
Field-Testing a Proteomics-Derived Machine-Learning Model for Predicting Coral Bleaching Susceptibility

Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
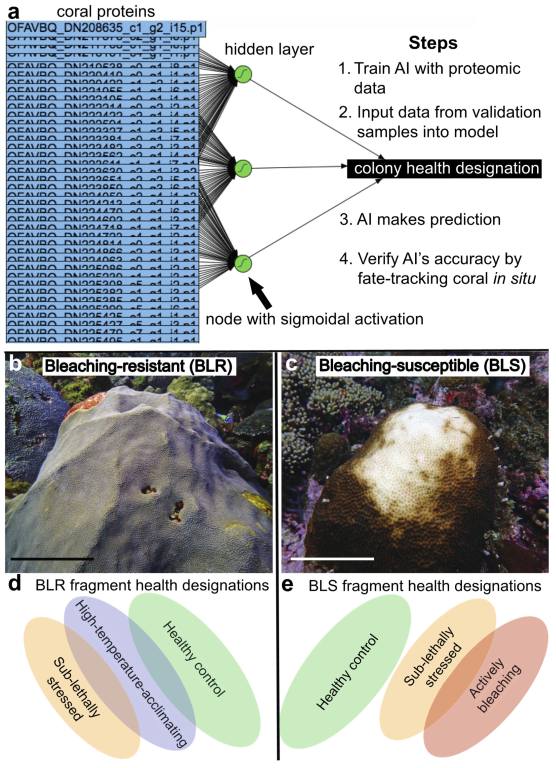
2.1. Approach and Terminology
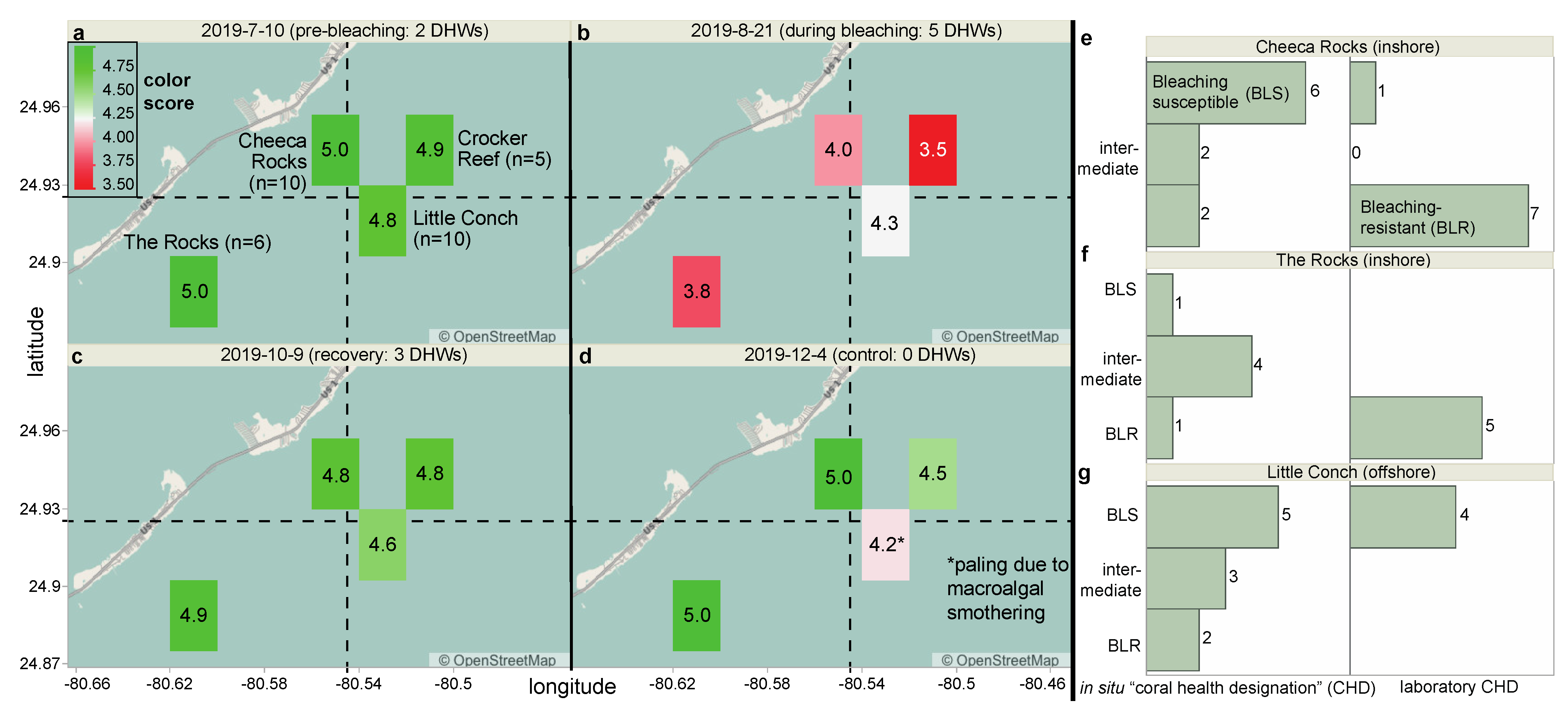
2.2. The Experiment

2.3. Field Coral Fate-Tracking and Sampling
2.4. Proteome Profiling
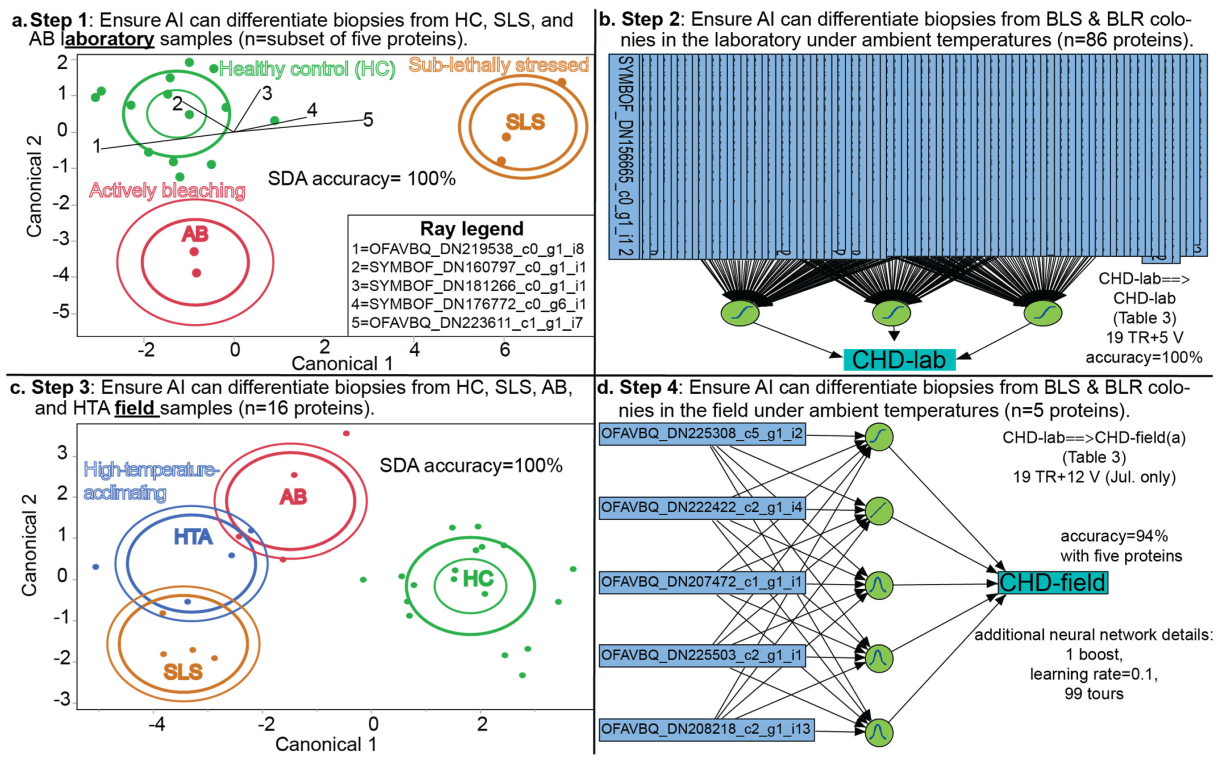
2.5. Proteomic Predictive Modeling
2.5.1. Lab-Trained Models
2.5.2. Field-Trained Models
3. Results and Discussion
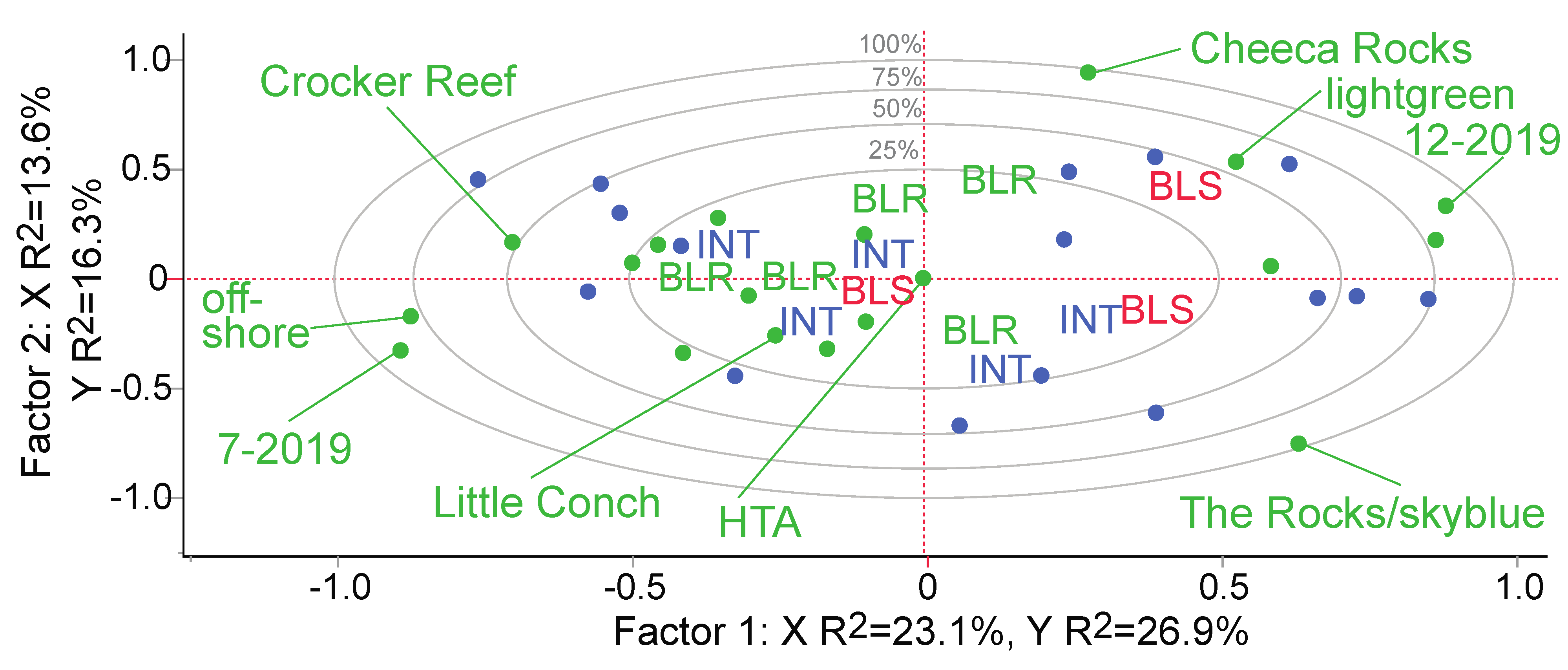
3.1. Overview of the Field Coral Proteomic Dataset

{kind=link}
{kind=link}
{kind=link}
{kind=link}
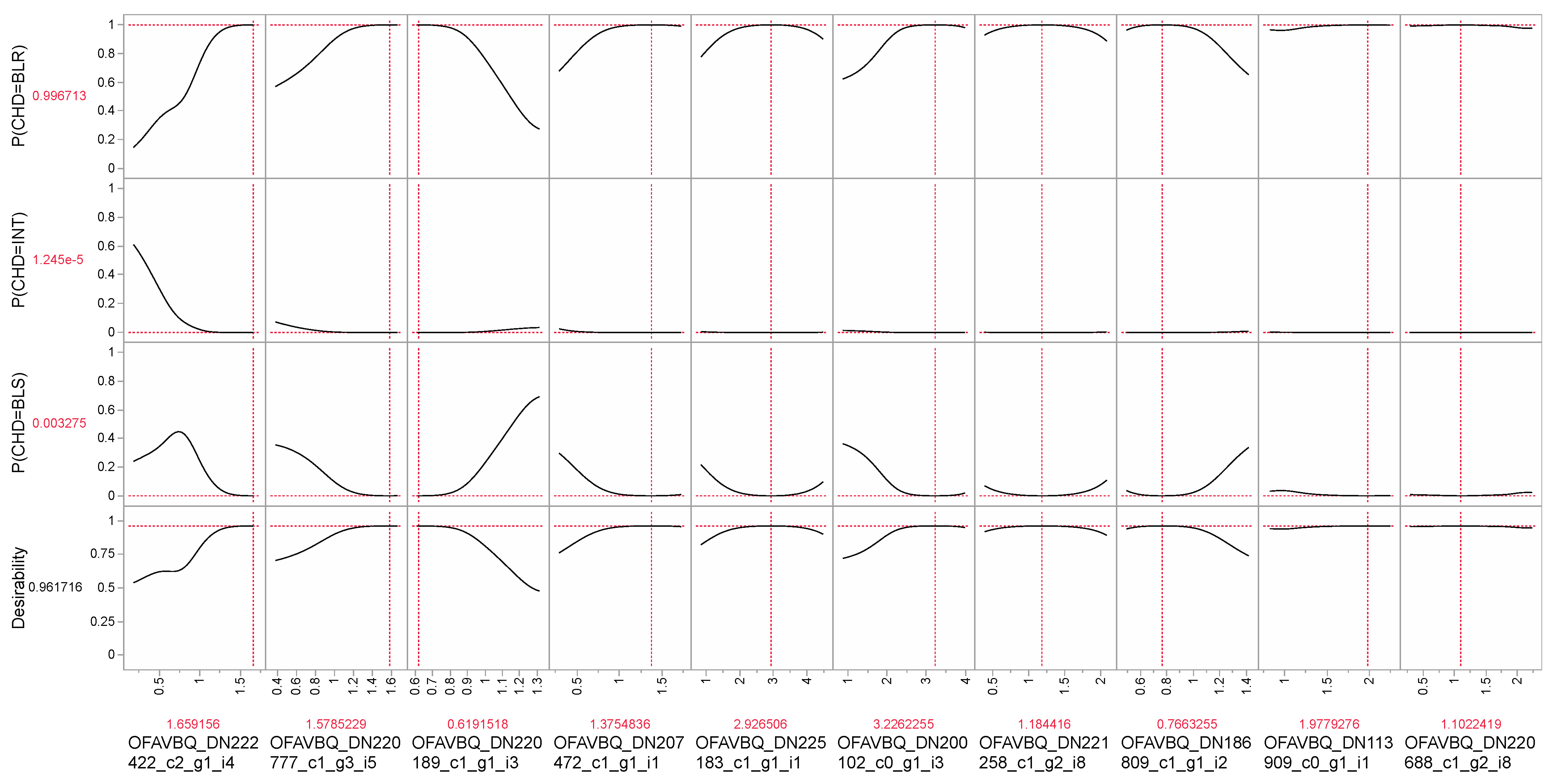{kind=link}
| Model Abbreviation | CHD-Lab→ CHD-Lab [11] | CHD-Lab→CHD-Field(a) | CHD-Lab→CHD-Field(c) |
|---|---|---|---|
| Model prediction goal | CHD-lab | CHD-field | CHD-field |
