2016, 11, e0152754. [Google Scholar] [CrossRef] [Green Version]
- Holding, B.C.; Sundelin, T.; Cairns, P.; Perrett, D.I.; Axelsson, J. The Effect of Sleep Deprivation on Objective and Subjective Measures of Facial Appearance. J. Sleep Res. 2019, 28, e12860. [Google Scholar] [CrossRef]
- De Almondes, K.M.; Júnior, F.W.N.H.; Leonardo, M.E.M.; Alves, N.T. Facial Emotion Recognition and Executive Functions in Insomnia Disorder: An Exploratory Study. Front. Psychol. 2020, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chan, A.B.; Lau, E.Y.Y.; Hsiao, J.H. Individuals with Insomnia Misrecognize Angry Faces as Fearful Faces While Missing the Eyes: An Eye-Tracking Study. Sleep 2019, 42, zsy220. [Google Scholar] [CrossRef] [Green Version]
- Mograss, M.; Godbout, R.; Guillem, F. The ERP Old-New Effect: A Useful Indicator in Studying the Effects of Sleep on Memory Retrieval Processes. Sleep 2006, 29, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Solomonova, E.; Stenstrom, P.; Schon, E.; Duquette, A.; Dubé, S.; O’Reilly, C.; Nielsen, T. Sleep-Dependent Consolidation of Face Recognition and Its Relationship to REM Sleep Duration, REM Density and Stage 2 Sleep Spindles. J. Sleep Res. 2017, 26, 318–321. [Google Scholar] [CrossRef] [Green Version]
- Wagner, U.; Kashyap, N.; Diekelmann, S.; Born, J. The Impact of Post-Learning Sleep vs. Wakefulness on Recognition Memory for Faces with Different Facial Expressions. Neurobiol. Learn. Mem. 2007, 87, 679–687. [Google Scholar] [CrossRef]
- Chee, M.W.L.; Tan, J.C.; Parimal, S.; Zagorodnov, V. Sleep Deprivation and Its Effects on Object-Selective Attention. NeuroImage 2010, 49, 1903–1910. [Google Scholar] [CrossRef]
- Mograss, M.A.; Guillem, F.; Stickgold, R. Individual Differences in Face Recognition Memory: Comparison among Habitual Short, Average, and Long Sleepers. Behav. Brain Res. 2010, 208, 576–583. [Google Scholar] [CrossRef]
- Peters, M.D.J.; Marnie, C.; Tricco, A.C.; Pollock, D.; Munn, Z.; Alexander, L.; McInerney, P.; Godfrey, C.M.; Khalil, H. Updated Methodological Guidance for the Conduct of Scoping Reviews. JBI Evid. Synth. 2020, 18, 2119–2126. [Google Scholar] [CrossRef]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—A Web and Mobile App for Systematic Reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, U.; Hallschmid, M.; Verleger, R.; Born, J. Signs of REM Sleep Dependent Enhancement of Implicit Face Memory: A Repetition Priming Study. Biol. Psychol. 2003, 62, 197–210. [Google Scholar] [CrossRef]
- Barton, S.; Pietrowsky, R. Selective Enhancement of Processing of Negative Faces During Slow-Wave Sleep. Sleep Hypn. 2019, 21, 328–338. [Google Scholar] [CrossRef]
- Frings, D. The Effects of Low Levels of Fatigue on Face Recognition among Individuals and Team Members. J. Appl. Soc. Psychol. 2015, 45, 461–470. [Google Scholar] [CrossRef]
- Harrison, Y.; Horne, J.A. Sleep Loss and Temporal Memory. Q. J. Exp. Psychol. 2000, 53, 271–279. [Google Scholar] [CrossRef]
- Hussain, Z.; Sekuler, A.B.; Bennett, P.J. Robust Perceptual Learning of Faces in the Absence of Sleep. Vision Res. 2008, 48, 2785–2792. [Google Scholar] [CrossRef] [Green Version]
- Martella, D.; Plaza, V.; Estévez, A.F.; Castillo, A.; Fuentes, L.J. Minimizing Sleep Deprivation Effects in Healthy Adults by Differential Outcomes. Acta Psychol. 2012, 139, 391–396. [Google Scholar] [CrossRef]
- Maurer, L.; Zitting, K.-M.; Elliott, K.; Czeisler, C.A.; Ronda, J.M.; Duffy, J.F. A New Face of Sleep: The Impact of Post-Learning Sleep on Recognition Memory for Face-Name Associations. Neurobiol. Learn. Mem. 2015, 126, 31–38. [Google Scholar] [CrossRef]
- Mograss, M.A.; Guillem, F.; Godbout, R. Event-Related Potentials Differentiates the Processes Involved in the Effects of Sleep on Recognition Memory. Psychophysiology 2008, 45, 420–434. [Google Scholar] [CrossRef]
- Mograss, M.A.; Guillem, F.; Brazzini-Poisson, V.; Godbout, R. The Effects of Total Sleep Deprivation on Recognition Memory Processes: A Study of Event-Related Potential. Neurobiol. Learn. Mem. 2009, 91, 343–352. [Google Scholar] [CrossRef]
