
Dose–Response Effect of Nitrogen on Microbial Community during Hydrocarbon Biodegradation in Simplified Model System

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Consortium Isolation
2.2. Experimental Design
2.3. Hydrocarbon Biodegradation Analysis
2.4. Metabolic Activity Analysis
2.5. Genetic Analysis of Microbial Population
2.5.1. DNA Isolation
2.5.2. NGS Sequencing
2.5.3. Bioinformatic Analysis
2.5.4. PICRUSt Analysis
2.6. Statistical Analysis
3. Results
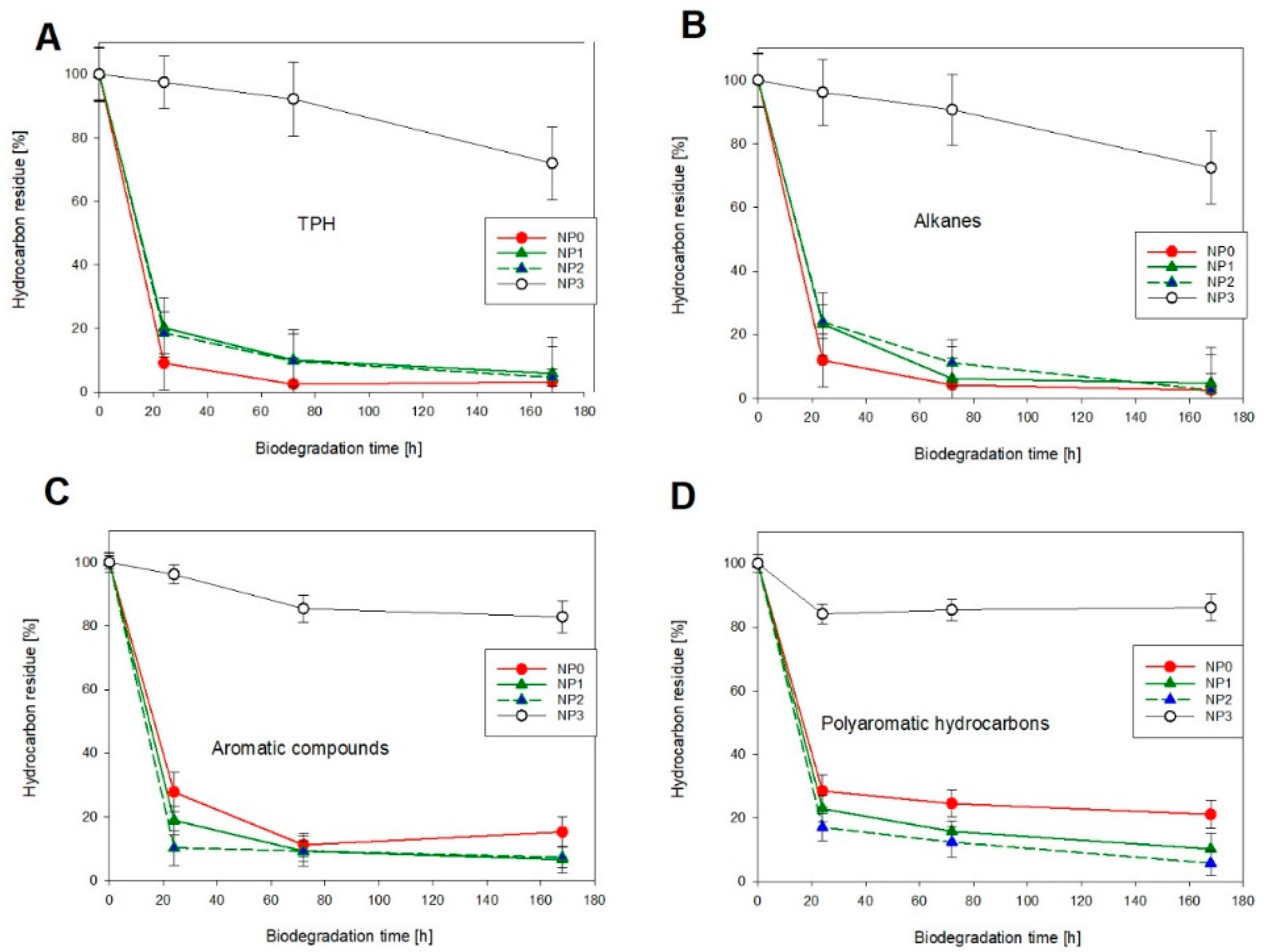
3.1. Hydrocarbon Biodegradation
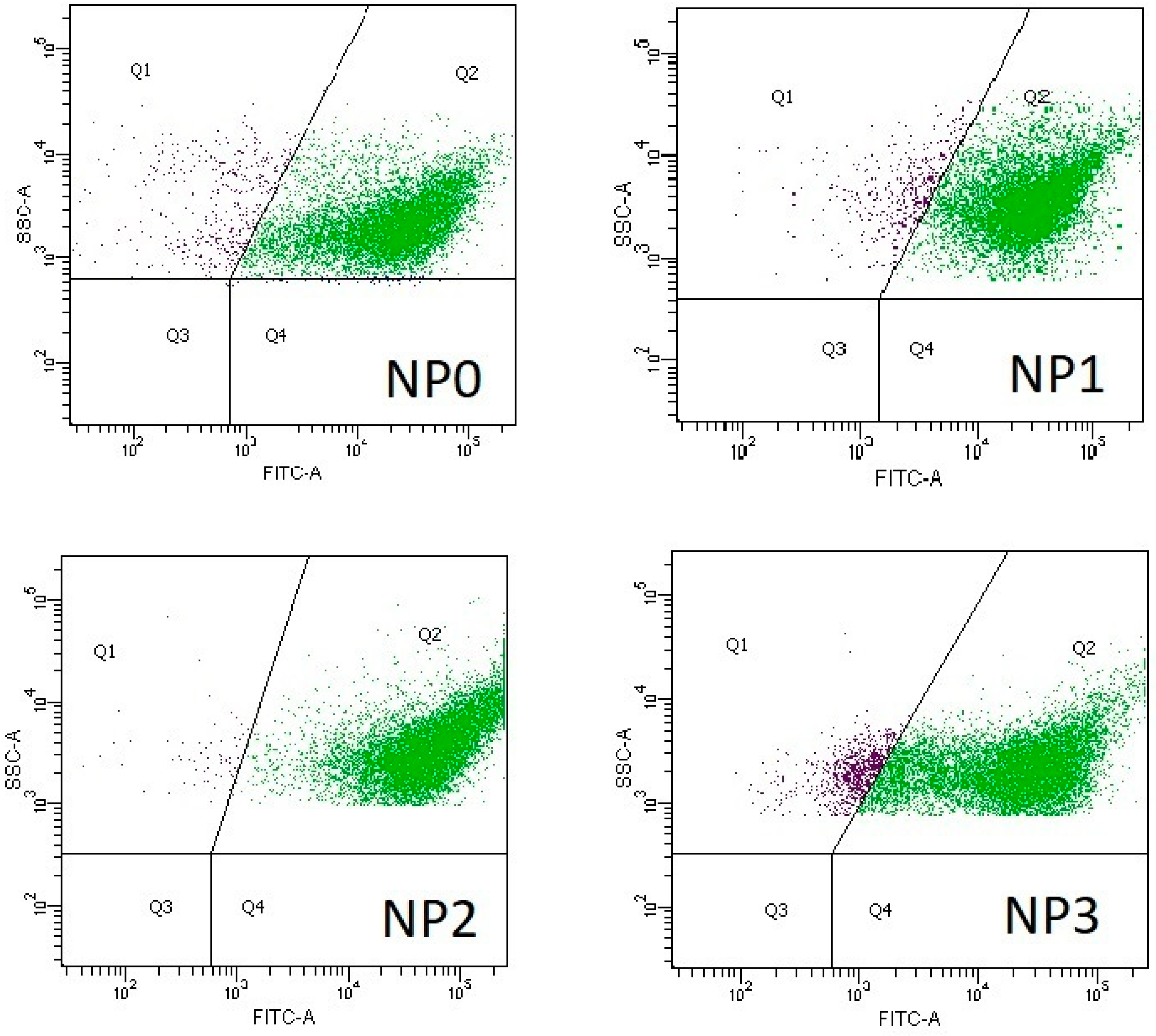
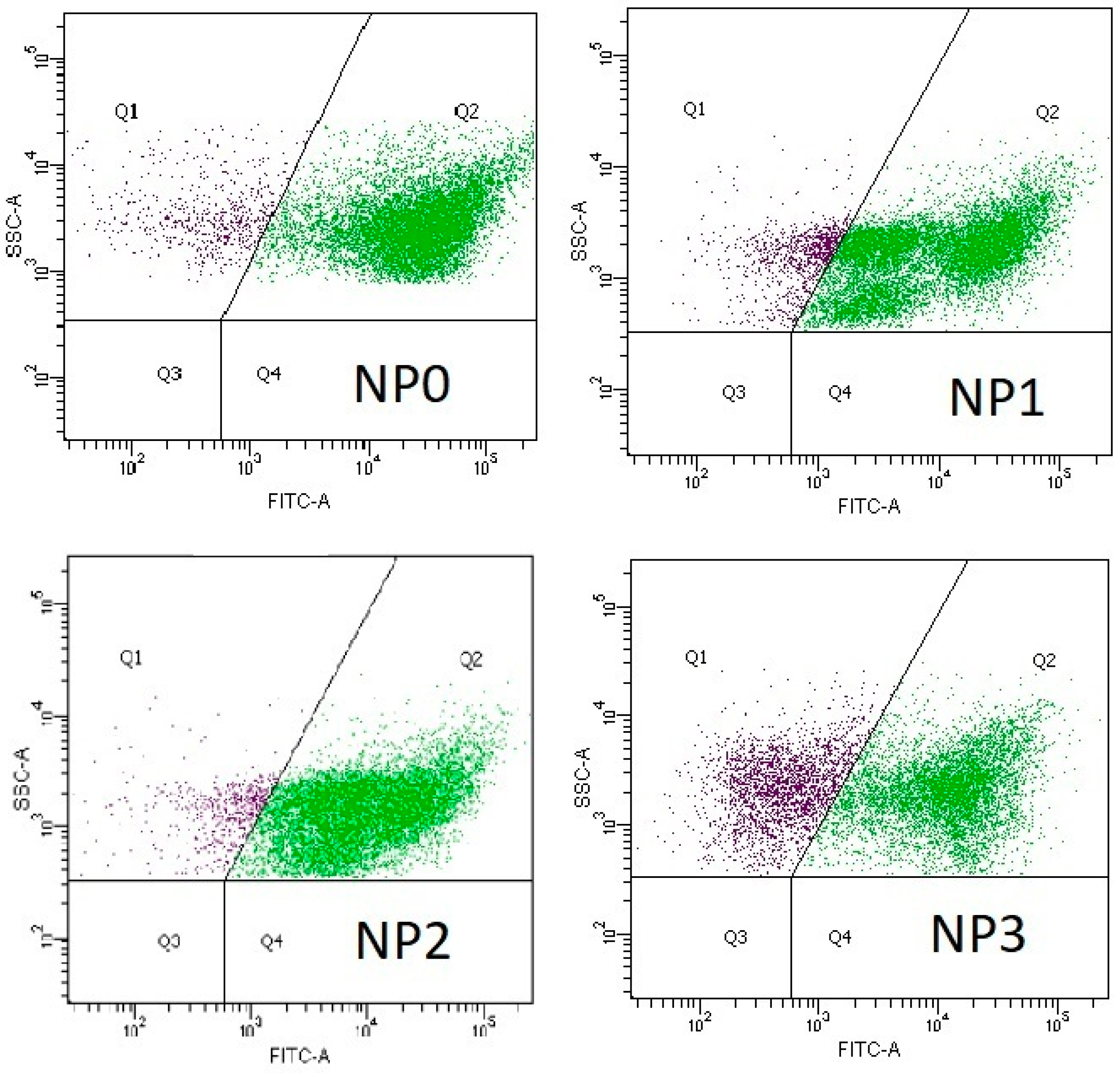
3.2. Microbial Activity
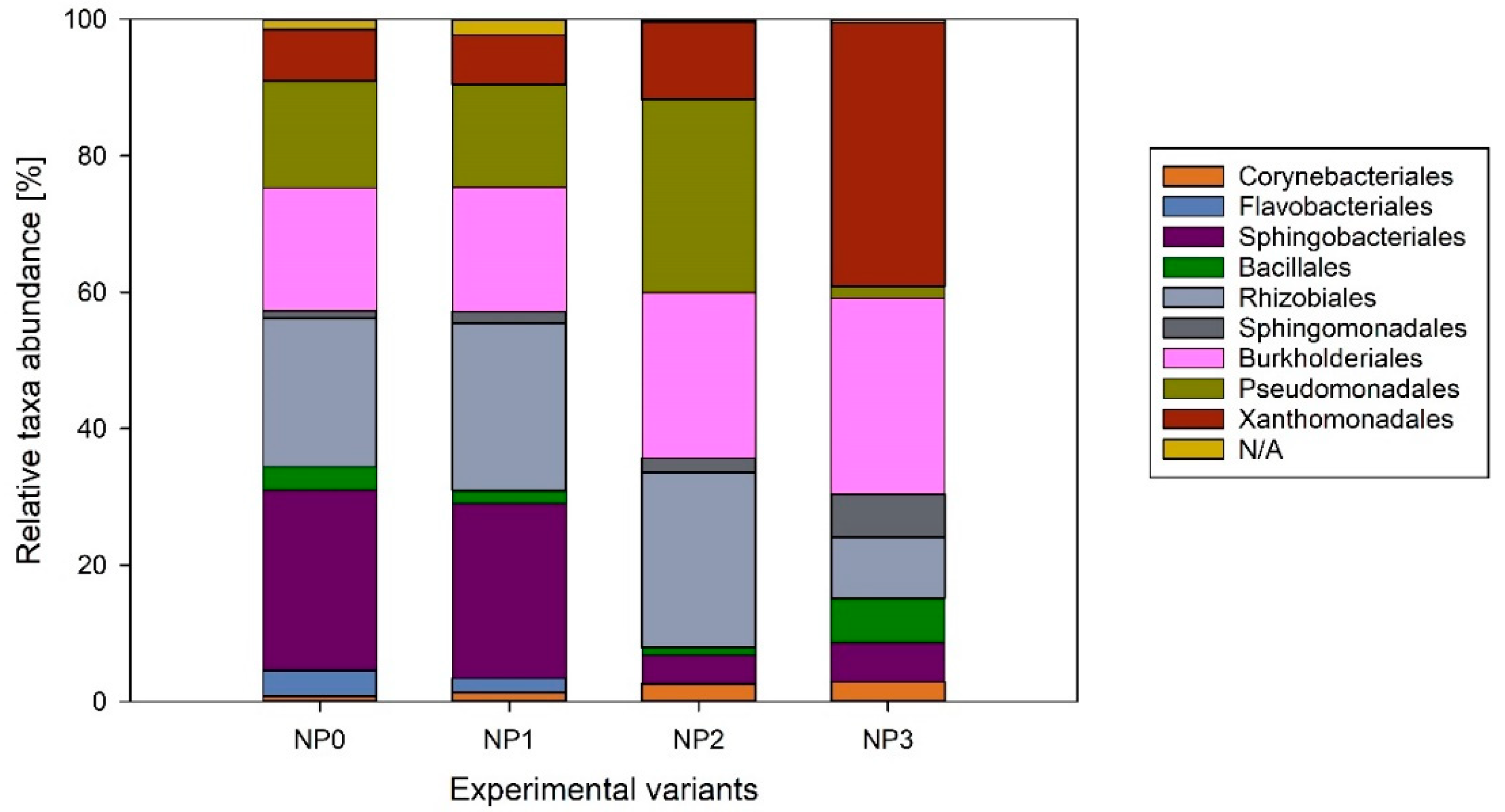
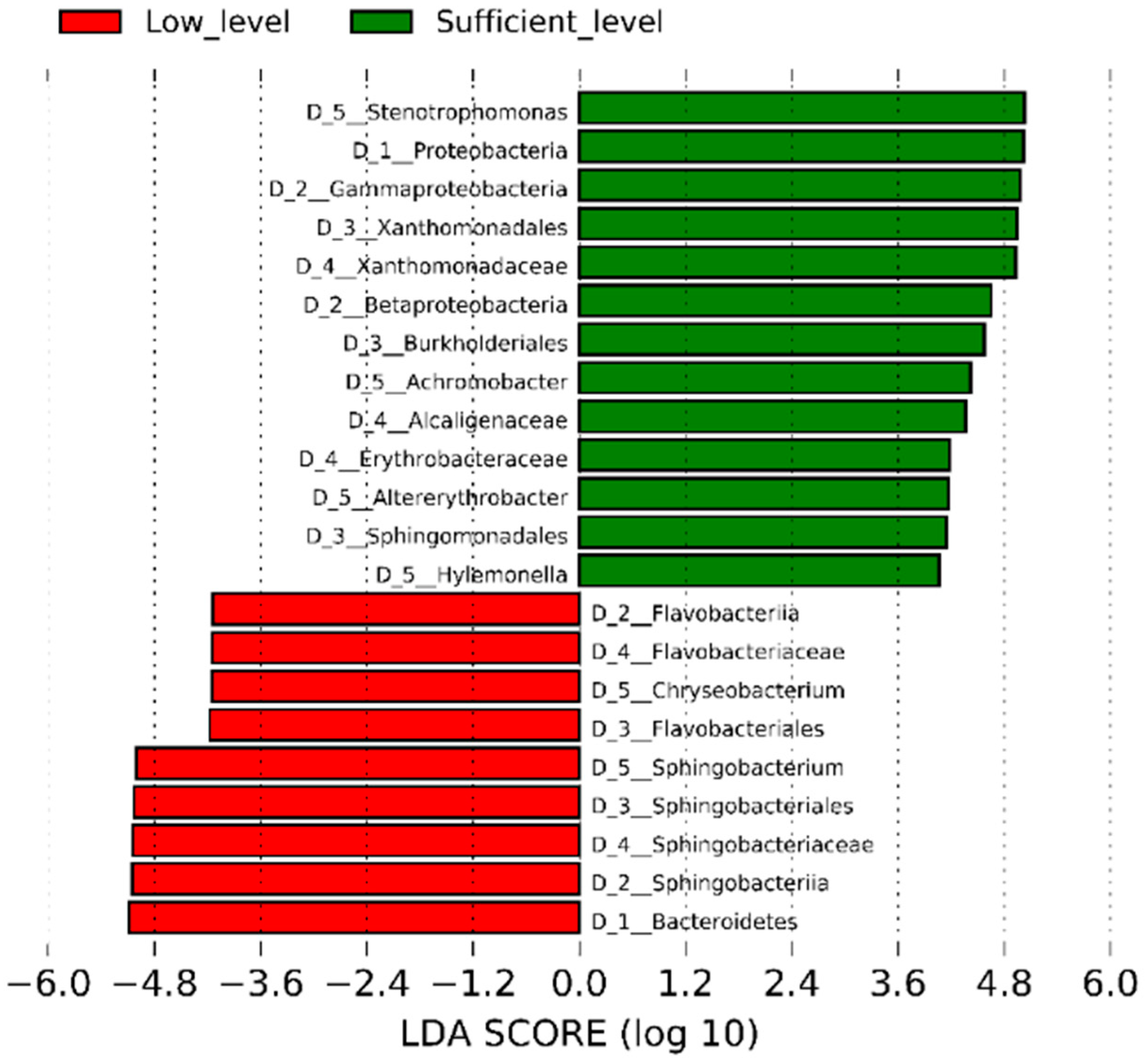
3.3. Biodiversity Analysis
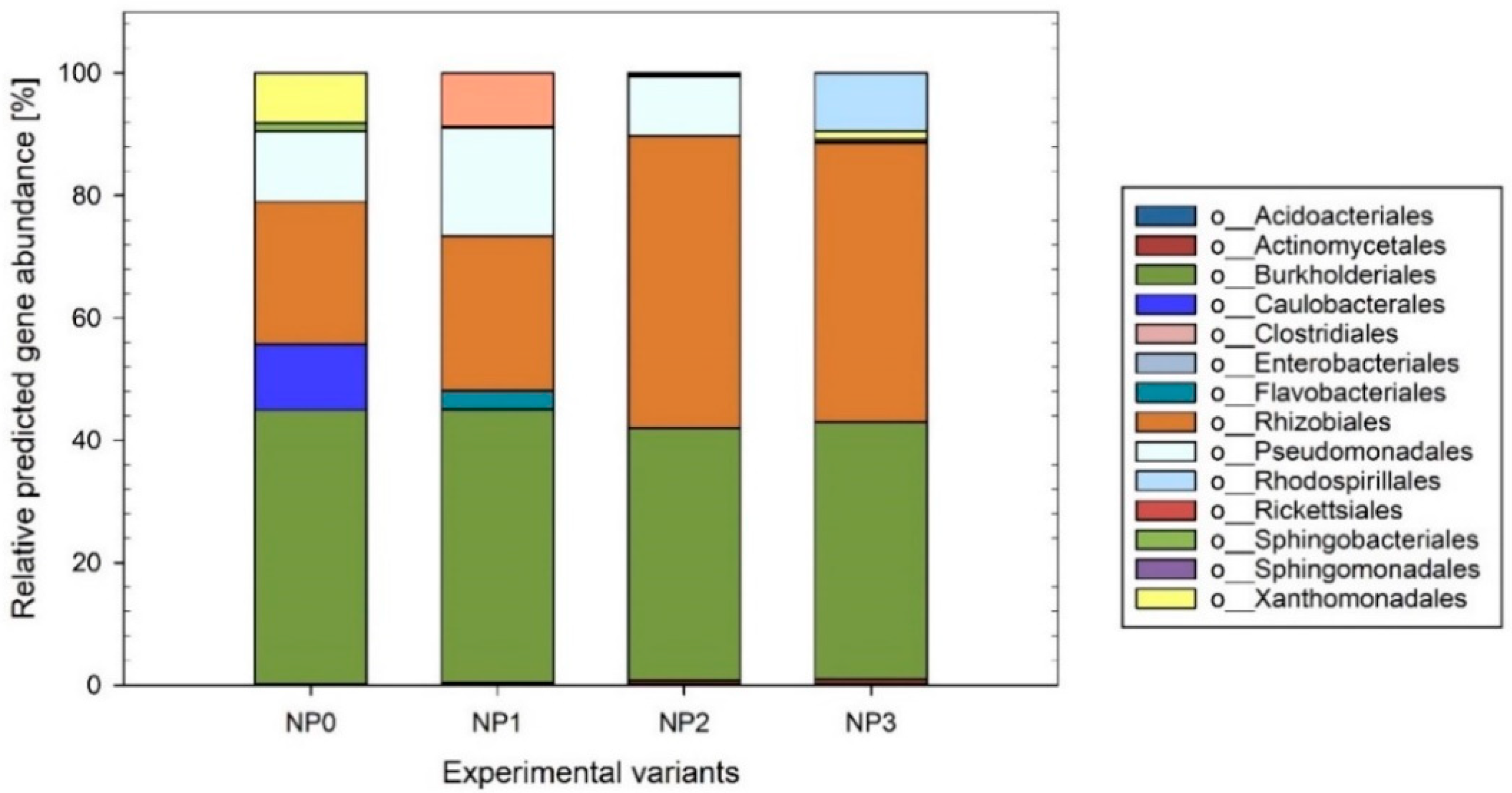
3.4. Prediction of Metabolic Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Sharma, R.; Sharma, A. Application of bioremediation technology in the environment contaminated with petroleum hydrocarbon. Ann. Microbiol. 2013, 63, 417–431. [Google Scholar] [CrossRef]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems-Fate and Microbial Responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef] [Green Version]
- Abha, S.; Singh, C.S. Hydrocarbon pollution: Effects on living organisms, remediation of contaminated environments, and effects of heavy metals co-contamination on bioremediation. In Introduction to Enhanced Oil Recovery (EOR) Processes and Bioremediation of Oil-Contaminated Sites; Romero-Zerón, L., Ed.; InTech Open: Rijeka, Croatia, 2012. [Google Scholar]
