
Topological Characteristics Associated with Intraoperative Stimulation Related Epilepsy of Glioma Patients: A DTI Network Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. MRI Acquisition
2.3. DTI Data Preprocessing
2.4. Operation and Stimulation Protocol
2.5. Tumor Region of Interest Extraction
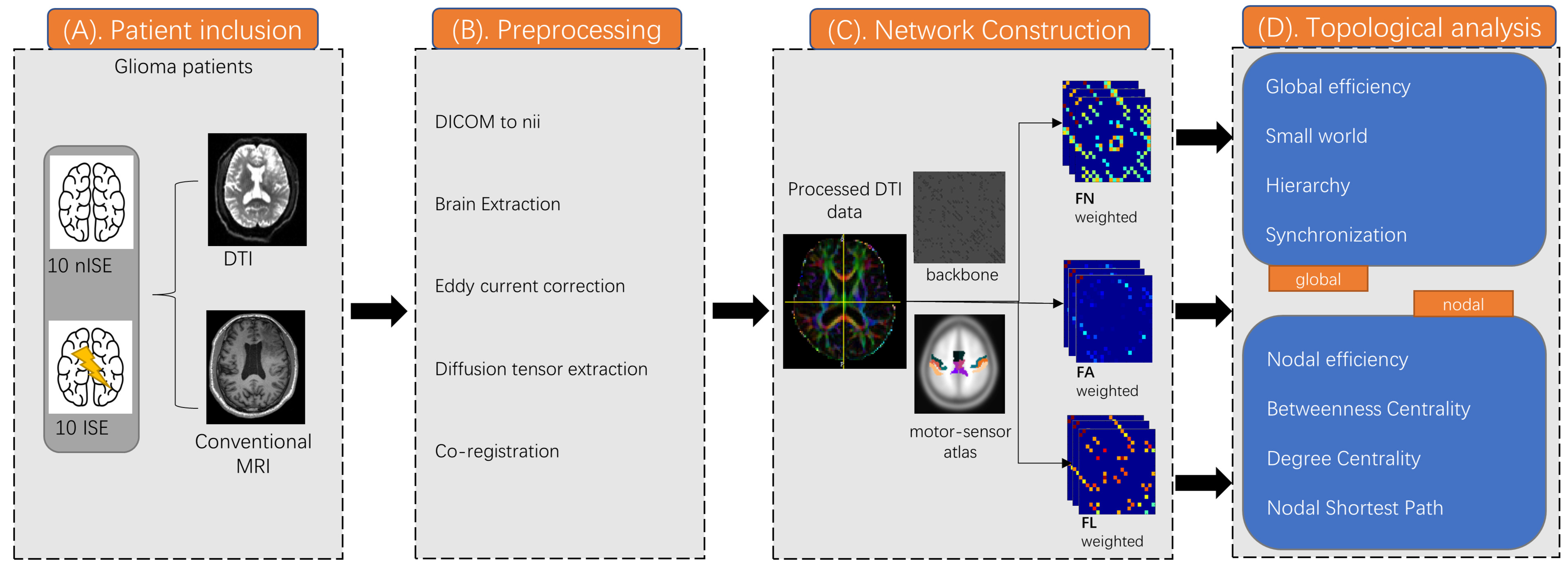
2.6. Network Construction
2.7. Graph Theoretical Measures
2.8. Statistical Analysis
3. Results
3.1. Demographic Characteristics
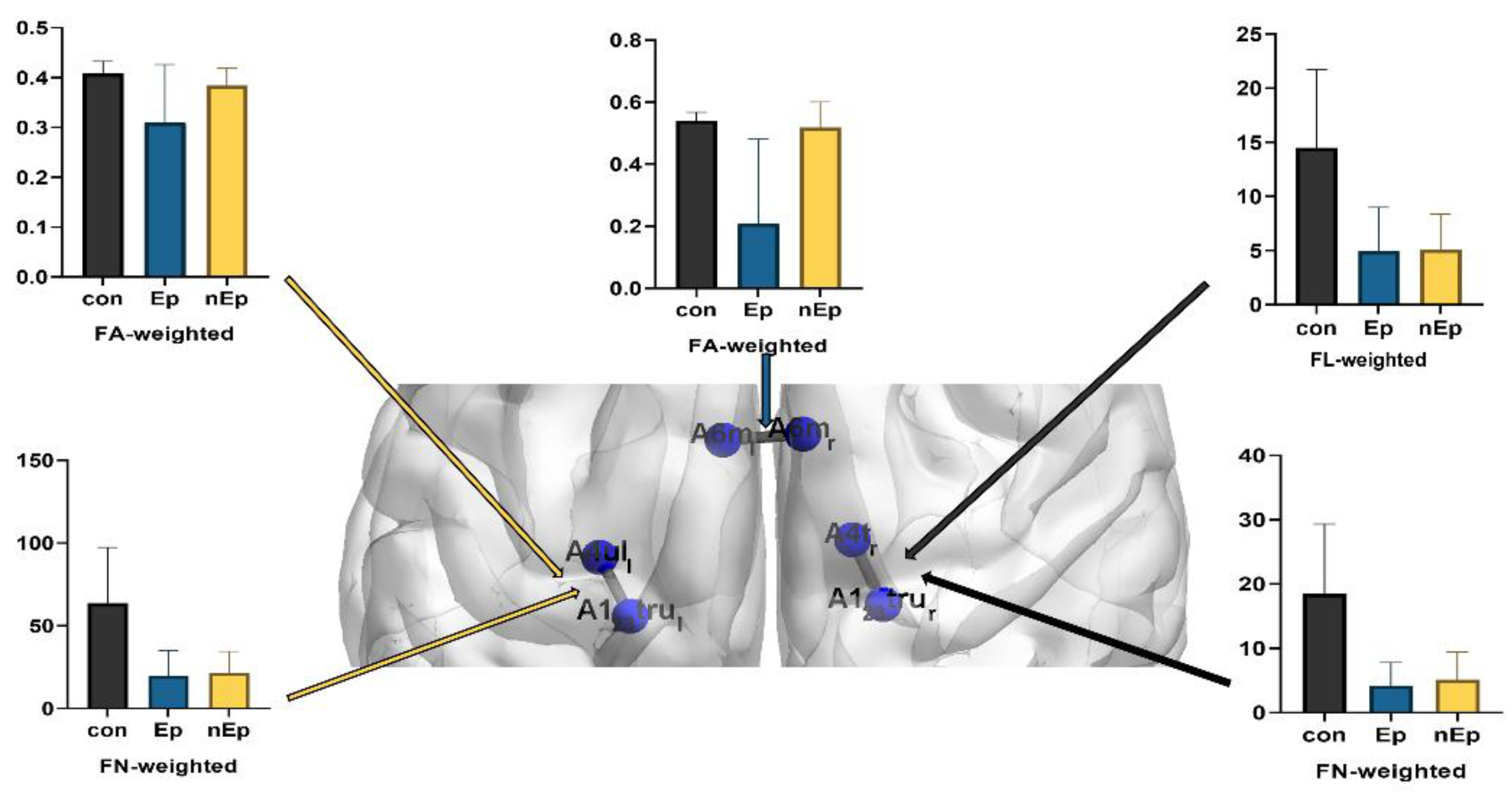
3.2. Connections Differences
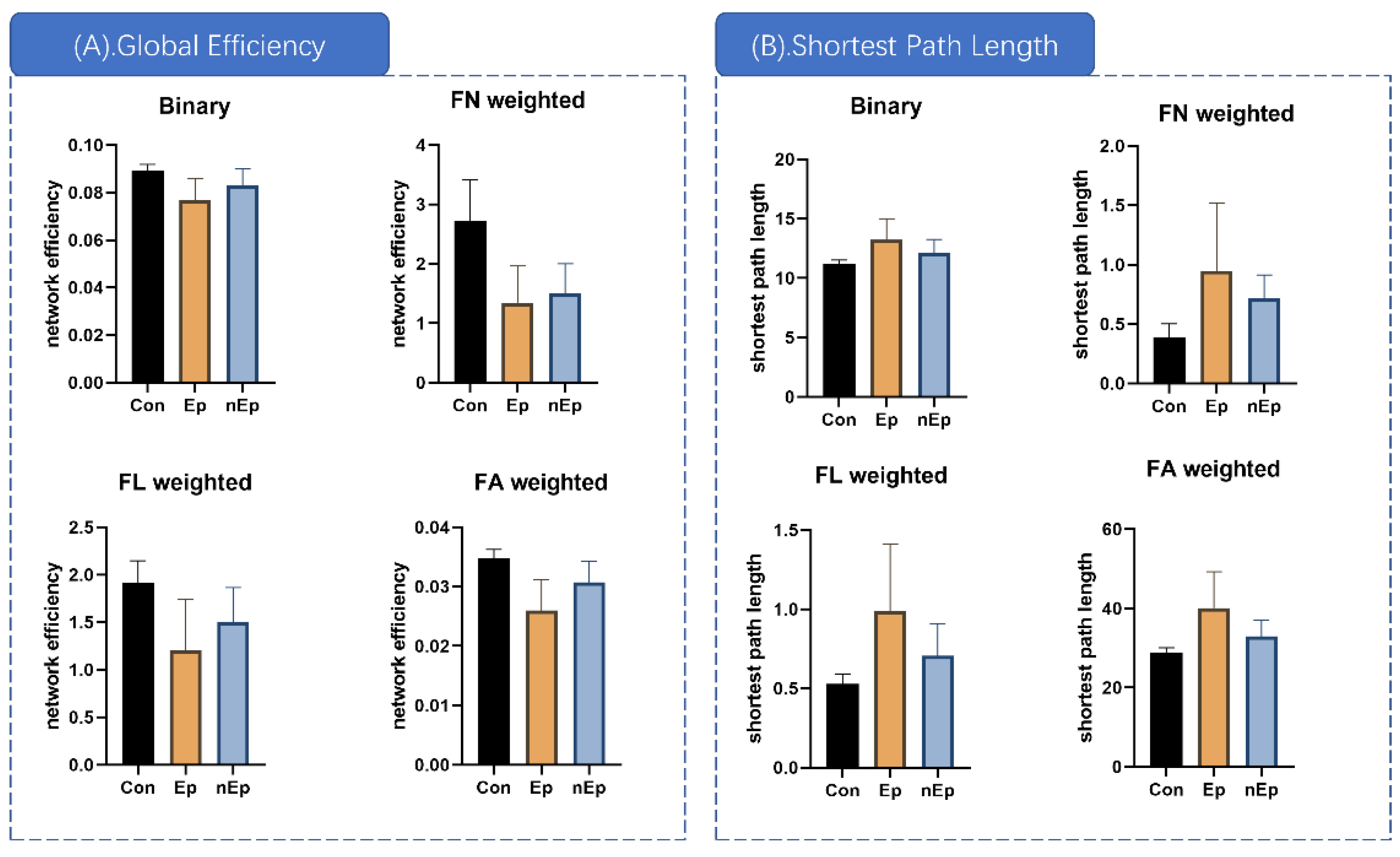
3.3. Global Properties Differences
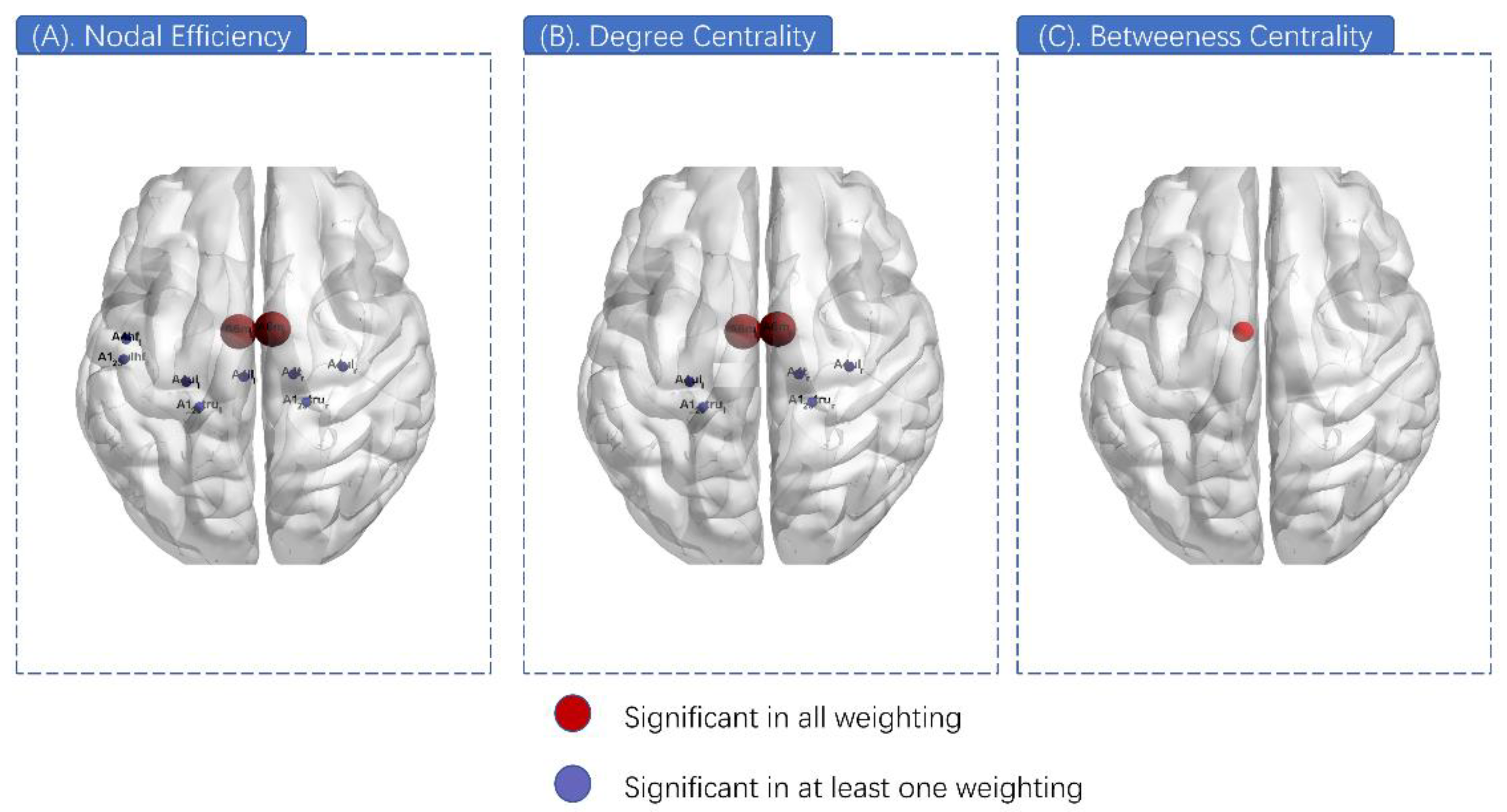
3.4. Nodal Properties Differences
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bampoe, J.; Laperriere, N.; Pintilie, M.; Glen, J.; Micallef, J.; Bernstein, M. Quality of life in patients with glioblastoma multiforme participating in a randomized study of brachytherapy as a boost treatment. J. Neurosurg. 2000, 93, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Hadjipanayis, C.J.N. Surgery of intrinsic cerebral tumors. Neurosurgery 2007, 61, 279–304; discussion 275–304. [Google Scholar] [CrossRef] [PubMed]
- Duffau, H. Lessons from brain mapping in surgery for low-grade glioma: Insights into associations between tumour and brain plasticity. Lancet Neurol. 2005, 4, 476–486. [Google Scholar] [CrossRef]
- Serletis, D.; Bernstein, M. Prospective study of awake craniotomy used routinely and nonselectively for supratentorial tumors. J. Neurosurg. 2007, 107, 1–6. [Google Scholar] [CrossRef]