- Alberca-Reina, E.; Cantero, J.L.; Atienza, M. Semantic Congruence Reverses Effects of Sleep Restriction on Associative Encoding. Neurobiol. Learn. Mem. 2014, 110, 27–34. [Google Scholar] [CrossRef]
- Alberca-Reina, E.; Cantero, J.L.; Atienza, M. Impact of Sleep Loss before Learning on Cortical Dynamics during Memory Retrieval. NeuroImage 2015, 123, 51–62. [Google Scholar] [CrossRef]
- Sheth, B.R.; Nguyen, N.; Janvelyan, D. Does Sleep Really Influence Face Recognition Memory? PLoS ONE 2009, 4, e5496. [Google Scholar] [CrossRef] [Green Version]
- Stare, C.J.; Gruber, M.J.; Nadel, L.; Ranganath, C.; Gómez, R.L. Curiosity-Driven Memory Enhancement Persists over Time but Does Not Benefit from Post-Learning Sleep. Cogn. Neurosci. 2018, 9, 100–115. [Google Scholar] [CrossRef]
- Monteiro, F.; Rodrigues, P.; Nascimento, C.S.; Simões, F.; Miguel, M. The Daily Rhythms of Working Memory and Their Methodological Constraints: A Critical Overview. Biol. Rhythm Res. 2021, 53, 1116–1143. [Google Scholar] [CrossRef]
- Valdez, P.; Ramírez, C.; García, A.; Talamantes, J.; Armijo, P.; Borrani, J. Circadian Rhythms in Components of Attention. Biol. Rhythm Res. 2005, 36, 57–65. [Google Scholar] [CrossRef]
- Walker, M.P. Cognitive Consequences of Sleep and Sleep Loss. Sleep Med. 2008, 9, S29–S34. [Google Scholar] [CrossRef]
- Pizinger, T.M.; Aggarwal, B.; St-Onge, M.-P. Sleep Extension in Short Sleepers: An Evaluation of Feasibility and Effectiveness for Weight Management and Cardiometabolic Disease Prevention. Front. Endocrinol. 2018, 9, 392. [Google Scholar] [CrossRef]
- Schmidt, C.; Collette, F.; Cajochen, C.; Peigneux, P. A Time to Think: Circadian Rhythms in Human Cognition. Cogn. Neuropsychol. 2007, 24, 755–789. [Google Scholar] [CrossRef]
- Wilmer, J.B. Individual Differences in Face Recognition: A Decade of Discovery. Curr. Dir. Psychol. Sci. 2017, 26, 225–230. [Google Scholar] [CrossRef]


{kind=link}
| First Author | Year | Sample Size | Age Mean (SD) [and/or Range] | Design (for the Sleep Manipulation) | Face Memory Task | Training/Encoding ToD | Testing/Recall ToD | Retention Interval | Prior Sleep Controlled? How? | Sleep Manipulation Control | Sleep Length | VDs | Main Results | Direction of Effect and Conclusion |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POSITIVE EFFECTS, i.e., Beneficial Effects of Sleep on Performance | ||||||||||||||
| Barton and Pietrowsky [86] | 2019 | 40 [22F + 18M] sleep group (SG) = 20 [8F + 12M]; wake group (WG) = 20 [14F + 6M] | SG: 25.05 (6.88), WG: 27.05 (6.82) | Between-subjects; 2 groups: SG (1 h nap during retention) vs. WG (awake during retention) | Old-new recognition task, with emotion faces; explicit memorization instructions at encoding | 12 pm or early afternoon | Afternoon (1–4 p.m.) | 1 h | Sleep diaries (day before), abstain from substance intake | Polysomnography (lab) | No sleep past 8 a.m. the night before; SG: 1 h nap during retention WG: no nap during retention | Memory accuracy [Pr = hit rate-false alarm rate]; response bias [Br = false alarm rate/(1 − Pr)]; RTs | No significant main effects of group, or interactions with group, for memory accuracy, response bias and RT; positive correlation between N3 sleep and negative face recognition | No direct effect of sleep on face memory; correlation with N3 sleep suggests more elaborate processing during SWS and subsequently better face recognition for negative faces |