- Szczepaniak, Z.; Cyplik, P.; Juzwa, W.; Czarny, J.; Staninska, J.; Piotrowska-Cyplik, A. Antibacterial effect of the Trichoderma viride fungi on soil microbiome during PAH’s biodegradation. Int. Biodeterior. Biodegrad. 2015, 104, 170–177. [Google Scholar] [CrossRef]
- Szczepaniak, Z.; Czarny, J.; Staninska-Pięta, J.; Lisiecki, P.; Zgoła-Grześkowiak, A.; Cyplik, P.; Chrzanowski, Ł.; Wolko, Ł.; Marecik, R.; Juzwa, W.; et al. Influence of soil contamination with PAH on microbial community dynamics and expression level of genes responsible for biodegradation of PAH and production of rhamnolipids. Environ. Sci. Pollut. Res. 2016, 23, 23043–23056. [Google Scholar] [CrossRef] [Green Version]
- Al-Hawash, A.B.; Dragh, M.A.; Li, S.; Alhujaily, A.; Abbood, H.A.; Zhang, X.; Ma, F. Principles of microbial degradation of petroleum hydrocarbons in the environment. Egypt. J. Aquat. Res. 2018, 44, 71–76. [Google Scholar] [CrossRef]
- Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Juzwa, W.; Zgoła-Grześkowiak, A.; Wolko, Ł.; Sydow, Z.; Kaczorowski, Ł.; Powierska-Czarny, J.; Cyplik, P. The impact of natural and synthetic surfactants on bacterial community during hydrocarbon biodegradation. Int. Biodeterior. Biodegrad. 2019, 142, 191–199. [Google Scholar] [CrossRef]
- Steliga, T. Ocena efektywności biodegradacji węglowodorów ropopochodnych w zastarzałym odpadzie z dołu urobkowego Graby-59 w warunkach przemysłowych metodą in-situ. Nafta-Gaz 2014, 70, 351–364. [Google Scholar]
- Sihag, S.; Pathak, H.; Jaroli, D.P. Factors affecting biodegradation of polyaromatic hydrocarbons. Int. J. Pure Appl. 2014, 2, 185–202. [Google Scholar]
- Ouriache, H.; Moumed, I.; Arrar, J.; Abdelkader, N.; Lounici, H. Influence of C/N/P ratio evolution on biodegradation of petroleum hydrocarbons-contaminated soil. ALJEST 2020, 6, 1604–1611. [Google Scholar]