- Szelenyi, A.; Bello, L.; Duffau, H.; Fava, E.; Feigl, G.C.; Galanda, M.; Neuloh, G.; Signorelli, F.; Sala, F. Workgroup for Intraoperative Management in Low-Grade Glioma Surgery within the European Low-Grade Glioma Network Intraoperative electrical stimulation in awake craniotomy: Methodological aspects of current practice. Neurosurg. Focus 2010, 28, E7. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, C.J.; Berger, M.S. Rapid termination of intraoperative stimulation-evoked seizures with application of cold Ringer’s lactate to the cortex. Technical note. J. Neurosurg. 1998, 88, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Nossek, E.; Matot, I.; Shahar, T.; Barzilai, O.; Rapoport, Y.; Gonen, T.; Sela, G.; Grossman, R.; Korn, A.; Hayat, D.; et al. Intraoperative seizures during awake craniotomy: Incidence and consequences: Analysis of 477 patients. Neurosurgery 2013, 73, 135–140; discussion 140. [Google Scholar] [CrossRef]
- Herrick, I.A.; Craen, R.A.; Gelb, A.W.; McLachlan, R.S.; Girvin, J.P.; Parrent, A.G.; Eliasziw, M.; Kirkby, J. Propofol sedation during awake craniotomy for seizures: Electrocorticographic and epileptogenic effects. Anesth. Analg. 1997, 84, 1280–1284. [Google Scholar] [CrossRef]
- Abecassis, Z.A.; Ayer, A.B.; Templer, J.W.; Yerneni, K.; Murthy, N.K.; Tate, M.C. Analysis of risk factors and clinical sequelae of direct electrical cortical stimulation-induced seizures and afterdischarges in patients undergoing awake mapping. J. Neurosurg. 2020, 134, 1610–1617. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Winston, G.P. The potential role of novel diffusion imaging techniques in the understanding and treatment of epilepsy. Quant. Imaging Med. Surg. 2015, 5, 279–287. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Coman, D.; Munbodh, R.; Dhaher, R.; Zaveri, H.; Hyder, F.; Eid, T.J.E. Network evolution in mesial temporal lobe epilepsy revealed by diffusion tensor imaging. Epilepsia 2017, 58, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Dyhrfjeld-Johnsen, J.; Santhakumar, V.; Morgan, R.J.; Huerta, R.; Tsimring, L.; Soltesz, I. Topological determinants of epileptogenesis in large-scale structural and functional models of the dentate gyrus derived from experimental data. J. Neurophysiol. 