
Expediting the Search for Climate-Resilient Reef Corals in the Coral Triangle with Artificial Intelligence
 ,
,
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
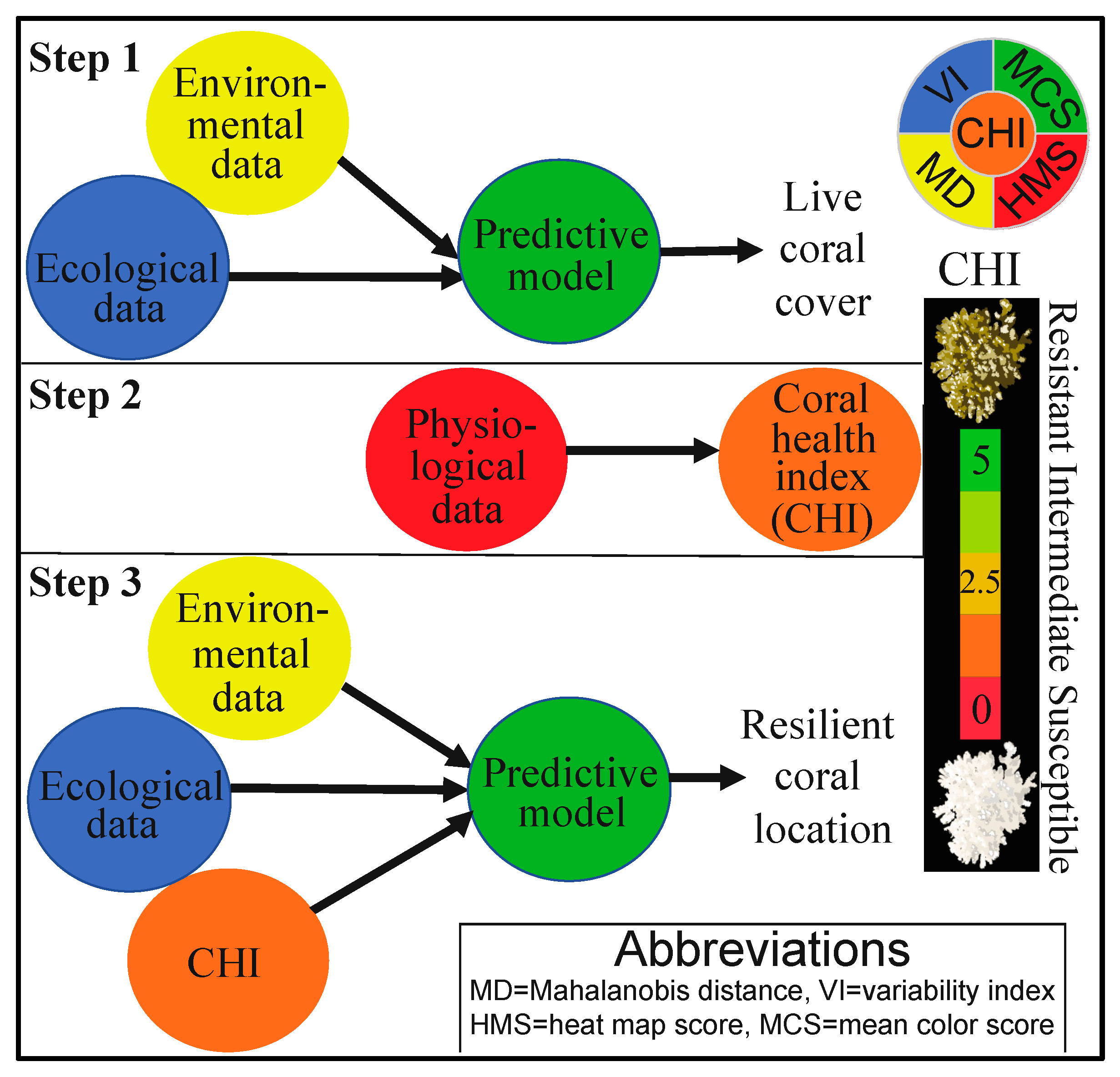
2.1. Overview of Approach
2.2. Field Surveys and Sampling
2.3. Coral Sample Analysis
2.4. Data Analysis I-Multivariate Physiology
2.5. Data Analysis II-CHI
3. Results and Discussion
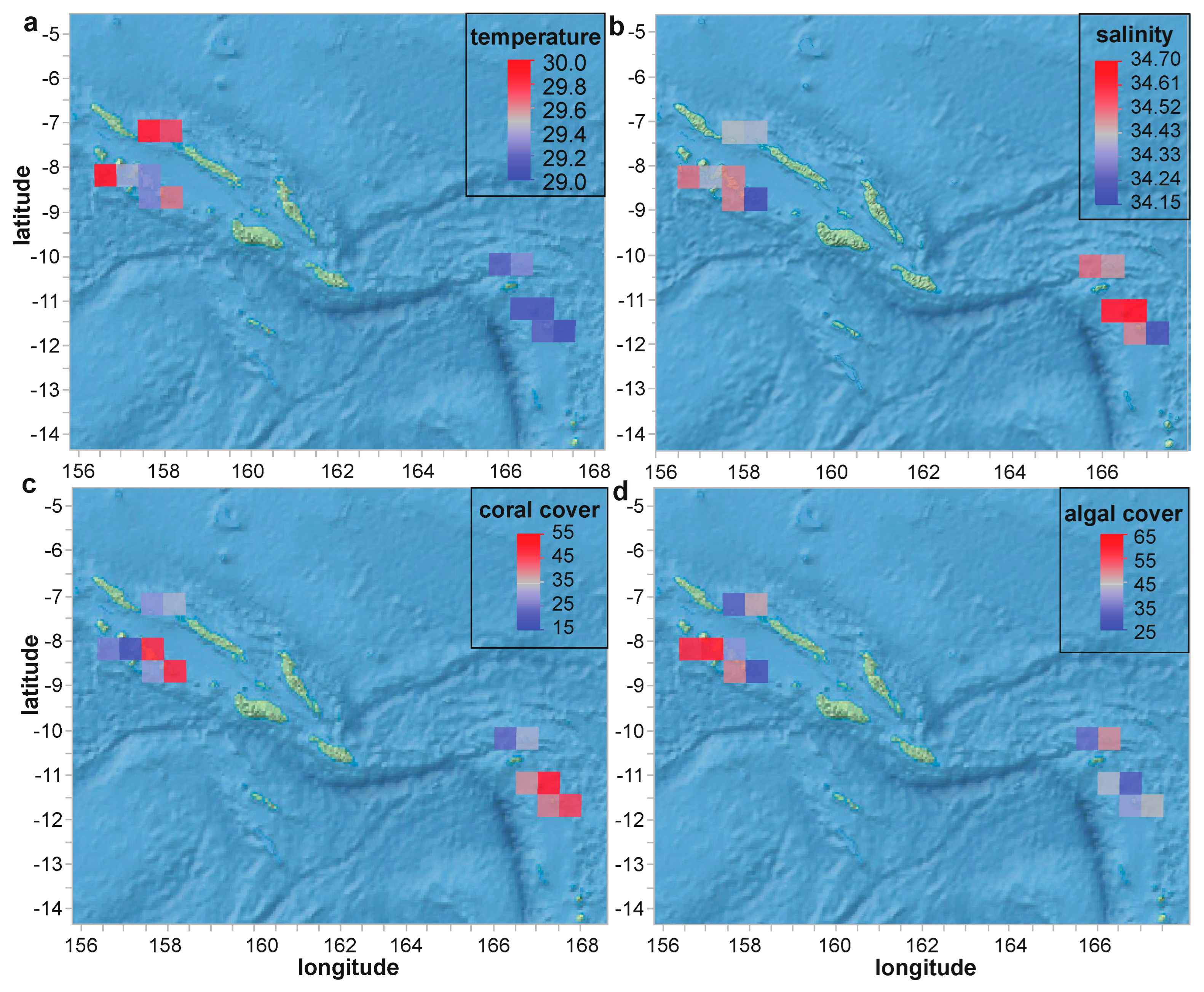
3.1. Overview of the Research Cruise

3.2. Coral Physiological Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Parameter | n | F | p |
|---|---|---|---|
| Island | 10 | 2.20 | <0.01a |
| Site | 33 | 1.56 | <0.01 |
| Latitude | 33 | 0.98 | 0.42 |
| Longitude | 33 | 3.24 | 0.02 |
| Survey date | 19 | 1.69 | 0.16 |
| Survey time | 3 | 0.78 | 0.54 |
| Survey depth | 4 | 1.31 | 0.21 |
| Reef exposure | 3 | 1.19 | 0.31 |
| Reef type | 3 | 1.94 | 0.06 |
| Lagoon | 2 | 2.36 | 0.06 |
| Reef emergence | 2 | 0.46 | 0.76 |
| Reef location | 3 | 1.85 | 0.07 |
| Temperature | 13 | 0.63 | 0.64 |
| Salinity | 10 | 0.99 | 0.42 |

| X’s (#) | Validation Type | Model Type | R2 | Model Details | Most Important Predictor |
|---|---|---|---|---|---|
| ENV(11) 1 | NA | Predictor screen | NA | 100 trees | Site (45%) |
| ENV(14) 1 | NA | Predictor screen | NA | 100 trees a | Site (37%) |
| ENV(14) 1 | Val w test | NN GUI-HL(1) | 0.83 | TanH(1)-Linear(1)-Gaussian(3)-Boost(6) b | Longitude |
| ENV(14) 1 | 20% HB | NN GUI-HL(1) | 0.86 | TanH(3)-Linear(1)-Gaussian(1)-Boost(11) c | Date |
| ECO(50) 1 | NA | Predictor screen | NA | 100 trees | Turf-sediment (19%) |
| ENV(11)+ECO(50) | 10% HB | NN GUI-HL(1) | 0.98 | HL1: TanH(4)-Linear(2)-Gaussian(3)-Boost(8) c,d,e | Reef emergence |
| ENV(14)+ECO(50) | 20% HB | NN GUI-HL(1) | 0.92 | TanH(4)-Linear(1)-Boost(11) f | Site |
3.3. Caveats and Future Work
