
Salinity Inhibition in Thermophilic Anaerobic Digestion of Organic Waste
by

Gregor Drago Zupančič
1,*,
Mario Panjičko
1,
Romana Marinšek Logar
2,
Lea Lavrič
3,
Maša Zorec
2 and
Lijana Fanedl
2 1
Croteh Ltd., Avenue Dubrovnik 15, HR10020 Zagreb, Croatia
2
Department for Microbiology, Biotechnical Faculty, University of Ljubljana, Jamnikarjeva 101, SI-1000 Ljubljana, Slovenia
3
KOTO Ltd., Agrokombinatska 80, SI-1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Appl. Sci. 2023, 13(11), 6590; https://doi.org/10.3390/app13116590
Submission received: 19 April 2023
/
Revised: 24 May 2023
/
Accepted: 25 May 2023
/
Published: 29 May 2023
(This article belongs to the Special Issue Production, Treatment, Utilization and Future Opportunities of Biogas)
Abstract
:Anaerobic digestion, despite its preferable use as a treatment for high organic matter polluted waste streams, is susceptible to inhibitors, salt included. Therefore, two different experiments were conducted to observe the responses of bacterial and archaeal communities to hypersaline environments. In the first experiment, salt was added gradually, while in the second experiment, salt was added rapidly (so-called salt shocks were performed). The results of the gradual addition of salt showed a recovery of methane production after the salt concentration decreased. The NaCl concentration of 28.2 g/L seems to be the limit between stable operation and occurrence inhibition. The specific biogas production varied between 0.490 and 0.562 m3/kgtCOD during the stepwise salt addition, depending on the salt concentration, while the maximal achieved COD removal was 79.8%. The results of the rapid salt addition showed good recovery of the bacterial community, while a reduction of salt-sensitive species was observed in the archaeal community. The trend of specific biogas production during rapid salt addition was stable with an average value of 0.590 m3/kgtCOD, and it was observed that higher concentrations of up to 39.4 g/L of NaCl were tolerated. The maximum COD removal achieved during rapid salt addition was 83.1%. In conclusion, certain bacterial and archaeal communities were well-adapted to the hypersaline environment and remained active during the anaerobic digestion of substrates with high salt concentration.
1. Introduction
Anaerobic digestion (AD) is a preferred treatment for waste streams heavily loaded with organic matter, allowing the waste volume to be reduced and energy to be recovered at the same time. According to current biogas trends in the literature reviewed by Abanades et al. [1], the main parameters that influence biogas production include process, pretreatment, and feedstock, which is why the proper balance of these parameters is needed to achieve the highest biogas yield. In addition, current trends in anaerobic digestion contribute to the circular economy, which can also, in turn, benefit wastewater treatment [2]. Much of the research on biogas production accentuates lab-scale production [3], currently used in biogas production, which is why the assessment of its applicability needs to be considered [4].
Advantages of anaerobic digestion over other treatment processes include low operating and maintenance costs, low excess biological sludge accumulation, low nutrient requirements, and high methane production efficiency. The disadvantage of anaerobic digestion also includes sensitivity to inhibitors. Many nutrients necessary for operation can have an inhibitory effect in excess amounts [5]. Regular table salt (NaCl) is one of the common compounds found in nature, especially when the substrates for anaerobic digestion come from the sea. When seaweed was used as a substrate for AD, the inhibitory effect of NaCl was found to be slightly below a salt concentration of 3% w/w (concentration of salt in seawater), as no biogas was produced under these conditions [6]. Salt-tolerant plants (halophytes) used as energy crops on saline soils also accumulate salt in their vacuoles, increasing the salt concentration in the biomass [7]. During the anaerobic digestion of food waste, table salt used as a food additive may interfere with anaerobic digestion, and therefore, the optimal NaCl concentration in the anaerobic digestion of food waste has been reported to be 2–4 g/L [8]. Wang et al. [9] found that a NaCl concentration of 20 g/L during anaerobic digestion of food waste did not affect acidogenesis, but it did affect methanogenesis. Adamietz et al. [6] found that increasing salinity up to 20 g/L NaCl had no significant effect on biogas production from oily halophilic algal residues.
Anaerobic treatment of saline waste streams is hindered by ions present in the substrates, especially monovalent cations such as Na+, which cause inhibition of methanogenic archaea [10]. All other influencing factors that inhibit the anaerobic digestion process in general and for different types of waste streams are detailed in the review article by Chen et al. [5]. Indeed, high salt concentrations lead to the plasmolysis of microbial cells due to strong osmotic stress on the cell wall, resulting in the desiccation of microbial cells or, to a greater extent, cell death [7]. Anaerobic bacteria have varying degrees of saline tolerance. Staphylococcus aureus can grow in solutions containing up to 65 g/L NaCl, while E. coli is inhibited at much lower levels [11]. The dominance of the order Clostridiales was observed at higher salinity in biogas reactors, and under the same conditions, a shift in the archaeal community from acetoclastic Methanosaeta to hydrogenotrophic Methanocorpusculum was detected [12]. According to Liu and Boone [13], NaCl toxicity increased in the following order: lignocelluloses-degrading bacteria > acetate-utilizing bacteria > propionate-utilizing bacteria > H2/CO2 utilizing organisms.
In studying waste degradation in landfills with leachate recirculation, Ogata et al. [14] found that a salt concentration of 35 mS/cm of electrical conductivity inhibited methane generation under mesophilic conditions, while salt concentrations below 21 mS/cm electrical conductivity did not affect methane production. It has been postulated previously that the total conductivity in AD should be kept below 30 mS/cm to avoid salt stress [5]. To overcome the negative effect of salinity on anaerobic digestion, Turcios et al. [7] proposed co-digestion with other biomass feedstocks to dilute the salt and/or acclimate the microbial biomass to operate at a higher salinity in the AD process by gradually optimizing the microbial community composition and metabolism. During this optimization process, salt-tolerant microorganisms are preferentially selected in the biomass. While short-term exposure to NaCl concentrations below 4 g/L improved the anaerobic digestion of kitchen waste, higher salt concentrations inhibited biogas production, although some microbial communities had the potential to adapt to a hypersaline environment [15].
Among the first studies on anaerobic treatment of highly saline wastewater were mainly experiments with wastewater from seafood processing [16]. De Baere et al. [17] studied the effects of high NaCl concentrations on methanogenic activity in semi-continuous flow-through reactor systems at mesophilic temperatures (30–35 °C). At 95 g/L of NaCl, 50% inhibition was observed after the adjustment period. Van den Heuvel et al. [18] studied the inhibitory effect of Cl ions on anaerobic up-flow digestion for acidified brine from reverse osmosis treatment of sewage at 20 °C. 2.1 g/L chloride ions in the brine did not negatively affect the treatment. Rinzema et al. [19] reported various inhibitory effects of sodium at neutral pH on the methanogenic activity of the granular sludge in the UASB reactor. Sodium concentrations of 5, 10, and 14 g/L of Na+ caused 10, 50 and 100% inhibition, respectively, relative to the maximum specific acetoclastic methanogenic activity of the granulated sludge containing Methanothrix sp. as predominant acetoclastic methanogen. In addition, no adaptation of Methanothrix sp. to high sodium concentrations was detected over a 12-week period. Feijoo et al. [10] studied the effect of sodium on the methanation of short-chain (volatile) fatty acid (SCFA) mixtures for three different sludges. Sodium concentrations that caused 50% inhibition ranged from 3 to 16 g/L in the absence of nutrients or other salts. Higher sodium tolerance was evident for sludges originating from the digesters that treated highly saline effluents after adaptation. In contrast, Gebauer [20] was unable to achieve sludge stabilization in the mesophilic anaerobic treatment of saline fish farm effluents when using 10–10.5 g/L sodium in the continuously stirred tank reactors (CSTR) at 35 °C. Fang et al. [21] investigated the anaerobic digestion of desugared molasses for biogas production in batch mode and in a CSTR under mesophilic (37 °C) and thermophilic (55 °C) conditions. The latter contains two to three times higher ion concentration than normal molasses, which could inhibit the biogas process. Furthermore, a 50% inhibition of manure digestion occurred at sodium and potassium concentrations of 11 and 28 g/L, respectively.
