
A Barrier to Entry: Examining the Bacterial Outer Membrane and Antibiotic Resistance

Department of Life Sciences and Chemistry, Jacobs University Bremen, 28719 Bremen, Germany
Appl. Sci. 2023, 13(7), 4238; https://doi.org/10.3390/app13074238
Submission received: 21 February 2023
/
Revised: 12 March 2023
/
Accepted: 14 March 2023
/
Published: 27 March 2023
(This article belongs to the Special Issue Bioactive Lipids: Chemical, Biological Properties and Pharmaceutical Applications)
Abstract
:Gram-negative bacteria can resist antibiotics by changing the permeability via their outer membrane. These bacteria have a complex cell envelope that incorporates an outer membrane separating the periplasm from the external environment. This outer membrane contains many protein channels, also known as porins or nanopores, which mainly allow the influx of hydrophilic compounds, including antibiotics. One probable way bacteria may possibly develop antibiotic resistance is by reworking to reduce the inflow through these outer membrane porins or nanopores. The challenge now is to recognize and potentially comprehend the molecular basis of permeability via the bacterial outer membrane. To address this challenge, this assessment builds upon the author’s previous work to develop a comprehensive understanding of membrane porins and their crucial role in the influx of antibiotics and solutes. Furthermore, the work aspires to investigate the bacterial response to antibiotic membrane permeability and nurture discussion toward further exploration of the physicochemical parameters governing the translocation/transport of antibiotics through bacterial membrane porins. By augmenting our understanding of these mechanisms, we may devise novel approaches to mitigate antibiotic resistance in Gram-negative bacteria.
1. Introduction
Despite recent efforts to strengthen the antibiotics development pipeline, it is insufficient to meet the increasing demand. Only few antibiotics in various stages of development are expected to become commercially viable products [1,2,3,4,5,6,7]. Many of today’s antibiotics lack novel mechanisms of action or fail to target the most resistant pathogens, such as avibactam ceftazidime-resistant Klebsiella pneumoniae (2015) [8], Daptomycin-resistant Staphylococcus aureus (2004) [9], Imipenem resistant Klebsiella pneumoniae (1996) [10], Azithromycin resistant Neisseria gonorrhoeae (2011) [11], Cephalosporins Extended spectrum inactivating beta-lactamase making Escherichia coli (1983), and the long-established Staphylococcus aureus resistant towards Methicillin (1960) [12]. The development of antibiotics is a challenging process, scientifically and clinically, and large pharmaceutical companies have shifted their focus to more lucrative endeavors, leaving small-scale and intermediate ventures with limited resources to carry out extensive research and development [1,2,3,8,9,10,11,12,13,14,15].
Public programs targeted toward incentivizing antibiotic research have been commissioned over the past decade. Despite political momentum to address antibiotic/antimicrobial resistance, the best utilization of these incentives remains unclear [1,2,3,13,14,16,17]. A framework has been developed to assess these initiatives and identify any gaps in provisions, providing recommendations for improving the scope of antibiotics in a workable and reasonable manner [1,2,15,18,19,20]. Optimism and concern have arisen in antibiotic/antimicrobial research and development in recent years. This optimistic enthusiasm is rooted in the growing global government and private sector commitment to address this issue [15,18,20]. However, there is also a concern due to the difficult task posed by Gram-negative bacterial antibiotic resistance in clinical settings, as Gram-negative bacteria are more difficult to treat because they have an outer membrane that makes it more challenging for antibiotics to penetrate and kill them [21]. Unfortunately, no new antibiotics/antimicrobials for Gram-negative bacteria have been discovered and used in recent decades [15,22,23]. This lack of progress in discovering and developing new antibiotics for Gram-negative bacteria is a significant concern, as there is a continuous evolution of resistant Gram-negative infections caused by bacteria such as carbapenem-resistant Enterobacteriaceae, multidrug-resistant Pseudomonas aeruginosa, Klebsiella pneumoniae, Staphylococcus aureus, Neisseria gonorrhoeae, and Escherichia coli. These infections are becoming increasingly difficult to treat and a major concern [8,9,10,11,12,15,18,19,24,25,26,27].
1.1. Membrane Proteins of Gram-Negative Bacterium Species
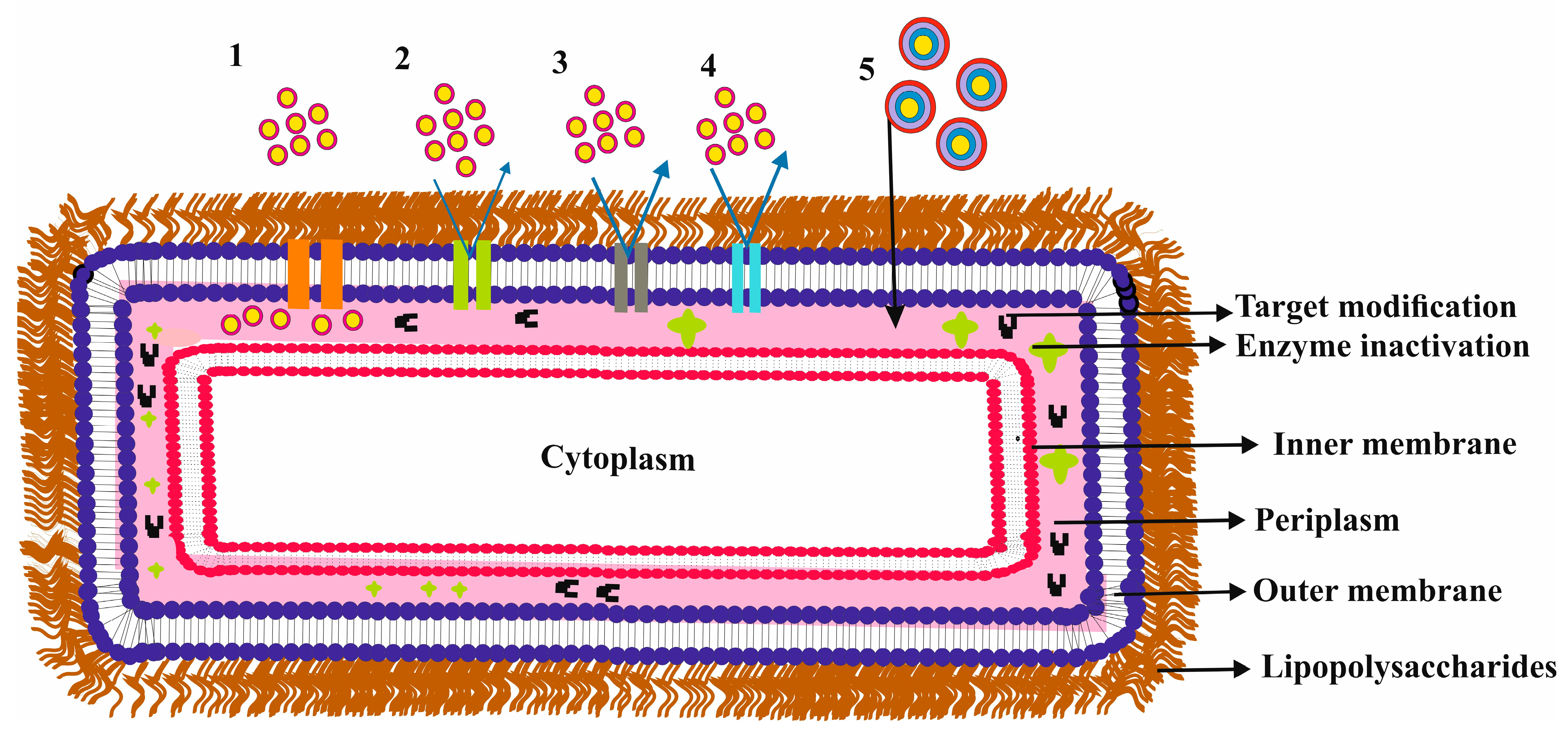
Gram-negative bacteria species are well-known and responsible for causing significant harm to mankind, making them one of the most notorious threats [28]. These bugs have an intricate cell cover with an outer membrane that acts as a molecular screen, forming an able selective permeation impediment. Like other biological membranes, this outer membrane, “see in Figure 1”, is mainly made of a bilayer of lipids, making it impermeable to hydrophilic molecules [7,15,21,24,29,30]. Certain water-filled membrane proteins primarily regulate the adequate inflow of these molecules. “see Table 1 for different membrane proteins studied across different research experimental studies and Table 2 for different membrane proteins classifications”.
These membrane proteins are found in various structures with different substrate specificities, regulations, and expressions [6,21,30]. They mostly form hollow beta-barrel structures with a hydrophobic outer surface [31]. The barrel structure makes a transmembrane pore that permits the passive flux of hydrophilic substances and acts as a functional diffusional block for nonpolar molecules/solutes [6,7]. These proteins also exhibit particular ion-selectivity for either cations or anions [32,33,34]. Bacteria can alter/modify these proteins to lower or inhibit influx through membrane channels, which, further along with efflux machinery systems, contributes towards antibiotic resistance [6,7,24,31,35]. An exhilarating task for drug design strategy is understanding membrane permeability at the molecular level to understand membrane transport’s starring role in bacterial resistance processes [15]. Antibiotics, such as penicillin, carbapenems, cephalosporins, fluoroquinolones, etc., enter Gram-negative bacteria mainly through membrane channels [29,36,37,38,39]. Every slight alteration by the bacteria of these membrane proteins can considerably affect antibiotic drug therapy [24]. Many clinically significant bacterial species have been sequenced to determine the key membrane channels in the Gram-negative bacterial outer membrane [6,7].
This reduced permeability might result from fewer membrane channels in the outer membrane and their distinct physical and chemical properties [17,24]. These channels have different structures and specificities but have been reported to form a diffusional barrier for nonpolar solutes and antibiotic molecules [7,15,24,29]. Bacteria can adapt and reduce influx through these membrane channels, leading to antibiotic resistance [7,15,24,29]. Thus, the study of membrane permeability is crucial in realizing the role of membrane transport in antibiotic resistance mechanisms. Research has shown that small polar molecules roughly below 600 Dalton have the highest activity critical towards Gram-negative bacteria, suggesting that porins may be the primary entry path [15,24,29,37,40]. However, low antibiotic permeability via the outer membrane requires high doses of antibiotics, which can result in toxic side effects.
Antibiotic uptake and antibiotic efficacy are complex phenomena that are influenced by various factors in bacteria [7,15,24,29,41,42]. While the outer membrane barrier of Gram-negative bacteria is a significant obstacle to the entry of antibiotics, small hydrophilic solutes are then facilitated through the porin or outer membrane protein pathways [6,15,17,24,29,35,43,44]. See Figure 1 for an overview. The cytoplasm and inner membrane also have mechanisms to efflux toxic compounds, reducing the dose at the site of action [29]. Overall, antibiotic resistance mechanisms include enzymes that inactivate antibiotics, changes in the bacterial outer membrane that decrease entry, and modifications in target proteins within the cell [27,45].
To overcome these defense mechanisms, it is crucial to recognize the mechanisms of antibiotic uptake, resistance, and delivery [27,45]. There is a need for more quantitative data on antibiotic uptake, especially for various compartments of bacteria, and innovative ways to improve the delivery and efficacy of antibiotics [24,29,45]. The outer membrane of bacteria is a crucial barrier that determines the organism’s susceptibility to antibiotics [24,29,45]. Structural changes to the outer membrane proteins can lead to antibiotic resistance, particularly when combined with beta-lactamases in the periplasmic space [10,46]. It is essential to study the transport mechanisms involved in membrane permeability to combat this issue, as this knowledge will inform the design of effective antibiotics. [24]. To this end, various membrane channels have been studied using different methodologies, and the results have been compiled in Table 2 to better understand the role of membrane channels in outer membrane permeability and their interaction with various antibiotics.
The central aim of this assessment is to expand upon the author’s prior [27,45] work by analyzing and summarizing the advancements made in comprehending the role of various outer membrane proteins in Gram-negative bacteria, which facilitate the influx of solutes and antibiotics. Furthermore, this review consolidates multiple research studies examining antibiotics and outer membrane proteins in diverse Gram-negative bacterial strains. The knowledge presented herein may be utilized to develop new antibiotics.
Understanding how antibiotics penetrate the cell wall of Gram-negative bacteria is a significant hurdle in the creation of novel drugs [15,24,35]. This understanding is critical towards developing effective antibiotics, and interpreting the molecular core of outer membrane penetrability is still the current challenge [6,7,15,24,29,35].
This review endeavors to compile state-of-the-art knowledge relevant to outer membrane proteins in Gram-negative bacteria and their crucial role in antibiotic influx. Ultimately, this study aims to stimulate a discussion about exploring and comprehending physicochemical parameters that govern antibiotic translocation via bacterial membrane proteins.

{kind=link}
{kind=link}
Table 1.
The tabulated different research studies investigated involving different membrane proteins from various species of Gram-negative bacteria, building upon and updating earlier published research by the author [27,45]: molecular Simulations (MS), outer membrane protein (Omp). The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
Table 1.