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, G.; Heron, S.F.; Eakin, C.M.; Muller-Karger, F.E.; Vega-Rodriguez, M.; Guild, L.S.; De La Cour, J.L.; Geiger, E.F.; Skirving, W.J.; Burgess, T.F.R.; et al. Reef-scale thermal stress monitoring of coral ecosystems: New 5-km global products from NOAA Coral Reef Watch. Remote Sens. 2014, 6, 11579–11606. [Google Scholar] [CrossRef] [Green Version]
- UNEP. Coral Bleaching Futures—Downscaled Projections of Bleaching Conditions for the World’s Coral Reefs, Implications of Climate Policy and Management Responses; United Nations Environment Programme: Nairobi, Kenya, 2017. [Google Scholar]
- Woesik, R.V.; Köksal, S.; Ünal, A.; Cacciapaglia, C.W.; Randall, C.J. Predicting coral dynamics through climate change. Sci. Rep. 2018, 8, 17997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sully, S.; Burkepile, D.E.; Donovan, M.K.; Hodgson, G.; van Woesik, R. A global analysis of coral bleaching over the past two decades. Nat. Commun. 2019, 10, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quigley, K.M.; van Oppen, M.J.H. Predictive models for the selection of thermally tolerant corals based on offspring survival. Nat. Commun. 2022, 13, 1543. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Donner, S.D.; Maynard, J.A.; MacNeil, M.A.; Graham, N.A.; Maina, J.; Baker, A.C.; Alemu I, J.B.; Beger, M.; Campbell, S.J.; et al. Prioritizing key resilience indicators to support coral reef management in a changing climate. PLoS ONE 2012, 7, e42884. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Eakin, C.M.; Sweatman, H.P.; Brainard, R.E. The 2014–2017 global-scale coral bleaching event: Insights and impacts. Coral Reefs 2019, 38, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Gintert, B.E.; Manzello, D.P.; Enochs, I.C.; Kolodziej, G.; Carlton, R.; Gleason, A.C.R.; Gracias, N. Marked annual coral bleaching resilience of an inshore patch reef in the Florida Keys: A nugget of hope, aberrance, or last man standing? Coral Reefs 2018, 37, 533–547. [Google Scholar] [CrossRef] [Green Version]