| Frings [87] | 2015 | 182 (36F + 146M) Alert = 103, Fatigued = 79 | [18,19,20,21,22,23,24] | Between-subjects; 2 groups: alert vs. fatigued (subdivided in individual participants and teams, but not relevant for review purposes) | Target detection task: two faces were initially shown and had to be subsequently identified among several sequentially presented quartets of faces, either target-present or target-absent | Alert: 7 p.m.; fatigued: 10–11 a.m. | Same as encoding | Immediate | No, but participants were instructed to be well rested before | Observational (field); Manipulation check with the Piper Fatigue Scale (confirmed) | Alert: prior sleep not controlled; fatigued: sleep duration <5 h for 2 consecutive nights, and systematically disturbed sleep | Hits, false alarms, sensitivity (A′), and response bias (B″) | Fatigued individuals had a higher response bias (i.e., a higher tendency towards identifying a target as absent) than alert individuals | No differences in detection ability. Sleep disruption interfered with face recognition, through response bias: sleep-disrupted individuals were more biased towards identifying the target as being absent (i.e., missing its recognition) than alert individuals. |
| Harrison and Horne [88] | 2000 | 40 (20F + 20M) | 23.4 [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] | Between-subjects; 4 groups: non-sleep-deprived (SD) with placed, non-SD with caffeine, SD with placebo, SD with caffeine. Total sleep deprivation for 35 h. | Old-new recognition task; two sequential sets of faces presented with a 5 min filler task between them; testing after a second 5 min. filler task. Assessed recognition (old-new?) and temporal memory (face belonged to 1st or 2nd set?). | Between 7:30 p.m. and 8:30 p.m. | Same as encoding | Almost immediate: 5 min between the two sets of 12 faces + 5 min after the second set, before testing | Actimetry and sleep diaries (3 nights before), controlled substance intake, normal sleepiness range and sleep habits (self-reported) | Non-SD groups: Actigraphy (home) SD groups: Observational (lab) | Non-SD groups: regular sleep duration; SD groups: no sleep | Recognition accuracy (d′), recency discrimination (z sensitivity), confidence for recency discrimination | No sig. effects of sleep condition on recognition; sig. effect of sleep condition on recency discrimination (SD worse) and confidence rating (SD groups were more confident about being correct when they were wrong) | No-sleep effects on face recognition accuracy, but a lack of sleep significantly impaired the temporal memory component for the recognition of faces |
| Hussain et al. [89] | 2008 | 103 (74F + 29M) (G1-4: 24 each, G5: 7) | 20.9 (3.31) | Between-subjects (for the sleep manipulation); 5 groups, with varying interval and sleep between sessions 1 and 2: G1 = 9 a.m. encoding—24 h interval with sleep; G2 = 9 p.m. encoding—24 h interval with sleep; G3 = 9 p.m. encoding—12 h interval with sleep; G4 = 9 a.m. encoding—12 h interval no sleep; G5 = 9 a.m. encoding—3 h interval no sleep | Explicit face identification task where a face that was presented at the beginning of each trial had to be identified amongst an array of 10 simultaneously presented faces; same stimuli in sessions 1 and 2 | Two sessions for each group: G1 = 9 a.m. and 9 a.m. (next day); G2 = 9 p.m. and 9 p.m. (next day); G3 = 9 p.m. and 9 a.m. (next day); G4 = 9 a.m. and 9 p.m. (same day); G5 = 9 a.m. and 12 p.m. (same day) | Same as training ToD | Immediate | No, but chronotype was assessed (participants were unbiased towards ToD and values did not differ between groups) | None; Sleep groups: instructed to sleep normally overnight (between sessions); No-Sleep groups: instructed to not sleep or nap between sessions | Not controlled | Proportion of correct responses | Marginally greater improvement from session 1 to 2 (learning) in the sleep than no-sleep groups; small drop in performance of no-sleep groups in session 2, but restricted to first 102 trials; larger improvement in the 12 h sleep groups than in the 24 h sleep groups | Small positive effect of sleep on face identification, but the authors conclude that the effect of sleep on between session learning is negligible and that “robust perceptual learning for a face identification task can be obtained in the absence of sleep” (p. 2792) |