Several papers in the older and recent literature deal with the effect of salinity on mesophilic anaerobic digestion, but very rarely with anaerobic digestion under thermophilic conditions [22,23]. A list of recent work in this area is given in Table 1. The thermophilic temperature range in anaerobic digestion has many advantages compared to the mesophilic range, such as a high level of pathogen elimination, improved hydrolysis of complex organic materials, better destruction of organic solids, and higher potential for biogas production [24]. However, a disadvantage is also that the process is more sensitive to inhibition than anaerobic digestion in the mesophilic range. Therefore, this study investigates the limits of successful thermophilic anaerobic digestion of organic wastes at high salt concentrations to determine the concentration limits for successful operation. Since substrates with high salinity present a challenge to established anaerobic digestion processes, the study was conducted using inoculum from a biogas plant that had not previously acclimated.
2. Materials and Methods
2.1. Substrate and Inoculum
The microbial inoculum and substrate (a combination of food waste, separately collected municipal organic waste, and slaughterhouse waste) were taken from a full-scale operating thermophilic anaerobic digester from KOTO Ltd. (Ljubljana, Slovenia), which treated the substrate in a conventional thermophilic anaerobic CSTR at 55 °C, with chloride concentrations ranging from 1 and 1.4 g/L, which corresponded to 2.3 g/L NaCl (assuming that almost all chloride was derived from NaCl). The average total solids (TS) concentration of the original substrate was 8.8 ± 0.8%, of which 84.6% were volatile solids (VS). The average total chemical oxygen demand (tCOD) was 100.722 ± 9.836 g/kg, and the total nitrogen (TN) concentration was 3.010 ± 0.420 g/kg. The substrate contained an average of 0.83 ± 0.12% protein and 13.8 ± 1.8% total fat. For the experiments, the substrate was prepared in one batch for all experimental series (approximately 400 kg) and stored in a cold room at 2 °C. Before use, it was diluted 1:2 to avoid interference with other possible inhibitors and to facilitate hydraulic handling in small tubing on lab-scale equipment. The average tCOD of the influent was 49.617 ± 4.918 g/kg with a corresponding concentration of 4.47 ± 0.38% of TS and TN of 1.518 ± 0.157 g/kg. The inoculum was collected from the operating full-scale digester and transferred to the experimental reactors immediately before starting. TS concentration of inoculum was 3.6%, of which 61.7% was VS. The salt content was provided by refined standard cooking salt (NaCl).
2.2. Experimental Setup
The experimental part of the study was conducted in two five-liter lab-scale anaerobic digesters operated at a constant temperature of 55 °C with a constant average organic loading rate (OLR) of 3.8 ± 0.3 kg/m3/d of added VS, corresponding to 5.0 ± 0.4 kg/m3/d of added tCOD. The hydraulic retention time (HRT) was 10 days. In the first digester—experiment, where salt addition was gradual, the pure substrate was started as a control for 24 days, a continuation of the full-scale digester operation where substrate and inoculum were collected. After 24 days, salt was added to a chloride concentration of 15 g/L (equivalent to approximately 21 g/L NaCl). Further, into the course of the experiment, the salt concentration (NaCl) was gradually increased stepwise by approximately 2.5 to 3.0 g/L. After reaching the desired chloride concentration (the targeted concentrations were 17.5, 20.0, 23.0, 25.0, and 28.0 g/L), the process was run for three HRTs. The objective was to test the anaerobic process during continuous—chronic exposure to high salt levels. In the second digester—experiment, where the salt shock was applied, the plain substrate was started as a control for 33 days, followed by shock loadings of salinity, with each desired concentration of salinity reached in 3 days. Shocks were introduced on the 34th, 67th, 100th, 132nd, 162nd, 195th, 235th, 263rd, 271st, 278th, and 285th days of the experiment. The first shock was applied with 21 g/L NaCl (15 g/L of chloride), and later the peak concentrations were increased by 1.4 g/L. Each time after the shock, the chloride concentration was slowly decreased to approximately the initial value of 1 g/L (equivalent to 1.4 g NaCl) by simply omitting the salt addition. After 260 days of operation, the shock loads were applied consecutively every 7 days, and the chloride concentration was not allowed to decrease to the initial value in order to obtain an inhibition effect. In this case, the objective was to test the anaerobic process at high salinity shock loads (acute exposure), as when batches of very saline organic waste substrates were introduced. The salinity levels used in this study were selected based on available information from the relevant literature. These salt contents are found in various substrates originating from the food industry, e.g., in the production of instant soups. A practical example of such batches may be concentrated soup substrates in kitchen waste.
2.3. Analytical Methods
TSS and VSS were analyzed according to APHA Standard Methods. The tCOD was measured according to the method of Zupančič and Roš [33]. TN and protein quantity were determined according to the method of ISO 16634-1. Total fat content was determined according to the method of ISO 11085. SCFAs were extracted from supernatants with diethyl ether according to Holdeman [34] and determined using a gas chromatograph with a flame ionization detector (FID) (Shimadzu GC14A, Kyoto, Japan). Quantification was performed by a standard internal method (crotonic acid 1 g/L). The amount of biogas was determined using a wet tip gas meter and the percent contents of nitrogen, methane, and carbon dioxide by a gas chromatograph equipped with a thermal conductivity detector (TCD) (Shimadzu GC14A, Kyoto, Japan). The chromatographic peaks were evaluated using the integrator Chromatopack CR-4A (Shimadzu, Japan) based on absolute calibration. Chlorides were determined according to ISO 9297:1998.
Changes in the microbial community structure were analyzed using the T-RFLP fingerprinting method on samples collected during the sampling period. Total genomic DNA was extracted from 500 mg of selected samples using the Ultra Clean Soil DNA Isolation Kit (MO-BIO Laboratories, Inc., Carlsbad, CA, USA) according to the manufacturer’s instructions. Then, 16S rRNA gene fragments were amplified using universal bacterial primer pairs 27F [35] and 926R [36], and archaeal primers A109F (5′-ACNGCTCAGTAACRCGYR-3′) (modification of Großkopf et al. [37]) and A915R [38]. Both forward primers were labeled with carboxyfluorescein (6-FAM) at the 5′-end. PCR reactions for bacterial 16S rRNA were performed in a MyCycler™ Thermal Cycler (Bio-Rad, Hercules, CA, USA) with the following parameters: initial preheating for 5 min at 95 °C, followed by 25 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 1 min; and a final extension at 72 °C for 10 min. The optimized PCR amplification conditions for archaeal 16S rDNA were as follows: an initial preheating for 5 min at 95 °C; followed by 28 cycles of denaturation at 95 °C for 30 s, annealing at 52 °C for 45 s, and extension at 72 °C for 90 s; and with a final extension at 72 °C for 10 min. Purification and quantification of amplicons were carried out. Screening for suitable restriction endonucleases was performed, and MspI (Thermo Fisher Scientific Inc., Waltham, MA, USA) was chosen for further analysis of bacteria and archaea. Purified PCR products (300 ng) were digested at 37 °C for 16 h, inactivated, and purified again using a High Pure PCR Purification Kit (Roche) according to the manufacturer’s instructions. Subsequently, 5 µL of the purified digestions were mixed with 0.5 µL internal-lane DNA standard GeneScanTM ROX-500TM (Applied Biosystems Inc., Waltham, MA, USA) and 8 µL HiDi formamide (Applied Biosystems Inc.). Fragment analysis was performed using capillary gel electrophoresis on an ABI PRISM 3130xl Genetic Analyser (Applied Biosystems, Waltham, MA, USA), and electropherograms were analyzed using BioNumerics software 5.1 (Applied Maths, Sint-Martens-Latem, Belgium). The Pearson correlation coefficient with the UPGMA (Unweighted Pair-Group Method with Arithmetic Averages) clustering method was used to assess community profile similarity in BioNumerics [39].