- McClanahan, T.R.; Darling, E.S.; Maina, J.M.; Muthiga, N.A.; D’agata, S.; Leblond, J.; Arthur, R.; Jupiter, S.D.; Wilson, S.K.; Mangubhai, S.; et al. Highly variable taxa-specific coral bleaching responses to thermal stresses. Mar. Ecol. Prog. Ser. 2020, 648, 135–151. [Google Scholar] [CrossRef]
- Keshavmurthy, S.; Meng, P.J.; Wang, J.T.; Kuo, C.Y.; Yang, S.Y.; Hsu, C.M.; Gan, C.H.; Dai, C.F.; Chen, C.A. Can resistant coral-Symbiodinium associations enable coral communities to survive climate change? A study of a site exposed to long-term hot water input. Peer J. 2014, 2, e327. [Google Scholar] [CrossRef]
- Rubin, E.; Enochs, I.C.; Foord, C.; Kolodziej, G.; Basden, I.; Manzello, D.P.; Mayfield, A.B. Molecular mechanisms of coral persistence within highly urbanized locations in the Port of Miami, Florida. Front. Mar. Sci. 2021, 8, 695236. [Google Scholar] [CrossRef]
- Mayfield, A.B. Uncovering spatio-temporal and treatment-derived differences in the molecular physiology of a model coral-dinoflagellate mutualism with multi-variate statistical approaches. J. Mar. Sci. Eng. 2016, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Putnam, H.M.; Mayfield, A.B.; Fan, T.Y.; Chen, C.S.; Gate, R.D. The physiological and molecular responses of larvae from the reef-building coral Pocillopora damicornis exposed to near-future increases in temperature and pCO2. Mar. Biol. 2013, 160, 2157–2173. [Google Scholar] [CrossRef]
- Cruz-García, R.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L.; Mayfield, A.B. Ephemeral effects of El Niño southern oscillation events on an eastern tropical Pacific coral community. Mar. Freshw. Res. 2020, 71, 1259–1268. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Chen, C.S. Enabling coral reef triage via molecular biotechnology and artificial intelligence. Platax 2019, 16, 23–47. [Google Scholar]
- Mayfield, A.B. Exploiting the power of multivariate statistics for probing the cellular biology of thermally challenged reef corals. Platax 2020, 17, 27–52. [Google Scholar]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C.; Bruckner, A.W. The molecular ecophysiology of closely related pocilloporids from the South Pacific: A case study from the Austral and Cook Islands. Platax 2016, 13, 1–25. [Google Scholar]