| Training sample size (lab corals) | 14 | 19 | 19 |
| Validation data type (#samples) | Lab corals (5) | Field corals (12) | Field corals (24) |
| Validation data months | Not applicable | Jul. | Jul. and Aug. |
| Training proteins (#) | 86 | 5 | 4 |
| Model details | TanH(3)-Boost(7) a,b | TanH(1)-Linear(1)-Gaussian(3) c | TanH(2)-Linear(2)-Gaussian(4) |
| Accuracy (%) ± std. dev. | 100 ± 1 (n = 20) | 86 ± 10 (n = 40) | 82 ± 7 (n = 40) |
3.2. Coral Health Predictions Overview
3.3. Coral Health Predictions from Lab Data
3.4. Coral Health Predictions from Field Data
3.5. Other Models
3.6. Caveats and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morikawa, M.K.; Palumbi, S.R. Using naturally occurring climate resilient corals to construct bleaching-resistant nurseries. Proc. Natl. Acad. Sci. USA 2019, 116, 10586–10591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.E. Coral bleaching: Causes and consequences. Coral Reefs 1997, 16s, 129–138. [Google Scholar] [CrossRef]
- Grottoli, A.G.; Toonen, R.J.; van Woesik, R.; Thurber, R.V.; Warner, M.E.; McLachlan, R.H.; Price, J.T.; Bahr, K.D.; Baums, I.B.; Castillo, K.D.; et al. Increasing comparability among coral bleaching experiments. Ecol. Appl. 2021, 31, e02262. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.B.; Lin, C. Leveraging the power of artificial intelligence to identify resilient reef corals. Platax 2022, 19, 43–57. [Google Scholar]
- McLachlan, R.H.; Price, J.; Solomon, S.; Grottoli, A.G. Thirty years of coral heat-stress experiments: A review of methods. Coral Reefs 2020, 39, 885–902. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Dupiol, J.; Ladrière, O.; Meistertzheim, A.L.; Fouré, L.; Adjeroud, M.; Mitta, G. Physiological responses of the scleractinian coral Pocillopora damicornis to bacterial stress from Vibrio corallilyticus. J. Exp. Biol. 2011, 214, 1533–1545. [Google Scholar] [CrossRef] [Green Version]
- Downs, C.A.; Mueller, E.; Phillips, S.; Fauth, J.E.; Woodley, C.M. A molecular biomarker system for assessing the health of coral (Montastrea faveolata) during heat stress. Mar. Biotechnol. 2020, 2, 533–544. [Google Scholar] [CrossRef]
- Parkinson, J.E.; Bartels, E.; Devlin-Durante, M.K.; Lustic, C.; Nedimyer, K.; Schopmeyer, S.; Lirman, D.; LaJeunesse, T.C.; Baums, I.B. Extensive transcriptional variation poses a challenge to thermal stress biomarker development for endangered coral. Mol. Ecol. 2018, 27, 1103–1119. [Google Scholar] [CrossRef]
- Aguilar, C.; Enochs, I.C.; Manzello, D.P.; Kolodziej, G.; Carlton, R.; Mayfield, A.B. Transcriptome profiling of thermotolerant corals of the Upper Florida Keys. Mol. Ecol. under review.