3. Results
3.1. Continuous Exposure to Salinity
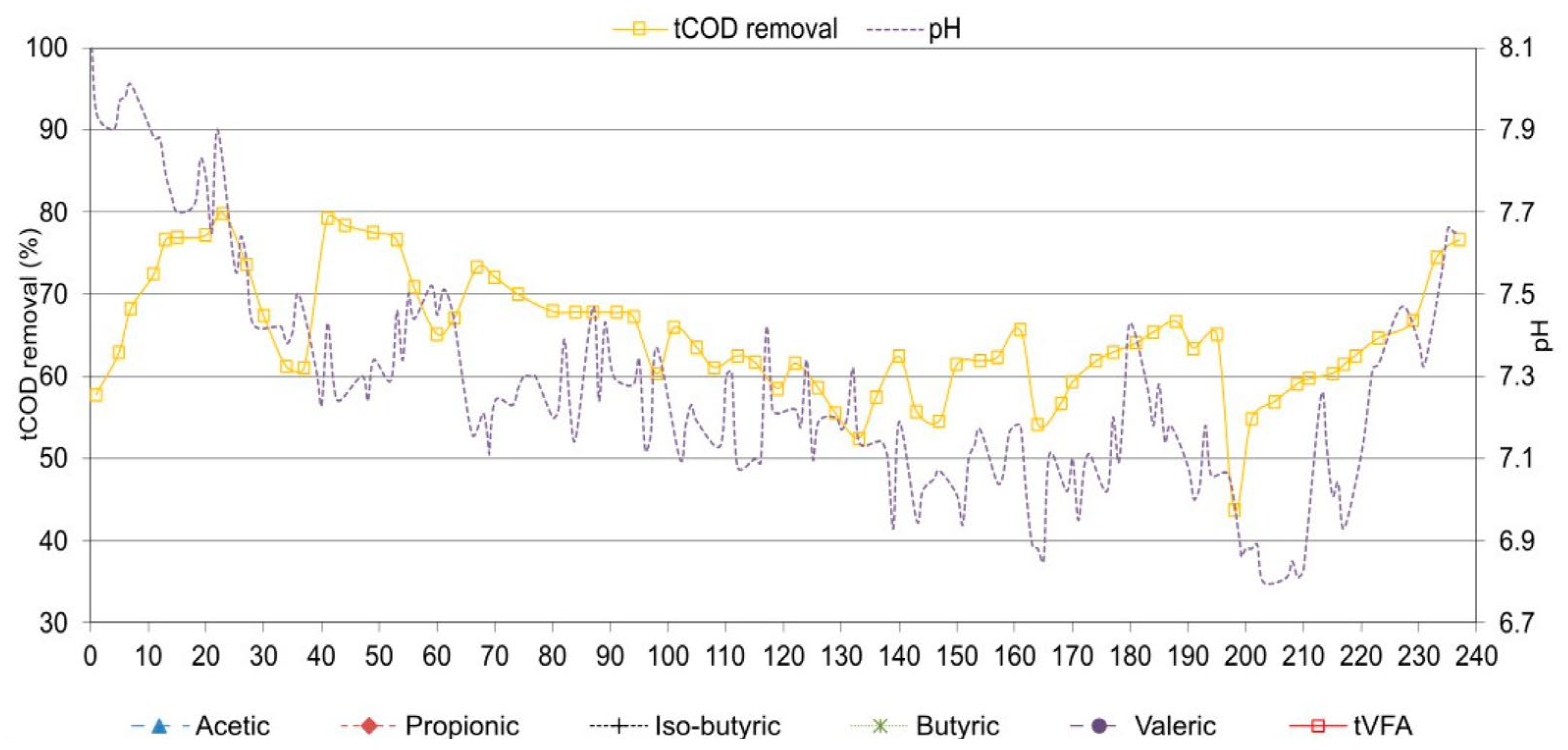
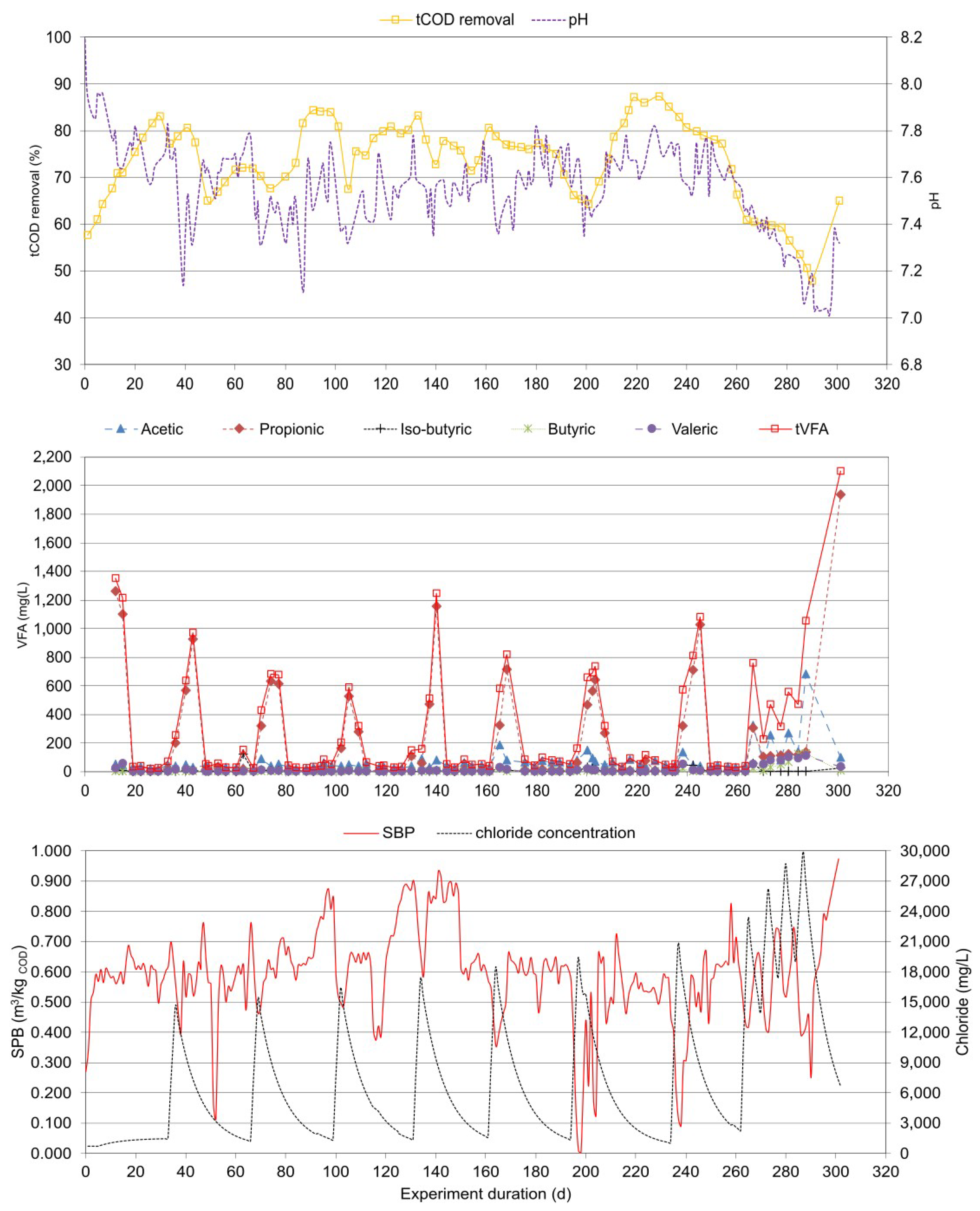
The experiment lasted 238 days (Figure 1). The first 24 days were operated without salt added, and the average SBP (specific biogas production) was 0.562 m3/kgtCOD with 62.0% methane. The maximum tCOD removal efficiency was 79.9%. No significant amounts of SCFAs were detected, and the pH was well above 7.7. After the addition of salt at a concentration of 15 g/L chloride (21 g/L NaCl), the system showed a slight shock, but operation was not significantly hindered. SBP decreased to 0.490 m3/kgtCOD, and tCOD degradation also decreased slightly. tSCFAs increased above 1 g/L, with propionic acid predominating. Concentrations peaked on day 43 with tSCFAs at 1.594 g/L (propionic acid 1.493 g/L and approximately equal traces of other SCFAs). After increasing salinity to chloride concentration of about 17.5 g/L (24.6 g/L NaCl) on day 61 and later to 20.0 g/L (28.2 g/L NaCl) on day 94, the operation did not change significantly. SBP was 0.512 m3/kgtCOD and 0.509 m3/kgtCOD, respectively. The average methane concentration was 58.3 ± 3.0% and did not vary significantly. Peak tSCFAs were 1.078 g/L (propionic acid 0.928 g/L) and 1.485 g/L (propionic acid 1.309 g/L), respectively. The pH ranged from 7.1 to 7.5. Looking at the SCFA data in Figure 1, it can be stated that before the NaCl concentration was increased to 24.6 g/L and 28.2 g/L, the SCFA values decreased significantly to values below 0.500 g/L, indicating that the process flow is stabilizing, and the microbial community is adapting to the conditions. Therefore, up to the NaCl concentration of 28.2 g/L, it can be concluded that no significant inhibition is observed, but the process is affected and requires about three HRTs to adapt to the salt conditions. The NaCl concentration of 28.2 g/L seems to be the boundary between stable operation and the occurrence of inhibition. Already at 28.2 mg/L NaCl (20.0 g/L chloride), the SCFA concentration no longer decreases.