- Mayfield, A.B.; Dempsey, A.C. Environmentally-driven physiological variation in a New Caledonian reef coral. Oceans 2022, 3, 15–29. [Google Scholar] [CrossRef]
- Bruckner, A.W. Global Reef Expedition: Solomon Islands; Field Report; Khaled bin Sultan Living Oceans Foundation: Landover, MD, USA, 2015; 39p. [Google Scholar]
- Mayfield, A.B.; Dempsey, A.C.; Chen, C.S. Predicting the abundance of corals from simple environmental predictors with a machine-learning approach. Platax 2022, 19, 1–24. [Google Scholar]
- Mayfield, A.B.; Tsai, S.; Lin, C. The Coral Hospital. Biopreserv. Biobank. 2019, 17, 355–369. [Google Scholar] [CrossRef]
- Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Mayfield, A.B.; Cupul-Magaña, A.L. Temporal variation in invertebrate recruitment on an Eastern Pacific coral reef. J. Sea Res. 2019, 145, 8–15. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Chen, C.S. A coral transcriptome in the Anthropocene as an “alternative stable state”. Platax 2020, 17, 1–26. [Google Scholar]
- Mayfield, A.B.; Bruckner, A.W.; Chen, C.H.; Chen, C.S. A survey of pocilloporids and their endosymbiotic dinoflagellate communities in the Austral and Cook Islands of the South Pacific. Platax 2015, 12, 1–17. [Google Scholar]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C. Biomarker profiling in reef corals of Tonga’s Ha’apai and Vava’u Archipelagos. PLoS ONE 2017, 12, e0185857. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C. Identifying corals displaying aberrant behavior in Fiji’s Lau Archipelago. PLoS ONE 2017, 12, e0177267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C. The molecular ecophysiology of closely related pocilloporid corals of New Caledonia. Platax 2017, 14, 1–45. [Google Scholar]