2007, 97, 1566–1587. [Google Scholar] [CrossRef] [Green Version]
- Moseley, M.E.; Cohen, Y.; Kucharczyk, J.; Mintorovitch, J.; Asgari, H.S.; Wendland, M.F.; Tsuruda, J.; Norman, D. Diffusion-weighted MR imaging of anisotropic water diffusion in cat central nervous system. Radiology 1990, 176, 439–445. [Google Scholar] [CrossRef]
- Winston, G.P. The physical and biological basis of quantitative parameters derived from diffusion MRI. Quant. Imaging Med. Surg. 2012, 2, 254–265. [Google Scholar] [CrossRef]
- Le Bihan, D.; Mangin, J.F.; Poupon, C.; Clark, C.A.; Pappata, S.; Molko, N.; Chabriat, H. Diffusion tensor imaging: Concepts and applications. J. Magn. Reson. Imaging 2001, 13, 534–546. [Google Scholar] [CrossRef]
- von Podewils, F.; Runge, U.; Krüger, S.; Geithner, J.; Wang, Z.I.; Khaw, A.V.; Angermaier, A.; Gaida, B.; Domin, M.; Kessler, C.; et al. Diffusion tensor imaging abnormalities in photosensitive juvenile myoclonic epilepsy. Eur. J. Neurol. 2015, 22, 1192–1200. [Google Scholar] [CrossRef]
- Park, K.M.; Lee, B.I.; Shin, K.J.; Ha, S.Y.; Park, J.; Kim, T.H.; Mun, C.W.; Kim, S.E. Progressive topological disorganization of brain network in focal epilepsy. Acta Neurol. Scand. 2018, 137, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.J.; Yu, Y.; Miao, H.H.; Wang, Z.J.; Tang, Y.L.; Liao, W. Decreased Network Efficiency in Benign Epilepsy with Centrotemporal Spikes. Radiology 2017, 283, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponten, S.C.; Bartolomei, F.; Stam, C.J. Small-world networks and epilepsy: Graph theoretical analysis of intracerebrally recorded mesial temporal lobe seizures. Clin. Neurophysiol. 