- Tyagi, M.; da Fonseca, M.R.; de Carvalho, C.C.C.R. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 2011, 22, 231–241. [Google Scholar] [CrossRef]
- Adams, G.O.; Fufeyin, P.T.; Okoro, S.E.; Ehinomen, I. Bioremediation, biostimulation and bioaugmentation: A review. Int. J. Environ. Bioremediat. Biodegrad. 2015, 3, 28–39. [Google Scholar] [CrossRef]
- Agarry, S.E.; Owabor, C.N. Anaerobic bioremediation of marine sediment artificially contaminated with anthracene and naphthalene. Environ. Technol. 2011, 32, 1375–1381. [Google Scholar] [CrossRef]
- Silva-Castro, G.A.; Rodelas, B.; Perucha, C.; Laguna, J.; González-López, J.; Calvo, C. Bioremediation of diesel-polluted soil using biostimulation as post-treatment after oxidation with Fenton-like reagents: Assays in a pilot plant. Sci. Total Environ. 2013, 15, 347–355. [Google Scholar] [CrossRef]
- Wu, M.; Dick, W.A.; Li, W.; Wang, X.; Yang, Q.; Wang, T.; Xu, L.; Zhang, M.; Chen, L. Bioaugmentation and biostimulation of hydrocarbon degradation and the microbial community in a petroleum-contaminated soil. Int. Biodeterior. Biodegrad. 2016, 107, 158–164. [Google Scholar] [CrossRef]
- Suja, F.; Rahim, F.; Taha, M.R.; Hambali, N.; Razali, M.R.; Khalid, A.; Hamzah, A. Effects of local microbial bioaugmentation and biostimulation on the bioremediation of total petroleum hydrocarbons (TPH) in crude oil contaminated soil based on laboratory and field observations. Int. Biodeterior. Biodegrad. 2014, 90, 115–122. [Google Scholar] [CrossRef]
- Hamoudi-Belarbi, L.; Demdoum, S.; Medjras, S.; Hamoudi, S. Combination of bioaugmentation and biostimulation as an oil-drilling mud contaminated soil bioremediation treatment. Appl. Ecol. Environ. Res. 2019, 17, 15463–15475. [Google Scholar] [CrossRef]