- Mayfield, A.B.; Hirst, M.B.; Gates, R.D. Gene expression normalization in a dual-compartment system: A real-time PCR protocol for symbiotic anthozoans. Mol. Ecol. Res. 2009, 9, 462–470. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Chen, C.S.; Liu, P.J. Decreased green fluorescent protein-like chromoprotein gene expression in specimens of the reef-building coral Pocillopora damicornis undergoing high temperature-induced bleaching. Platax 2014, 11, 1–23. [Google Scholar]
- Mayfield, A.B.; Wang, Y.B.; Chen, C.S.; Chen, S.H.; Lin, C.Y. Compartment-specific transcriptomics in a reef-building coral exposed to elevated temperatures. Mol. Ecol. 2014, 23, 5816–5830. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.B.; Chen, C.S.; Dempsey, A.C. Modeling environmentally-mediated variation in reef coral physiology. J. Sea Res. 2019, 145, 44–54. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Dempsey, A.C.; Inamdar, J.; Chen, C.S. A statistical platform for assessing coral health in an era of changing global climate-I: A case study from Fiji’s Lau Archipelago. Platax 2018, 15, 1–35. [Google Scholar]
- Mayfield, A.B.; Dempsey, A.C.; Chen, C.S. Assessing coral health in the Kingdom of Tonga with a coral health index. Platax 2021, 18, 53–78. [Google Scholar]
- Mayfield, A.B. Machine-learning-based proteomic predictive modeling with thermally-challenged Caribbean reef corals. Diversity 2022, 14, 33. [Google Scholar] [CrossRef]
- Chen, C.C.; Hsieh, H.Y.; Mayfield, A.B.; Chang, C.M.; Wang, J.T.; Meng, P.J. The key impact of water quality on the coral reefs of Kenting National Park. J. Mar. Sci. Eng. 2022, 10, 270. [Google Scholar] [CrossRef]
- Lin, H.J.; Lee, C.L.; Peng, S.E.; Hung, M.C.; Liu, P.J.; Mayfield, A.B. Anthropogenic nutrient enrichment may exacerbate the impacts of El Niño-Southern Oscillation (ENSO) events on intertidal seagrass beds. Glob. Chang. Biol. 2018, 24, 4566–4580. [Google Scholar] [CrossRef]
- Huang, Y.L.; Mayfield, A.B.; Fan, T.Y. Effects of feeding on the physiological performance of the stony coral Pocillopora acuta. Sci. Rep. 2020, 10, 19988. [Google Scholar] [CrossRef]