The tabulated different research studies investigated involving different membrane proteins from various species of Gram-negative bacteria, building upon and updating earlier published research by the author [27,45]: molecular Simulations (MS), outer membrane protein (Omp). The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
| Species | Concluding Analysis | Membrane Proteins/Channels/Porins |
|---|---|---|
| Escherichia coli | Showed the structure and function of porin. The work focused on the annulus of basic and acidic patches. Showed the effect of mutations in the annulus to affect ion selectivity and sugar transport. These findings provide insights into the mechanisms underlying the permeation of molecules through bacterial porins [47]. | OmpG |
| Escherichia coli | Examined and showed effects of Loop L3 from OmpF membrane protein dynamics toward antibiotic translocation [48]. | OmpF |
| Escherichia coli | Explored and showed the mechanism of TolC–AcrA complex formation [49]. * E. coli AcrAB-TolC is the ternary efflux pump, implicated in drug resistance by removing antimicrobials/antibiotics out of the cell. TolC obstruction by numerous cations, for example, hexaamminecobalt and others, have been reported and therefore signifies a prospective target for reducing antibiotic resistance [50]. | TolC * |
| Escherichia coli | This study presents an innovative approach for investigating the role of membrane channels in antibiotic enrofloxacin, kanamycin sulfate, norfloxacin, and ciprofloxacin uptake in Gram-negative bacteria. The researchers were able to characterize membrane protein channels in their native environment [51]. | OmpF, OmpC |
| Escherichia coli | This study presents a novel approach to distinguish between translocation and binding processes in biological channels using an additional barrier at the channel exit. By functionalizing the channel with covalent blockers, they were able to identify the successful permeation of model peptides and an antibiotic through the well-studied outer membrane channel [52]. | OmpF |
| Escherichia coli | Reported the interface of gentamicin, kanamycin, and polymyxin B with purified lipopolysaccharide from the outer membrane [53]. | OmpF |
| Escherichia coli | Reported ampicillin, benzylpenicillin permeation, and the effect of ampicillin degradation on interaction with channel [54]. | OmpF |
| Escherichia coli | This study demonstrates an improved method for analyzing small molecule penetration and permeation through nanopores. The analysis of the single-channel ion current of meropenem interacting with porin suggests sub-microsecond-scale gating kinetic parameters are accessible with existing experimental equipment [55]. | OmpF |
| Escherichia coli | Reported the consequences and impacts of access resistance on membrane proteins [56]. | OmpF |
| Escherichia coli | Reported the transportation of divalent ions and its effect on pore conductance and selectivity [57]. | OmpF |
| Escherichia coli | Reported the impact of diverse divalent cation salts on channel conductance, the function of electrolytes and the accumulation of counterions caused by channel charges, and other effects not present in monovalent cation salts [58]. | OmpF |
| Escherichia coli | Explored the transport of different aminoglycosides through porins [39]. | OmpF, OmpC, LamB, ChiP |
| Escherichia coli | Reported the influence of divalent cationic ions concerning the pH sensitivity of membrane channel via causing the pK(a) shift of chief acidic residues [59]. | OmpF |
| Escherichia coli | Reported the process of selectivity inversion in the membrane protein [60]. | OmpF |
| Escherichia coli | Reported insights into the diffusion route of ciprofloxacin across the outer membrane porin, which is crucial for developing effective antibiotics against Gram-negative bacteria [61]. | OmpC |
| Escherichia coli | Reported the recombinant type of the membrane channel and revealed the monomeric nature of the membrane channel [62]. | OmpG |
| Escherichia coli | Reported crystal structure (X-ray) of the membrane channel [63]. | OmpG |
| Escherichia coli | Reported membrane channel crystal structure in different dimensions [64]. | OmpG |
| Escherichia coli | Reported the folding mechanism of the membrane channel in detergent solution [65]. | OmpG |
| Escherichia coliAcinetobacter baumannii Porphyromonas asaccharolytica Photobacterium damsela Serratia liquefaciens | The reported structural configuration of various membrane proteins and measured penetration rates for different β-lactams ampicillin, cephalexin, cefuroxime, cefotaxime, ceftazidime, and ceftriaxone [46]. | OmpA, Omp-ECB HMP-AB HMP-PA Omp-PD Omp-SL |
| Escherichia coli | Reported different binding regions of membrane proteins [66]. | OmpA |
| Escherichia coli | Reported the function of outer membrane proteins in stress survival [67]. | OmpA |
| Escherichia coli | Reported the crystal structure of membrane proteins and further explained possible virulence mechanisms [68]. | OmpX |
| Escherichia coli | Explored the outer membrane protein behavior and explained the impact of channel expansion [69]. | FhuA |
| Escherichia coli | Showed transfer of DNA via membrane proteins [70]. | FhuA |
| Escherichia coli | Examined structural parameters of membrane proteins [71]. | FhuA |
| Escherichia coli | Reported different structural elements and the interface of proteins and protein subdomains also exhibited the role of the membrane proteins in uptake [72]. | FhuA, |
| Escherichia coli | Studied and demonstrated effective transport of the ferrichrome complex through the membrane [73]. | FhuA |
| Escherichia coli | Studied and demonstrated antibiotic uptake (norfloxacin, ciprofloxacin, and enoxacin) and showed the electroosmotic flow/electrophoretic force’s involvement in transport/translocation [74]. | OmpF |
| Escherichia coli | Studied kanamycin uptake effectively facilitated by membrane channels [75]. | OmpF, OmpC |
| Escherichia coli | Studied fosfomycin transport and uptake via outer membrane porin [76]. | OmpF |
| Escherichia coli | Demonstrated the movement of cephalosporin through the channels [38]. | OmpF, OmpC |
| Escherichia coli | Studied and showed for the first time and determined the transport of charged molecules sulbactam/tazobactam/avibactam β-lactamase inhibitors [40]. | OmpF |
| Escherichia coli | Measured transport for the first-time using membrane pore innate ion selectivity, employed for charged inhibitors/molecules, sulbactam/tazobactam/avibactam β-lactamase inhibitors, respectively, using novel electrophysiology zero-current/reversal-potential assay [37]. | OmpC |
| Escherichia coli | Examined the permeability of carbapenems through membrane channels from distinctive clinical isolates [77]. | OmpC |
| Escherichia coli | Showed quantification of fluoroquinolone norfloxacin transport/uptake via membrane pore [36]. | OmpF |
| Escherichia coli | Showed the process of fluoroquinolone enrofloxacin molecule transport [24]. | OmpF |
| Escherichia coli | Evaluated the uptake of cefepime, ceftazidime, and imipenem [78]. | OmpF |
| Escherichia coli | Showed meropenem interaction with the channel and elaborated the binding activity [79]. | OmpF |
| Escherichia coli | Reported translocation of polypeptides through membrane protein [80]. | OmpF |
| Escherichia coli | Reported enrofloxacin’s effective interaction across the channel and effective modulation of the affinity site in the existence of magnesium ion [81,82,83]. | OmpF |
| Escherichia coli | Reported ampicillin and benzyl-penicillin uptake and the effective mechanism involved in the uptake across the outer membrane [84]. | OmpF |
| Escherichia coli | Described and explained the charged residues role in membrane channel constriction implicating, channel conductance, ionic selectivity, and voltage-gating [85]. | OmpF |
| Escherichia coli | Reported involvement of electrostatic interactions amongst ions and charge distribution in the interior of the channel regulating effective ion permeation and selectivity [32]. | OmpF |
| Escherichia coli | Reported different amino acid residues ionization states and evaluated self-consistently electric potential distribution within the channel [86]. | OmpF |
| Escherichia coli | Explained particular ampicillin molecule movement within the membrane pore [87]. | OmpF |
| Escherichia coli | Demonstrated cation selectivity within membrane porin/protein by identifying the impact of alkali-metal ions atomic-radii on the site affinity binding [88]. | OmpF |
| Escherichia coli | Described particular interaction stuck between grepafloxacin, ciprofloxacin, nalidixic acid, and moxifloxacin with membrane protein [89]. | OmpF |
| Escherichia coli | Explained active permeation of moxifloxacin across membrane protein and showed protein/antibiotic interaction [90]. | OmpF |
| Escherichia coli | The effects of four types of polyamines (spermidine, putrescine, cadaverine, and spermine) on membrane protein activity were demonstrated and explained [91]. | OmpF |
| Escherichia coli | The functional characteristics of four individual amino acid substitutions and the impact of a deletion mutant in constriction loop L3 on specific channels were demonstrated and explained [92,93]. | OmpF |
| Escherichia coli | The important role of the constriction loop in regulating the voltage-dependent gating of intrinsic membrane proteins was demonstrated and explained [94]. | OmpF |
| Escherichia coli | The interactions between nalidixic acid and moxifloxacin antibiotics with bacterial proteins were demonstrated and explained [95]. | OmpF |
| Escherichia coli | Demonstrated and explained effective ampicillin translocation across the membrane protein. Further illustrated the impact of mutations within pores impacting molecular passage [96]. | OmpF |
| Escherichia coli | Demonstrated ion pathways along the channel surface [97]. | OmpF |
| Escherichia coli | Explained the interface of particular (HPA3P and magainin 2) peptides with the membrane protein and the impact of electric field on pore and peptide geometry [98]. | OmpF |
| Escherichia coli | Reported a particular mechanism of translocation of ampicillin via membrane protein [99]. | OmpF |
| Escherichia coli | Reported particular interaction/binding of carbenicillin, and ampicillin ertapenem, within the membrane protein [100]. | OmpF |
| Escherichia coli | Explained specific interactions engaged in the transport/translocation of amoxicillin, ampicillin, azlocillin, carbenicillin, and piperacillin [44]. | OmpF |
| Escherichia coli | Explained the influence of particular acid residue (D113A) substitution on ceftazidime, cefotaxime, cefpirome, cefepime, cefoxitin, and ampicillin bacterial susceptibility [101]. | OmpF |
| Escherichia coli | Reported the operational anti-loop 3 residue (Lys-16) position in cefepime transport across membrane protein [102]. | OmpF |
| Escherichia coli | Explained ionic concentration and charged residues in membrane protein present at constriction zone and their impact over intrinsic gating behavior of membrane protein [103]. | OmpC |
| Escherichia coli | Explained trio mutations in the interior of isolated porins from multi-drug resistant bacterium. Further, demonstrated the resulting mutations affecting the transport of cefotaxime [104]. | OmpC |
| Escherichia coli | Demonstrated and explained effective interaction of ceftriaxone, cefpirome, and ceftazidime with membrane protein [105]. | OmpC, OmpF |
| Escherichia coli | Reported an effective influx of cefepime, enrofloxacin, norfloxacin, ciprofloxacin, ceftazidime, and ceftriaxone [106]. | OmpC, OmpF |
| Escherichia coli | Demonstrated the impact of culture medium over effective membrane protein expression and showed the effect on tazobactam-piperacillin susceptibility [107]. | OmpC, OmpF |
| Escherichia coli | Demonstrated by exploiting water as an analytical tool, a distinctive macroscopic electric field within water-filled channels [108]. | OmpF, OmpC |
| Escherichia coli | The successful penetration of meropenem and imipenem was demonstrated and explained. It was also shown that their uptake relies on the alignment of the molecule’s electric dipole with the membrane protein’s interior electric field. Moreover, certain regions within the membrane protein were identified as preorientation sites that affect the pathway of the molecules [109]. | OmpF, OmpC |
| Escherichia coli | Demonstrated and explained excess of positively charged fixed ions within the interior of the membrane protein. As a result, the protein exhibits a characteristic selectivity towards anions [110]. | PhoE |
| Escherichia coli | The influence of lysine and carboxyl groups on the inherent ionic selectivity of the membrane pore was demonstrated and explained [111]. | PhoE |
| Enterobacter aerogenes, Enterobacter cloacae Klebsiella pneumoniae | Demonstrated and explained the transport of different antibiotic membrane proteins [23]. | (Omp35, Omp36), (OmpE35, OmpE36), (OmpK35 OmpK36) |
| Klebsiella pneumoniae, Escherichia coli | Demonstrated possible system of antibiotic resistance by mutational loss of membrane proteins. Further, effectively demonstrated quantitative influx rates for different antibiotics such as erythromycin, novobiocin, cloxacillin, oxacillin, benzylpenicillin, ampicillin, cephaloridine, cefoxitin, cephalothin, ceftazidime, cefepime, cefotaxime, cefamandole, imipenem, ceftriaxone and ertapenem [112]. | OmpK35/OmpK36 OmpF, OmpC |
| Enterobacter aero-genesE. Aerogenes, Klebsiella pneumoniae, Escherichia coli | Demonstrated outer membrane porins role over avibactam-ceftazidime diffusion via outer membrane [113]. | Omp35, Omp36 OmpK35, OmpK36 OmpF, OmpC |
| Enterobacter aerogenes, | The mechanism resulting in imipenem resistance by reducing outer membrane permeability was demonstrated and explained for distinct resistant clinical isolated strains [114,115]. | Omp36, Omp35 |
| Enterobacter aerogenes, | The reported effective function of membrane channel towards susceptibility in relation to ceftriaxone [116]. | Omp35 |
| Enterobacter aerogenes, | Reported the impact of porin over the uptake/influx of cefepime and ertapenem [117]. | Omp36 |
| Enterobacter aerogenes, | Reported the process of resistance to be appropriate to membrane channel mutation, affecting cefepime, imipenem, and cefpirome permeability in clinical strains [118]. | Omp36 |
| Providencia stuartii | Explained different β-lactam molecules cefoxitin, cefepime, and ertapenem interactions [119]. | OmpPst1 and OmpPst2 |
| Providencia stuartii | Demonstrated and explained membrane protein’s structure surface-exposed loop function and specific interaction with diverse membrane modules (e.g., lipopolysaccharides) [120]. | OmpPst1 and OmpPst2 |
| Providencia stuartii | Showed and explained the role of membrane proteins in carbapenem transport across the outer membrane [121]. | OmpPst1 |
| Klebsiella pneumoniae | Demonstrated and explained the effect of outer membrane proteins over bacterial resistance towards tigecycline, avibactam ceftazidime combination, along with colistin in clinical strains [122] | OmpK36, OmpK35 |
| Pseudomonas aeruginosa | Demonstrated and explained the process involved in the effective uptake of fosmidomycin across phosphate-specific proteins/transporters [123]. | OprO, OprP |