- Vidali, M. Bioremediation. An overview. Pure Appl. Chem. 2001, 73, 1163–1172. [Google Scholar] [CrossRef]
- Kumavath, R.N.; Deverapalli, P. Scientific swift in bioremediation: An overview. In Bioremediation; Patil, Y., Ed.; InTech Open: Rijeka, Croatia, 2013. [Google Scholar]
- Walworth, J.; Pond, A.; Snape, I.; Rayner, J.; Ferguson, S.; Harvey, P. Nitrogen Requirements for Maximizing Petroleum Bioremediation in a Sub-Antarctic Soil. Cold Reg. Sci. Technol. 2007, 48, 84–91. [Google Scholar] [CrossRef]
- Ruberto, L.; Vazquez, S.C.; Mac Cormack, W.P. Effectiveness of the natural bacterial flora, biostimulation and bioaugmentation on the bioremediation of a hydrocarbon contaminated Antarctic soil. Int. Biodeterior. Biodegrad. 2003, 52, 115–125. [Google Scholar] [CrossRef]
- Liu, P.W.; Chang, T.C.; Whang, L.M.; Kao, C.H.; Pan, P.T.; Cheng, S.S. Bioremediation of petroleum hydrocarbon contaminated soil: Effects of strategies and microbial community shift. Int. Biodeterior. Biodegrad. 2011, 65, 1119–1127. [Google Scholar] [CrossRef]
- Dolinšek, J.; Goldschmidt, F.; Johnson, D.R. Synthetic microbial ecology and the dynamic interplay between microbial genotypes. FEMS Microbiol. Rev. 2016, 40, 961–979. [Google Scholar] [CrossRef] [Green Version]
- Said, S.B.; Or, D. Synthetic microbial ecology: Engineering habitats for modular consortia. Front. Microbiol. 2017, 8, 1125. [Google Scholar] [CrossRef] [Green Version]
- Czarny, J.; Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Juzwa, W.; Wolniewicz, A.; Marecik, R.; Ławniczak, Ł.; Chrzanowski, Ł. Acinetobacter sp. as the key player in diesel oil degrading community exposed to PAHs and heavy metals. J. Hazard. Mater. 2020, 383, 121168. [Google Scholar] [CrossRef]