As far as the change in the concentration of organic acids, the following was observed. Acetic acid varied between 18.4 and 2023 mg/L, propionic acid between 2.4 and 1693 mg/L, and tSCFA between 28.8 and 3849 mg/L. It was also noted that there were periods when iso-butyric, butyric and valeric acids were not detected, while the highest concentrations observed were 117, 374 and 121 mg/L, respectively.
After further increasing the chloride concentration, the process showed a partial inhibition, increasing tSCFA to more than 2.000 g/L and peaking at 2.986 g/L and 3.416 g/L at NaCl concentrations of 31.7 (22.5 g/L chloride) and 35.2 g/L (25.0 g/L chloride), respectively. During most of this experimental period (from day 127 to day 190), propionic acid was still predominant (average at 1.462 ± 0.126 g/L), but acetic acid began to increase to equal concentrations. Other SCFAs were less significant in the range between 0.060 and 0.150 g/L. Partial inhibition also decreased the SBP to 0.368 m3/kgtCOD and 0.296 m3/kgtCOD at NaCl concentrations 31.7 and 35.2 g/L, respectively. The tCOD removal also decreased below 60%. The pH was in the range between 6.9 and 7.3, and the methane concentration decreased to 51.1 ± 2.3%.
Further increase of salinity to NaCl concentration of 39.4 g/L (28.0 g/L chloride) initially increased the symptoms of partial inhibition. SBP decreased to 0.256 m3/kgtCOD, tCOD removal continued to decrease (minimal peak at day 199, where removal was 43.6%), and tSCFAs continued to increase. Propionic acid was in the same range as before, and acetic acid increased massively, reaching a peak concentration of 2.023 g/L on day 210. In addition, significant amounts of butyric acid occurred, reaching a peak of 0.374 g/L, confirming the strong inhibition of AD. The other acids were still in the range of 0.060 to 0.150 g/L. On day 210, the process failed, and the SPB dropped below 0.100 m3/kgtCOD (minimum peak 0.075 m3/kgtCOD), which is less than 15% of the initial value. The minimum methane concentration was 47.2%. The pH also decreased below 6.9 (minimum peak 6.8). Recovery of the process was initiated immediately by stopping the addition of substrate. From day 215 to 217, only substrate without salt was added to lower the NaCl concentration to about 32 g/L. After day 226, the process began to recover as biogas production increased. From day 227, the substrate continued to be added daily. While the chloride concentration was decreased, the process fully recovered to an SBP of 0.542 m3/kgtCOD, with a methane concentration of 60.0%, which is practically in the same range as the initial value.
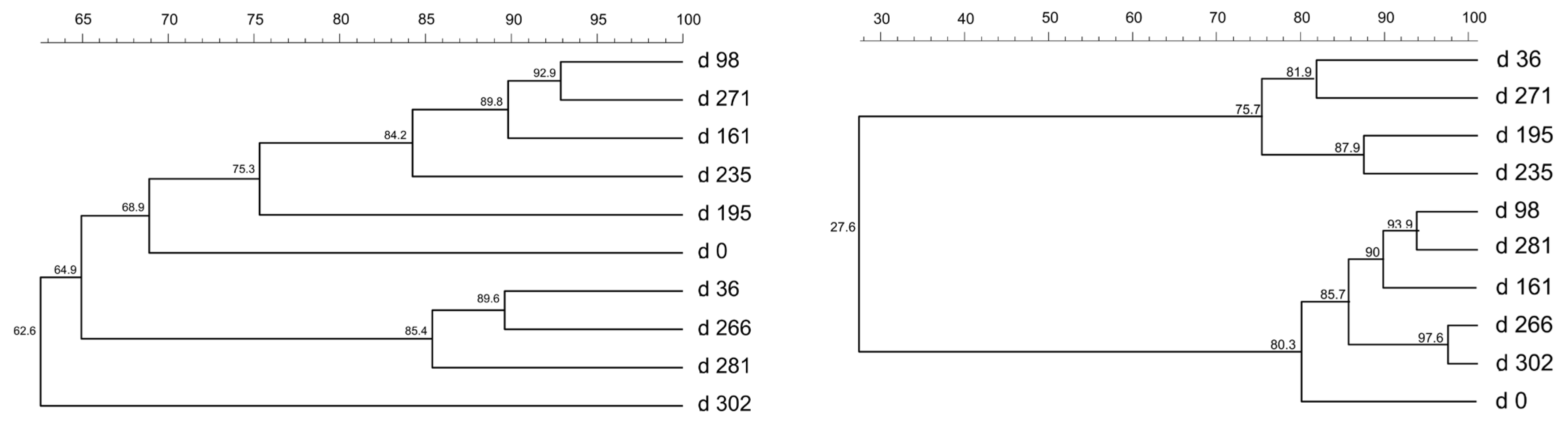
In Figure 2 the T-RFLP analysis at continuous exposure to salinity is shown. Bacteria and archaea in the experiment responded rapidly to salt addition, as indicated by the fact that the samples taken at the chloride concentration of less than 22.5 g/L (31.6 g/L NaCl) were clustered in one group, while samples taken at higher chloride concentrations were clustered separately. Also, at chloride concentration above 22.5 g/L showed higher acetic and propionic acid concentrations and lower methane production.
3.2. Shock Exposure to Salinity
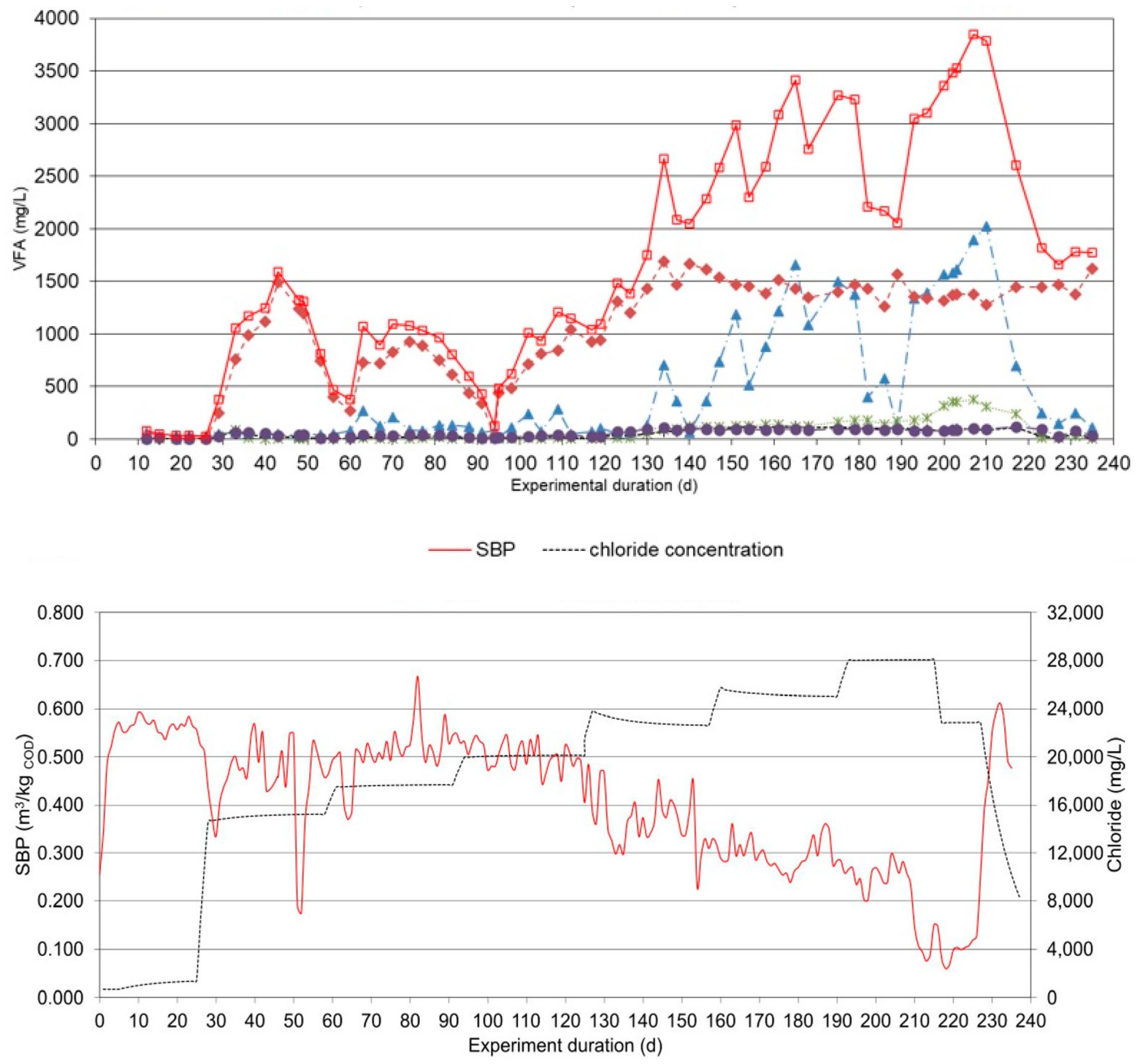
The experiment lasted 303 days (Figure 3). The first 33 days were operated without the addition of salt, and the average SBP was 0.595 m3/kgtCOD with 63.2% methane. The maximum tCOD removal efficiency was 83.1%. No significant amounts of SCFAs were detected, and the pH was well above 7.6.