| Pseudomonas aeruginosa | Demonstrated and explained single-channel characteristics of protein [124]. | Occk8 |
| Pseudomonas aeruginosa | Demonstrated and explained membrane protein activity, broad range conductance, gating dynamics, and cation selectivity for outer membrane proteins [33]. | OccD1–OccD6 |
| Pseudomonas aeruginosa | The uptake of outer membrane proteins was demonstrated and described, highlighting the contact between carboxylate groups and central residues of the basic ladder (lysine and arginine) residues [125]. | OccD1–OccD6, OccK1–OccK7 |
| Pseudomonas aeruginosa | Channel activity was demonstrated and explained, including the conductance, gating transitions, different channel sub-states (K3 for one-open sub-state, K4–K6 for two-open sub-state, and K1, K2, K7 for three-open sub-state kinetics). Additionally, the study revealed the channel’s anion selectivity and identified the positive residues within the central constriction of the membrane proteins [126]. | OccK1–OccK7 (K1–K7) |
| Pseudomonas aeruginosa | Showed gating physical characteristics, conductance, and the consequent impression of inner constriction loop omission on gating transitions [127]. | OccK1 |
| Pseudomonas aeruginosa | The gating dynamics, including enthalpy-driven and entropy-driven current transitions, were demonstrated and described. Additionally, the effect of loop deletion on activation enthalpies and entropies over channel transitions was explained [128]. | OccK1 |
| Pseudomonas aeruginosa | Reported effect of concentrations of ions on channel gating transitions [129]. | OccK1 |
| Pseudomonas aeruginosa | Demonstrated and explained the effects of structural insight toward substrate specificity. Further also showed channel composition with monomeric 18-stranded beta-barrel consequent to slender constriction inside the pore [130]. | OccD1 |
| Pseudomonas aeruginosa | Demonstrated and explained the imipenem passage through the pore and the position of particular surface loop areas in the pore establishing passage [131]. | OccD1 |
| Pseudomonas aeruginosa | Showed and explained amino acid substrates uptake/translocation in-order to recognize the structure and dynamics of pore [132]. | OccD1 |
| Pseudomonas aeruginosa | Showed and explained imipenem and meropenem uptake [42]. | OccD3 |
| Pseudomonas aeruginosa | Showed and explained membrane protein’s role in citrate, isocitrate, and tricarboxylate uptake [133]. | OccK5 |
| Pseudomonas aeruginosa | Showed and explained the different distinct gating mechanisms of the channel [134]. | OccK5 |
| Pseudomonas aeruginosa | Examined and described the contribution of the membrane proteins in the transport of temocillin [135]. | OccK1, OccK2 |
| Pseudomonas aeruginosa | The ionic selectivity of the phosphate-specific pore was investigated and elucidated. The energetic properties, particularly for phosphate, chloride, potassium, and sulfate ions, were determined [34,136]. | OprP |
| Pseudomonas aeruginosa | Explored, confirmed, and elucidated the significance of negatively charged residue (D94) in central binding for specific phosphate binding and selectivity [137]. | OprP |
| Pseudomonas aeruginosa | The role of the central arginine (R133) in determining channel selectivity and ion transport properties has been investigated and established [138]. | OprP |
| Pseudomonas aeruginosa | The impact of important residues on channel constriction and their influence on the substrate specificity of the membrane protein has been investigated [139]. | OprP and OprO |
| Pseudomonas aeruginosa | Demonstrated declined membrane protein production to be only of the causative considerations for carbapenem hetero-resistance [140]. | OprD |
| Pseudomonas aeruginosa | The role of membrane proteins in increasing the minimal inhibitory concentrations of carbapenems in clinical isolates has been investigated and confirmed [141]. | OprD |
| Pseudomonas aeruginosa | Examined and recognized membrane protein levels in distinct carbapenem-resistant isolates [142]. | OprD |
| Pseudomonas aeruginosa | Recognized conflicting susceptibility of carbapenem together with alterations in outer membrane permeability [143]. | OprD |
| Pseudomonas aeruginosa | The in vitro activity of ceftolozane-tazobactam and ceftazidime-avibactam against meropenem-resistant bacterial isolates was examined and confirmed [144]. | OprD |
| Pseudomonas aeruginosa | Examined and recognized distinctive membrane proteins in-frame deletions amongst particular clinical isolates [145]. | OprD |
| Pseudomonas aeruginosa | Researched distinct adaptations of membrane proteins domination in -isolated resistant stains, particularly towards imipenem [146]. | OprD |
| Pseudomonas aeruginosa | Investigated the structure-porin permeation relationships of a group of novel carbapenem analogs across the membrane protein [147]. | OprD |
| Pseudomonas aeruginosa | The influence of membrane protein polymorphisms, particularly amino acid substitutions or replacements at codon 170, on carbapenem resistance was examined [148]. | OprD |
| Pseudomonas aeruginosa | Investigated the influence of single swaps of amino acid in outer membrane proteins on carbapenem resistance strains [149]. | OprD |
| Pseudomonas aeruginosa | The study examined and uncovered that debilitating mutations and decreased expression of outer membrane proteins are among the factors that contribute to resistance against imipenem and meropenem [150]. | OprD |
| Pseudomonas aeruginosa | Showed the existence of membrane proteins in distinct (seventy) clinical bacterial isolates resistant to carbapenem [151]. | OprD |
| Pseudomonas aeruginosa | The study’s results demonstrated the effects of double mutations, which resulted in an interchange of phosphate and diphosphate specificities of membrane proteins, and were evaluated [152]. | OprP, OprO |
| Pseudomonas aeruginosa | Demonstrated organizational components accountable for the conveyance of (amino acid residues) via substrate-specific channels [153]. | OccK8 |
| Acinetobacter baumannii | The study investigated outer membrane proteins’ channel-forming properties and other physicochemical attributes [154]. | CarO and Omp25 |
| Acinetobacter baumannii | Identified different antibiotic scaffolds [22]. | DcaP |
| Acinetobacter baumannii | Analyzed uptake of L-ornithine through outer membrane proteins and exhibited L-ornithine influence over pathogen sensitivity in relation to imipenem [155]. | CarO |
| Acinetobacter baumannii | Demonstrated membrane protein uptake intended for ornithine and glycine and nil uptake in support of glucose, glutamic acid, and imipenem via membrane channel [156]. | CarO isoforms CarO1, CarO2, CarO3 |
| Acinetobacter baumannii | Reported channel conductance, identifiable cationic selectivity, and particular specificity regarding meropenem, imipenem glutamic acid, and arginine [157]. | CarO |
| Acinetobacter baumannii | The study demonstrated the imperative function and role of membrane proteins in relation to imipenem, meropenem, colistin, ceftazidime, and ciprofloxacin [158]. | rOprD homologue |
| Acinetobacter baumannii | Showed membrane proteins specificities towards substrate arginine, glutamic acid, glycine, ornithine, maltose, putrescine, glucose, phenylalanine, tryptophan, benzoic acid, ampicillin, meropenem, imipenem, ceftazidime, and fosfomycin [159]. | OccAB1–OccAB4 |
| Campylobacter jejuni | Analyzed and demonstrated imperative properties of the membrane protein [160]. | MOMP |
| Campylobacter jejuni | Analyzed and demonstrated the imperative three-dimensional structure of outer membrane proteins and further elucidated the underlying molecular mechanisms [161]. | MOMP |
| Campylobacter jejuni | Analyzed crucial sequence polymorphism and revealed the membrane protein’s secondary structures and surface-exposed conformational epitopes [162]. | MOMP |
| Campylobacter jejuni | The study elucidated the critical channel-forming properties of membrane proteins as both trimers and monomers and the transition from trimer to monomer [163]. | MOMP |
1.2. Computing Influx
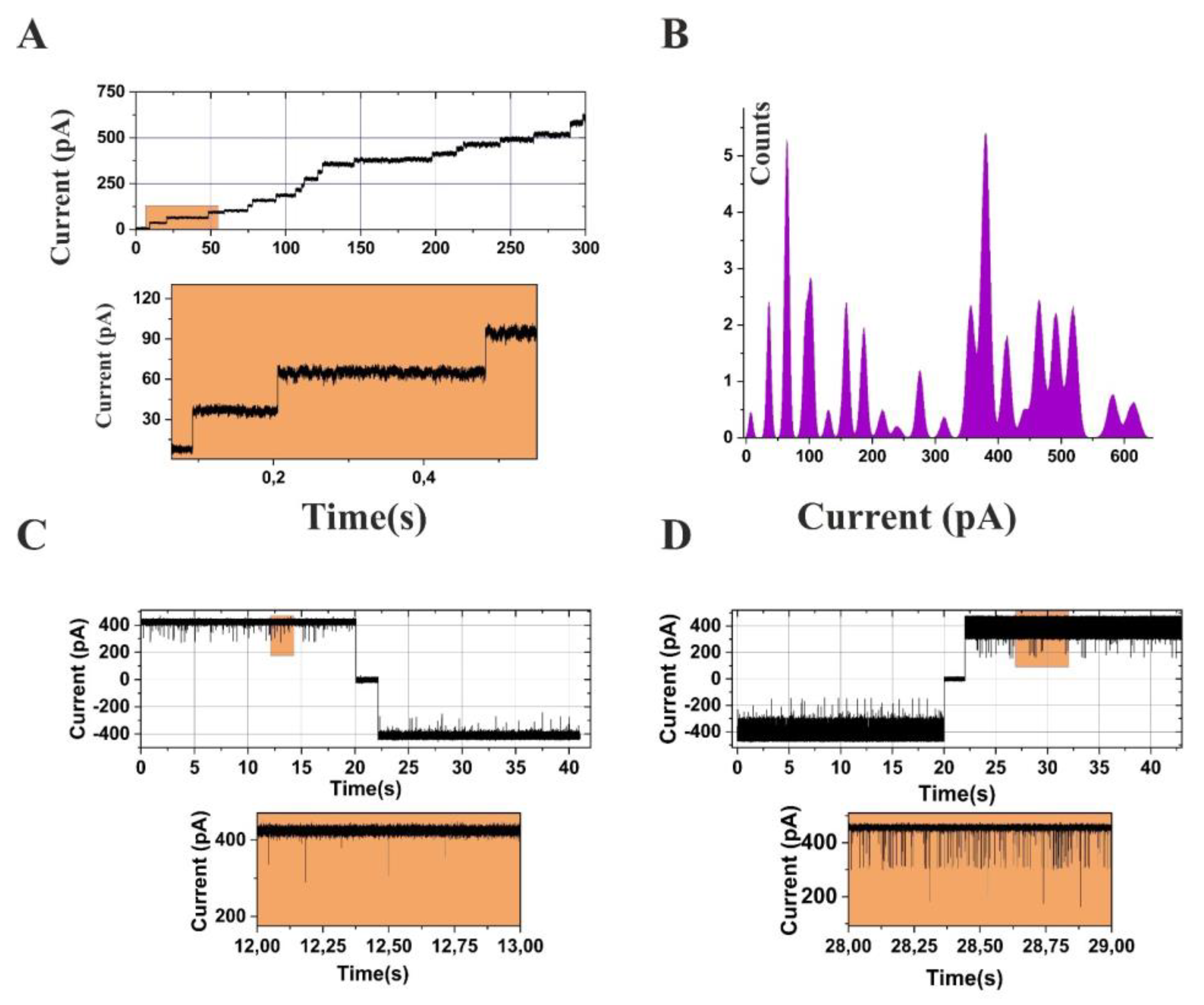
The creation of specific inhibitors for emerging Gram-negative bacterial targets is a vital element in treating bacterial infections. Nonetheless, a significant hurdle in achieving this goal is the low permeability of antibiotics and the need for high doses to reach the intended target. This can result in off-target effects. To address this challenge, a thorough understanding and quantification of the permeability barrier of Gram-negative bacteria are imperative [7,24,29,36,37]. Measuring the uptake of antibiotics across the cell envelope of Gram-negative bacteria is a complicated process [24,29]. Various experimental techniques have been employed to quantify this process, such as mass spectrometry and fluorescence, which measure uptake at a cellular level, and electrophysiology, which measures uptake at a single-channel stage (see Figure 2). For antibiotics with channel-limited permeation, the flux across a single OmpF or OmpC channel has been measured, with the results indicating uptake rates varying from one to a few hundred molecules per second [37,38,39,40,54,164]. However, the lack of appropriate direct assays to measure permeability presents a significant challenge in understanding this barrier. To assist researchers, critical information has compiled in Table 2, including data from different studies involving various membrane proteins from different Gram-negative bacterial species. A larger dataset of single-channel permeabilities under various conditions would be a powerful tool. Additionally, real-time flux rate data across different membrane proteins measured using reversal potential measurements have compiled in Table 3 [38,39,153,164,165].
Further, this electrophysiology assay offers an essential methodology for quantifying the flux of charged antibiotics and antimicrobials through the bacterial cell membrane proteins [37,40]. This methodology entails the incorporation of one or more membrane channels into classical experiments utilizing an artificial planar lipid bilayer and utilizing the transmembrane potential-driven ion current as a detection probe [37,38,39,40,54,153,164,165]. An overview of this methodology is provided in Figure 2B. The electrophysiological characteristics of the membrane channels, such as their size, single-channel conductance, channel ion selectivity, and channel gating dynamics, have been well-characterized and provide valuable information for comprehending the transport of small molecules across the bacterial cell membrane [37,40]. Moreover, the size of the membrane channels plays a crucial role in transporting molecules through the channel and, thus, in antibiotic susceptibility.
Developing antibiotics that effectively target Gram-negative bacteria is a complex and challenging undertaking due to the multi-layered cell envelope and the various obstacles active compounds must overcome. Understanding and quantifying the permeability barrier is vital for designing and developing new antibiotics, and compiling experimental data, including electrophysiology measurements, can play a critical role in this process. Figure 2A,B depicts how membrane channels get inserted into a planar lipid bilayer. Figure 2C,D clearly distinguishes when a channel interacts with a particular compound. Please be advised that Figure 2A–D is a unique representation created solely for explanatory purposes. For a comprehensive understanding of the research, kindly refer to the cited sources [40,54].
Furthermore, the sizing of membrane channels using electrophysiology can provide insights into the maximum size of the molecule it can transport and help evaluate the inner structure, including the constriction site [24,69,79,80,81,83]. In addition, the electrophysiological technique enables the real-time quantification of the transport rate of individual molecules, which is a potent approach for exploring the kinetics of the transport process [15,83]. This methodology is not constrained to antibiotic transport but can also be extended to investigating the transport of other small molecules, such as sugars and amino acids [15,83,117]. In conclusion, assessing the flux of small ions/molecules across the bacterial cell membrane is pivotal for comprehending the mechanisms of antibiotic uptake [43,44,88].
Additionally, Molecular Simulations (MS) have emerged as a powerful tool to gain atomic-scale information on the translocation of antibiotics across the outer membrane of Gram-negative bacteria [24,40,79,153]. These simulations offer valuable insights into potential transport mechanisms, including diffusion with molecule binding, dehydration-induced pore, and slow diffusion with molecule binding. However, it is essential to acknowledge that MS is only one aspect of the complex puzzle of understanding antibiotic transport. In addition, MS typically investigates transport under highly controlled conditions that may not perfectly reflect the in vivo environment’s complexity [24,40,79,153]. Nonetheless, MS has contributed significantly to understanding antibiotic transport and identifying potential targets for overcoming resistance mechanisms and enhancing antibiotic permeation [6,24,40,76,79,117,153]. Furthermore, by providing information on the dynamic and structural properties of Membrane channels, MS can aid in designing new antibiotics with improved transport properties. Given the continuing challenge of antibiotic resistance, using MS and other techniques will be crucial in discovering new approaches to combat bacterial infections [6,15,24,36,40,76,77,78,79,117,153].
Figure 2.
Electrophysiology experiment overview (A) Current records from a bilayer in symmetric salt solution at t = 0. After 300 s., Stepwise channel insertion into the bilayer. (B) Current amplitude histogram of the ion current trace (A). (C) Current recordings from a bilayer containing single membrane OmpF channel with Ampicillin. (D) Current recordings from a bilayer containing single membrane OmpF channel with degraded Ampicillin. Original figures by author. The figures are only for representations, not experimental data explained here. For the experimental explanation for (A,B), please see references [69]; for (C,D), see reference [54].
Figure 2.
Electrophysiology experiment overview (A) Current records from a bilayer in symmetric salt solution at t = 0. After 300 s., Stepwise channel insertion into the bilayer. (B) Current amplitude histogram of the ion current trace (A). (C) Current recordings from a bilayer containing single membrane OmpF channel with Ampicillin. (D) Current recordings from a bilayer containing single membrane OmpF channel with degraded Ampicillin. Original figures by author. The figures are only for representations, not experimental data explained here. For the experimental explanation for (A,B), please see references [69]; for (C,D), see reference [54].

Table 2.
Crucial Membrane Proteins and antibiotic membrane protein (Omps) combinations were studied in different Species of Gram-negative bacteria, building upon and updating earlier published research by the author [27,45]. The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
Table 2.