- Cheng, X.; Zhao, T.; Fu, X.; Hu, Z. Identification of nitrogen compounds in RFCC diesel oil by mass spectrometry. Fuel Process. Technol. 2004, 85, 1463–1472. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Brg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betly, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garret, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Staley, C.; Gould, T.J.; Wang, P.; Phillips, J.; Cotner, J.B.; Sadowsky, M.J. Core functional traits of bacterial communities in the Upper Mississippi River show limited variation in response to land cover. Front. Microbiol. 2014, 5, 414. [Google Scholar] [CrossRef]
- Czarny, J.; Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Wolko, Ł.; Staninski, K.; Hornik, B.; Cyplik, P. Assessment of soil potential to natural attenuation and autochthonous bioaugmentation using microarray and functional predictions from metagenome profiling. Ann. Microbiol. 2019, 9, 945–955. [Google Scholar] [CrossRef]
- Staninska-Pięta, J.; Czarny, J.; Piotrowska-Cyplik, A.; Juzwa, W.; Wolko, Ł.; Nowak, J.; Cyplik, P. Heavy Metals as a Factor Increasing the Functional Genetic Potential of Bacterial Community for Polycyclic Aromatic Hydrocarbon Biodegradation. Molecules 2020, 25, 319. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, T.; Gupta, G.; Sharma, A.; Kaur, B.; El-Sheikh, M.A.; Alyemeni, M.N. Metagenomic analysis exploring taxonomic and functional diversity of bacterial communities of a Himalayan urban fresh water lake. PLoS ONE 2021, 16, e0248116. [Google Scholar] [CrossRef]