In terms of the concentration of organic acid, the following was observed. Acetic acid varied between 17.2 and 680 mg/L and tSCFA between 20.1 and 2101 mg/L. It was also noted that there were periods when propionic, iso-butyric, butyric and valeric acids were not detected, while the highest concentrations were 1938, 119, 126, and 112 mg/L, respectively.
The microbial community handled shocks of salinity much better than continuous load. For each shock, an increase in tSCFAs up to 1248 mg/L was observed, with propionic acid (1153 mg/L) being the most dominant. Of the other SCFAs, acetic was the most prevalent, with the others present at less significant concentrations. Each spike showed a decrease in biogas production with a brief inhibition followed by a rapid recovery. The trend in SBP was stable, with an average value of 0.590 ± 0.159 m3/kgtCOD. The average methane concentration was 63.2 ± 2.7%. The degradation of tCOD was stable at 75.2%. The pH varied from 7.1 when the salt shock occurred to 7.8, indicating normal operation. Only after four consecutive shocks did prolonged inhibition occur, which was a consequence of the build-up of salt concentration. The minimum SBP was 0.250 m3/kgtCOD and the pH decreased to 7.0, and the corresponding methane concentration was 50.8%. After the salinity was reduced, the process recovered by itself and reached its initial parameters. This clearly shows that only prolonged and continuous exposure to salinity strongly affects the anaerobic process. It can be concluded that concentrations up to 25.4 g/L NaCl (18 g/L chloride) allow safe operation in the thermophilic range. Thermophilic AD can tolerate shock loading at higher levels of up to 39.4 g/L NaCl (28 g/L chloride) if the salt concentration drops to baseline between shocks. It takes 1 to 1.5 HRT to completely normalize after a shock.
In this experiment, shifts in bacterial communities were clearly influenced by salt addition (Figure 4, left). Obvious shifts were detected one to three days after the salt shock, as indicated by the clustering of samples taken on the first and third days after the salt shock. During the first 260 days, the bacterial community recovered and reached up to 80% similarity to the community prior to the salt shock. The response of the bacterial community to the salt shock was correlated with a rapid partial inhibition of SBP and increased propionic acid concentrations, followed by rapid recovery after approximately 3 days. A similar trend was evident during the shock period from day 260 until the end of the experiment. Although the final bacterial community differed from the initial one by about 34%, it did not lose functionality and provided continuous methane production throughout the experiment (303 days). The diversity of the bacterial community was not highly reduced during the experiment. TRFLP electropherograms showed that two dominant peaks persisted throughout the experiment, indicating the existence of halotolerant species that may have originated from the initial inoculum.
4. Discussion
The results of T-RFLP analysis (Figure 2) showed that bacteria and archaea in the experiment in which salt addition was gradual responded rapidly to salt addition, as indicated by the fact that the samples taken at the chloride concentration of less than 22.5 g/L (31.6 g/L NaCl) were clustered in one group, while samples taken at higher chloride concentrations (above 22.5 g/L and up to 60 g/L NaCl) were clustered separately. Changes in the composition of both bacterial and archaeal microbial communities and lower diversity of both communities as a consequence of high salinity have already been noted by several authors [1,9,12,23]. The increase in NaCl concentration above 31.6 g/L correlates with higher acetic and propionic acid concentrations and lower methane production, too. As reported by Westerholm et al. [40], propionate is an important precursor for methane production, and its accumulation which means hindered conversion to acetate, results in lower SBP. Propionate accumulation is the result of salt stress for propionate-oxidizing bacteria at NaCl concentrations above 31.6 g/L.
As for the changes in the composition of organic acids, it is noted that higher concentrations of acetic acid, butyric acid, valeric acid, and tSCFA were obtained under continuous salt exposure than under salt shock conditions. Higher concentrations of propionic acid and isobutyric acid were obtained under salt shock conditions than under continuous salt exposure. Acclimation of the bacterial and archaeal microbiome to increasing salinity occurs gradually and successfully up to the NaCl concentration of 31.6 g/L so that SBP is almost the same level as at a NaCl concentration of 1.4 g/L. Higher salinity does not reduce effective microbial acclimation (Figure 1). Although the bacterial community at the end of the experiment differs by about 35% and the archaeal community by about 25% from the initial one, and the diversity of both microbial communities is reduced, they still allow the resumption of the methane production process after the salinity decrease at the end of the experiment. The archaeal community is usually considered to be less diverse than that of the bacterial community [9]. The TRFLP electropherograms in the experiment with gradual salt addition showed three common peaks from the beginning to the end of the experiment, one of which became dominant with increasing salinity, while the other two became less distinct, suggesting that one of the archaeal species (presumably halotolerant) was enriched by the high salinity (Figure 2, right). Wilson et al. [41] explained the maintenance of biogas production under saline conditions by acclimation of the microbial community to elevated salt concentrations. De Vrieze et al. [42] concluded from their experiments with hypersalinity in AD, where they analyzed the microbial community composition that the reason for the process recovery after exposure to salinity could be the redundancy of the microbial community. Jabari et al. [23] conducted the only relatively comparable experiment to the presented one. They tested the anaerobic degradation of slaughterhouse wastewater with the enriched (adapted) anaerobic cultures under mesophilic and thermophilic conditions. They found that the bacterial community was strongly affected by the high salinity of 20 and 40 g/L, and bacterial diversity decreased with increasing salinity, which is consistent with the present experiment. The effects of high salt concentration were more evident in thermophilic bacterial cultures than in mesophilic ones.
The archaeal community in the experiment in which salt shocks were applied was more uneven than the bacterial community and less diverse (Figure 4, right). The archaeal community responded rapidly to the salinity shocks. The obtained TRFLP results showed that on the first day after salt shocks, the same peaks in the TRFLP electropherograms disappeared, decreased (two peaks), or increased noticeably (one peak). Salinity shocks likely reduced the salt-sensitive species to levels not detectable by the TRFLP method, resulting in lower archaeal diversity after the shock. The dominant microbial groups are present throughout the shock period (three peaks) and may be the key groups for the resilience of AD. It seems that both bacterial and archaeal microbiomes acclimated well to increased salinity and recovered quite well between the salt shocks. These TRFLP results are in good agreement with Wilson et al. [41] and can be compared to the results of Gao et al. [26], although these experiments with increased salinity were performed under mesophilic conditions and the microbial communities were studied in more detail by metagenomic sequencing.
Allison et al. [12] reported that microbial community redundancy, rather than resilience or resistance, was the main strategy behind anaerobic digestion process recovery. This idea is also supported by De Vrieze et al. [42]. However, some methanogens, such as Methanosarcina spp., form irregular cell clusters that could increase their tolerance to high salt concentrations, and some of them (such as Methanosaeta) can adapt to higher salt concentrations [15]. The salt-adapted archaeal community showed up to 83% similarity with the community before each salt shock. The archaeal community at the end of the experiment with salt shocks differed by 25% from the initial community and, together with the bacterial community, allowed relatively effective methane production until the end of the experiment, except for short periods immediately after the salt shocks. SPB levels partially decreased immediately after salt addition but recovered quickly and remained at virtually the same level until the end of the experiment (Figure 3).