- Lin, C.; Tsai, S.; Mayfield, A.B. Physiological differences between cultured and wild coral eggs. Biopreserv. Biobank. 2019, 17, 370–371. [Google Scholar] [CrossRef]
- Horwitz, R.; Hoogenboom, M.; Fine, M. Spatial competition dynamics between reef corals under ocean acidification. Sci. Rep. 2017, 7, 40288. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Fan, T.Y.; Chen, C.S. Physiological acclimation to elevated temperature in a reef-building coral from an upwelling environment. Coral Reefs 2013, 32, 909–921. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Fan, T.Y.; Chen, C.S. Real-time PCR-based gene expression analysis in the model reef-building coral Pocillopora damicornis: Insight from a salinity stress study. Platax 2013, 10, 1–29. [Google Scholar]
- Mayfield, A.B.; Chen, Y.J.; Lu, C.Y.; Chen, C.S. The proteomic response of the reef coral Pocillopora acuta to experimentally elevated temperature. PLoS ONE 2018, 13, e0192001. [Google Scholar] [CrossRef] [PubMed]
- McRae, C.; Mayfield, A.B.; Fan, T.Y.; Huang, W.B.; Cote, I. Differing proteomic responses to high-temperature exposure between adult and larval reef corals. Front. Mar. Sci. 2021, 8, 716124. [Google Scholar] [CrossRef]
- Enochs, I.C.; Formel, N.; Manzello, D.P.; Morris, J.; Mayfield, A.B.; Boyd, A.; Kolodziej, G.; Adams, G. Coral persistence despite extreme periodic pH fluctuations at a volcanically acidified Caribbean reef. Coral Reefs 2020, 39, 523–528. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Wang, L.H.; Tang, P.C.; Hsiao, Y.Y.; Fan, T.Y.; Tsai, C.L.; Chen, C.S. Assessing the impacts of experimentally elevated temperature on the biological composition and molecular chaperone gene expression of a reef coral. PLoS ONE 2011, 6, e26529. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, A.B. Proteomic signature of corals from thermodynamic reefs. Microorganisms 2020, 8, 1171. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Aguilar, C.; Enochs, I.C.; Kolodziej, G.; Manzello, D.P. Shotgun proteomics of thermally challenged Caribbean reef corals. Front. Mar. Sci. 2021, 8, 660153. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Chan, P.H.; Putnam, H.M.; Chen, C.S.; Fan, T.Y. The effects of a variable temperature regime on the physiology of the reef-building coral Seriatopora hystrix: Results from a laboratory-based reciprocal transplant. J. Exp. Biol. 2012, 215, 4183–4195. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.Y.; Hwang, G.W.; Lin, H.J.; Mayfield, A.B.; Chen, C.P. The development of a habitat suitability model for sub-tropical tidal flat fiddler crabs. Ocean Coast. Manag. 2019, 182, 104931. [Google Scholar] [CrossRef]
- Chang, T.C.; Mayfield, A.B.; Fan, T.Y. Culture systems influence the physiological performance of the soft coral Sarcophyton glaucum. Sci. Rep. 2020, 10, 20200. [Google Scholar] [CrossRef]




| Model/Analysis Type | Model Y(’s) | Model X’s | Validation Type | Data Location |
|---|---|---|---|---|
| CORAL PHYSIOLOGICAL DATASET EXPLORATION | ||||
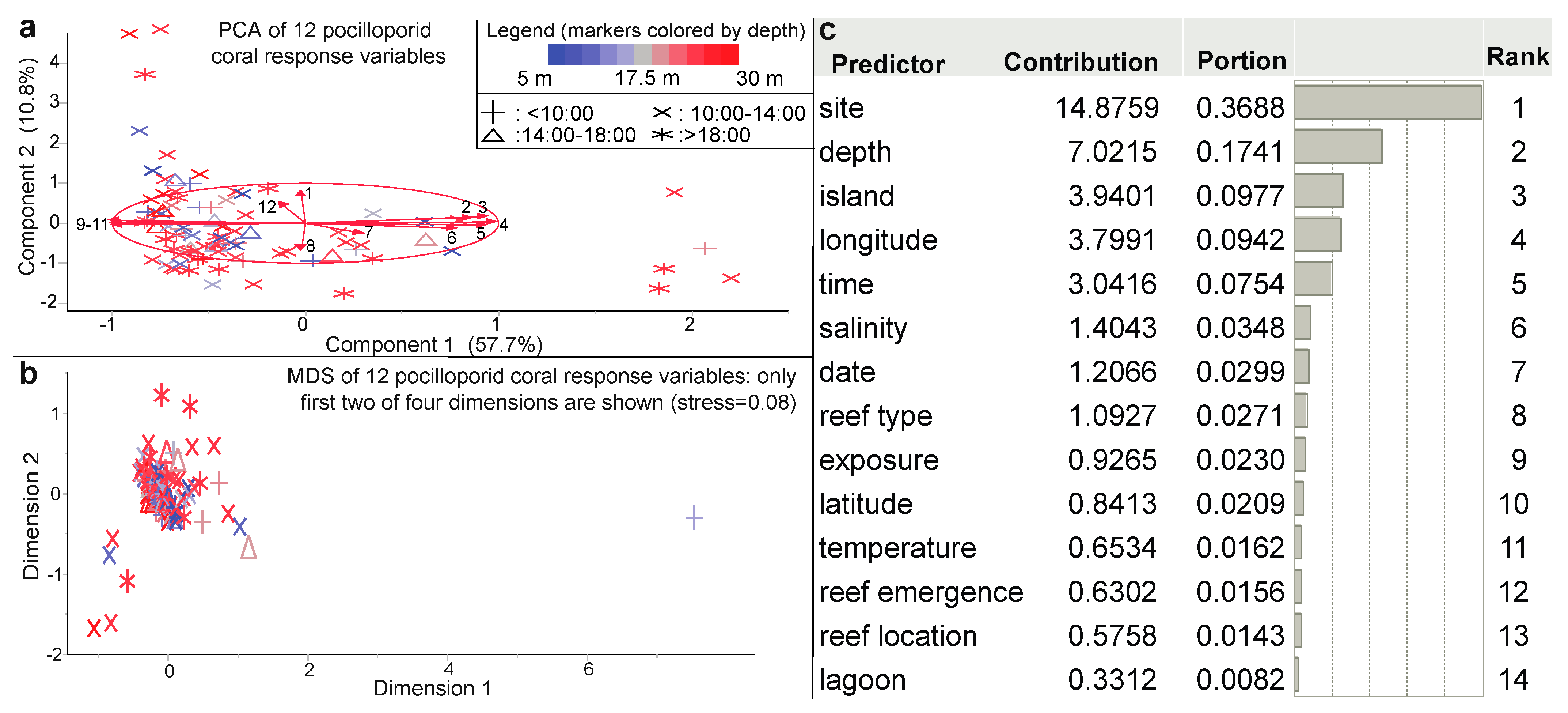
| PCA | 12 PRV a | NA | NA | Figure 4a |
| MDS | 12 PRV a | NA | NA | Figure 4b |
| Factor analysis | 16 PRV | NA | NA | Not shown |
| ENVIRONMENTAL EFFECTS ON CORAL PHYSIOLOGY | ||||
| NP-MANOVA | 4 PRV factors | ENV 1 | NA | Table 2 |
| NIPALS | 4 PRV factors | ENV 1, ENV 2, ENV 3, ENV-RS | Kfold7, val col, val w/test | Table S1 |
| NIPALS | 4 PRV factors | ECO 1, ECO 2, ECO-RS | Kfold7, val col, val w/test | Table S1 |
| NIPALS | 4 PRV factors | (ENV + ECO) 1, (ENV + ECO) 2 | Kfold7, val col, val w/test | Table S1 |
| NIPALS | 16 PRV | ENV 1, ENV 2, ENV 3, ENV-RS | Kfold7, val col, val w/test | Table S1 |
| NIPALS | 16 PRV | ECO 1, ECO 2, ECO-RS | Kfold7, val col, val w/test | Table S1 |
| NIPALS | 16 PRV | (ENV + ECO) 1, (ENV + ECO) 2 | Kfold7, val col, val w/test | Table S1 |
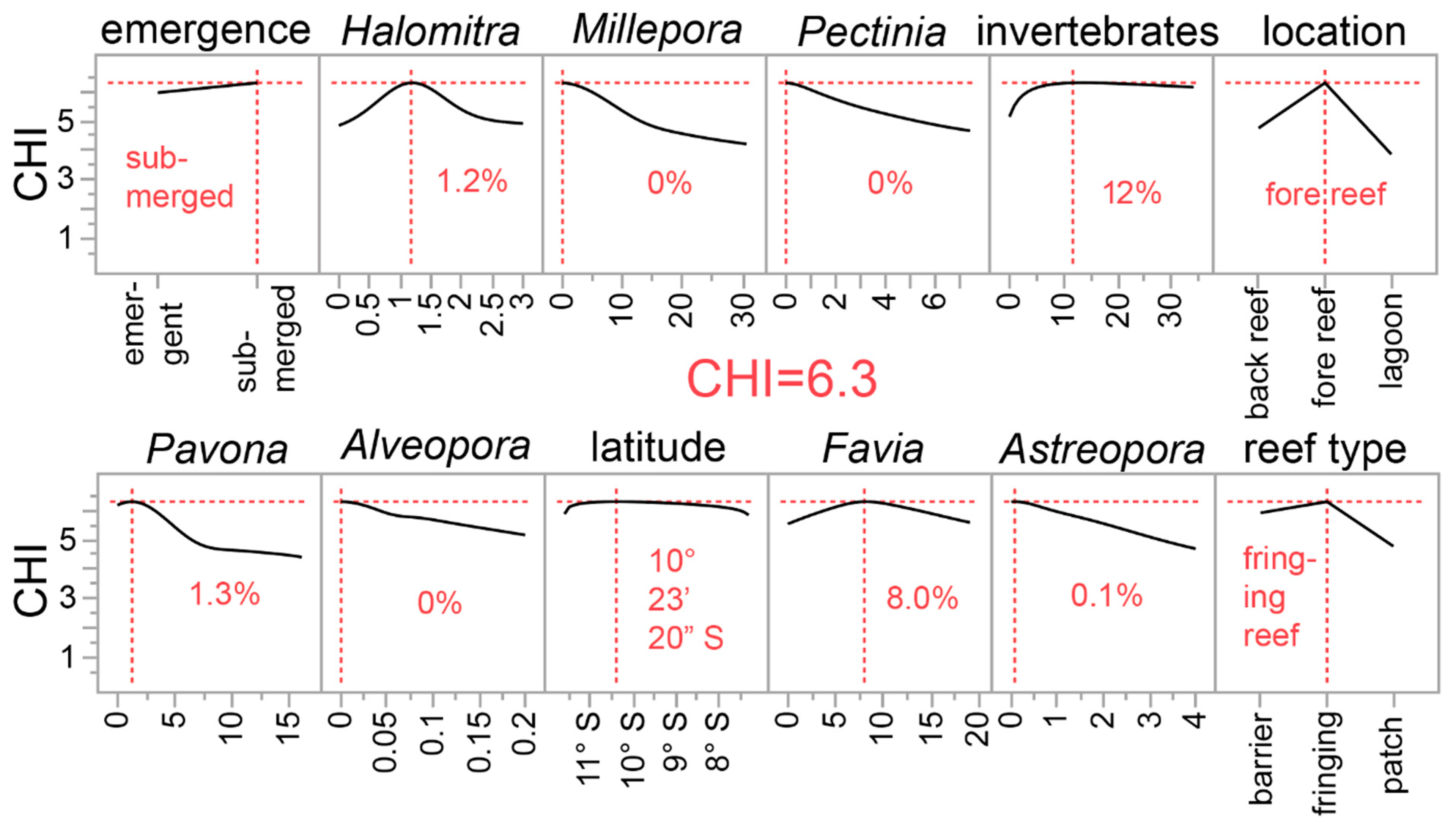
| Predictor screen | CHI | ENV 1 | NA | Figure 4c |
| Model screen | CHI | ENV 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NN GUI-HL(1) | CHI | ENV 1, ENV 2, ENV 3, ENV-RS | Kfold7, val col, val w/test, 20% HB | Table 3 and Table S2 |