| Martella et al. [90] | 2012 | 60 (34F + 26M) (Total sleep deprivation group = 26; control/sleep group = 34) | 21.67 (5.54) | Between-subjects; 2 groups: sleep deprivation (SD) group vs. control/sleep group (CG) | Adapted old-new recognition task: training on a 6-face array, followed by individual faces and asked to indicate whether each face was in the previous array | CG: 8 p.m. SD: 4 a.m. | Same as encoding | Almost immediate: 5, 10, 25, or 32 s. random intervals | Daily sleep questionnaire (1 week), normal sleep duration and schedule | Observational (staying in the lab) | CG: unrestricted regular sleep schedule and duration; SD: no sleep | % correct responses; median correct RTs | Accuracy sig. higher for CG than SD; no effects for RTs | Detrimental effect of sleep deprivation on face recognition memory |
| Maurer et al. [91] | 2015 | 22 (11F + 11M) Exp. 1: 14 (8F + 6M) Exp. 2: 8 (3F + 5M) (data from 2 studies pooled for analysis) | Exp. 1: 23.33 [21,22,23,24,25,26,27,28] Exp. 2: 24.75 (3.37) [22,23,24,25,26,27,28,29,30] | Within-subjects; 2 conditions: sleep vs. wake In Exp. 2, only sleep condition | Face-name task (explicit): learn to associate names to faces and later required to indicate if a specific pairing is correct or incorrect + rate confidence in the response (scale 1–9) | Sleep condition: evening, 2.5 h prior to scheduled sleep; wake condition: morning, 1.5 h after waking | Sleep condition: morning, 1.5 h after scheduled wake time; wake condition: evening, 12 h later, during the same wake episode | 12 h | Actigraphy, sleep-wake diaries, kept a regular 8 h sleep schedule (1 week) | In the lab: 13-day inpatient circadian rhythm study; polysomnography during some sleep episodes | Approx. 8 h in the sleep condition | Proportion of correct responses; confidence rating; high confidence (rating 7–9) correct responses; RTs | Sig. higher proportion of correct responses in the sleep than the wake condition; 12% more highly confident responses in the sleep condition; 30% less incorrect responses in the sleep condition; no sig. effects for RTs | Post-learning sleep (in the retention period) had a significantly positive effect on recognition memory for face-name associations |
| Mograss et al. [77] | 2006 | 13 (7F + 6M) | [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39] | Within-subjects; 2 conditions: sleep between learn and test vs. wake between learn and test; 2 sessions, 3–7 days apart | Old-new recognition task; explicit memory task | Sleep condition: acquisition 5–7 p.m.; wake condition: acquisition 7–9 a.m. | Sleep condition: test/recall 7–9 a.m.; wake condition: test/recall 5–7 p.m. | Sleep condition: 12–16 h; wake condition: 8–12 h | Sleep agenda with questions about sleep habits and sleep quality for 3 days prior and during the experiment; Stanford Sleepiness Scale | Self-report questionnaires; observational (night of sleep spent in the lab) | Between 7.0 and 7.8 h/night on the various measures of sleep; no significant differences between sleep at home and in the lab | % of hits; RTs of correct responses | Significantly lower recognition of “old” items in the wake than in the sleep condition; no-sleep effects on RTs | Better performance after a night of sleep compared with daytime wake |
| Mograss et al. [92] | 2008 | 18 (9F + 9M) | 29 [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39] | Within-subjects; 2 conditions: sleep between learn and test vs. wake between learn and test; 2 sessions, 4–7 days apart | Old-new recognition task; explicit memory task | Sleep condition: acquisition 4–6 pm; wake condition: acquisition 7–9 a.m. | Sleep condition: test/recall 7–9 a.m.; wake condition: test/recall 4–6 pm | Sleep condition: 13–17 h; wake condition: 7–11 h | Participants asked to keep regular sleep cycles for at least 3 days prior to the experiment, fill sleep diaries; refrain from taking naps during the day of testing | Sleep agenda; night of sleep in the lab (sleep condition) Stanford Sleepiness Scale (SSS) prior to the testing | Between 7.0 and 7.8 h/night on the various measures of sleep; no significant differences between sleep at home and in the lab | % of hits; % of misses; RTs of correct responses | Marginally more hits and significantly fewer misses to “old” stimuli in the sleep compared to the wake condition; no-sleep effects on RTs | More accurate performance after sleep compared to wakefulness during the retention period, suggesting a positive role of sleep in memory consolidation; fewer misses suggest that less information is forgotten after sleep |