The results of the presented experiments on thermophilic anaerobic digestion under high salinity conditions are difficult to compare with the results of other authors for several reasons. There are only a few studies dealing with the influence of high salinity conditions on thermophilic anaerobic digestion. Most of the experiments were performed under mesophilic conditions. The salinity is represented in different ways in the different experiments: as NaCl concentration, chloride concentration, Na+ concentration or conductivity. The majority of the experiments are short-term (approximately 30 days), while the experiments presented in this study are long-term experiments (238 and 303 days). The volumes of experimental bioreactors range from a few milliliters to several liters, and the experiments are performed at different ORLs, etc.
For anaerobic degradation under high salinity at mesophilic conditions, it was observed that a NaCl concentration of 5 g/L inhibits the CH4 formation rate [9], although the studies on anaerobic treatment of seafood processing wastewater indicate the possibility of anaerobic digestion even at salt concentrations close to the seawater salinity of 30 g/L NaCl [10]. Wang et al. [9] observed that during the mesophilic process, an increase in NaCl concentration from 0 to 20 g/L inhibited methanogenesis but did not affect acidogenesis when the NaCl concentration was below 20 g/L. Alhraishavi and Alani [43] reported that methanogens are affected at NaCl concentrations as low as 6 g/L, whereas acidogenic bacteria are affected only at NaCl concentrations higher than 20 g/L. NaCl concentrations greater than 28.2 g/L were found to inhibit methane production under thermophilic conditions and continuous salt loading in the experiment. The results of the experiment with salt shocks showed that thermophilic AD could even tolerate shock loadings with high levels up to 39.4 g/L NaCl when the salt concentration drops to baseline between shocks and that concentrations up to 25.4 g/L NaCl do not cause inhibition of methane production. These high inhibitory concentrations (compared with mesophilic concentrations) were not expected for the thermophilic process, which is normally considered more susceptible to inhibition than the mesophilic process. The low inhibition could be due to the very robust and flexible microbiome with high redundancy, which originated from a long-standing anaerobic reactor where the microbial biomass could be exposed to several salt shocks before being used for the experiments. Therefore, the microbiome could already be partially acclimated to the high salinity.
The papers of Akinshina et al. [22,44] are some of the few scientific reports on thermophilic AD under saline conditions. They tested the anaerobic degradation of halophyte biomass in a mixture with organic waste for biogas production comparatively at mesophilic and thermophilic temperatures. They obtained biogas yields (equal to SBP) of up to 0.366 m3/kg dry matter at mesophilic conditions and up to 0.480 m3/kg dry matter at thermophilic conditions with organic matter degradation of 60–70% at 55 °C during the 30 days of the incubation period, although thermophilic AD is generally considered to be more sensitive to different inhibition factors [45]. Their results show slightly lower values than in the present work (SBP was 0.562 m3/kgtCOD, and the tCOD removal efficiency was 79.9%), which is probably due to the fact that more degradable organic waste is mixed with less degradable halophyte biomass.
According to Soto et al. [46], a comparison of thermophilic and mesophilic anaerobic treatment of waste from a mussel cooking factory exhibited better performance of the mesophilic reactor due to the faster adaptation of the mesophilic sludge to the high salinity. In addition, sodium is more toxic to propionic acid-utilizing microorganisms than to acetic-acid-utilizing ones. According to Kugelman and McCarty [47], sodium proved to be the most potent inhibitor on a molar basis among the metal cations. Sodium showed moderate inhibition at 3.5–5.5 g/L and strong inhibition at 8 g/L. The authors reviewed by Singh [48] showed different limits of salt inhibition and acclimation, with IC50s in the range of 5.6–53 g/L depending on the adaptation period, antagonistic/synergistic effects, substrate, and reactor configuration. Nevertheless, it can be concluded, from this research, that the process adapts very well up to 20 g/L chloride. Even at higher concentrations, only partial inhibition occurs, while immediate recovery occurs when the salinity is reduced. Alhraishawi and Aslan [29] conducted a review study in which they concluded that the right amount of salt could boost microbial activity, which in turn can increase biogas production, and that sodium salt concentration should be below 8 g/L to prevent inhibition of methane production. Furthermore, Gagliano et al. [30] studied the effects of a complex protein substrate and individual amino acids on anaerobic digestion in a saline environment and found that the addition of individual amino acids in the saline environment disrupted the granule form. The methane content produced during the anaerobic digestion of acidic wastewater can be optimized by dilution with municipal wastewater [31]. Finally, Da Borso et al. [32] studied the anaerobic digestion of brackish water fish sludge (salinity 12 g/L) and observed the highest biomethane potential of 564.2 NmL CH4/g VS.
In summary, based on the conducted studies and in comparison, with the results found in the relevant literature, it can be concluded that a saline environment imposes some limitations on anaerobic digestion. However, the presence of salt is not lethal to the microorganisms, suggesting that careful optimization and future research are needed for better understanding.
5. Conclusions and Future Perspectives
The results of the experiment, in which salt addition was gradual, show that bacteria and archaea responded rapidly to salt addition. Although the structure of bacterial and archaeal microbiomes differed and the diversity decreased at the end of the experiment, the resumption of methane production was still possible after the decrease of salt concentration at the end of the experiment. The results of the experiment in which there was rapid salt addition show that the shift of the bacterial community was significantly affected by the salt addition. However, it was also observed that the bacterial community recovered and reached a great similarity with the original community before the salt shocks. As for the archaeal community, the results show non-uniformity and lower diversity compared to the bacterial community. The archaeal microbiome responded rapidly to the salt shocks by reducing the salt-sensitive species. The maximum COD removal achieved in the gradual salt addition and rapid salt addition experiments was 79.8% and 83.1%, respectively. In the first case, the maximum NaCl concentration was 28.2 g/L, whereas in the second case, higher concentrations of up to 39.4 g/L were tolerated. SPB production with gradual salt addition varied between 0.490 and 0.562 m3/kgtCOD depending on the salt concentration, while SPB production with rapid salt addition was relatively stable at 0.590 m3/kgtCOD. This was also underlined by the fact that short-term rapid salt shocks were not lethal to the microbial community, which quickly recovered and adapted to the above conditions. This suggests that both the bacterial and archaeal communities adapted well to the increased salinity and recovered quite well between salt shocks. These observations suggest that the selected bacterial and archaeal communities are robust and well adapted to high salt concentrations under the conditions tested, which in turn implies that these microbial communities have the potential for anaerobic digestion in hypersaline environments.
We can argue that our thermophilic methanogenic microbiota, resistant to high salinity, developed during lab-scale experiments one and two have great potential. Following the pilot experiments with the developed salt-resistant biomass, we plan to use it as a source of microorganisms for bioaugmentation of anaerobic digestion processes on an industrial scale where high salinity substrates are degraded under thermophilic conditions like waste from the fish industry and food waste. Successful bioaugmentation experiments under mesophilic conditions were conducted by Van Duc et al. [28], using microbial consortia derived from marine sediments to reduce inhibition of methane production under NaCl stress. There is almost no way to find natural microbial consortia for bioaugmentation of thermophilic digestion processes, but there is a way to develop them, as shown in the present work and proposed in one of the recent articles on high salinity wastewater treatment technology [48]. When the results of future pilot scale bioaugmentation experiments are collected, the sustainability assessment of the developed process for anaerobic degradation of high saline organic substrates needs to be performed using tools suggested by Aghbashlo et al. [49].
In future experiments, the metagenomic sequencing of the developed microbial consortium is planned, as performed by Gao et al. [26], to identify the predominant thermophilic salt-resistant bacterial and methanogenic species in order to explain and understand the phenomenon of microbial acclimation to high salinity stress.
The results suggest that anaerobic digestion has great potential for high-salinity wastes. Given the relatively sparse literature on this topic, further investigation of the effect of salinity on anaerobic digestion will provide insights that will optimize waste management for these specific properties. There are certain limitations with respect to immediate application in the industry. Namely, the experiments conducted in this study dealt with uniform, chemically defined salt environments. Therefore, in the case of the application of anaerobic digestion of complex waste streams (shellfish, fish, etc.), additional research is required to gain a more detailed understanding of the eventual application of anaerobic digestion in such an industry characterized by high salinity.
Like the results presented by Fazzino et al. [2], our results confirm the possibility of the production of renewable energy from problematic substrates containing inhibitory substances. We can conclude that anaerobic digestion is a promising technique to introduce a circular economy to the existing platforms. It is very important to upgrade the sustainability features of existing waste management systems in the future as it also contributes to the mitigation of climate change.