2007, 118, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhong, S.; Xu, P.; He, Y.; Gong, G. PANDA: A pipeline toolbox for analyzing brain diffusion images. Front. Hum. Neurosci. 2013, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A graph theoretical network analysis toolbox for imaging connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Vulliemoz, S.; Vollmar, C.; Koepp, M.J.; Yogarajah, M.; O’Muircheartaigh, J.; Carmichael, D.W.; Stretton, J.; Richardson, M.P.; Symms, M.R.; Duncan, J.S. Connectivity of the supplementary motor area in juvenile myoclonic epilepsy and frontal lobe epilepsy. Epilepsia 2011, 52, 507–514. [Google Scholar] [CrossRef]
- Mallela, A.; Peck, K.; Petrovich-Brennan, N.; Zhang, Z.; Lou, W.; Holodny, A.I. Altered Resting-State Functional Connectivity in the Hand Motor Network in Glioma Patients. Brain Connect. 2016, 6, 587–595. [Google Scholar] [CrossRef]
- Mohan, A.; Roberto, A.; Mohan, A.; Lorenzo, A.; Jones, K.; Carney, M.; Liogier-Weyback, L.; Hwang, S.; Lapidus, K.A. The Significance of the Default Mode Network (DMN) in Neurological and Neuropsychiatric Disorders: A Review. Yale J. Biol. Med. 2016, 89, 49–57. [Google Scholar] [PubMed]
- Miro, J.; Gurtubay-Antolin, A.; Ripolles, P.; Sierpowska, J.; Juncadella, M.; Fuentemilla, L.; Sanchez, V.; Falip, M.; Rodriguez-Fornells, A. Interhemispheric microstructural connectivity in bitemporal lobe epilepsy with hippocampal sclerosis. Cortex 2015, 67, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Netoff, T.; Clewley, R.; Arno, S.; Keck, T.; White, J.A. Epilepsy in small-world networks. J. Neurosci. 2004, 24, 8075–8083. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.; Powell, F. Models of Network Spread and Network Degeneration in Brain Disorders. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 788–797. [Google Scholar] [CrossRef]