| Mograss et al. [93] | 2009 | 18 (9F + 9M) | 21.9 (2.8) [18,19,20,21,22,23,24,25,26,27,28,29] | Within-subjects; 2 conditions: sleep vs. total sleep deprivation (TSD) | Old-new recognition task; explicit memory task | 4–6 p.m. | 7–9 a.m. | 13–17 h | Participants asked to keep regular sleep cycles for at least 3 days prior to the experiment, fill sleep diaries; refrain from taking naps during the day of testing | Observational (two nights spent in the lab); Sleep quality questionnaire; vigilance scale | Sleep session: 7.6 h (0.32) TSD session: no sleep | % of hits; % of misses; RTs of correct responses and RTs of errors analyzed separately | Trend towards higher correct rejection of new stimuli following sleep compared to TSD (p = 0.07); sig. slower RTs on false alarms (FAs) after TSD compared to sleep | General tendency towards worse performance (increased difficulty to discriminate old and new items) following TSD, compared to normal sleep |
| Solomonova et al. [78] | 2017 | 14 (Gender distribution not reported) REMD = 7 CG = 7 | Not reported | Between-subjects; 2 groups: Partial REM sleep deprivation (REMD) group vs. control group (CG) | Old-new recognition task; incidental learning task: learning occurred through VR interactions with 3D characters (not real faces) | Evening (before bed time) | Morning | Not reported (one night) | Not reported | Observational (in the lab); EEG, electrooculogram | Mean = 5.41 h (groups differed on the amount of REM sleep, but not in the total sleep duration) | Proportion of correct responses | REMD performed significantly worse than CG; differential relationship between face recognition and fast and slow sleep spindles | REM-deprived participants showed worse recognition than non-REM-deprived participants; relationship with sleep spindles still unclear |
| Wagner et al. [85] | 2003 (Main experiment) | 24 (all male) | Not reported | Mixed design: within-subjects manipulation: relevant retention interval early sleep (SWS) vs. late sleep (REM); between-subjects manipulation: sleeping or being wake during the retention interval | Repetition priming task (implicit task): during study, participants indicated the sex of the faces; during test, with the same faces among distractors, participants indicated the viewing direction of the faces | Early-sleep (ES) condition: 10:30 p.m. Late-sleep (LS) condition: 2:15–2:45 a.m. | Early-sleep (ES) condition: 2:15–2:45 a.m. Late-sleep (LS) condition: 6:00–6:30 a.m. | 3 h | Healthy, regular sleepers, no smokers, medication-free | Polysomnography | Sleep groups ES: 3 h between learning and testing; LS: 3 h prior to learning, plus 3 h between learning and testing. Wake groups ES: no sleep; LS: 3 h prior to learning, no sleep between learning and testing | RTs of correct responses; priming effect calculated as the difference between the mean RT for ‘new’ and ‘old’ faces | No main effect of sleep-wake; significant interaction sleep-wake x ES/LS: increased RT for old faces compared to new faces (i.e., an inverse priming effect) in the LS condition | No main effects of sleep; however, REM sleep apparently impaired the expected priming effect. A possible explanation was that, instead of meaning impaired face memory, the inverse priming could indicate improved recognition, which was confirmed in the supp. experiment. |