Crucial Membrane Proteins and antibiotic membrane protein (Omps) combinations were studied in different Species of Gram-negative bacteria, building upon and updating earlier published research by the author [27,45]. The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
| Species | Membrane Proteins (Omp) | Reported Antibiotic Study with (Omps) |
|---|---|---|
| Porphyromonase asaccharolytica | HMP-PA [46] |
|
| Photobacterium damsela | Omp-PD [46] |
|
| Serratia liquefaciens | Omp-SL [46] |
|
| Acinetobacter baumannii | (OccAB1-OccAB5) [159], rOprD [158], CarO [156,157], Omp25 [154].HMP-AB [46]DcaP [22] |
|
| Escherichia coli | OmpX [68], OmpA [66,67,166], OmpT [167], Tsx [168], FadL [169], OmpF [26,27,39,56,170], OmpC [103,104,105,106,107,108,109,112,113], PhoE [171], LamB [172,173], BtuB [174], FepA [175], FhuA [5,176]OmpC [39,103,104,105,106,107,108,109,112,113], OmpF [24,32,36,37,40,44,77,78,79,80,81,82,83,84,85,86,87,88,89,90,95,96,97,98,99,100,101,102,105,106,107,108,109,112,113], PhoE [110,111]ChiP [39] |
|
| Enterobacter aerogenes | Omp36 [112,114,115,117,118], Omp35 [27,112,114,115,116] | |
| Enterobacter cloacae | OmpE36 [178], OmpE35 [27] |
|
| Klebsiella pneumoniae | OmpK36 [112,113,122], OmpK35 [27,112,113,122] | |
| Providencia stuartii | OmpPst1 [119,120], OmpPst2 [119] | |
| Pseudomonas aeruginosa | (OpdC/OccD2, OprDOccD1, OpdP/OccD3, OpdB/OccD7, OpdI/OccD5, OprQ/OccD6, OpdT/OccD4, OpdJ/OccD8) [27,33,34,125,126,130,131,132,179] OpdF/OccK2, OpdO/OccK3, OpdH/OccK5, OpdQ/OccK6, OpdL/OccK4, OpdK/OccK1, OpdD/OccK7, OprE/OccK8/[27,96,126,127,128,129,133,134,135,153,179], OprP [34,136,137,138,152], OprO [152] |
Table 3.
Reported turnover rates molecules/second at a given concentration gradient for different outer membrane proteins (Omps) with different substrates. The values are approximate, and for detailed calculations, please see the research publications mentioned with each value [26,27,39,54,153]. Substrates are shown in one column on the right, followed by reported porins, Gram-negative bacterial species, recalculated flux rates molecules/second at 1 μM for comparison purposes only, approximate values, and Reported Flux Molecules/second at specifically mentioned gradient. The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
Table 3.
Reported turnover rates molecules/second at a given concentration gradient for different outer membrane proteins (Omps) with different substrates. The values are approximate, and for detailed calculations, please see the research publications mentioned with each value [26,27,39,54,153]. Substrates are shown in one column on the right, followed by reported porins, Gram-negative bacterial species, recalculated flux rates molecules/second at 1 μM for comparison purposes only, approximate values, and Reported Flux Molecules/second at specifically mentioned gradient. The author has endeavored to gather as much data as possible, and the citations provided are accurate to the best of the knowledge. However, any errors in the citations or table are beyond the author’s control. Additionally, it should be noted that while this Table is included, the list may not be limited.
| Molecules | Omps | Species | Recalculated Flux at 1 μM Molecules/Second | Reported Flux Molecules/Second at the Specific Mentioned Gradient |
|---|---|---|---|---|
| Avibactam | OmpF | E. coli | 62 | ≈620 at gradient10 μM [40] |
| OmpC | E. coli | 23 | ≈229 at gradient10 μM [37] | |
| Tazobactam | OmpF | E.coli | 20 | ≈200 at gradient10 μM [40] |
| OmpC | E. coli | 20 | ≈200 at gradient10 μM [37] | |
| Sulbactam | OmpF | E. coli | 20 | ≈200 at gradient10 μM [40] |
| OmpC | E. coli | 18 | ≈187 at gradient10 μM [37] | |
| Fosfomycin | OprO | P. aeruginosa | 28 | ≈280 at gradient10 μM [165] |
| OprP | P. aeruginosa | ≤1 | ≈2.2 at gradient10 μM [165] | |
| Ceftazidime | OmpF | E. coli | 34 | ≈1000 at gradient 30 μM [38] |
| OmpC | E. coli | 17 | ≈500 at gradient 30 μM [38] | |
| OprE | P. aeruginosa | ≤1 | ≈0.4 at gradient 10 μM [153] | |
| Cefotaxime | OprE | P. aeruginosa | ≤1 | ≈0.1 at gradient 10 μM [153] |
| Ampicillin | OmpF | E. coli | 24 | ≈237 at gradient10 μM [54] |
| Benzylpenicillin | OmpF | E. Coli | 12 | ≈120 at gradient 10 μM [54] |
| Carbenicilin | OprE | P. aeruginosa | ≤1 | 0.04 at gradient 10 μM [153] |
| Gentamicin | OmpF | E. Coli | 1.5 | ≈15 at gradient 10 μM [39] |
| OmpC | E. Coli | ≤1 | ≈8 at gradient 10 μM [39] | |
| LamB | E. Coli | ≤1 | ≤1 at gradient 10 μM [39] | |
| Chip | E. Coli | ≤1 | ≈3 at gradient 10 μM [39] | |
| Kanamycin | OmpF | E. Coli | 1 | ≈10 at gradient 10 μM [39] |
| OmpC | E. Coli | 1 | ≈11 at gradient 10 μM [39] | |
| LamB | E. Coli | ≤1 | ≤ 1 at gradient 10 μM [39] | |
| Chip | E. Coli | ≤1 | ≈5 at gradient 10 μM [39] | |
| Amikacin | OmpF | E. Coli | ≤1 | ≤1 at gradient 10 μM [39] |
| OmpC | E. Coli | ≤1 | ≤1 at gradient 10 μM [39] | |
| LamB | E. Coli | ≤1 | ≤1 at gradient 10 μM [39] | |
| Chip | E. Coli | ≤1 | ≈2 at gradient 10 μM [39] | |
| Sodium Glutamate | OprE | P. aeruginosa | ≤1 | ≈0.6 at gradient 10 μM [153] |
| Arginine Chloride | OprE | P. aeruginosa | ≤1 | ≈0.1 at gradient 10 μM [153] |
2. Discussion
The phenomenon of antibiotic/antimicrobial resistance due to deficient, lost, or altered membrane channels present in the bacterial outer membrane has remained well-documented and evidenced [6,7,29,31,37,40,107]. This can occur due to regulatory or mutational events in the porin gene or the regulatory cascade controlling bacterial envelope permeability [6,7,31,107]. During antibiotic treatments, porin exchange and decreased porin production have been observed in several species, including E. coli, E. aerogenes, E. cloacae, and K. pneumoniae [7,31]. Continuous antibiotic exposure leads to successive changes in porin expression, resulting in reduced influx and increased resistance to antibiotics or antimicrobials [7,23,31,38]. When porin is entirely absent in resistant bacteria, it may results in complete impermeability to β-lactams and possibly other antimicrobials, representing an effective adaptive response of bacteria to antibiotics [7,31]. Moreover, Mutations in porin channels, particularly near or at the constriction region of the porin channel, may lead to altered antibiotic translocation [6,7]. This has been observed in OmpC-type porin mutants isolated from chemotherapy patients, causing significant conformational changes in the OmpC pore lumen, reducing β-lactam translocation, and increased bacterial resistance [6,7,77]. Site-directed mutagenesis research into specific amino acid residues has emphasized the crucial role played by the constriction region of the porin channel. These changes result in a reduction in β-lactam translocation and increased bacterial resistance [77]. Additionally, research into specific amino acid residues has been conducted through site-directed mutagenesis and has emphasized the crucial role played by the constriction region of the porin channel [6,7,23,31].
Recent studies have further highlighted the direct role of outer membrane porins in transporting antibiotics across bacterial outer membranes [6,7,23,38]. The combination of various membrane porins and antibiotics were studied in different studies, such as Ampicillin, Cephalexin, Cefuroxime, Cefotaxime, Ceftazidime, Ceftriaxone/HMP-PA [46], Omp-PD [46], Omp-SL [46], HMP-AB [46], Imipenem/CarO isoforms CarO1, CarO2, CarO3 [155,156,157], Imipenem, Meropenem, Colistin, Ceftazidime, Ciprofloxacin/rOprD homolog [158], Imipenem, Meropenem, Ceftazidime, Ampicillin, Fosfomycin/OccAB1–OccAB4 [159], Sulbactam, Tazobactam, Ticarcillin, Ampicillin, Piperacillin/DcaP [22], Ampicillin, Cephalexin, Cefuroxime, Cefotaxime, Ceftazidime, Ceftriaxone/OmpA, and Omp-ECB [46], Meropenem/OmpF [55], Ampicillin, Benzylpenicillin/OmpF [26], Ciprofloxacin/OmpC [61] Sulbactam, Tazobactam, Avibactam/OmpF [40], OmpC [37], Imipenam, Meropenam/OmpC [77] Norfloxacin/OmpF [36], Enrofloxacin/OmpF [24,81,82,83], Imipenem, Ceftazidime, Cefepime/OmpF [78,79] Ampicillin, benzylpenicillin/OmpF [84,87,96,99], Grepafloxacin, Ciprofloxacin, Moxifloxacin, Nalidixic acid/Ompf [89,90,95], Carbenicillin, Ertapenem, Ampicillin/Ompf [100], Ampicillin, Amoxicillin, Carbenicillin, Azlocillin, Piperacillin/Ompf [44], Cefepime, Cefpirome, Cefotaxime, Ceftazidime, Cefoxitin, Ampicillin/OmpF [101], Cefepime/OmpF [102], Cefotaxime/OmpF [104], Ceftriaxone, Cefpirome, Ceftazidime/OmpF, OmpC [105], Ceftriaxone, Cefepime, Ceftazidime, Norfloxacin, Ciprofloxacin, Enrofloxacin/OmpF, OmpC [106], Piperacillin-Tazobactam/OmpF [107], Imipenem, Meropenem,/OmpF, OmpC [109], Benzylpenicillin, Ampicillin, Cefoxitin, Cefamandole, Cefotaxime, Ceftazidime, Ceftriaxone, Cephaloridine, Oxacillin, Cloxacillin, Cephalothin, Cefepime, Imipenem, Ertapenem, Novobiocin, Erythromycin/OmpF, OmpC [112], Fosfomycin/OmpF [76], Ceftazidime, Cefotaxime/OmpF, OmpC [38], Ciprofloxacin/OmpF [48]. Kanamycin/OmpF, OmpC [75], Kanamycin, Norfloxacin, Ciprofloxacin/OmpF, OmpC [51], Norfloxacin/OmpF, OmpC [177], Norfloxacin, Ciprofloxacin, Enoxacin/OmpF [74], Norfloxacin/OmpF [52], Kanamycin, Gentamicin, Amikacin/Ompf, OmpC, LamB, ChiP [39], Gentamicin, kanamycin, Polymyxin B/OmpF [53], Ceftazidime-Avibactam/Omp35, Omp36 [113], Imipenem, Meropenem/Omp35, Omp36 [114,115], Ceftriaxone/Omp35, Omp36 [116], Ertapenem, Cefepime/Omp36 [117], Imipenem, Cefepime, Cefpirome/Omp36 [118], Ampicillin, Ticarcillin, Piperacillin, Ertapenem, Imipenem, Meropenem, Cefotaxime, Cefepime, Ceftazidime/Omp35, Omp36 [23], Norfloxacin/Omp36, Omp35 [177], Ampicillin, Ticarcillin, Piperacillin, Ertapenem, Imipenem, Meropenem, Cefotaxime, Cefepime, Ceftazidime/OmpE36 and OmpE35 [23], Ceftazidime-Avibactam, Tigecycline, Colistin/OmpK36, OmpK35 [122], Ampicillin, Benzylpenicillin, Oxacillin, Cephaloridine, Cefoxitin, Cloxacillin, Cephalothin, Ceftriaxone, Cefotaxime, Cefamandole, Ceftazidime, Erythromycin Imipenem, Ertapenem, Cefepime, Novobiocin/OmpK35, OmpK36 [112,113], Ampicillin, Ticarcillin, Piperacillin, Ertapenem, Imipenem, Meropenem, Cefotaxime, Cefepime, Ceftazidime/OmpK36, OmpK35 [23], Ertapenem, Cefepime, Cefoxitin/OmpPst1 OmpPst2 [119], Carbapenem/OmpPst1 [121], Fosfomycin/OprO, Oprp [165], Meropenem, imipenem/OprD [140,143] Ceftazidime-Avibactam, Ceftolozane-Tazobactam/OprD [144], Imipenem, Meropenem/OccD3 [42], Temocillin/OccK1, OccK2 [135], Fosmidomycin/OprO, OprP [123], ceftazidime, cefotaxime, carbenicillin/Occk8(OprE) [164]. Note that the list is not limited to the mentioned names. These findings indicate that outer membrane porins can have an impending responsibility towards the efficacy of antimicrobials/antibiotics and may serve as potential targets to support the progression of innovative and novel antimicrobial approaches [6,31,35,180]. As per the studies, transmembrane proteins present in the outer membrane of the Gram-negative bacterium might be directly responsible for the transport of small molecules such as nutrients and antibiotics [6,35,180]. The porin channels are selective for specific types of molecules, and their size and charge selectivity determine the types of molecules that can cross the outer membrane [35,180]. The porins that have been shown to play a direct role or have shown interaction with antibiotics are HMP-PA [46],Omp-PD [46], Omp-SL [46], HMP-AB [46],/CarO isoforms [155,156,157], rOprD homologue [158], OccAB1–OccAB4 [159], DcaP [22], Omp-ECB [46], OmpF and OmpC [22,24,26,38,39,44,48,51,52,53,55,61,67,72,74,75,76,77,78,79,81,82,83,84,87,89,90,94,95,96,99,100,101,102,104,105,106,107,109,112,177] Omp35, Omp36 [21,23,66,113,114,115,116,117,118,177], OmpE36 and OmpE35 [23], OmpK36, OmpK35 [112,113,122], OmpPst1/OmpPst2 [119,121]/OprO, Oprp [123,165], OprD [140,141,142,143,144], OccD3 [42], OccK1, OccK2 [135], Occk8(OprE) [164]. From the studies it has been shown that the transport of Fosfomycin occurs through OmpF porins [76], while the transport of Cephalosporin occurs through OmpF and OmpC porins [23]. Fosfomycin, and Fosmidomycin transport through OprO/OprP [123,165] porins, and the dynamics of loop L3 from OmpF have been demonstrated to influence antibiotic translocation [48,101]. Additionally, ampicillin permeation has been demonstrated to occur through the major outer-membrane channel OmpF [26,54,181], and the fluoroquinolones interaction in conjunction with magnesium ions thru OmpF, OmpC, Omp36, and Omp35 porins [177]. Furthermore, Kanamycin and other aminoglycosides (see also Table 3) transport/uptake into the bacterium peremptorily facilitated by OmpF/OmpC channels [39], and the transport of small beta-lactamase inhibitors through OmpF/OmpC [26,37] (see Table 3) for other molecule/omps flux rates). As it is challenging to measure the flux of these tiny molecules, reported turnover rates molecules/second (Table 3) at a given concentration gradient for different outer membrane proteins (Omps) with different substrates have been given [26,27,39,54,153].