- Hornik, B.; Czarny, J.; Staninska-Pięta, J.; Wolko, Ł.; Cyplik, P.; Piotrowska-Cyplik, A. The Raw Milk Microbiota from Semi-Subsistence Farms Characteristics by NGS Analysis Method. Molecules 2021, 26, 5029. [Google Scholar] [CrossRef]
- Ijoma, G.N.; Nkuna, R.; Mutungwazi, A.; Rashama, C.; Matambo, T.S. Applying PICRUSt and 16S rRNA functional characterisation to predicting co-digestion strategies of various animal manures for biogas production. Sci. Rep. 2021, 11, 19913. [Google Scholar] [CrossRef]
- Li, J.; Huang, B.; Long, J. Effects of different antimony contamination levels on paddy soil bacterial diversity and community structure. Ecotoxicol. Environ. Saf. 2021, 220, 112339. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Seo, J.S.; Keum, Y.S.; Li, Q.X. Bacterial degradation of aromatic compounds. Int. J. Environ. Res. 2009, 6, 278–309. [Google Scholar] [CrossRef]
- Staninska, J.; Szczepaniak, Z.; Staninski, K.; Czarny, J.; Piotrowska-Cyplik, A.; Nowak, J.; Marecik, R.; Chrzanowski, Ł.; Cyplik, P. High Voltage Electrochemiluminescence (ECL) as a New Method for Detection of PAH During Screening for PAH-Degrading Microbial Consortia. Water Air Soil Pollut. 2015, 226, 270. [Google Scholar] [CrossRef] [Green Version]
- Czarny, J.; Staninska-Pięta, J.; Powierska-Czarny, J.; Nowak, J.; Wolko, Ł.; Piotrowska-Cyplik, A. Metagenomic analysis of soil bacterial community and level of genes responsible for biodegradation of aromatic hydrocarbons. Pol. J. Microbiol. 2017, 66, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, L.M.; Pfaender, F.K. The effect of inorganic and organic supplements on the microbial degradation of phenanthrene and pyrene in soils. Biodegradation 1997, 8, 1–13. [Google Scholar] [CrossRef]