- Bernhardt, B.C.; Chen, Z.; He, Y.; Evans, A.C.; Bernasconi, N. Graph-theoretical analysis reveals disrupted small-world organization of cortical thickness correlation networks in temporal lobe epilepsy. Cereb. Cortex 2011, 21, 2147–2157. [Google Scholar] [CrossRef] [PubMed]
- Vlooswijk, M.C.; Vaessen, M.J.; Jansen, J.F.; de Krom, M.C.; Majoie, H.J.; Hofman, P.A.; Aldenkamp, A.P.; Backes, W.H. Loss of network efficiency associated with cognitive decline in chronic epilepsy. Neurology 2011, 77, 938–944. [Google Scholar] [CrossRef]
- Eddin, A.S.; Wang, J.; Wu, W.; Sargolzaei, S.; Bjornson, B.; Jones, R.A.; Gaillard, W.D.; Adjouadi, M. The effects of pediatric epilepsy on a language connectome. Hum. Brain Mapp. 2014, 35, 5996–6010. [Google Scholar] [CrossRef]
- Widjaja, E.; Zamyadi, M.; Raybaud, C.; Snead, O.C.; Doesburg, S.M.; Smith, M.L. Disrupted Global and Regional Structural Networks and Subnetworks in Children with Localization-Related Epilepsy. Am. J. Neuroradiol. 2015, 36, 1362–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Chu, L.; Liu, C.; Huang, M.; Wang, H. Alterations of white matter network in patients with left and right non-lesional temporal lobe epilepsy. Eur. Radiol. 2019, 29, 6750–6761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liao, W.; Chen, H.; Mantini, D.; Ding, J.R.; Xu, Q.; Wang, Z.; Yuan, C.; Chen, G.; Jiao, Q.; et al. Altered functional-structural coupling of large-scale brain networks in idiopathic generalized epilepsy. Brain 2011, 134, 2912–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Jiao, D.; Zhang, X.; Lin, X. Altered degree centrality in childhood absence epilepsy: A resting-state fMRI study. J. Neurol. Sci. 2017, 373, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Chaichana, K.L.; Parker, S.L.; Olivi, A.; Quinones-Hinojosa, A. Long-term seizure outcomes in adult patients undergoing primary resection of malignant brain astrocytomas. Clinical article. J. Neurosurg. 