In general, these studies have provided insights into the explicit role of outer membrane proteins/porins/channels/transporters in the haulage of antibiotics and have highlighted the importance of these proteins in the efficacy of antibiotic therapy. Targeting these porins may provide a promising strategy for advancing and improving novel current antimicrobial/antibiotic agents and therapy. However, further extensive research and data sets are needed to comprehend the mechanisms and systems involved in antibiotic transport through porins. Further to fully identify new targets for the progress of novel antimicrobial strategies. Electrophysiology techniques, such as planar lipid bilayer, have provided valuable insights into Membrane channels’ structural and functional properties [7,24,29,49]. These techniques have also been used to examine the interface of antibiotics with the pore-forming complex, providing a molecular-level understanding of the transport mechanisms [7,24,29,37,42,49]. Despite these advances, a significant amount of information has been gathered toward understanding the correlation between pore geometry and antibiotic transport using electrophysiology alone [24,37,42,47,48,51,54,55,75,76,78,79,83,87,96,98,124,134,153,177].
Furthermore, the molecular transport mechanism through membrane channels is essential in developing targeted therapies against antimicrobial resistance [15,24]. Understanding how antibiotics interact with the bacteria at a molecular level makes it possible to design strategies that increase permeation and avoid resistance mechanisms based on decreased permeability. This can further take the lead toward the development of more efficient antibiotic therapeutics and systems to combat bacterial resistance [7,15,24,29,49]. It is also vital to note that the study of membrane channels is not constrained to their role in antimicrobial resistance. These proteins also play a crucial role in transporting essential nutrients and ions across the membrane, which is critical to the survival and growth of bacteria [7,24,29,49]. Therefore, studying membrane channels can also provide insights into the fundamental biology of bacteria [6,7,15,23,24,29,37,49].
Moreover, the study of membrane channels can also have implications in other fields, such as biotechnology and bioengineering [29,48]. The ability to manipulate the conveyance of solutes across the membrane can have applications in creating various products, such as enzymes and biosurfactants [7,29,31]. Additionally, the use of membrane channels as a standard system for studying protein–lipid interfaces and interactions and thus can also have implications in developing new drug delivery systems [15,24,40]. In conclusion, this review highlights the current scientific understanding of the role of membrane proteins/channels in controlling permeation [7,15,24,27,45].
Conclusive Remarks
The bacterial envelope’s outer membrane is a crucial component that is vital in controlling bacterial metabolism and adaptation [7,27,29,37,45]. One of the essential parts of the outer membrane is the outer membrane proteins, which regulate the diffusion of vital nutrients and other solutes across the membrane, thereby influencing bacterial survival and growth [7,29,182]. The unique structural and functional properties of outer membrane proteins, such as the constriction region and associated amino acids, may play a crucial role in determining the rate at which solutes diffuse across the bacterial envelope [7,27,29,37,45,48]. Outer membrane proteins have been extensively researched for many years, and recent advancements in methodology have provided new insights into the process of antibiotic translocation [15,29,30]. Specifically, membrane channels such as HMP-PA [46], Omp-PD [46], Omp-SL [46], HMP-AB [46],/CarO isoforms [155,157], rOprD homologue [158], OccAB1–OccAB4 [159], DcaP [22], Omp-ECB [46], OmpF and OmpC [24,26,38,39,44,48,51,52,53,55,61,74,75,76,77,78,79,81,82,83,84,87,89,90,95,96,99,100,101,102,104,105,106,107,109,112,177] Omp35, Omp36 [23,113,114,115,116,117,118,177], OmpE36 and OmpE35 [23], OmpK36, OmpK35 [112,113,122], OmpPst1/OmpPst2 [119,121]/OprO, Oprp [123,165], OprD [140,141,142,143,144], OccD3 [42], OccK1, OccK2 [135], Occk8(OprE) [164] have shown a critical role in controlling the membrane’s permeability in Gram-negative bacteria [23,27,37,38,45]. The β-lactam antibiotics have been considered and have significantly received the most attention regarding their translocation across outer membranes [7,23,38,87]. Over the last three decades, many studies have demonstrated the function of membrane transporters/porins in β-lactam uptake/translocation and the underlying characteristics required for uptake/translocation [7,23,31,37,38,42,77]. The effective flux all through the bacterial membrane and different transporters/porins can now be quantified in real-time using innovative techniques such as electrophysiology reversal potential assay, mass spectrometry, and other assays [7,31,37,45,54]. This has resulted in a better understanding of the structural and functional properties that determine outer membrane permeability and the intracellular concentration of permeating compounds, which ultimately impact bacterial susceptibility [7,27,31,37,38,40,54,153]. Studies have shown that permeation across porins occurs through passive diffusion for zwitterionic compounds through OmpF [44,100]. Further, it has been determined that OmpF may have higher permeability rates than OmpC, with the latter’s pore interior being more negative, making it more cation selective than OmpF [33,36,38,42,48]. In conjunction with the current structural determination of many bacterial membrane transporters/porins, numerous studies investigate their structural and physical properties to envisage small molecule translocation [44,46,77,100]. The work employs MS to understand the potential trajectories of small molecules to traverse porins and have produced essential observations [44,48,76,77,132,177]. The study of porins using electrophysiology gives a better understanding of the uptake of drugs through different porins, which can be readily measured by changes in ion flux [181,183]. The example has shown that the ion-current through with the OmpF channel decreases temporarily dependent on pH in the presence of ampicillin molecule, the maximum decrease occurring at the range of pH where ampicillin is zwitterionic [181,183].
MS suggests that ampicillin may obstructs the OmpF pore by interacting with charged residues of the membrane pore [87]. Similar outcomes were found for amoxicillin, a zwitterionic β-lactam [84,99,181,183]. However, no interaction was observed with carbenicillin, azlocillin, and piperacillin [84,99,181,183]. This implies that interactions at the channel’s recognition site correspond to increased diffusion of particular compounds through OmpF [84,99,181,183], as shown by previous studies indicating that mutations of critical residues in the porin affect β-lactam uptake and susceptibility [7,84,88,99,181,183]. An alternative method involves introducing the charged compound of interest to one side of the channel to create a concentration-driven flux, which can be assessed by a diffusion potential [43,44,88,181]. The permeation assay, based on the conventional electrophysiology method, primarily entails the application of concentration gradients of charged molecules with different electrophoretic mobilities through a membrane channel [26,39,54,153].
Consequently, the unequal flux generates an electrostatic potential that provides direct evidence of relative ion fluxes [37,38,40,54,153]. The investigative methodology utilized here involves measuring zero-current-reversal potentials and the subsequent channel conductance of a single pore [37,38,40,54,153]. Combined with single-channel conductance measurement, this technique allows for more quantitative analysis [37,40]. In summary, electrophysiology and its integration with other techniques have yielded valuable insights into the uptake of drugs through porins, enabling the development of new methods to differentiate between molecular binding and transport [37,40]. These techniques can potentially aid in discovering new scaffolds to enhance the permeability of antibiotics [37,39,40,54,153,164,165]. Recently, the efficacy of these approaches has been validated using in vivo susceptibility and accumulation assays on intact bacterial cells by comparing different methodologies to measure the transport of ceftazidime and cefotaxime [38].
Likewise, these new methodologies offer the opportunity to study membrane porins and their role in influx and environmental stressors affecting them [7,29]. In real-time, measurements of the interbacterial accumulation of individual molecules, such as antibiotics, inhibitors, antimicrobials, and more, provide valuable insights into the early phases of envelope changes during bacterial life [7,29].
These studies are essential for understanding the relationships between bacterial adaptation and various environmental stressors, which could provide new targets for developing antibiotics [7]. In conclusion, the advances toward understanding bacterial porins can contribute to a deeper understanding of the transport of hydrophilic molecules across lipid bilayers, a fundamental process in living cells [7]. By investigating the molecular events that regulate the efficiency of diffusion rate, these studies will aid in our efforts to understand how gram-negative bacteria manage to transport essential molecules while adapting to changing environmental conditions [7,15,24,29].
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Gootz, T.D. The global problem of antibiotic resistance. Crit. Rev. Immunol. 2010, 30, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Dever, L.A.; Dermody, T.S. Mechanisms of bacterial resistance to antibiotics. Arch. Intern. Med. 1991, 151, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Hofmann, E.; Coulton, J.W.; Diederichs, K.; Welte, W. Siderophore-mediated iron transport: Crystal structure of FhuA with bound lipopolysaccharide. Science 1998, 282, 2215–2220. [Google Scholar] [CrossRef] [Green Version]
- Pages, J.M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Winterhalter, M.; Pages, J.M. Outer Membrane Porins. Subcell. Biochem. 2019, 92, 79–123. [Google Scholar] [CrossRef]
- Humphries, R.M.; Yang, S.; Hemarajata, P.; Ward, K.W.; Hindler, J.A.; Miller, S.A.; Gregson, A. First Report of Ceftazidime-Avibactam Resistance in a KPC-3-Expressing Klebsiella pneumoniae Isolate. Antimicrob. Agents Chemother. 2015, 59, 6605–6607. [Google Scholar] [CrossRef] [Green Version]
- Mangili, A.; Bica, I.; Snydman, D.R.; Hamer, D.H. Daptomycin-resistant, methicillin-resistant Staphylococcus aureus bacteremia. Clin. Infect. Dis. 2005, 40, 1058–1060. [Google Scholar] [CrossRef] [Green Version]
- Yigit, H.; Queenan, A.M.; Anderson, G.J.; Domenech-Sanchez, A.; Biddle, J.W.; Steward, C.D.; Alberti, S.; Bush, K.; Tenover, F.C. Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Soge, O.O.; Harger, D.; Schafer, S.; Toevs, K.; Raisler, K.A.; Venator, K.; Holmes, K.K.; Kirkcaldy, R.D. Emergence of increased azithromycin resistance during unsuccessful treatment of Neisseria gonorrhoeae infection with azithromycin (Portland, OR, 2011). Sex Transm. Dis. 2012, 39, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Jevons, M.P.; Rolinson, G.N.; Knox, R. Celbenin-Resistant Staphylococci. Br. Med. J. 1961, 1, 124. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. P T 2015, 40, 277–283. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Stavenger, R.A.; Winterhalter, M. TRANSLOCATION project: How to get good drugs into bad bugs. Sci. Transl. Med. 2014, 6, 228ed227. [Google Scholar] [CrossRef]
- Coculescu, B.I. Antimicrobial resistance induced by genetic changes. J. Med. Life 2009, 2, 114–123. [Google Scholar]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Et Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Simpkin, V.L.; Renwick, M.J.; Kelly, R.; Mossialos, E. Incentivising innovation in antibiotic drug discovery and development: Progress, challenges and next steps. J. Antibiot. 2017, 70, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koizumi, R.; Kusama, Y.; Muraki, Y.; Ishikane, M.; Yamasaki, D.; Tanabe, M.; Ohmagari, N. Effect of population inflow and outflow between rural and urban areas on regional antimicrobial use surveillance. PLoS ONE 2021, 16, e0248338. [Google Scholar] [CrossRef]
- Schirmer, T. General and specific porins from bacterial outer membranes. J. Struct. Biol. 1998, 121, 101–109. [Google Scholar] [CrossRef]
- Nikaido, H.; Nakae, T. The outer membrane of Gram-negative bacteria. Adv. Microb. Physiol. 1979, 20, 163–250. [Google Scholar] [CrossRef]
- Bhamidimarri, S.P.; Zahn, M.; Prajapati, J.D.; Schleberger, C.; Soderholm, S.; Hoover, J.; West, J.; Kleinekathofer, U.; Bumann, D.; Winterhalter, M.; et al. A Multidisciplinary Approach toward Identification of Antibiotic Scaffolds for Acinetobacter baumannii. Structure 2019, 27, 268–280.E6. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Gutierrez, S.; Ferrara, L.; Pathania, M.; Masi, M.; Wang, J.; Bodrenko, I.; Zahn, M.; Winterhalter, M.; Stavenger, R.A.; Pages, J.M.; et al. Getting Drugs into Gram-Negative Bacteria: Rational Rules for Permeation through General Porins. ACS Infect. Dis. 2018, 4, 1487–1498. [Google Scholar] [CrossRef] [Green Version]
- Winterhalter, M.; Ceccarelli, M. Physical methods to quantify small antibiotic molecules uptake into Gram-negative bacteria. Eur. J. Pharm Biopharm 2015, 95, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Kostyanev, T.; Bonten, M.J.; O’Brien, S.; Steel, H.; Ross, S.; Francois, B.; Tacconelli, E.; Winterhalter, M.; Stavenger, R.A.; Karlen, A.; et al. The Innovative Medicines Initiative’s New Drugs for Bad Bugs programme: European public-private partnerships for the development of new strategies to tackle antibiotic resistance. J. Antimicrob. Chemother. 2016, 71, 290–295. [Google Scholar] [CrossRef]