- Mayfield, A.B.; Aguilar, C.; Enochs, I.C.; Kolodziej, G.; Manzello, D.P. Shotgun proteomics of thermally challenged Caribbean reef corals. Front. Mar. Sci. 2021, 8, 660153. [Google Scholar] [CrossRef]
- Mayfield, A.B. Machine-learning-based proteomic predictive modeling with thermally-challenged Caribbean reef corals. Diversity 2022, 14, 33. [Google Scholar] [CrossRef]
- Manzello, D.P.; Matz, M.V.; Enochs, I.C.; Valentino, L.; Carlton, R.D.; Kolodziej, G.; Serrano, X.; Towle, E.K.; Jankulak, M. Role of host genetics and heat-tolerant algal symbionts in sustaining populations of the endangered coral Orbicella faveolata in the Florida Keys with ocean warming. Glob. Chang. Biol. 2019, 25, 1016–1031. [Google Scholar] [CrossRef] [PubMed]
- Gintert, B.E.; Manzello, D.P.; Enochs, I.C.; Kolodziej, G.; Carlton, R.; Gleason, A.C.R.; Gracias, N. Marked annual coral bleaching resilience of an inshore patch reef in the Florida Keys: A nugget of hope, aberrance, or last man standing? Coral Reefs 2018, 37, 533–547. [Google Scholar] [CrossRef] [Green Version]
- Siebeck, U.; Marshall, N.; Klüter, A.; Hoegh-Guldberg, O. Monitoring coral bleaching using a colour (sp.) reference card. Coral Reefs 2006, 25, 453–460. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Wang, L.H.; Tang, P.C.; Fan, T.Y.; Hsiao, Y.Y.; Tsai, C.L.; Chen, C.S. Assessing the impacts of experimentally elevated temperature on the biological composition and molecular chaperone gene expression of a reef coral. PLoS ONE 2011, 6, e26529. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.B.; Tsai, S.; Lin, C. The Coral Hospital. Biopreserv. Biobank 2019, 17, 355–369. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Chen, Y.J.; Lu, C.Y.; Chen, C.S. Exploring the environmental physiology of the Indo-Pacific reef coral Seriatopora hystrix using differential proteomics. Open J. Mar. Sci. 2018, 8, 223–252. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C. The molecular ecophysiology of closely related pocilloporid corals of New Caledonia. Platax 2017, 14, 1–45. [Google Scholar]
- Mayfield, A.B.; Wang, Y.B.; Chen, C.S.; Chen, S.H.; Lin, C.Y. Compartment-specific transcriptomics in a reef-building coral exposed to elevated temperatures. Mol. Ecol. 2014, 23, 5816–5830. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.H.; Lee, H.H.; Fang, L.S.; Mayfield, A.B.; Chen, C.S. Normal fatty acid and phospholipid synthesis are prerequisites for the cell cycle of Symbiodinium and their endosymbiosis with sea anemones. PLoS ONE 2013, 8, e72486. [Google Scholar]
- Bay, R.A.; Rose, N.H.; Logan, C.A.; Palumbi, S.R. Genomic models predict successful coral adaptation if future ocean warming rates are reduced. Sci. Adv. 2017, 3, e1701413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, Z.; Mocellin, V.J.L.; Morris, L.A.; Cantin, N.; Shepherd, J.; Sarre, L.; Peng, J.; Liao, Y.; Pickrell, J.; Przeworski, M. Population genetics of the cooral (sp.). Acropora millepora: Toward genomic prediction of bleaching. Science 2020, 369, eaba4674. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.N.F.; Dilworth, J.; Christian, M.H.; Jones, D.; Quinn, R.A.; Drury, C. Metabolomic signatures of coral bleaching history. Nat. Ecol. Evol. 2021, 5, 495–503. [Google Scholar] [PubMed]
- van Hooidonk, R.; Maynard, J.; Tamelander, J.; Gove, J.; Ahmadia, G.; Raymundo, L.; Williams, G.; Heron, S.F. Local-scale projections of coral reef futures and implications of the Paris Agreement. Sci. Rep. 2016, 6, 39666. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Dempsey, A.C.; Chen, C.S.; Lin, C. Expediting the search for climate-resilient reef corals in the Coral Triangle with artificial intelligence. Appl. Sci. 2022, 12, 12955. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Dempsey, A.C.; Chen, C.S. Predicting the abundance of corals from simple environmental predictors with a machine-learning approach. Platax 2022, 19, 43–57. [Google Scholar]