2009, 111, 282–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, H.; Roos, D.; Blümcke, I.; Pietsch, T.; Wiestler, O.D. Perilesional neurochemical changes in focal epilepsies. Acta Neuropathol. 1996, 91, 376–384. [Google Scholar] [CrossRef]
- Pallud, J.; Capelle, L.; Huberfeld, G.J.E. Tumoral epileptogenicity: How does it happen? Epilepsia 2013, 54, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Williamson, A.; Patrylo, P.R.; Lee, S.; Spencer, D.D. Physiology of human cortical neurons adjacent to cavernous malformations and tumors. Epilepsia 2003, 44, 1413–1419. [Google Scholar] [CrossRef] [Green Version]
- Humphries, M.D.; Gurney, K.; Prescott, T.J. The brainstem reticular formation is a small-world, not scale-free, network. Proc. R. Soc. Biol. Sci. 2006, 273, 503–511. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ep | nEp | Con | p-Value | |
|---|---|---|---|---|
| Final sample size (n) | 10 | 10 | 10 | - |
| Age range (mean ± SE) | 40.50 ± 4.50 | 43.80 ± 3.74 | 40.20 ± 2.41 | 0.580 * |
| Sex (female/male) | 6/4 | 5/5 | 5/5 | 0.711 ^ |
| Education level (yrs) | 14.71 ± 1.02 | 15.20 ± 0.91 | 16.44 ± 0.83 | 0.232 * |
| Language deficits (Y/N) | 3/7 | 3/7 | / | >0.999 ^ |
| Motor deficits (Y/N) | 3/7 | 3/7 | / | >0.999 ^ |
| Diagnosed time (day) | 60.10 ± 22.60 | 65.10 ± 20.96 | / | 0.873 $ |
| Preoperational KPS | 94.00 ± 2.21 | 93.00 ± 2.13 | / | 0.749 $ |
| Histology (HGG/LGG) | 4/6 | 5/5 | / | >0.999 ^ |
| IDH status (MU/WT) | 4/6 | 3/7 | / | >0.999 ^ |
| Tumor volume (mL) | 30.17 ± 5.15 | 31.25 ± 4.81 | / | 0.880 $ |