- Ghai, I. Quantifying the Flux of Charged Molecules through Bacterial Membrane Proteins. J. Phys. Chem. Lett. 2017, 8, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Ghai, I.; Ghai, S. Exploring bacterial outer membrane barrier to combat bad bugs. Infect. Drug Resist. 2017, 10, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, J.; Reygaert, W.C. Gram Negative Bacteria. In StatPearls; StatPearls Publishing Copyright © 2022; StatPearls Publishing LLC: Treasure Island, FL, USA, 2022. [Google Scholar]
- Winterhalter, M. Antibiotic uptake through porins located in the outer membrane of Gram-negative bacteria. Expert Opin. Drug Deliv. 2021, 18, 449–457. [Google Scholar] [CrossRef]
- Nikaido, H. Prevention of drug access to bacterial targets: Permeability barriers and active efflux. Science 1994, 264, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Pages, J.M. Structure, Function and Regulation of Outer Membrane Proteins Involved in Drug Transport in Enterobactericeae: The OmpF/C-TolC Case. Open Microbiol. J. 2013, 7, 22–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, W.; Roux, B. Ion permeation and selectivity of OmpF porin: A theoretical study based on molecular dynamics, Brownian dynamics, and continuum electrodiffusion theory. J. Mol. Biol. 2002, 322, 851–869. [Google Scholar] [CrossRef]
- Liu, J.; Wolfe, A.J.; Eren, E.; Vijayaraghavan, J.; Indic, M.; van den Berg, B.; Movileanu, L. Cation selectivity is a conserved feature in the OccD subfamily of Pseudomonas aeruginosa. Biochim. Biophys. Acta 2012, 1818, 2908–2916. [Google Scholar] [CrossRef] [Green Version]
- Modi, N.; Benz, R.; Hancock, R.E.; Kleinekathofer, U. Modeling the Ion Selectivity of the Phosphate Specific Channel OprP. J. Phys. Chem. Lett. 2012, 3, 3639–3645. [Google Scholar] [CrossRef] [PubMed]
- Weingart, H.; Petrescu, M.; Winterhalter, M. Biophysical characterization of in- and efflux in Gram-negative bacteria. Curr. Drug Targets 2008, 9, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Cama, J.; Bajaj, H.; Pagliara, S.; Maier, T.; Braun, Y.; Winterhalter, M.; Keyser, U.F. Quantification of Fluoroquinolone Uptake through the Outer Membrane Channel OmpF of Escherichia coli. J. Am. Chem Soc. 2015, 137, 13836–13843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghai, I.; Winterhalter, M.; Wagner, R. Probing transport of charged beta-lactamase inhibitors through OmpC, a membrane channel from E. coli. Biochem. Biophys. Res. Commun. 2017, 484, 51–55. [Google Scholar] [CrossRef]
- Masi, M.; Vergalli, J.; Ghai, I.; Barba-Bon, A.; Schembri, T.; Nau, W.M.; Lafitte, D.; Winterhalter, M.; Pages, J.M. Cephalosporin translocation across enterobacterial OmpF and OmpC channels, a filter across the outer membrane. Commun. Biol. 2022, 5, 1059. [Google Scholar] [CrossRef]
- Paul, E.; Ghai, I.; Hörömpöli, D.; Brötz-Oesterhelt, H.; Winterhalter, M.; Bafna, J.A. Uptake of aminoglycosides through outer membrane porins in Escherichia coli. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ghai, I.; Pira, A.; Scorciapino, M.A.; Bodrenko, I.; Benier, L.; Ceccarelli, M.; Winterhalter, M.; Wagner, R. General Method to Determine the Flux of Charged Molecules through Nanopores Applied to beta-Lactamase Inhibitors and OmpF. J. Phys. Chem. Lett 2017, 8, 1295–1301. [Google Scholar] [CrossRef]
- Bajaj, H.; Acosta Gutierrez, S.; Bodrenko, I.; Malloci, G.; Scorciapino, M.A.; Winterhalter, M.; Ceccarelli, M. Bacterial Outer Membrane Porins as Electrostatic Nanosieves: Exploring Transport Rules of Small Polar Molecules. ACS Nano 2017, 11, 5465–5473. [Google Scholar] [CrossRef] [Green Version]
- Soundararajan, G.; Bhamidimarri, S.P.; Winterhalter, M. Understanding Carbapenem Translocation through OccD3 (OpdP) of Pseudomonas aeruginosa. ACS Chem. Biol. 2017, 12, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.; Danelon, C.; Winterhalter, M. On translocation through a membrane channel via an internal binding site: Kinetics and voltage dependence. Biophys. J. 2003, 84, 2990–2998. [Google Scholar] [CrossRef] [Green Version]
- Danelon, C.; Nestorovich, E.M.; Winterhalter, M.; Ceccarelli, M.; Bezrukov, S.M. Interaction of zwitterionic penicillins with the OmpF channel facilitates their translocation. Biophys. J. 2006, 90, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Ghai, I.; Ghai, S. Understanding antibiotic resistance via outer membrane permeability. Infect. Drug Resist. 2018, 11, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Nitzan, Y.; Deutsch, E.B.; Pechatnikov, I. Diffusion of beta-lactam antibiotics through oligomeric or monomeric porin channels of some gram-negative bacteria. Curr. Microbiol. 2002, 45, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.; Bafna, J.A.; Schmid, B.; Klingl, S.; Baier, S.; Hemmis, B.; Wagner, R.; Winterhalter, M.; Voll, L.M. Manipulation of charge distribution in the arginine and glutamate clusters of the OmpG pore alters sugar specificity and ion selectivity. Biochim. Biophys. Acta Biomembr. 2019, 1861, 183021. [Google Scholar] [CrossRef]
- Acharya, A.; Ghai, I.; Piselli, C.; Prajapati, J.D.; Benz, R.; Winterhalter, M.; Kleinekathofer, U. Conformational Dynamics of Loop L3 in OmpF: Implications toward Antibiotic Translocation and Voltage Gating. J. Chem. Inf. Model 2023, 63, 910–927. [Google Scholar] [CrossRef] [PubMed]
- Bodrenko, I.V.; Zewdie, T.A.; Wang, J.; Paul, E.; Witt, S.; Winterhalter, M. TolC-AcrA complex formation monitored by time dependent single-channel electrophysiology. Biochimie 2023, 205, 102–109. [Google Scholar] [CrossRef]
- Gilardi, A.; Bhamidimarri, S.P.; Bronstrup, M.; Bilitewski, U.; Marreddy, R.K.R.; Pos, K.M.; Benier, L.; Gribbon, P.; Winterhalter, M.; Windshugel, B. Biophysical characterization of E. coli TolC interaction with the known blocker hexaamminecobalt. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2702–2709. [Google Scholar] [CrossRef]
- Wang, J.; Terrasse, R.; Bafna, J.A.; Benier, L.; Winterhalter, M. Electrophysiological Characterization of Transport Across Outer-Membrane Channels from Gram-Negative Bacteria in Presence of Lipopolysaccharides. Angew. Chem. Int. Ed. 2020, 59, 8517–8521. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Bafna, J.A.; Bhamidimarri, S.P.; Winterhalter, M. Small-Molecule Permeation across Membrane Channels: Chemical Modification to Quantify Transport across OmpF. Angew. Chem. Int. Ed. 2019, 58, 4737–4741. [Google Scholar] [CrossRef]
- Hancock, R.E.; Farmer, S.W.; Li, Z.S.; Poole, K. Interaction of aminoglycosides with the outer membranes and purified lipopolysaccharide and OmpF porin of Escherichia coli. Antimicrob. Agents Chemother. 1991, 35, 1309–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghai, I.; Bajaj, H.; Arun Bafna, J.; El Damrany Hussein, H.A.; Winterhalter, M.; Wagner, R. Ampicillin permeation across OmpF, the major outer-membrane channel in Escherichia coli. J. Biol. Chem. 2018, 293, 7030–7037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodrenko, I.V.; Wang, J.; Salis, S.; Winterhalter, M.; Ceccarelli, M. Sensing Single Molecule Penetration into Nanopores: Pushing the Time Resolution to the Diffusion Limit. ACS Sens. 2017, 2, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, A.; Lopez, M.L.; Queralt-Martin, M.; Aguilella, V.M. Ion Transport in Confined Geometries below the Nanoscale: Access Resistance Dominates Protein Channel Conductance in Diluted Solutions. ACS Nano 2017, 11, 10392–10400. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gimenez, E.; Alcaraz, A.; Aguilella, V.M. Divalent Metal Ion Transport across Large Biological Ion Channels and Their Effect on Conductance and Selectivity. Biochem. Res. Int. 2012, 2012, 245786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gimenez, E.; Lopez, M.L.; Aguilella, V.M.; Alcaraz, A. Linearity, saturation and blocking in a large multiionic channel: Divalent cation modulation of the OmpF porin conductance. Biochem. Biophys. Res. Commun. 2011, 404, 330–334. [Google Scholar] [CrossRef]
- Queralt-Martin, M.; Garcia-Gimenez, E.; Mafe, S.; Alcaraz, A. Divalent cations reduce the pH sensitivity of OmpF channel inducing the pK(a) shift of key acidic residues. Phys. Chem. Chem. Phys. PCCP 2011, 13, 563–569. [Google Scholar] [CrossRef]
- Alcaraz, A.; Nestorovich, E.M.; Lopez, M.L.; Garcia-Gimenez, E.; Bezrukov, S.M.; Aguilella, V.M. Diffusion, exclusion, and specific binding in a large channel: A study of OmpF selectivity inversion. Biophys. J. 2009, 96, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, J.D.; Fernandez Solano, C.J.; Winterhalter, M.; Kleinekathofer, U. Characterization of Ciprofloxacin Permeation Pathways across the Porin OmpC Using Metadynamics and a String Method. J. Chem. Theory Comput. 2017, 13, 4553–4566. [Google Scholar] [CrossRef]
- Conlan, S.; Zhang, Y.; Cheley, S.; Bayley, H. Biochemical and biophysical characterization of OmpG: A monomeric porin. Biochemistry 2000, 39, 11845–11854. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, G.V.; van den Berg, B. Crystal structure of the monomeric porin OmpG. J. Mol. Biol. 2006, 360, 750–759. [Google Scholar] [CrossRef]
- Behlau, M.; Mills, D.J.; Quader, H.; Kuhlbrandt, W.; Vonck, J. Projection structure of the monomeric porin OmpG at 6 A resolution. J. Mol. Biol. 2001, 305, 71–77. [Google Scholar] [CrossRef]
- Conlan, S.; Bayley, H. Folding of a monomeric porin, OmpG, in detergent solution. Biochemistry 2003, 42, 9453–9465. [Google Scholar] [CrossRef]
- Qu, J.; Behrens-Kneip, S.; Holst, O.; Kleinschmidt, J.H. Binding regions of outer membrane protein A in complexes with the periplasmic chaperone Skp. A site-directed fluorescence study. Biochemistry 2009, 48, 4926–4936. [Google Scholar] [CrossRef]
- Wang, Y. The function of OmpA in Escherichia coli. Biochem. Biophys. Res. Commun. 2002, 292, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Vogt, J.; Schulz, G.E. The structure of the outer membrane protein OmpX from Escherichia coli reveals possible mechanisms of virulence. Structure 1999, 7, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ghai, I.; Winterhalter, M.; Schwaneberg, U. Engineering Enhanced Pore Sizes Using FhuA Delta1-160 from E. coli Outer Membrane as Template. ACS Sens. 2017, 2, 1619–1626. [Google Scholar] [CrossRef]
- Plancon, L.; Chami, M.; Letellier, L. Reconstitution of FhuA, an Escherichia coli outer membrane protein, into liposomes. Binding of phage T5 to Fhua triggers the transfer of DNA into the proteoliposomes. J. Biol. Chem. 1997, 272, 16868–16872. [Google Scholar] [CrossRef] [Green Version]
- Bonhivers, M.; Plancon, L.; Ghazi, A.; Boulanger, P.; le Maire, M.; Lambert, O.; Rigaud, J.L.; Letellier, L. FhuA, an Escherichia coli outer membrane protein with a dual function of transporter and channel which mediates the transport of phage DNA. Biochimie 1998, 80, 363–369. [Google Scholar] [CrossRef]
- Braun, V.; Endriss, F. Energy-coupled outer membrane transport proteins and regulatory proteins. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2007, 20, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Braun, V.; Killmann, H.; Benz, R. Energy-coupled transport through the outer membrane of Escherichia coli small deletions in the gating loop convert the FhuA transport protein into a diffusion channel. FEBS Lett. 1994, 346, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Bafna, J.A.; Pangeni, S.; Winterhalter, M.; Aksoyoglu, M.A. Electroosmosis Dominates Electrophoresis of Antibiotic Transport Across the Outer Membrane Porin F. Biophys. J. 2020, 118, 2844–2852. [Google Scholar] [CrossRef] [PubMed]
- Bafna, J.A.; Sans-Serramitjana, E.; Acosta-Gutierrez, S.; Bodrenko, I.V.; Horompoli, D.; Berscheid, A.; Brotz-Oesterhelt, H.; Winterhalter, M.; Ceccarelli, M. Kanamycin Uptake into Escherichia coli Is Facilitated by OmpF and OmpC Porin Channels Located in the Outer Membrane. ACS Infect. Dis. 2020, 6, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Golla, V.K.; Sans-Serramitjana, E.; Pothula, K.R.; Benier, L.; Bafna, J.A.; Winterhalter, M.; Kleinekathofer, U. Fosfomycin Permeation through the Outer Membrane Porin OmpF. Biophys. J. 2019, 116, 258–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, H.; Scorciapino, M.A.; Moynie, L.; Page, M.G.; Naismith, J.H.; Ceccarelli, M.; Winterhalter, M. Molecular Basis of Filtering Carbapenems by Porins from beta-Lactam-resistant Clinical Strains of Escherichia coli. J. Biol. Chem. 2016, 291, 2837–2847. [Google Scholar] [CrossRef] [Green Version]
- Weichbrodt, C.; Bajaj, H.; Baaken, G.; Wang, J.; Guinot, S.; Kreir, M.; Behrends, J.C.; Winterhalter, M.; Fertig, N. Antibiotic translocation through porins studied in planar lipid bilayers using parallel platforms. Analyst 2015, 140, 4874–4881. [Google Scholar] [CrossRef]
- Bodrenko, I.; Bajaj, H.; Ruggerone, P.; Winterhalter, M.; Ceccarelli, M. Analysis of fast channel blockage: Revealing substrate binding in the microsecond range. Analyst 2015, 140, 4820–4827. [Google Scholar] [CrossRef]
- Lamichhane, U.; Islam, T.; Prasad, S.; Weingart, H.; Mahendran, K.R.; Winterhalter, M. Peptide translocation through the mesoscopic channel: Binding kinetics at the single molecule level. Eur. Biophys. J. 2013, 42, 363–369. [Google Scholar] [CrossRef]
- Singh, P.R.; Ceccarelli, M.; Lovelle, M.; Winterhalter, M.; Mahendran, K.R. Antibiotic permeation across the OmpF channel: Modulation of the affinity site in the presence of magnesium. J. Phys. Chem. B 2012, 116, 4433–4438. [Google Scholar] [CrossRef]