- Chaîneau, C.H.; Rougeux, G.; Yéprémian, C.; Oudot, J. Effects of nutrient concentration on the biodegradation of crude oil and associated microbial populations in the soil. Soil Biol. Biochem. 2005, 37, 1490–1497. [Google Scholar] [CrossRef]
- Müller, T.; Walter, B.; Wirtz, A.; Burkovski, A. Ammonium toxicity in bacteria. Curr. Microbiol. 2006, 52, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Leejeerajumnean, A.; Ames, J.M.; Owens, J.D. Effect of ammonia on the growth of Bacillus species and some other bacteria. Lett. Appl. Microbiol. 2000, 30, 385–389. [Google Scholar] [CrossRef]
- Avila-Jimenez, M.-L.; Burns, G.; He, Z.; Zhou, J.; Hodson, A.; Avila-Jimenez, J.M.-L.; Pearce, D. Functional Associations and Resilience in Microbial Communities. Microorganisms 2020, 8, 951. [Google Scholar] [CrossRef]
- Li, C.; Yan, K.; Tang, L.; Jia, Z.; Li, Y. Change in deep soil microbial communities due to long-term fertilization. Soil Biol. Biochem. 2014, 75, 264–272. [Google Scholar] [CrossRef]
- Sun, R.; Wang, F.; Hu, C.; Liu, B. Metagenomics reveals taxon-specific responses of the nitrogen-cycling microbial community to long-term nitrogen fertilization. Soil Biol. Biochem. 2021, 156, 108214. [Google Scholar] [CrossRef]
- Shin, B.; Bociu, I.; Kolton, M.; Huettel, M.; Kostka, J.E. Succession of microbial populations and nitrogen-fixation associated with the biodegradation of sediment-oil-agglomerates buried in a Florida sandy beach. Sci. Rep. 2019, 9, 19401. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.; Kumar, V.; Pal, A.K. Microbial Degradation of High Molecular Weight Polycyclic Aromatic Hydrocarbons with Emphasis on Pyrene. Polycycl. Aromat. Compd. 2019, 39, 124–138. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, Z.; Wang, H. Metagenomic analysis exhibited the co-metabolism of polycyclic aromatic hydrocarbons by bacterial community from estuarine sediment. Environ. Int. 2019, 129, 308–319. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Estimated C:N Ratio | Experimental Variant Description |
|---|---|---|
| NP0 | No nitrogen supplementation (control sample) | |
| NP1 | 1:1 | Low nitrogen supplementation |
| NP2 | 10:1 | Moderate nitrogen supplementation |
| NP3 | 30:1 | Excessive nitrogen supplementation |
| Diversity Index | NP0 | NP1 | NP2 | NP3 |
|---|---|---|---|---|
| OTU number | 96 ± 4 | 101 ± 5 | 100 ± 2 | 85 ± 4 |
| Simpson’s index | 0.85 ± 0.02 | 0.84 ± 0.02 | 0.84 ± 0.01 | 0.79 ± 0.02 |
| Phylogenetic diversity | 4.41 ± 0.07 | 4.57 ± 0.11 | 4.43 ± 0.08 | 4.26 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staninska-Pięta, J.; Czarny, J.; Juzwa, W.; Wolko, Ł.; Cyplik, P.; Piotrowska-Cyplik, A. Dose–Response Effect of Nitrogen on Microbial Community during Hydrocarbon Biodegradation in Simplified Model System. Appl. Sci. 2022, 12, 6012. https://doi.org/10.3390/app12126012
Staninska-Pięta J, Czarny J, Juzwa W, Wolko Ł, Cyplik P, Piotrowska-Cyplik A. Dose–Response Effect of Nitrogen on Microbial Community during Hydrocarbon Biodegradation in Simplified Model System. Applied Sciences. 2022; 12(12):6012. https://doi.org/10.3390/app12126012
Chicago/Turabian StyleStaninska-Pięta, Justyna, Jakub Czarny, Wojciech Juzwa, Łukasz Wolko, Paweł Cyplik, and Agnieszka Piotrowska-Cyplik. 2022. "Dose–Response Effect of Nitrogen on Microbial Community during Hydrocarbon Biodegradation in Simplified Model System" Applied Sciences 12, no. 12: 6012. https://doi.org/10.3390/app12126012