| NN GUI-HL(2) | CHI | ENV 1, ENV 2, ENV 3, ENV-RS | Kfold7, val col, val w/test, 20% HB | Table 3 and Table S2 |
| NIPALS | CHI | ENV 1, ENV 2, ENV 3, ENV-RS | Kfold7, val col, val w/test | Table 3 and Table S2 |
| ECOLOGICAL EFFECTS ON CORAL PHYSIOLOGY | ||||
| Predictor screen | CHI | ECO 1 | NA | Table 3 and Table S2 |
| Model screen | CHI | ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NN GUI-HL(1) | CHI | ECO 1, ECO(15) 1 | Kfold7, val col, val w/test, 20% HB | Table 3 and Table S2 |
| NN GUI-HL(2) | CHI | ECO 1, ECO(15) 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NIPALS | CHI | ECO 1, ECO(15) 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| ENVIRONMENTAL + ECOLOGICAL EFFECTS ON CORAL PHYSIOLOGY | ||||
| Predictor screen | CHI | ENV 1 + ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| Model screen | CHI | ENV 1 + ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NN GUI-HL(1) | CHI | ENV 1 + ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NN GUI-HL(2) | CHI | ENV 1 + ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
| NIPALS | CHI | ENV 1 + ECO 1 | Kfold7, val col, val w/test | Table 3 and Table S2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayfield, A.B.; Dempsey, A.C.; Chen, C.-S.; Lin, C. Expediting the Search for Climate-Resilient Reef Corals in the Coral Triangle with Artificial Intelligence. Appl. Sci. 2022, 12, 12955. https://doi.org/10.3390/app122412955
Mayfield AB, Dempsey AC, Chen C-S, Lin C. Expediting the Search for Climate-Resilient Reef Corals in the Coral Triangle with Artificial Intelligence. Applied Sciences. 2022; 12(24):12955. https://doi.org/10.3390/app122412955
Chicago/Turabian StyleMayfield, Anderson B., Alexandra C. Dempsey, Chii-Shiarng Chen, and Chiahsin Lin. 2022. "Expediting the Search for Climate-Resilient Reef Corals in the Coral Triangle with Artificial Intelligence" Applied Sciences 12, no. 24: 12955. https://doi.org/10.3390/app122412955