- Brauser, A.; Schroeder, I.; Gutsmann, T.; Cosentino, C.; Moroni, A.; Hansen, U.P.; Winterhalter, M. Modulation of enrofloxacin binding in OmpF by Mg2+ as revealed by the analysis of fast flickering single-porin current. J. Gen. Physiol. 2012, 140, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Mahendran, K.R.; Hajjar, E.; Mach, T.; Lovelle, M.; Kumar, A.; Sousa, I.; Spiga, E.; Weingart, H.; Gameiro, P.; Winterhalter, M.; et al. Molecular basis of enrofloxacin translocation through OmpF, an outer membrane channel of Escherichia coli--when binding does not imply translocation. J. Phys. Chem. B 2010, 114, 5170–5179. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, E.; Bessonov, A.; Molitor, A.; Kumar, A.; Mahendran, K.R.; Winterhalter, M.; Pages, J.M.; Ruggerone, P.; Ceccarelli, M. Toward screening for antibiotics with enhanced permeation properties through bacterial porins. Biochemistry 2010, 49, 6928–6935. [Google Scholar] [CrossRef] [PubMed]
- Phale, P.S.; Philippsen, A.; Widmer, C.; Phale, V.P.; Rosenbusch, J.P.; Schirmer, T. Role of charged residues at the OmpF porin channel constriction probed by mutagenesis and simulation. Biochemistry 2001, 40, 6319–6325. [Google Scholar] [CrossRef]
- Aguilella-Arzo, M.; Garcia-Celma, J.J.; Cervera, J.; Alcaraz, A.; Aguilella, V.M. Electrostatic properties and macroscopic electrodiffusion in OmpF porin and mutants. Bioelectrochemistry 2007, 70, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Nestorovich, E.M.; Danelon, C.; Winterhalter, M.; Bezrukov, S.M. Designed to penetrate: Time-resolved interaction of single antibiotic molecules with bacterial pores. Proc. Natl. Acad. Sci. USA 2002, 99, 9789–9794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danelon, C.; Suenaga, A.; Winterhalter, M.; Yamato, I. Molecular origin of the cation selectivity in OmpF porin: Single channel conductances vs. free energy calculation. Biophys. Chem. 2003, 104, 591–603. [Google Scholar] [CrossRef]
- Neves, P.; Berkane, E.; Gameiro, P.; Winterhalter, M.; de Castro, B. Interaction between quinolones antibiotics and bacterial outer membrane porin OmpF. Biophys. Chem. 2005, 113, 123–128. [Google Scholar] [CrossRef]
- Mach, T.; Neves, P.; Spiga, E.; Weingart, H.; Winterhalter, M.; Ruggerone, P.; Ceccarelli, M.; Gameiro, P. Facilitated permeation of antibiotics across membrane channels--interaction of the quinolone moxifloxacin with the OmpF channel. J. Am. Chem. Soc. 2008, 130, 13301–13309. [Google Scholar] [CrossRef]
- Iyer, R.; Delcour, A.H. Complex inhibition of OmpF and OmpC bacterial porins by polyamines. J. Biol. Chem. 1997, 272, 18595–18601. [Google Scholar] [CrossRef] [Green Version]
- Saint, N.; Lou, K.L.; Widmer, C.; Luckey, M.; Schirmer, T.; Rosenbusch, J.P. Structural and functional characterization of OmpF porin mutants selected for larger pore size. II. Functional characterization. J. Biol. Chem. 1996, 271, 20676–20680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, K.L.; Saint, N.; Prilipov, A.; Rummel, G.; Benson, S.A.; Rosenbusch, J.P.; Schirmer, T. Structural and functional characterization of OmpF porin mutants selected for larger pore size. I. Crystallographic analysis. J. Biol. Chem. 1996, 271, 20669–20675. [Google Scholar] [CrossRef] [Green Version]
- Phale, P.S.; Schirmer, T.; Prilipov, A.; Lou, K.L.; Hardmeyer, A.; Rosenbusch, J.P. Voltage gating of Escherichia coli porin channels: Role of the constriction loop. Proc. Natl. Acad. Sci. USA 1997, 94, 6741–6745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, P.; Sousa, I.; Winterhalter, M.; Gameiro, P. Fluorescence quenching as a tool to investigate quinolone antibiotic interactions with bacterial protein OmpF. J. Membr. Biol. 2009, 227, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, E.; Mahendran, K.R.; Kumar, A.; Bessonov, A.; Petrescu, M.; Weingart, H.; Ruggerone, P.; Winterhalter, M.; Ceccarelli, M. Bridging timescales and length scales: From macroscopic flux to the molecular mechanism of antibiotic diffusion through porins. Biophys. J. 2010, 98, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Pezeshki, S.; Chimerel, C.; Bessonov, A.N.; Winterhalter, M.; Kleinekathofer, U. Understanding ion conductance on a molecular level: An all-atom modeling of the bacterial porin OmpF. Biophys. J. 2009, 97, 1898–1906. [Google Scholar] [CrossRef] [Green Version]
- Apetrei, A.; Asandei, A.; Park, Y.; Hahm, K.S.; Winterhalter, M.; Luchian, T. Unimolecular study of the interaction between the outer membrane protein OmpF from E. coli and an analogue of the HP(2-20) antimicrobial peptide. J. Bioenerg. Biomembr. 2010, 42, 173–180. [Google Scholar] [CrossRef]
- Kumar, A.; Hajjar, E.; Ruggerone, P.; Ceccarelli, M. Molecular simulations reveal the mechanism and the determinants for ampicillin translocation through OmpF. J. Phys. Chem. B 2010, 114, 9608–9616. [Google Scholar] [CrossRef]
- Ziervogel, B.K.; Roux, B. The binding of antibiotics in OmpF porin. Structure 2013, 21, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Vidal, S.; Bredin, J.; Pages, J.M.; Barbe, J. Beta-lactam screening by specific residues of the OmpF eyelet. J. Med. Chem. 2005, 48, 1395–1400. [Google Scholar] [CrossRef]
- Bredin, J.; Saint, N.; Mallea, M.; De, E.; Molle, G.; Pages, J.M.; Simonet, V. Alteration of pore properties of Escherichia coli OmpF induced by mutation of key residues in anti-loop 3 region. Biochem. J. 2002, 363, 521–528. [Google Scholar] [CrossRef]
- Liu, N.; Samartzidou, H.; Lee, K.W.; Briggs, J.M.; Delcour, A.H. Effects of pore mutations and permeant ion concentration on the spontaneous gating activity of OmpC porin. Protein Eng. 2000, 13, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.; Chen, M.; Black, S.S.; Bushell, S.R.; Ceccarelli, M.; Mach, T.; Beis, K.; Low, A.S.; Bamford, V.A.; Booth, I.R.; et al. Altered antibiotic transport in OmpC mutants isolated from a series of clinical strains of multi-drug resistant E. coli. PLoS ONE 2011, 6, e25825. [Google Scholar] [CrossRef] [Green Version]
- Lovelle, M.; Mach, T.; Mahendran, K.R.; Weingart, H.; Winterhalter, M.; Gameiro, P. Interaction of cephalosporins with outer membrane channels of Escherichia coli. Revealing binding by fluorescence quenching and ion conductance fluctuations. Phys. Chem. Chem. Phys. PCCP 2011, 13, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, K.R.; Kreir, M.; Weingart, H.; Fertig, N.; Winterhalter, M. Permeation of antibiotics through Escherichia coli OmpF and OmpC porins: Screening for influx on a single-molecule level. J. Biomol. Screen. 2010, 15, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Pinet, E.; Franceschi, C.; Davin-Regli, A.; Zambardi, G.; Pages, J.M. Role of the culture medium in porin expression and piperacillin-tazobactam susceptibility in Escherichia coli. J. Med. Microbiol. 2015, 64, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Acosta Gutierrez, S.; Bodrenko, I.; Scorciapino, M.A.; Ceccarelli, M. Macroscopic electric field inside water-filled biological nanopores. Phys. Chem. Chem. Phys. PCCP 2016, 18, 8855–8864. [Google Scholar] [CrossRef] [PubMed]
- Scorciapino, M.A.; D’Agostino, T.; Acosta-Gutierrez, S.; Malloci, G.; Bodrenko, I.; Ceccarelli, M. Exploiting the porin pathway for polar compound delivery into Gram-negative bacteria. Future Med. Chem. 2016, 8, 1047–1062. [Google Scholar] [CrossRef] [Green Version]
- Benz, R.; Darveau, R.P.; Hancock, R.E. Outer-membrane protein PhoE from Escherichia coli forms anion-selective pores in lipid-bilayer membranes. Eur. J. Biochem. 1984, 140, 319–324. [Google Scholar] [CrossRef]
- Darveau, R.P.; Hancock, R.E.; Benz, R. Chemical modification of the anion selectivity of the PhoE porin from the Escherichia coli outer membrane. Biochim. Biophys. Acta 1984, 774, 67–74. [Google Scholar] [CrossRef]
- Sugawara, E.; Kojima, S.; Nikaido, H. Klebsiella pneumoniae Major Porins OmpK35 and OmpK36 Allow More Efficient Diffusion of beta-Lactams than Their Escherichia coli Homologs OmpF and OmpC. J. Bacteriol. 2016, 198, 3200–3208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pages, J.M.; Peslier, S.; Keating, T.A.; Lavigne, J.P.; Nichols, W.W. Role of the Outer Membrane and Porins in Susceptibility of beta-Lactamase-Producing Enterobacteriaceae to Ceftazidime-Avibactam. Antimicrob. Agents Chemother. 2015, 60, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Bornet, C.; Davin-Regli, A.; Bosi, C.; Pages, J.M.; Bollet, C. Imipenem resistance of enterobacter aerogenes mediated by outer membrane permeability. J. Clin. Microbiol. 2000, 38, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Lavigne, J.P.; Sotto, A.; Nicolas-Chanoine, M.H.; Bouziges, N.; Pages, J.M.; Davin-Regli, A. An adaptive response of Enterobacter aerogenes to imipenem: Regulation of porin balance in clinical isolates. Int. J. Antimicrob. Agents 2013, 41, 130–136. [Google Scholar] [CrossRef]
- Bornet, C.; Saint, N.; Fetnaci, L.; Dupont, M.; Davin-Regli, A.; Bollet, C.; Pages, J.M. Omp35, a new Enterobacter aerogenes porin involved in selective susceptibility to cephalosporins. Antimicrob. Agents Chemother. 2004, 48, 2153–2158. [Google Scholar] [CrossRef] [Green Version]
- James, C.E.; Mahendran, K.R.; Molitor, A.; Bolla, J.M.; Bessonov, A.N.; Winterhalter, M.; Pages, J.M. How beta-lactam antibiotics enter bacteria: A dialogue with the porins. PLoS ONE 2009, 4, e5453. [Google Scholar] [CrossRef] [PubMed]
- Thiolas, A.; Bornet, C.; Davin-Regli, A.; Pages, J.M.; Bollet, C. Resistance to imipenem, cefepime, and cefpirome associated with mutation in Omp36 osmoporin of Enterobacter aerogenes. Biochem. Biophys. Res. Commun. 2004, 317, 851–856. [Google Scholar] [CrossRef]
- Tran, Q.T.; Mahendran, K.R.; Hajjar, E.; Ceccarelli, M.; Davin-Regli, A.; Winterhalter, M.; Weingart, H.; Pages, J.M. Implication of porins in beta-lactam resistance of Providencia stuartii. J. Biol. Chem. 2010, 285, 32273–32281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, Q.T.; Maigre, L.; D’Agostino, T.; Ceccarelli, M.; Winterhalter, M.; Pages, J.M.; Davin-Regli, A. Porin flexibility in Providencia stuartii: Cell-surface-exposed loops L5 and L7 are markers of Providencia porin OmpPst1. Res. Microbiol. 2017, 168, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, H.; Tran, Q.T.; Mahendran, K.R.; Nasrallah, C.; Colletier, J.P.; Davin-Regli, A.; Bolla, J.M.; Pages, J.M.; Winterhalter, M. Antibiotic uptake through membrane channels: Role of Providencia stuartii OmpPst1 porin in carbapenem resistance. Biochemistry 2012, 51, 10244–10249. [Google Scholar] [CrossRef]
- Castanheira, M.; Mendes, R.E.; Sader, H.S. Low Frequency of Ceftazidime-Avibactam Resistance among Enterobacteriaceae Isolates Carrying bla(KPC) Collected in U.S. Hospitals from 2012 to 2015. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piselli, C.; Benz, R. Fosmidomycin transport through the phosphate-specific porins OprO and OprP of Pseudomonas aeruginosa. Mol. Microbiol. 2021, 116, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Dogan Guzel, F.; Pletzer, D.; Norouz Dizaji, A.; Al-Nahas, K.; Bajrai, M.; Winterhalter, M. Towards understanding single-channel characteristics of OccK8 purified from Pseudomonas aeruginosa. Eur. Biophys. J. 2021, 50, 87–98. [Google Scholar] [CrossRef]
- Eren, E.; Parkin, J.; Adelanwa, A.; Cheneke, B.; Movileanu, L.; Khalid, S.; van den Berg, B. Toward understanding the outer membrane uptake of small molecules by Pseudomonas aeruginosa. J. Biol. Chem. 2013, 288, 12042–12053. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Eren, E.; Vijayaraghavan, J.; Cheneke, B.R.; Indic, M.; van den Berg, B.; Movileanu, L. OccK channels from Pseudomonas aeruginosa exhibit diverse single-channel electrical signatures but conserved anion selectivity. Biochemistry 2012, 51, 2319–2330. [Google Scholar] [CrossRef] [Green Version]
- Pothula, K.R.; Kleinekathofer, U. Theoretical analysis of ion conductance and gating transitions in the OpdK (OccK1) channel. Analyst 2015, 140, 4855–4864. [Google Scholar] [CrossRef]
- Cheneke, B.R.; Indic, M.; van den Berg, B.; Movileanu, L. An outer membrane protein undergoes enthalpy- and entropy-driven transitions. Biochemistry 2012, 51, 5348–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheneke, B.R.; van den Berg, B.; Movileanu, L. Analysis of gating transitions among the three major open states of the OpdK channel. Biochemistry 2011, 50, 4987–4997. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Mohammad, M.M.; Patel, D.R.; Movileanu, L.; van den Berg, B. Structural insight into OprD substrate specificity. Nat. Struct. Mol. Biol. 2007, 14, 1108–1109. [Google Scholar] [CrossRef]
- Huang, H.; Hancock, R.E. The role of specific surface loop regions in determining the function of the imipenem-specific pore protein OprD of Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 3085–3090. [Google Scholar] [CrossRef] [Green Version]
- Samanta, S.; Scorciapino, M.A.; Ceccarelli, M. Molecular basis of substrate translocation through the outer membrane channel OprD of Pseudomonas aeruginosa. Phys. Chem. Chem. Phys. PCCP 2015, 17, 23867–23876. [Google Scholar] [CrossRef]
- Tamber, S.; Maier, E.; Benz, R.; Hancock, R.E. Characterization of OpdH, a Pseudomonas aeruginosa porin involved in the uptake of tricarboxylates. J. Bacteriol. 2007, 189, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothula, K.R.; Dhanasekar, N.N.; Lamichhane, U.; Younas, F.; Pletzer, D.; Benz, R.; Winterhalter, M.; Kleinekathofer, U. Single Residue Acts as Gate in OccK Channels. J. Phys. Chem. B 2017, 121, 2614–2621. [Google Scholar] [CrossRef]
- Chalhoub, H.; Pletzer, D.; Weingart, H.; Braun, Y.; Tunney, M.M.; Elborn, J.S.; Rodriguez-Villalobos, H.; Plesiat, P.; Kahl, B.C.; Denis, O.; et al. Mechanisms of intrinsic resistance and acquired susceptibility of Pseudomonas aeruginosa isolated from cystic fibrosis patients to temocillin, a revived antibiotic. Sci. Rep. 2017, 7, 40208. [Google Scholar] [CrossRef]
- Pongprayoon, P.; Beckstein, O.; Wee, C.L.; Sansom, M.S. Simulations of anion transport through OprP reveal the molecular basis for high affinity and selectivity for phosphate. Proc. Natl. Acad. Sci. USA 2009, 106, 21614–21618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modi, N.; Barcena-Uribarri, I.; Bains, M.; Benz, R.; Hancock, R.E.; Kleinekathofer, U. Tuning the affinity of anion binding sites in porin channels with negatively charged residues: Molecular details for OprP. ACS Chem. Biol. 2015, 10, 441–451. [Google Scholar] [CrossRef]
