
Study on the Development and Growth of Coral Larvae

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Species
2.2. Sample Collection
2.3. Fertilization Experiment
2.4. Cultivation and Feeding
2.5. Observation Records
3. Results
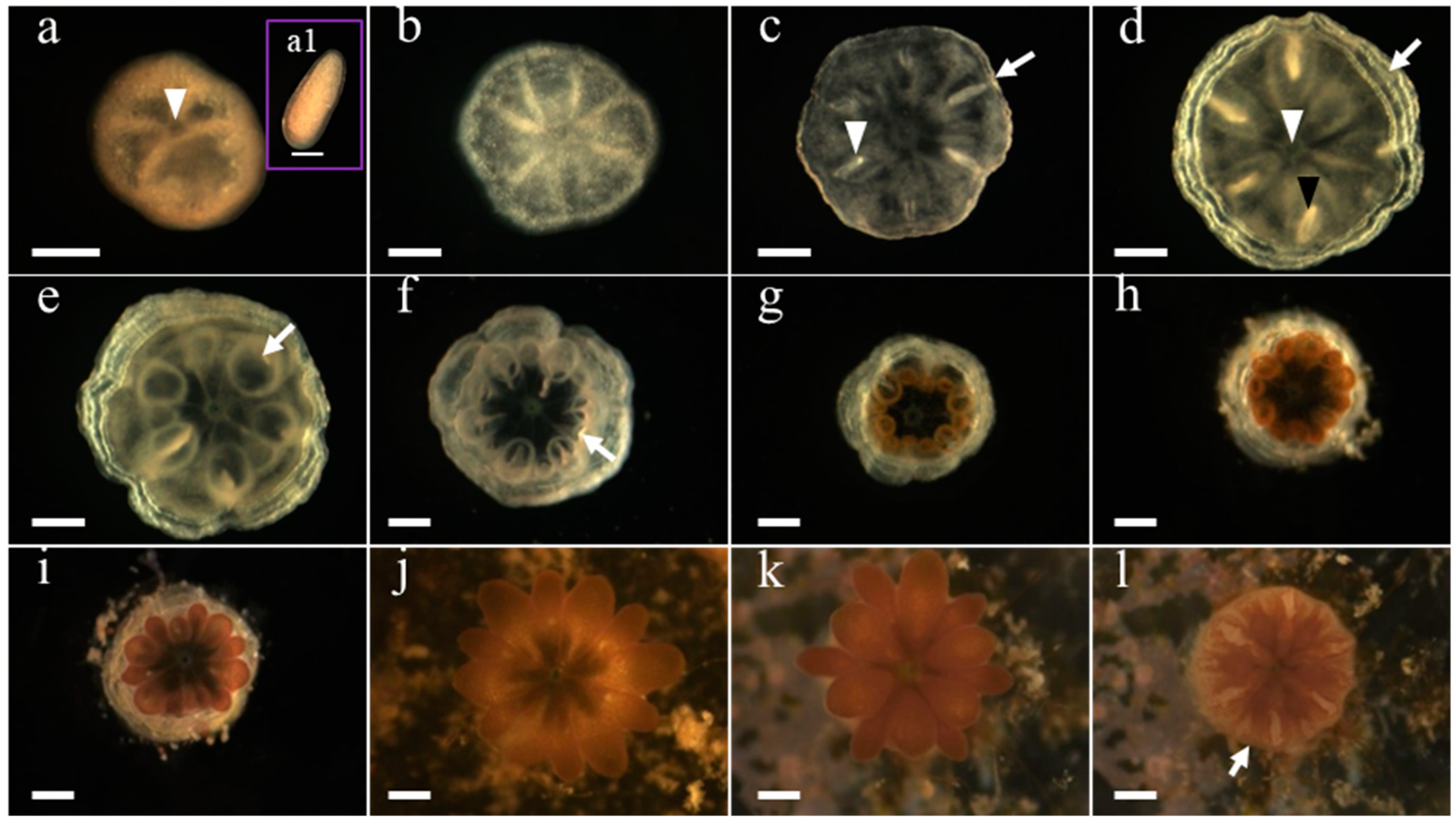
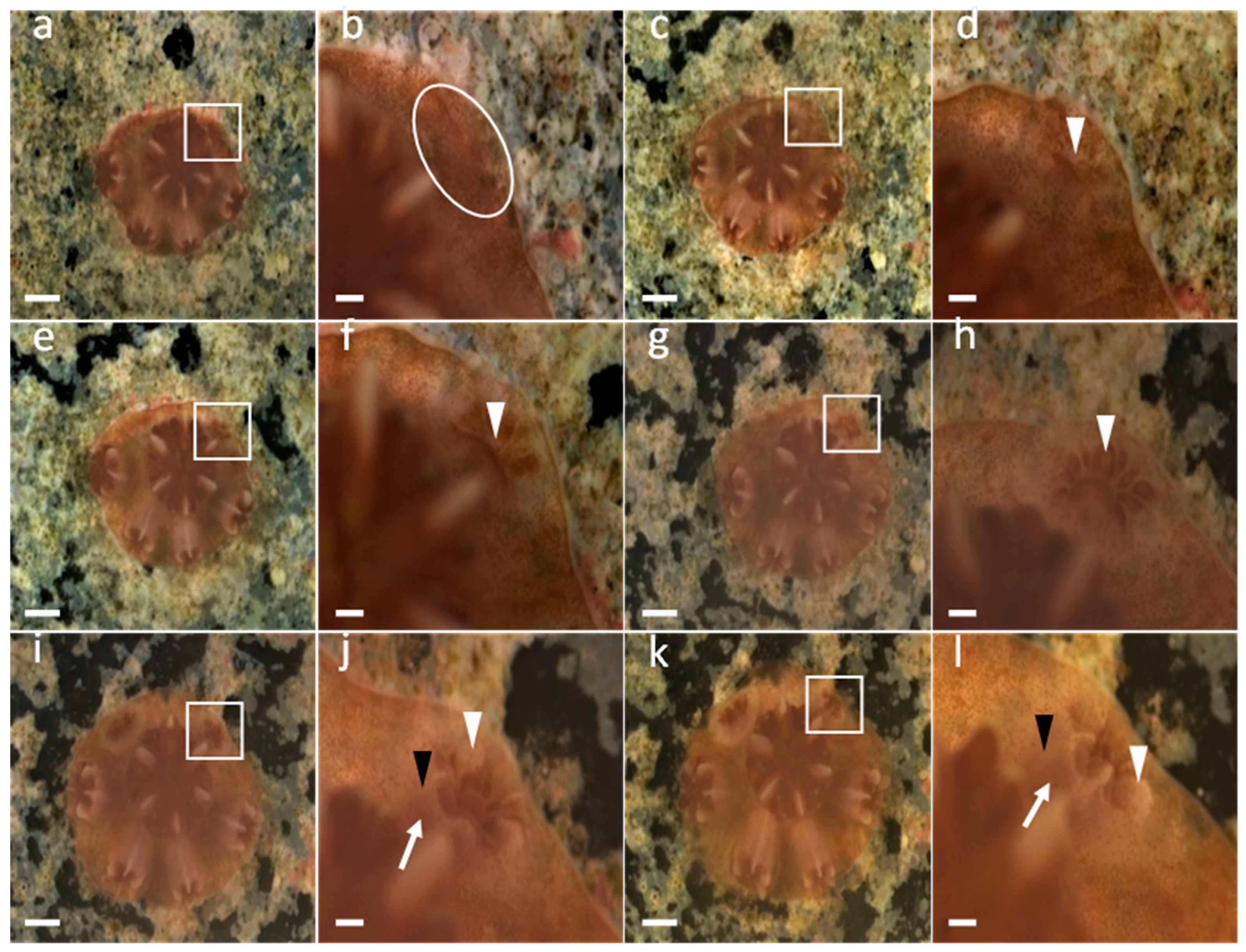