| Wagner et al. [85] | 2003 (Supplementary experiment) | 19 (all male) | Not reported | Between-subjects: relevant retention interval (early sleep (SWS) vs. late sleep (REM)) | Repetition priming with explicit recognition task: study—indicates the sex of the faces; test—old-new recognition task | Early-sleep (ES) condition: 10:30 p.m. Late-sleep (LS) condition: 2:15–2:45 a.m. | Early-sleep (ES) condition: 2:15–2:45 a.m. Late-sleep (LS) condition: 6:00–6:30 a.m. | 3 h | Healthy, regular sleepers, no smokers, medication-free | Polysomnography | ES: 3 h between learning and testing; LS: 3 h prior to learning, plus 3 h between learning and testing. | RTs of correct responses; priming effect calculated as the difference between the mean RT for ‘new’ and ‘old’ faces | VD: RTs significant positive priming effect, i.e., faster RT for old faces, more pronounced in the LS condition | Positive effect of REM sleep on memory, which strengthened automatic face recognition (this interfered with the implicit task used in the main experiment) |
| Wagner et al. [79] | 2007 | 12 (5F + 7M) | [19,20,21,22,23,24,25,26,27,28,29,30] | Within-subjects; 2 conditions: sleep in the night following learning vs. wake in the night following learning; 2 sessions, at least 2 weeks apart | Old-new recognition task, with emotion faces; at encoding, participants had to indicate the emotional valence of the expression; no explicit memorization instruction | 10:30–11:00 p.m. | 6:00–6:30 p.m. (on the second evening after learning, allowing a full night of sleep at home) | Approx. 43 h | Regular sleep habits and normal sleep duration, night of adaptation in the lab | Night after encoding spent in the lab (asleep or awake); Polysomnography in the sleep condition; daytime sleep not allowed, and controlled by actigraphy in the wake condition | Sleep condition: approx. 8 h after learning, plus 1 night before testing; wake condition: total sleep deprivation after learning, plus 1 night before testing | Hit rate (HR); false alarm rate (FAR); memory accuracy, Pr [= HR - FAR]; response bias, Br [= FAR/(1 − Pr)]; RTs | Memory accuracy (hit rate-FA rate) was enhanced by sleep compared to wakefulness; sleep did not affect hits, false alarms, response bias, and RTs. Sleep condition: total sleep duration and amount of non-REM sleep in the consolidation night was significantly correlated with memory accuracy. | Sleep during consolidation improved face recognition memory; positive relation between memory accuracy and non-REM sleep and total sleep (but not REM sleep) duration |
| NEGATIVE EFFECTS, i.e., the detrimental effects of sleep on performance | ||||||||||||||
| Alberca-Reina et al. [94] | 2014 | 60 (31F + 29M) | 22 (2.7) [18,19,20,21,22,23,24,25,26,27] | Between-subjects; 3 groups: CG—8 h of sleep before and after training; SRPre-T—only 4 h of sleep pre-training; SRPost-T—only 4 h of sleep post-training (i.e., acute sleep restriction) | Day 1 (6:30 p.m.): semantic-perceptual matching task; informed of subsequent memory test. Day 2 (9 a.m.): retroactive interference task. Day 2 (11:30 am): associative memory test to the initial face pairs. Day 3 (11:30 am): associative memory test without interference | 6:30 pm day 1), following a familiarization procedure at 5 pm | 11:30 a.m. (day 2), 11:30 a.m. (day 3) | 17 h (day 2), 41 h since the initial encoding (day 3) | Sleep diaries (1 week before experiment), structured interview, abstain from substance intake | Observational (sleep in the lab) on the night before and the first night after training; slept at home on the second night | CG: 8 h (days 1–3) SRPre-T: 4 h (day 1), 8 h (days 2–3) SRPost-T: 8 h (days 1 and 3), 4 h (day 2) | RTs to correctly recognized stimuli; d′; estimates of recollection and familiarity processes derived from the dual-process signal detection model (Yonelinas et al., 1998) | Session 2: In comparison with the CG, the SRPre-T showed enhanced recognition for semantically congruent faces and decreased for semantically incongruent faces; SRPost-T - trend for enhanced memory for both semantically congruent and incongruent associations | The results seem to suggest that sleep restriction exerted a beneficial effect on associative recognition memory for face pairs |