- Modi, N.; Barcena-Uribarri, I.; Bains, M.; Benz, R.; Hancock, R.E.; Kleinekathofer, U. Role of the central arginine R133 toward the ion selectivity of the phosphate specific channel OprP: Effects of charge and solvation. Biochemistry 2013, 52, 5522–5532. [Google Scholar] [CrossRef]
- Ganguly, S.; Kesireddy, A.; Barcena-Uribarri, I.; Kleinekathofer, U.; Benz, R. Conversion of OprO into an OprP-like Channel by Exchanging Key Residues in the Channel Constriction. Biophys. J. 2017, 113, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Jia, X.; Yang, S.; Xu, X.; Sun, K.; Li, C.; Yang, T.; Zhang, L. Heteroresistance to carbapenems in invasive Pseudomonas aeruginosa infections. Int. J. Antimicrob. Agents 2018, 51, 413–421. [Google Scholar] [CrossRef]
- Hirabayashi, A.; Kato, D.; Tomita, Y.; Iguchi, M.; Yamada, K.; Kouyama, Y.; Morioka, H.; Tetsuka, N.; Yagi, T. Risk factors for and role of OprD protein in increasing minimal inhibitory concentrations of carbapenems in clinical isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2017, 66, 1562–1572. [Google Scholar] [CrossRef] [Green Version]
- Agah Terzi, H.; Kulah, C.; Riza Atasoy, A.; Hakki Ciftci, I. Investigation of OprD Porin Protein Levels in Carbapenem-Resistant Pseudomonas aeruginosa Isolates. Jundishapur. J. Microbiol. 2015, 8, e25952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pragasam, A.K.; Raghanivedha, M.; Anandan, S.; Veeraraghavan, B. Characterization of Pseudomonas aeruginosa with discrepant carbapenem susceptibility profile. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buehrle, D.J.; Shields, R.K.; Chen, L.; Hao, B.; Press, E.G.; Alkrouk, A.; Potoski, B.A.; Kreiswirth, B.N.; Clancy, C.J.; Nguyen, M.H. Evaluation of the In Vitro Activity of Ceftazidime-Avibactam and Ceftolozane-Tazobactam against Meropenem-Resistant Pseudomonas aeruginosa Isolates. Antimicrob. Agents Chemother. 2016, 60, 3227–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kos, V.N.; McLaughlin, R.E.; Gardner, H.A. Identification of unique in-frame deletions in OprD among clinical isolates of Pseudomonas aeruginosa. Pathog. Dis. 2016, 74, ftw031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, C.Y.; Chen, S.S.; Hung, K.H.; Wu, H.M.; Hsueh, P.R.; Yan, J.J.; Wu, J.J. Overproduction of active efflux pump and variations of OprD dominate in imipenem-resistant Pseudomonas aeruginosa isolated from patients with bloodstream infections in Taiwan. BMC Microbiol. 2016, 16, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, R.; Sylvester, M.A.; Velez-Vega, C.; Tommasi, R.; Durand-Reville, T.F.; Miller, A.A. Whole-Cell-Based Assay To Evaluate Structure Permeation Relationships for Carbapenem Passage through the Pseudomonas aeruginosa Porin OprD. ACS Infect. Dis. 2017, 3, 310–319. [Google Scholar] [CrossRef]
- Shu, J.C.; Kuo, A.J.; Su, L.H.; Liu, T.P.; Lee, M.H.; Su, I.N.; Wu, T.L. Development of carbapenem resistance in Pseudomonas aeruginosa is associated with OprD polymorphisms, particularly the amino acid substitution at codon 170. J. Antimicrob. Chemother. 2017, 72, 2489–2495. [Google Scholar] [CrossRef] [Green Version]
- Richardot, C.; Plesiat, P.; Fournier, D.; Monlezun, L.; Broutin, I.; Llanes, C. Carbapenem resistance in cystic fibrosis strains of Pseudomonas aeruginosa as a result of amino acid substitutions in porin OprD. Int. J. Antimicrob. Agents 2015, 45, 529–532. [Google Scholar] [CrossRef]
- Choudhury, D.; Talukdar, A.D.; Choudhury, M.D.; Maurya, A.P.; Chanda, D.D.; Chakravorty, A.; Bhattacharjee, A. Carbapenem nonsusceptibility with modified OprD in clinical isolates of Pseudomonas aeruginosa from India. Indian J. Med. Microbiol. 2017, 35, 137–139. [Google Scholar] [CrossRef]
- Naenna, P.; Noisumdaeng, P.; Pongpech, P.; Tribuddharat, C. Detection of outer membrane porin protein, an imipenem influx channel, in Pseudomonas aeruginosa clinical isolates. Southeast Asian J. Trop. Med. Public Health 2010, 41, 614–624. [Google Scholar]
- Modi, N.; Ganguly, S.; Barcena-Uribarri, I.; Benz, R.; van den Berg, B.; Kleinekathofer, U. Structure, Dynamics, and Substrate Specificity of the OprO Porin from Pseudomonas aeruginosa. Biophys. J. 2015, 109, 1429–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, S.; D’Agostino, T.; Ghai, I.; Pathania, M.; Acosta Gutierrez, S.; Andrea Scorciapino, M.; Bodrenko, I.; Wagner, R.; van den Berg, B.; Winterhalter, M.; et al. How to Get Large Drugs through Small Pores? Exploiting the Porins Pathway in Pseudomonas Aeruginosa. Biophys. J. 2017, 112, 416a. [Google Scholar] [CrossRef]
- Siroy, A.; Molle, V.; Lemaitre-Guillier, C.; Vallenet, D.; Pestel-Caron, M.; Cozzone, A.J.; Jouenne, T.; De, E. Channel formation by CarO, the carbapenem resistance-associated outer membrane protein of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 4876–4883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussi, M.A.; Relling, V.M.; Limansky, A.S.; Viale, A.M. CarO, an Acinetobacter baumannii outer membrane protein involved in carbapenem resistance, is essential for L-ornithine uptake. FEBS Lett. 2007, 581, 5573–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahn, M.; D’Agostino, T.; Eren, E.; Basle, A.; Ceccarelli, M.; van den Berg, B. Small-Molecule Transport by CarO, an Abundant Eight-Stranded beta-Barrel Outer Membrane Protein from Acinetobacter baumannii. J. Mol. Biol. 2015, 427, 2329–2339. [Google Scholar] [CrossRef]
- Catel-Ferreira, M.; Coadou, G.; Molle, V.; Mugnier, P.; Nordmann, P.; Siroy, A.; Jouenne, T.; De, E. Structure-function relationships of CarO, the carbapenem resistance-associated outer membrane protein of Acinetobacter baumannii. J. Antimicrob. Chemother. 2011, 66, 2053–2056. [Google Scholar] [CrossRef] [Green Version]
- Catel-Ferreira, M.; Nehme, R.; Molle, V.; Aranda, J.; Bouffartigues, E.; Chevalier, S.; Bou, G.; Jouenne, T.; De, E. Deciphering the function of the outer membrane protein OprD homologue of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 3826–3832. [Google Scholar] [CrossRef] [Green Version]
- Zahn, M.; Bhamidimarri, S.P.; Basle, A.; Winterhalter, M.; van den Berg, B. Structural Insights into Outer Membrane Permeability of Acinetobacter baumannii. Structure 2016, 24, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, L.G.M.; Wallat, G.D.; Moynie, L.; Dhanasekar, N.N.; Aliouane, S.; Acosta-Gutierrez, S.; Pages, J.M.; Bolla, J.M.; Winterhalter, M.; Ceccarelli, M.; et al. MOMP from Campylobacter jejuni Is a Trimer of 18-Stranded beta-Barrel Monomers with a Ca(2+) Ion Bound at the Constriction Zone. J. Mol. Biol. 2016, 428, 4528–4543. [Google Scholar] [CrossRef] [Green Version]
- Bolla, J.M.; Saint, N.; Labesse, G.; Pages, J.M.; Dumas, C. Crystallization and preliminary crystallographic studies of MOMP (major outer membrane protein) from Campylobacter jejuni. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2349–2351. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Meitzler, J.C.; Huang, S.; Morishita, T. Sequence polymorphism, predicted secondary structures, and surface-exposed conformational epitopes of Campylobacter major outer membrane protein. Infect. Immun. 2000, 68, 5679–5689. [Google Scholar] [CrossRef] [Green Version]
- De, E.; Jullien, M.; Labesse, G.; Pages, J.M.; Molle, G.; Bolla, J.M. MOMP (major outer membrane protein) of Campylobacter jejuni; a versatile pore-forming protein. FEBS Lett. 2000, 469, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, S.; Bodrenko, I.; Acosta-Gutierrez, S.; D’Agostino, T.; Pathania, M.; Ghai, I.; Schleberger, C.; Bumann, D.; Wagner, R.; Winterhalter, M.; et al. Getting Drugs through Small Pores: Exploiting the Porins Pathway in Pseudomonas aeruginosa. ACS Infect. Dis. 2018, 4, 1519–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citak, F.; Ghai, I.; Rosenkotter, F.; Benier, L.; Winterhalter, M.; Wagner, R. Probing transport of fosfomycin through substrate specific OprO and OprP from Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2018, 495, 1454–1460. [Google Scholar] [CrossRef] [PubMed]
- Pautsch, A.; Schulz, G.E. Structure of the outer membrane protein A transmembrane domain. Nat. Struct. Biol. 1998, 5, 1013–1017. [Google Scholar] [CrossRef]
- Vandeputte-Rutten, L.; Kramer, R.A.; Kroon, J.; Dekker, N.; Egmond, M.R.; Gros, P. Crystal structure of the outer membrane protease OmpT from Escherichia coli suggests a novel catalytic site. EMBO J. 2001, 20, 5033–5039. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; van den Berg, B. Crystal structure of the bacterial nucleoside transporter Tsx. EMBO J. 2004, 23, 3187–3195. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, B.; Black, P.N.; Clemons, W.M., Jr.; Rapoport, T.A. Crystal structure of the long-chain fatty acid transporter FadL. Science 2004, 304, 1506–1509. [Google Scholar] [CrossRef] [Green Version]
- Cowan, S.W.; Schirmer, T.; Rummel, G.; Steiert, M.; Ghosh, R.; Pauptit, R.A.; Jansonius, J.N.; Rosenbusch, J.P. Crystal structures explain functional properties of two E. coli porins. Nature 1992, 358, 727–733. [Google Scholar] [CrossRef]
- Monier, R. Wilhelm Bernhard (1920–1978). Biomedicine 1979, 30, 16. [Google Scholar]
- Meyer, J.E.; Hofnung, M.; Schulz, G.E. Structure of maltoporin from Salmonella typhimurium ligated with a nitrophenyl-maltotrioside. J. Mol. Biol. 1997, 266, 761–775. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, T.; Keller, T.A.; Wang, Y.F.; Rosenbusch, J.P. Structural basis for sugar translocation through maltoporin channels at 3.1 A resolution. Science 1995, 267, 512–514. [Google Scholar] [CrossRef]
- Chimento, D.P.; Mohanty, A.K.; Kadner, R.J.; Wiener, M.C. Substrate-induced transmembrane signaling in the cobalamin transporter BtuB. Nat. Struct. Biol. 2003, 10, 394–401. [Google Scholar] [CrossRef]
- Buchanan, S.K.; Smith, B.S.; Venkatramani, L.; Xia, D.; Esser, L.; Palnitkar, M.; Chakraborty, R.; van der Helm, D.; Deisenhofer, J. Crystal structure of the outer membrane active transporter FepA from Escherichia coli. Nat. Struct. Biol. 1999, 6, 56–63. [Google Scholar] [CrossRef]
- Thakur, A.K.; Larimi, M.G.; Gooden, K.; Movileanu, L. Aberrantly Large Single-Channel Conductance of Polyhistidine Arm-Containing Protein Nanopores. Biochemistry 2017, 56, 4895–4905. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Prajapati, J.D.; Kleinekathofer, U.; Winterhalter, M. Dynamic interaction of fluoroquinolones with magnesium ions monitored using bacterial outer membrane nanopores. Chem. Sci. 2020, 11, 10344–10353. [Google Scholar] [CrossRef]
- Arunmanee, W.; Pathania, M.; Solovyova, A.S.; Le Brun, A.P.; Ridley, H.; Basle, A.; van den Berg, B.; Lakey, J.H. Gram-negative trimeric porins have specific LPS binding sites that are essential for porin biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E5034–E5043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eren, E.; Vijayaraghavan, J.; Liu, J.; Cheneke, B.R.; Touw, D.S.; Lepore, B.W.; Indic, M.; Movileanu, L.; van den Berg, B. Substrate specificity within a family of outer membrane carboxylate channels. PLoS Biol. 2012, 10, e1001242. [Google Scholar] [CrossRef] [Green Version]
- Winterhalter, M.; Hilty, C.; Bezrukov, S.M.; Nardin, C.; Meier, W.; Fournier, D. Controlling membrane permeability with bacterial porins: Application to encapsulated enzymes. Talanta 2001, 55, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Danelon, C.; Laio, A.; Parrinello, M. Microscopic Mechanism of Antibiotics Translocation through a Porin. Biophys. J. 2004, 87, 58–64. [Google Scholar] [CrossRef]
- Nestorovich, E.M.; Sugawara, E.; Nikaido, H.; Bezrukov, S.M. Pseudomonas aeruginosa porin OprF: Properties of the channel. J. Biol. Chem. 2006, 281, 16230–16237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahendran, K.R.; Chimerel, C.; Mach, T.; Winterhalter, M. Antibiotic translocation through membrane channels: Temperature-dependent ion current fluctuation for catching the fast events. Eur. Biophys. J. 2009, 38, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Several systems pertaining to antibiotic/antimicrobial resistance that Gram-negative bacteria may be utilized as an overview (structural interpretation) and antibiotic/antimicrobial resistance mechanism correlated with outer membrane proteins modification. 1. Solute transport via outer membrane proteins, 2. Loss of functional outer membrane proteins, 3. Modification of outer membrane proteins, 4. Expression of constricted outer membrane proteins, 5. Partitioning of hydrophobic molecules via the outer layer.
Figure 1.
Several systems pertaining to antibiotic/antimicrobial resistance that Gram-negative bacteria may be utilized as an overview (structural interpretation) and antibiotic/antimicrobial resistance mechanism correlated with outer membrane proteins modification. 1. Solute transport via outer membrane proteins, 2. Loss of functional outer membrane proteins, 3. Modification of outer membrane proteins, 4. Expression of constricted outer membrane proteins, 5. Partitioning of hydrophobic molecules via the outer layer.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ghai, I. A Barrier to Entry: Examining the Bacterial Outer Membrane and Antibiotic Resistance. Appl. Sci. 2023, 13, 4238. https://doi.org/10.3390/app13074238
AMA Style
Ghai I. A Barrier to Entry: Examining the Bacterial Outer Membrane and Antibiotic Resistance. Applied Sciences. 2023; 13(7):4238. https://doi.org/10.3390/app13074238
Chicago/Turabian StyleGhai, Ishan. 2023. "A Barrier to Entry: Examining the Bacterial Outer Membrane and Antibiotic Resistance" Applied Sciences 13, no. 7: 4238. https://doi.org/10.3390/app13074238
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.