| Stimulation current (mA) | 3.40 ± 0.49 | 2.85 ± 0.50 | / | 0.444 $ |
| Connections | Weighting | Post-Hoc p Value | ||
|---|---|---|---|---|
| Ep vs. Con | Ep vs. nEp | nEp vs. Con | ||
| A123truL and A4ulL | FA | 0.017 | 0.071 | 0.089 |
| FN | 0.001 | 0.781 | 0.002 | |
| A123truR and A4tR | FN | 0.001 | 0.619 | 0.002 |
| FL | 0.002 | 0.927 | 0.001 | |
| A6mL and A6mR | FA | 0.001 | 0.003 | 0.432 |
| Global Properties | Weighting | p (ANOVA) | Post-Hoc p Value | ||
|---|---|---|---|---|---|
| Ep vs. Con | nEp vs. Con | Ep vs. nEp | |||
| Global efficiency | Binary | 0.001 | <0.001 | 0.015 | 0.105 |
| FA | <0.001 | <0.001 | 0.004 | 0.031 | |
| FN | <0.001 | <0.001 | <0.001 | 0.530 | |
| FL | 0.002 | 0.001 | 0.007 | 0.166 | |
| Shortest path length | Binary | 0.003 | 0.003 | 0.020 | 0.110 |
| FA | <0.001 | 0.001 | 0.006 | 0.036 | |
| FN | 0.006 | 0.007 | <0.001 | 0.245 | |
| FL | 0.003 | 0.003 | 0.016 | 0.073 | |
| Node | ANOVA p Value (FDR Corrected) | |||
|---|---|---|---|---|
| Binary (0.00083) | FA Weighted (0.014) | FN Weighted (0.002) | FL Weighted (0.002) | |
| A6m_L | 0.00026 | <0.001 | 0.002 | 0.002 |
| A6m_R | 0.00078 | <0.001 | 0.002 | 0.002 |
| A4t_R | 0.013 | 0.002 | <0.001 | <0.001 |
| A4ul_L | 0.153 | 0.010 | 0.001 | 0.026 |
| A4ul_R | 0.019 | 0.011 | <0.001 | 0.017 |
| A4ll_L | 0.00083 | 0.002 | 0.271 | 0.028 |
| A123tru_L | 0.233 | 0.014 | 0.001 | 0.004 |
| A123tru_R | 0.185 | 0.068 | <0.001 | 0.005 |
| A123ulhf_L | 0.126 | 0.006 | 0.384 | 0.168 |
| A4hf_L | 0.126 | 0.003 | 0.203 | 0.502 |
| Node | ANOVA p Value (FDR Corrected) | |||
|---|---|---|---|---|
| Binary (0.0037) | FA Weighted (0.013) | FN Weighted (0.002) | FL Weighted (0.006) | |
| A6m_L | <0.001 | <0.001 | 0.002 | 0.002 |
| A6m_R | <0.001 | <0.001 | 0.001 | 0.002 |
| A4t_R | 0.085 | 0.005 | <0.001 | 0.002 |