| Mograss et al. [81] | 2010 | 24 (12F + 12M) Short sleepers (SSs) = 8 (3F/5M). Average sleepers (ASs) = 9 (4F/5M). Long sleepers (LSs) = 7 (4F/3M). | [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39] SSs: 30.3 (5.8); ASs: 23.1 (4.0); LSs: 21.7 (2.1) | Between-subjects 3 groups: SSs vs. ASs vs. LSs | Old-new recognition task; explicit memory task | 4–6 pm | 7–9 a.m. | 13–17 h | Participants asked to keep regular sleep cycles for at least 3 days prior to the experiment, fill sleep diaries and diverse questionnaires, interview | Night of sleep in the lab; sleep questionnaire; 7–10 day sleep log | Home: SSs: 6.8 h (0.23) ASs: 8.1 h (0.45) LSs: 9.1 h (0.52) Lab: SSs: 6.9 h (1.2) ASs: 8.3 h (0.70) LSs: 8.9 h (0.39) | % hits; % false alarms; % misses; RTs to correct responses for “old” and “new” items separately | % hits: SSs significantly better than ASs and LSs; ASs and LSs did not differ; the retention of old items in LSs did not differ from chance, while SSs were sig higher; % of hits decreased with sleep duration in SSs, but it increased with sleep duration in ASs and LSs | Longer sleep duration yielded worse recognition memory compared to average and short sleep durations Individual differences in sleep duration might be related to individual differences in face recognition memory |
| NULL EFFECTS, i.e., sleep manipulation has no effect on performance | ||||||||||||||
| Alberca-Reina et al. [95] | 2015 | 40 (21F + 19M) | 21.8 (2.7) [18,19,20,21,22,23,24,25,26,27] | Between-subjects; 2 groups: control group (CG): sleep 8 h a night pre-training; acute sleep restriction (ASR): sleep 4 h a night pre-training | Day 1 (6:30 pm): training—semantic-perceptual matching task; informed of subsequent memory test. Day 2 (9:30 a.m.): retroactive interference task. Day 2 (12:00 p.m.): associative memory test to the initial face pairs. | 6:30 pm (Day 1) | 12 pm (Day 2, after a full night of sleep) | 17 h 30 min | Sleep diaries (1 week before experiment), structured interview, abstain from substance intake | Observational (sleep in the lab) on the nights before and after training | CG.: 8 h ASR: 4 h | Hit rate; false alarm rate; RTs; d′ (associative d′ and semantic d′) | No significant differences between groups in any memory indices and no interactions with the semantic congruence of the face pairs | No effect of sleep restriction on recognition memory for face-face associations |
| Chee et al. [80] | 2010 | 26 (14F + 12M) | 20.7 (1.9) | Within-subjects; 2 sessions: rested wakefulness (RW) vs. sleep deprivation (SD) (1 week apart) | Old-new recognition task; incidental learning task: encoding occurred during a selective attention task with 3 conditions (attend faces, scenes, or both) | RW: 8 a.m. (after a night of sleep); SD: 5 a.m. (after a whole day and night awake without napping) | Same as encoding (shortly after) | 10–15 min. after training | Actigraphy for 2 weeks, only participants with good sleeping habits; no extreme chronotype, controlled substance intake (24 h) | SD: observational (lab); RW: actigraphy (home) | RW: not reported (regular sleep); SD: no sleep | Analysis only of valid trials with responses to target stimuli during the encoding phase; RTs; response accuracy with A′ | The effect of sleep on face recognition accuracy was not significant (contrary to what happen for the recognition of scenes, which was sig. reduced in SD) | No effect of sleep: face recognition was not affected by sleep deprivation; the authors suggest that it may be a more automatic process |