- Maynard, J.A.; Anthony, K.R.N.; Marshall, P.A.; Masiri, I. Major bleaching events can lead to increased thermal tolerance in corals. Mar. Biol. 2008, 155, 173–182. [Google Scholar] [CrossRef]
- Liu, G.; Heron, S.F.; Eakin, C.M.; Muller-Karger, F.E.; Vega-Rodriguez, M.; Guild, L.S.; De La Cour, J.L.; Geiger, E.F.; Skirving, W.J.; Burgess, T.F.R.; et al. Reef-scale thermal stress monitoring of coral ecosystems: New 5-km Global Products from NOAA Coral Reef Watch. Remote Sens. 2014, 6, 11579–11606. [Google Scholar] [CrossRef] [Green Version]
- McClanahan, T.R.; Darling, E.S.; Maina, J.M.; Muthiga, N.A.; N’agata, S.D.; Jupiter, S.D.; Arthur, R.; Wilson, S.K.; Mangubhai, S.; Nand, Y.; et al. Temperature patterns and mechanisms influencing coral bleaching during the 2016 El Niño. Nat. Clim. Chang. 2019, 9, 845–851. [Google Scholar] [CrossRef]
- Logan, C.A.; Dunne, J.P.; Eakin, C.M.; Donner, S.D. Incorporating adaptive responses into future projections of coral bleaching. Glob. Chang. Biol. 2014, 20, 125–139. [Google Scholar] [CrossRef]
- Mayfield, A.B. Multi-macromolecular extraction from endosymbiotic anthozoans. In Lipidomics; Bhattacharya, S., Ed.; SpringerS: Berlin/Heidelberg, Germany, 2023. [Google Scholar]
- Desoubeaux, G.; Chauvin, D.; Piqueras, M.C.; Bronson, E.; Bhattacharya, S.K.; Sirpenski, G.; Sirpenski, G.; Bailly, E.; Cray, C. Translational proteomic study to address host protein changes during aspergillosis. Dev. Neurobiol. 2018, 13, e0200843. [Google Scholar]
- Musada, G.R.; Dvoriantchikova, G.; Myer, C.; Ivanov, D.; Bhattacharya, S.K.; Hackam, A.S. The effect of extrinsic Wnt/β-catenin signaling in Muller glia on retinal ganglion cell neurite growth. Dev. Neurobiol. 2020, 80, 98–110. [Google Scholar]




| Sample Name | Reef of Origin | Shelf | Treatment (Temp.-Time) | Genotype | CHD Field | CHD Lab | FHD Lab | Protein (µg) | iTRAQ Tag | iTRAQ Batch |
|---|---|---|---|---|---|---|---|---|---|---|
| Normalizer | mix of all | mix of both | mix of all | mix of all | NA | NA | NA | 22 | 113 | A |
| B5-7 a | Cheeca Rocks | inshore | 30-5 | lightyellow | BLS | BLR | HC | 22 | 114 | A |
| C5-1 | Little Conch | offshore | 30-5 | black(e) | BLS | BLS | HC | 22 | 115 | A |
| B5-4 | Cheeca Rocks | inshore | 33-5 | lightyellow | BLS | BLR | SLS | 22 | 116 | A |
| A2-2 | The Rocks | inshore | 30-31 | skyblue * | BLR | BLS | HC | 22 | 117 | A |
| A4-5 | The Rocks | inshore | 32-31 | skyblue * | INT | BLR | HTA | 22 | 118 | A |
| B3-1 | Cheeca Rocks | inshore | 32-31 | black(c) | BLR | BLR | HTA | 22 | 119 | A |
| C5-2 | Little Conch | offshore | 32-31 | black(e) | BLS | BLS | AB | 22 | 121 | A |
| Normalizer | mix of all | mix of both | mix of all | mix of all | NA | NA | 22 | 113 | B | |
| A4-1 | The Rocks | inshore | 30-5 | skyblue * | INT | BLR | HC | 22 | 114 | B |
| C2-2 | Little Conch | offshore | 30-5 | black(b) | BLS | BLS | HC | 22 | 115 | B |
| D5-2 | Cheeca Rocks | inshore | 30-5 | grey60 | BLS | BLR | HC | 22 | 116 | B |
| D6-6 | Cheeca Rocks | inshore | 33-5 | grey60 | BLS | ID | SLS | 22 | 117 | B |
| A4-8 | The Rocks | inshore | 30-31 | skyblue * | INT | BLR | HC | 22 | 118 | B |
| C5-7 | Little Conch | offshore | 30-31 | black(e) | BLS | BLS | HC | 22 | 119 | B |
| D5-3 | Cheeca Rocks | inshore | 32-31 | grey60 | BLS | BLR | HTA | 22 | 121 | B |
| Normalizer | mix of all | mix of both | mix of all | mix of all | NA | NA | 22 | 113 | C | |
| A4-7 | The Rocks | inshore | 33-5 | skyblue * | INT | BLR | HTA | 22 | 114 | C |
| C5-8 | Little Conch | offshore | 33-5 | black(e) | BLS | BLS | AB | 22 | 115 | C |