Author Contributions
Conceptualization and methodology, G.D.Z. and R.M.L.; methodology, G.DZ., L.L., M.Z. and L.F.; validation, G.D.Z. and L.L.; formal analysis, M.P., M.Z. and L.F.; investigation, G.D.Z., M.P. and L.F.; resources, L.L.; data curation, G.D.Z. and L.F.; writing—original draft preparation, G.D.Z., R.M.L. and L.F.; writing—review and editing, G.D.Z., R.M.L. and L.F.; supervision, L.L.; project administration, L.L.; funding acquisition, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by Slovenian Technology Agency, Project SRRP TIA, Grant No. P-SRRP-09/II/90, Priority scope: Competitiveness of companies and research excellence—improving the competitive abilities of companies and research excellence, Project title: Development of technology to produce biogas from municipal and industrial waste.
Data Availability Statement
The datasets generated from this study are available on request to the corresponding author.
Acknowledgments
The authors would like to thank the Koto maintenance team for support on research equipment service and the Koto lab team for providing analytical support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AD | Anaerobic Digestion |
| CSTR | Completely STirred Reactor |
| HRT | Hydraulic Retention Time |
| OLR | Organic Loading Rate |
| SCFA’s | Short Chain Fatty Acids |
| SBP | Specific Biogas Production |
| TRFLP | Terminal restriction fragment length polymorphism |
| tCOD | Total Chemical Oxygen Demand |
| TN | Total Nitrogen |
| TS | Total Solids |
| UPGMA | Unweighted Pair-Group Method with Arithmetic Averages |
| VS | Volatile Solids |
References
- Abanades, S.; Abbaspour, H.; Ahmadi, A.; Das, B.; Aliehyaei, M.; Esmaeilion, F.; El Haj Assad, M.; Hajilounezhad, T.; Jamali, H.; Hmida, A.; et al. A critical review of biogas production and usage with legislations framework across the globe. Int. J. Environm. Sci. Technol. 2021, 19, 3377–3400. [Google Scholar] [CrossRef]
- Fazzino, F.; Pedullà, A.; Calabrò, P. Boosting the circularity of waste management: Pretreated mature landfill leachate enhances the anaerobic digestion of market waste. Biofuel Res. J. 2023, 10, 1764–1773. [Google Scholar] [CrossRef]
- Tabatabaei, M.; Aghbashlo, M.; Valijanian, E.; Kazemi Shariat Panahi, H.; Nizami, A.-S.; Ghanavati, H.; Sulaiman, A.; Mirmohamadsadeghi, S.; Karimi, K. A comprehensive review on recent biological innovations to improve biogas production, Part 1: Upstream strategies. Renew. Energy 2020, 146, 1204–1220. [Google Scholar] [CrossRef]
- Tabatabaei, M.; Aghbashlo, M.; Valijanian, E.; Kazemi Shariat Panahi, H.; Nizami, A.-S.; Ghanavati, H.; Sulaiman, A.; Mirmohamadsadeghi, S.; Karimi, K. A comprehensive review on recent biological innovations to improve biogas production, Part 2: Mainstream and downstream strategies. Renew. Energy 2020, 146, 1392–1407. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- Adamietz, T.; Jurkowski, W.; Adolph, J.; Brück, T.B. Biogas yields and composition from oil-extracted halophilic algae residues in conventional biogas plants operated at high salinities. Bioproc. Biosyst. Eng. 2019, 42, 1915–1922. [Google Scholar] [CrossRef]
- Turcios, A.E.; Cayenne, A.; Uellendahl, H.; Papenbrock, J. Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Appl. Sci. 2021, 11, 2746. [Google Scholar] [CrossRef]
- Li, X.; Huang, J.; Liu, Y.; Huang, T.; Maurer, C.; Kranert, M. Effects of Salt on Anaerobic Digestion of Food Waste with Different Component Characteristics and Fermentation Concentrations. Energies 2019, 12, 3571. [Google Scholar] [CrossRef]
- Wang, S.; Hou, X.; Su, H. Exploration of the relationship between biogas production and microbial community under high salinity conditions. Sci. Rep. 2017, 7, 1149. [Google Scholar] [CrossRef]
- Feijoo, G.; Soto, M.; Méndez, R.; Lema, J.M. Sodium inhibition in the anaerobic digestion process: Antagonism and adaptation phenomena. Enzyme Microb. Technol. 1995, 17, 180–188. [Google Scholar] [CrossRef]
- Brock, T.D. Biology of Microorganisms; Prentice-Hall: Englewood Cliffs, NJ, USA, 1970. [Google Scholar]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef]
- Liu, Y.; Boone, D.R. Effects of salinity on Methanogenic Decomposition. Bioresour. Technol. 1990, 35, 271–273. [Google Scholar] [CrossRef]
- Ogata, Y.; Ishigaki, T.; Nakagawa, M.; Yamada. Effect of increasing salinity on biogas production in waste landfills with leachate recirculation: A lab-scale model study. Biotechnol. Rep. 2016, 10, 111–116. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, R.; He, Q.; Ji, B.; Wang, H.; Yang, K. Adaptation to salinity: Response of biogas production and microbial communities in anaerobic digestion of kitchen waste to salinity stress. J. Biosci. Bioeng. 2020, 130, 173–178. [Google Scholar] [CrossRef]
- Omil, F.; Mendez, R.; Lema, J.M. Anaerobic treatment of saline wastewaters under high sulphide and ammonia content. Bioresour. Technol. 1995, 54, 269–278. [Google Scholar] [CrossRef]
- De Baere, L.A.; Devocht, M.; Van Assche, P.; Verstraete, W. Influence of high NaCl and NH4Cl salt levels on methanogenic associations. Water Res. 1984, 18, 543–548. [Google Scholar] [CrossRef]
- Van Den Heuvel, J.C.; Zoetemeyer, R.J.; Boelhouwer, C. Purification of municipal wastewater by subsequent reverse osmosis and anaerobic digestion. Biotechnol. Bioeng. 1981, 23, 2001–2008. [Google Scholar] [CrossRef]
- Rinzema, A.; van Lier, J.; Lettinga, G. Sodium inhibition of acetoclastic methanogens in granular sludge from a UASB reactor. Enzyme Microbiol. Technol. 1988, 10, 24–32. [Google Scholar] [CrossRef]
- Gebauer, R. Mesophilic anaerobic treatment of sludge from saline fish farm effluents with biogas production. Bioresour. Technol. 2004, 93, 155–167. [Google Scholar] [CrossRef]
- Fang, C.; Boe, K.; Angelidaki, I. Anaerobic co-digestion of desugared molasses with cow manure; focusing on sodium and potassium inhibition. Bioresour. Technol. 2011, 102, 1005–1011. [Google Scholar] [CrossRef]
- Akinshina, N.; Naka, D.; Toderich, K.; Azizov, A.; Yasui, H. Anaerobic degradation of halophyte biomass for biogas production. J. Arid. Land 2012, 22, 227–230. [Google Scholar]
- Jabari, L.; Khelifi, E.; Gannoun, H.; Fardeu, M.-L.; Godon, J.-J.; Hamdi, M. Effect of salinity and temperature on the bacterial diversity shift of anaerobic batch cultures treating abattoir wastewater. Desalination Water Treat. 2016, 57, 13909–13915. [Google Scholar] [CrossRef]
- Siles, J.A.; Brekelmans, J.; Martín, M.A.; Chica, A.F.; Martín, A. Impact of ammonia and sulphate concentration on thermophilic anaerobic digestion. Bioresour. Technol. 2010, 101, 9040–9048. [Google Scholar] [CrossRef]
- Lee, J.; Kim, E.; Hwang, S. Effects of inhibition by sodium ion and ammonia and different inocula on acetate-utilizing methanogenesis: Methanogenic activity and succession of methanogens. Bioresour. Technol. 2021, 334, 125202. [Google Scholar] [CrossRef]
- Gao, M.; Yang, J.; Liu, Y.; Zhang, J.; Li, Y.; Liu, Y.; Wu, B.; Gu, L. Deep insights into the anaerobic co-digestion of waste activated sludge with concentrated leachate under different salinity stresses. Sci. Total Environ. 2022, 838, 155922. [Google Scholar] [CrossRef]
- Guo, L.; Xie, L.; Sun, W.; Xu, Y.; Sun, Y. Research progress of high-salinity wastewater treatment technology. Water 2023, 15, 684. [Google Scholar] [CrossRef]