| Sheth et al. [96] | 2009 | 112 (55F + 57M) 7 groups (16 participants in each) | 25.25 | Between-subjects; 7 groups that varied on acquisition and test times, as well as the retention period, intervening sleep, and intervening wake | Old-new recognition task + Confidence rating; Explicit memory task; Stimuli were computer-generated faces | G1: 9 p.m. G2: 9 p.m. G3: 9 p.m. G4: 9 a.m. G5: 9 a.m. G6: 9 a.m. G7: 9 p.m. | G1: 9 a.m. G2: 9 a.m. G3: 9 p.m. G4: 9 a.m. G5: 9 p.m. G6: 9 a.m. G7: 9 p.m. | G1: 12 h G2: 36 h G3: 24 h G4: 24 h G5: 12 h G6: 5 min G7: 5 min | Not controlled, but participants were selected on the basis of a screening questionnaire for substance intake, sleep habits and sleep duration | Actigraphy with a limited number of participants; sleep diaries | G1: sleep G2: sleep x2 G3: sleep G4: sleep G5: awake G6: awake G7: awake Sleep consisted in 1 or 2 normal nights of sleep (around 7.5 h) | Memory accuracy (d′); response bias (c) | Effect of sleep on d′ was not significant; effect on response bias: intervening wake during retention rendered the subject less likely to report seeing a test face before | No effect: sleep during retention did not appear to improve face recognition memory; however, the intervening wake time seems to impair memory strength |
| Stare et al. [97] | 2018 | 93 (62F + 29M + 2 unknown) | 21.38 (5.01) [18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47] | Between-subjects (4 groups: wakefulness, sleep, morning control, evening control) | Old-new recognition task; answer given on a confidence scale of 1 (confident not seen) to 4 (confident seen). Incidental memory task. | 9 a.m. for the wakefulness and the morning control group; 9 p.m. for the sleep and the evening control group | 9 p.m. for the wakefulness and the evening control group; 9 a.m. for the sleep and the morning control group | 12 h for the wakefulness and sleep groups; immediately for the morning and evening control groups | Screening/demographics form; Stanford Sleepiness Scale | Polysomnography for participants staying in the lab; oral instructions to sleep or not sleep to those not staying in the lab | Participants in the sleep group were asked to try to sleep for at least 6 h | Corrected recognition scores (hits-false alarms) | No main effect of group (p = 0.36). No correlation of memory for faces with time spent in SWS, N2 and REM, total sleep time, or sleepiness. | No effect of sleep on memory for faces |
| Sleep Manipulation | Sleep Restriction | Amount of Restriction | Task | Encoding | Testing | VD | ||
|---|---|---|---|---|---|---|---|---|
| Alberca-Reina et al. [95] | 2015 | Between | Pre-training | 4 h of sleep, night | Associative memory with interference | Evening | Morning | Various (RT, hits, FA, d′, associative d′, semantic d′) |
| Chee et al. [80] | 2010 | Within | Pre-training | Total sleep deprivation, night | Old-new; incidental learning | Morning | Morning | RT, A′ |
| Sheth et al. [96] | 2009 | Between | Post-encoding | Day time awake (12 h) | Old-new; explicit memory | Varied | Varied | d′, c |
| Stare [97] | 2018 | Between | Post-encoding | Day time awake (12 h) | Old-new; incidental learning | Morning (wake)/evening (sleep) | Evening (wake)/morning (sleep) | Corrected recognition scores (hits-false alarms) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, I.M.; Silva, A.; Bem-Haja, P.; Rosa, C.; Cerri, L.; Queiroz, D.F.; Barroso, T.; Alves, M.F.; Silva, C.F. The Impact of Sleep on Face Recognition Memory: A Scoping Review. Brain Sci. 2022, 12, 1385. https://doi.org/10.3390/brainsci12101385
Santos IM, Silva A, Bem-Haja P, Rosa C, Cerri L, Queiroz DF, Barroso T, Alves MF, Silva CF. The Impact of Sleep on Face Recognition Memory: A Scoping Review. Brain Sciences. 2022; 12(10):1385. https://doi.org/10.3390/brainsci12101385
Chicago/Turabian StyleSantos, Isabel M., André Silva, Pedro Bem-Haja, Catarina Rosa, Luíza Cerri, Diâner F. Queiroz, Talles Barroso, Miguel F. Alves, and Carlos F. Silva. 2022. "The Impact of Sleep on Face Recognition Memory: A Scoping Review" Brain Sciences 12, no. 10: 1385. https://doi.org/10.3390/brainsci12101385