| D5-5 | Cheeca Rocks | inshore | 33-5 | grey60 | BLS | BLR | HTA | 22 | 116 | C |
| B5-1 | Cheeca Rocks | inshore | 30-31 | lightyellow | BLS | BLR | HC | 22 | 117 | C |
| D4-8 | Cheeca Rocks | inshore | 30-31 | grey60 | BLS | BLR | HC | 22 | 118 | C |
| D5-8 | Cheeca Rocks | inshore | 30-31 | grey60 | BLS | BLR | HC | 22 | 119 | C |
| B5-2 | Cheeca Rocks | inshore | 32-31 | lightyellow | BLS | BLR | SLS | 22 | 121 | C |
| Sample Name | Reef of Origin | Shelf | Sampling Date | Genotype | CHD Field | FHD Field | Protein (µg) | iTRAQ Tag |
|---|---|---|---|---|---|---|---|---|
| Batch A | ||||||||
| Normalizer-A | mix of all | mix of both | mix of all | mix of all | NA | NA | 58 | 113 |
| Cheeca-3132-7/19 | Cheeca Rocks | inshore | 2019-7 | black(a) | BLR | HC | 70 | 114 |
| Cheeca-3916-7/19 | Cheeca Rocks | inshore | 2019-7 | lightgreen | BLS | HC | 70 | 115 |
| Cheeca-3939-7/19 | Cheeca Rocks | inshore | 2019-7 | darkred | INT | HC | 70 | 116 |
| CR-3165-7/19 | Crocker Reef | offshore | 2019-7 | black(r) | BLR | HC | 70 | 117 |
| CR-3679-7/19 | Crocker Reef | offshore | 2019-7 | not genotyped | INT | HC | 70 | 118 |
| CR-3986-7/19 | Crocker Reef | offshore | 2019-7 | black(aa) | BLS | HC | 70 | 119 |
| Batch B | ||||||||
| Normalizer-B | mix of all | mix of both | mix of all | mix of all | NA | NA | 58 | 113 |
| LC-3694-7/19 | Little Conch | offshore | 2019-7 | black(f) | BLS | HC | 70 | 114 |
| LC-3921-7/19 | Little Conch | offshore | 2019-7 | black(i) | BLR | HC | 70 | 115 |
| LC-3989-7/19 | Little Conch | offshore | 2019-7 | black(g) | INT | HC | 70 | 116 |
| Rocks-3105-7/19 | The Rocks | inshore | 2019-7 | skyblue | BLS | HC | 70 | 117 |
| Rocks-3148-7/19 | The Rocks | inshore | 2019-7 | skyblue | INT | HC | 70 | 118 |
| Rocks-3906-7/19 | The Rocks | inshore | 2019-7 | skyblue | BLR | HC | 70 | 119 |
| Batch C | ||||||||
| Normalizer-C | mix of all | mix of both | mix of all | mix of all | NA | NA | 58 | 113 |
| Cheeca-3132-8/19 | Cheeca Rocks | inshore | 2019-8 | black(a) | BLR | HTA | 70 | 114 |
| Cheeca-3916-8/19 | Cheeca Rocks | inshore | 2019-8 | lightgreen | BLS | AB | 70 | 115 |
| Cheeca-3939-8/19 | Cheeca Rocks | inshore | 2019-8 | darkred | INT | SLS | 70 | 116 |
| LC-3694-8/19 | Little Conch | offshore | 2019-8 | black(f) | BLS | AB | 70 | 117 |
| LC-3921-8/19 | Little Conch | offshore | 2019-8 | black(i) | BLR | HTA | 70 | 118 |
| LC-3989-8/19 | Little Conch | offshore | 2019-8 | black(g) | INT | SLS | 70 | 119 |
| Batch D | ||||||||
| Normalizer-D | mix of all | mix of both | mix of all | mix of all | NA | NA | 58 | 113 |
| CR-3165-8/19 | Crocker Reef | offshore | 2019-8 | black(r) | BLR | HTA | 70 | 114 |
| CR-3679-8/19 | Crocker Reef | offshore | 2019-8 | not genotyped | INT | SLS | 70 | 115 |
| CR-3986-8/19 | Crocker Reef | offshore | 2019-8 | black(aa) | BLS | AB | 70 | 116 |
| Rocks-3105-8/19 | The Rocks | inshore | 2019-8 | skyblue | BLS | AB | 70 | 117 |
| Rocks-3148-8/19 | The Rocks | inshore | 2019-8 | skyblue | INT | SLS | 70 | 118 |
| Rocks-3906-8/19 | The Rocks | inshore | 2019-8 | skyblue | BLR | HTA | 70 | 119 |
| Batch E | ||||||||
| Normalizer-E | mix of all | mix of both | mix of all | mix of all | NA | NA | 58 | 113 |
| Cheeca-3132-12/19 | Cheeca Rocks | inshore | 2019-12 | black(a) | BLR | HC | 70 | 114 |
| Cheeca-3916-12/19 | Cheeca Rocks | inshore | 2019-12 | lightgreen | BLS | HC | 70 | 115 |
| Cheeca-3939-12/19 | Cheeca Rocks | inshore | 2019-12 | darkred | INT | HC | 70 | 116 |
| Rocks-3105-12/19 | The Rocks | inshore | 2019-12 | skyblue | BLS | HC | 70 | 121 * |
| Rocks-3148-12/19 | The Rocks | inshore | 2019-12 | skyblue | INT | HC | 70 | 118 |
| Rocks-3906-12/19 | The Rocks | inshore | 2019-12 | skyblue | BLR | HC | 70 | 119 |
| Batch F | ||||||||
| Normalizer-F | mix of all | mix of both | mix of all | mix of all | NA | 58 | 113 | |