- Van Duc, L.; Nagao, S.; Mojarrad, M.; Miyagawa, Y.; Li, Z.-Y.; Inoue, D.; Tajima, T.; Ike, M. Bioaugmentation with marine-derived microbial consortia in mesophilic anaerobic digestion for enhancing methane production under ammonium or salinity stress. Bioresour. Technol. 2023, 376, 128853. [Google Scholar] [CrossRef]
- Alhraishawi, A.; Aslan, S. Effect of Salt Content on Biogas Production and Microbial Activity: Review Study. Eur. Sci. J. 2023, 19, 17. [Google Scholar] [CrossRef]
- Gagliano, M.C.; Sudmalis, D.; Pei, R.; Temmink, H.; Plugge, C.M. Microbial Community Drivers in Anaerobic Granulation at High Salinity. Front. Microbiol. 2020, 11, 235. [Google Scholar] [CrossRef]
- Mazioti, A.A.; Vyrides, I. Anaerobic digestion of high strength bilgewater with granular sludge: Confronting salinity and investigating biomass adaptation. In Proceedings of the 8th International Conference on Sustainable Solid Waste Management, Thessaloniki, Greece, 23–26 June 2021. [Google Scholar]
- Da Borso, F.; Chiumenti, A.; Fait, G.; Mainardis, M.; Goi, D. Biomethane Potential of Sludges from a Brackish Water Fish Hatchery. Appl. Sci. 2021, 11, 552. [Google Scholar] [CrossRef]
- Zupančič, G.D.; Roš, M. Determination of chemical oxygen demand in substrates from anaerobic treatment of solid organic waste. Waste Biomass Valor. 2012, 1, 89–98. [Google Scholar] [CrossRef]
- Holdeman, L.V.; Cato, E.P.; Moore, W.E.C. Anaerobe Laboratory Manual, 4th ed.; VPI: Blacksburg, VA, USA, 1977; pp. 1–156. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acids Techniques in Bacterial Systematics; Stackebrant, E., Goodfellov, M., Eds.; Wiley: Chichester, UK, 1991; pp. 115–148. [Google Scholar]
- Watanabe, K.; Kodama, Y.; Harayama, S. Design and evaluation of PCR primers to amplify bacterial 16S ribosomal DNA fragments used for community fingerprinting. J. Microbiol. Methods 2001, 44, 253–262. [Google Scholar] [CrossRef]
- Grosskopf, P.H.; Janssen, W.; Liesack. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl. Environ. Microbiol. 1998, 64, 960–969. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef] [PubMed]
- Novak, D.; Franke-Whittle, I.H.; Tratar-Pirc, E.; Jerman, V.; Insam, H.; Marinšek-Logar, R.; Stres, B. 2013. Biotic and abiotic processes contribute to successful anaerobic degradation of cyanide by UASB reactor biomass treating brewery treating brewery wastewater. Water Res. 2013, 47, 3644–3653. [Google Scholar] [CrossRef]
- Westerholm, M.; Calusinska, M.; Dolfing, J. Syntrophic propionate-oxidizing bacteria in methanogenic systems. FEMS Microbiol. Rev. 2022, 46, fuab057. [Google Scholar] [CrossRef]
- Wilson, L.P.; Loetscher, L.H.; Sharvelle, S.E.; De Long, S.K. Microbial community acclimation enhances waste hydrolysis rates under elevated ammonia and salinity conditions. Bioresour. Technol. 2013, 146, 15–22. [Google Scholar] [CrossRef]
- De Vrieze, J.; Christiaens, M.E.R.; Walraedt, D.; Devoogt, A. Microbial community redundance in anaerobic digestion drives process recovery after salinity exposure. Water Res. 2017, 111, 109–117. [Google Scholar] [CrossRef]
- Alhraishawi, A.A.; Alani, W.K. The cofermentation of organic substrates: A review. Performance of biogas production under different salt content. IOP Conf. Ser. J. Phys. Conf. Ser. 2018, 1032, 012041. [Google Scholar] [CrossRef]
- Akinshina, N.; Azizov, A.; Karasyova, T.; Klose, E. On the issue of halophytes as energy plants in saline environment. Biomass Bioenerg. 2016, 91, 306–311. [Google Scholar] [CrossRef]
- Gebreeyesus, G.D.; Janicek, P. Thermophilic versus mesophilic anaerobic digestion of sewage sludge: A comparative review. Bioeng. 2016, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.; Mendez, R.; Lema, J.M. Biodegradability and toxicity in the anaerobic treatment of fish canning wastewaters. Environ. Technol. 1991, 12, 669–677. [Google Scholar] [CrossRef]
- Kugelman, I.J.; McCarty, P.L. Cation Toxicity and Stimulation in Anaerobic Waste Treatment. J. Water Pollut. Control Fed. 1965, 37, 97–116. [Google Scholar]
- Singh, M. Metal Ion Inhibitors and Other Inorganic Toxicants in Anaerobic Biological Treatment Process of Industrial Effluents: A Critical Review. Int. J. Sci. Res. 2017, 6, 2353–2363. [Google Scholar]
- Aghbashlo, M.; Hosseinzadeh-Batha, H.; Shahbeik, H.; Tabatabaei, M. The role of sustainability assessment tools in realizing bioenergy and byproduct systems. Biofuel Res. J. 2022, 3, 1697–1706. [Google Scholar] [CrossRef]
Figure 1.
tCOD removal, pH, SBP, chloride concentration and SCFAs at continuous exposure to salinity.
Figure 1.
tCOD removal, pH, SBP, chloride concentration and SCFAs at continuous exposure to salinity.


Figure 2.
UPGMA dendrogram of T-RFLP profiles of bacterial (left) and archaeal (right) communities at continuous exposure to salinity. The numbers above the dendrogram represent the percent of similarity.
Figure 2.
UPGMA dendrogram of T-RFLP profiles of bacterial (left) and archaeal (right) communities at continuous exposure to salinity. The numbers above the dendrogram represent the percent of similarity.

Figure 3.
tCOD removal, pH, SBP, chloride concentration and SCFAs at Shock exposure to salinity.

Figure 4.
UPGMA dendrogram of T-RFLP profiles of bacterial (left) and archaeal (right) communities at shock exposure to salinity. The numbers above the dendrogram represent the percent of similarity.
Figure 4.
UPGMA dendrogram of T-RFLP profiles of bacterial (left) and archaeal (right) communities at shock exposure to salinity. The numbers above the dendrogram represent the percent of similarity.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of latest work in the relevant field.
| Authors | Literature |
|---|---|
| Lee et al. [25] | Effects of inhibition by sodium ion and ammonia and different inocula on acetate-utilizing methanogenesis: Methanogenic activity and succession of methanogens. |
| Gao et al. [26] | Deep insights into the anaerobic co-digestion of waste activated sludge with concentrated leachate under different salinity stresses |
| Guo et al. [27] | Research progress of high-salinity wastewater treatment technology |
| Van Duc et al. [28] | Bioaugmentation with marine-derived microbial consortia in mesophilic anaerobic digestion for enhancing methane production under ammonium or salinity stress |
| Alhraishawi and Aslan [29] | Effect of salt content on biogas production and microbial activity |
| Gagliano et al. [30] | Microbial community drivers in anaerobic granulation at high salinity |
| Mazioti and Vyrides [31] | Anaerobic digestion of high strength bilgewater with granular sludge: confronting salinity and investigating biomass adaptation |
| Da Borso et al. [32] | Biomethane potential of sludges from a brackish water fish hatchery |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zupančič, G.D.; Panjičko, M.; Marinšek Logar, R.; Lavrič, L.; Zorec, M.; Fanedl, L. Salinity Inhibition in Thermophilic Anaerobic Digestion of Organic Waste. Appl. Sci. 2023, 13, 6590. https://doi.org/10.3390/app13116590
AMA Style
Zupančič GD, Panjičko M, Marinšek Logar R, Lavrič L, Zorec M, Fanedl L. Salinity Inhibition in Thermophilic Anaerobic Digestion of Organic Waste. Applied Sciences. 2023; 13(11):6590. https://doi.org/10.3390/app13116590
Chicago/Turabian StyleZupančič, Gregor Drago, Mario Panjičko, Romana Marinšek Logar, Lea Lavrič, Maša Zorec, and Lijana Fanedl. 2023. "Salinity Inhibition in Thermophilic Anaerobic Digestion of Organic Waste" Applied Sciences 13, no. 11: 6590. https://doi.org/10.3390/app13116590
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.