| A4ul_L | 0.381 | 0.027 | <0.001 | 0.027 |
| A4ul_R | 0.085 | 0.010 | 0.001 | 0.003 |
| A123tru_L | 0.381 | 0.013 | 0.002 | 0.004 |
| A123tru_R | 0.213 | 0.027 | 0.001 | 0.006 |
| Node | Weighting | Post-Hoc p Value | ||
|---|---|---|---|---|
| Ep vs. Con | nEp vs. Con | Ep vs. nEp | ||
| A6m_L | Binary | 0.003 | 0.556 | 0.004 |
| FA | 0.002 | 0.401 | 0.004 | |
| FN | 0.004 | 0.011 | 0.241 | |
| FL | 0.004 | 0.402 | 0.002 | |
| A6m_R | Binary | 0.002 | 0.556 | 0.007 |
| FA | 0.001 | 0.264 | 0.005 | |
| FN | 0.004 | 0.009 | 0.332 | |
| FL | 0.003 | 0.211 | 0.023 | |
| Node | Weighting | Post-Hoc p Value | ||
|---|---|---|---|---|
| Ep vs. Con | nEp vs. Con | Ep vs. nEp | ||
| A6m_L | Binary | 0.004 | >0.999 | 0.004 |
| FA | 0.003 | 0.828 | 0.004 | |
| FN | 0.004 | 0.010 | 0.24 | |
| FL | 0.004 | 0.906 | 0.001 | |
| A6m_R | Binary | 0.006 | 0.556 | 0.017 |
| FA | 0.001 | 0.245 | 0.007 | |
| FN | 0.004 | 0.006 | 0.413 | |
| FL | 0.003 | 0.069 | 0.034 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhou, C.; Liang, Y.; Wang, Y.; Wang, L. Topological Characteristics Associated with Intraoperative Stimulation Related Epilepsy of Glioma Patients: A DTI Network Study. Brain Sci. 2022, 12, 60. https://doi.org/10.3390/brainsci12010060
Yang J, Zhou C, Liang Y, Wang Y, Wang L. Topological Characteristics Associated with Intraoperative Stimulation Related Epilepsy of Glioma Patients: A DTI Network Study. Brain Sciences. 2022; 12(1):60. https://doi.org/10.3390/brainsci12010060
Chicago/Turabian StyleYang, Jianing, Chunyao Zhou, Yuchao Liang, Yinyan Wang, and Lei Wang. 2022. "Topological Characteristics Associated with Intraoperative Stimulation Related Epilepsy of Glioma Patients: A DTI Network Study" Brain Sciences 12, no. 1: 60. https://doi.org/10.3390/brainsci12010060