| CR-3165-12/19 | Crocker Reef | offshore | 2019-12 | blackI | BLR | HC | 70 | 114 |
| CR-3679-12/19 | Crocker Reef | offshore | 2019-12 | not genotyped | INT | HC | 70 | 115 |
| CR-3986-12/19 | Crocker Reef | offshore | 2019-12 | black(aa) | BLS | SCTLD | 70 | 116 |
| LC-3694-12/19 | Little Conch | offshore | 2019-12 | black(f) | BLS | SLS | 70 | 117 |
| LC-3921-12/19 | Little Conch | offshore | 2019-12 | black(i) | BLR | HC | 70 | 118 |
| LC-3989-12/19 | Little Conch | offshore | 2019-12 | black(g) | INT | HC | 70 | 119 |
| Accession | Protein Name | Top BLAST Hit Accession | Protein Function | CHD-Lab→CHD-Field(a) Model Total Effect |
|---|---|---|---|---|
| Proteins featured in model CHD-lab→CHD-field(a) (Table 3) | ||||
| OFAVBQ_DN222422_c2_g1_i4 a | DELTA-actitoxin | XP_044182609.1 | prey capture | 0.44 |
| OFAVBQ_DN225308_c5_g1_i2 | E3 ubiquitin protein ligase | XP_020616505.1 | protein degradation | 0.25 |
| OFAVBQ_DN208218_c2_g1_i13 | histone-lysine N- methyltransferase SETD1B | XP_020616327.1 | transcription | 0.20 |
| OFAVBQ_DN225503_c2_g1_i1 b | concanavalin A-like lectin/glucanase | XP_020611325.1 | cell binding/immunity | 0.19 |
| OFAVBQ_DN207472_c1_g1_i1 | histone H2A | XP_020604133.1 | chromatin | 0.18 |
| Other proteins of interest | Finding | |||
| OFAVBQ_DN223604_c1_g1_i2 c | unknown | XP_020618813.1 | unknown | 50% lower in Aug. |
| SYMBOF_DN75265_c0_g1_i2 | chromosome segregation protein SMC | CAI3980295.1 | cell division | see Table 5. |
| OFAVBQ_DN221258_c1_g2_i8 | F-actin-methionine sulfoxide oxidase | XP_020606640.1 | molecular trafficking | see Table 5. |
| OFAVBQ_DN220777_c1_g3_i5 | titin-like | XP_020600883.1 | various d | see Table 5. |
| OFAVBQ_DN220189_c1_g1_i3 | unknown | XP_020619646.1 | unknown | see Table 5. |
| OFAVBQ_DN186152_c0_g1_i1 | transcription factor AP1 | XP_020624642.1 | transcription | see Table 5. |
| Model Input Type | Field Corals Sampled across Bleaching Event in 2019 (Figure 1) | ||||
|---|---|---|---|---|---|
| Model Name | CHD-Field→ CHD-Field(a) | CHD-Field→ CHD-Field(b) | CHD-Field→CHD-Field(c) | CHD-Field→CHD-Field(e) | CHD-Field→CHD-Field(f) |
| Training # | 19 | 18 | 11 | 12 | 15 |
| Training data months | Jul., Aug., Dec. | Jul. and Dec. | Jul. and Dec. | Jul. | Jul. and Aug. |
| Validation # | 8 | 12 | 4 | 12 | 6 |
| Validation data months | Jul., Aug., Dec. | Aug. | Jul. and Dec. | Aug. | Jul. and Aug. |
| Test sample # | 3 | NA | 3 | Not applicable (NA) | 3 |
| Test data months | Jul., Aug., Dec. | NA | Jul. and Dec. | NA | Jul. and Aug. |
| Training proteins | 16 | 16 | 16 | 28 | 28 |
| Model type #1 (accuracy [%]) | SVM: 90%a | Naïve Bayes: 94% | Gen-reg lasso: 95% | NN: 95 ± 6% (n = 20) | SVM: 94% |
| Model type #2 (accuracy [%]) | NN: 84 ± 9% (n = 40) a | NN: 87 ± 13% (n = 40) a | NN: 87 ± 14% (n = 20) | Naïve Bayes: 92% | NA |
| Most important protein(s) | OFAVBQ_DN221258_ c1_g2_i8 | OFAVBQ_DN222422_ c2 _g1_i4 | OFAVBQ_DN222422_c2_g1_i4 SYMBOF_DN75265_c0_g1_i2 | OFAVBQ_DN186152_ c0_g1_i1 b OFAVBQ_DN220777_c1_g3_i5 | OFAVBQ_DN222422_c2_g1_i4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayfield, A.B.; Lin, C. Field-Testing a Proteomics-Derived Machine-Learning Model for Predicting Coral Bleaching Susceptibility. Appl. Sci. 2023, 13, 1718. https://doi.org/10.3390/app13031718
Mayfield AB, Lin C. Field-Testing a Proteomics-Derived Machine-Learning Model for Predicting Coral Bleaching Susceptibility. Applied Sciences. 2023; 13(3):1718. https://doi.org/10.3390/app13031718
Chicago/Turabian StyleMayfield, Anderson B., and Chiahsin Lin. 2023. "Field-Testing a Proteomics-Derived Machine-Learning Model for Predicting Coral Bleaching Susceptibility" Applied Sciences 13, no. 3: 1718. https://doi.org/10.3390/app13031718