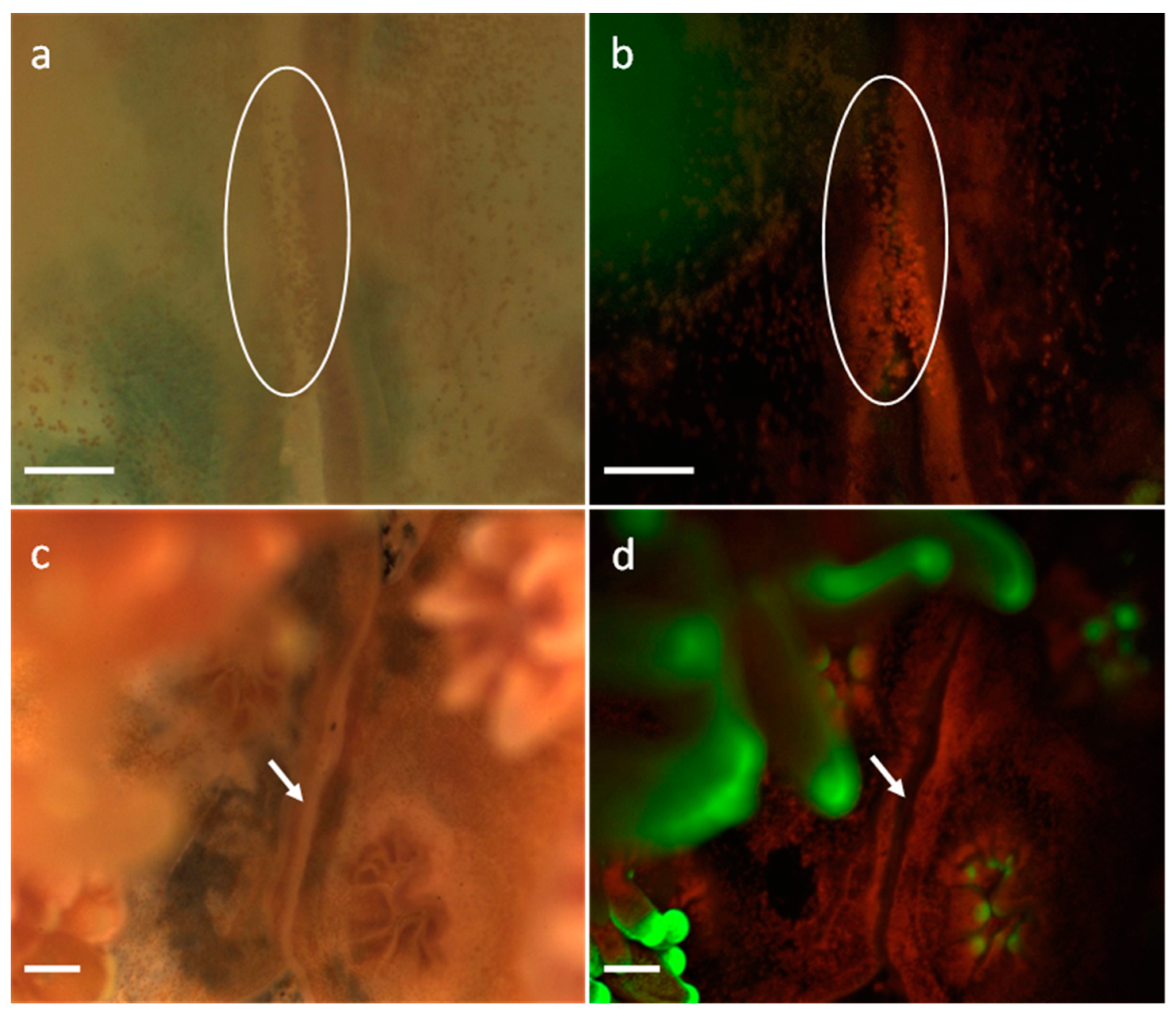
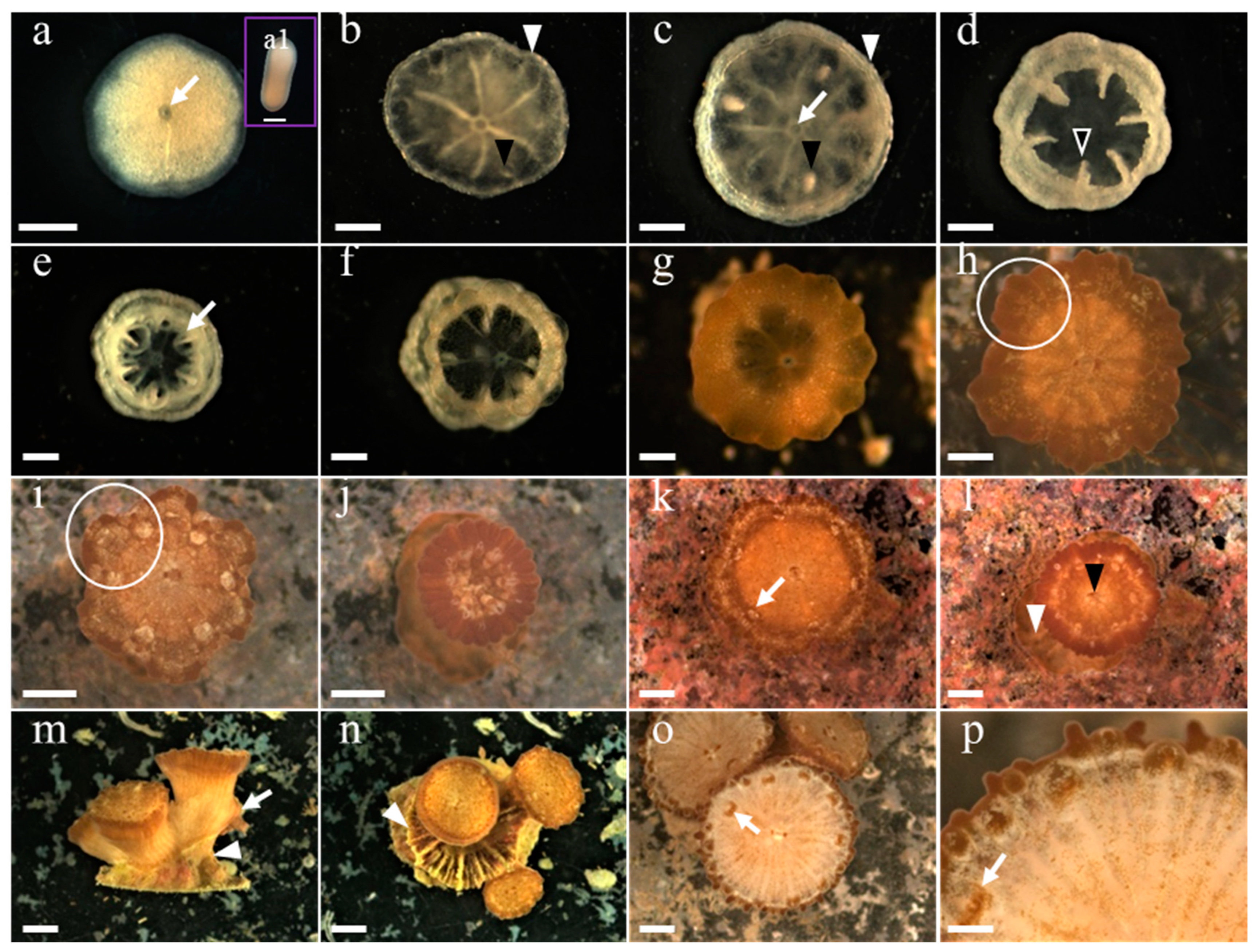
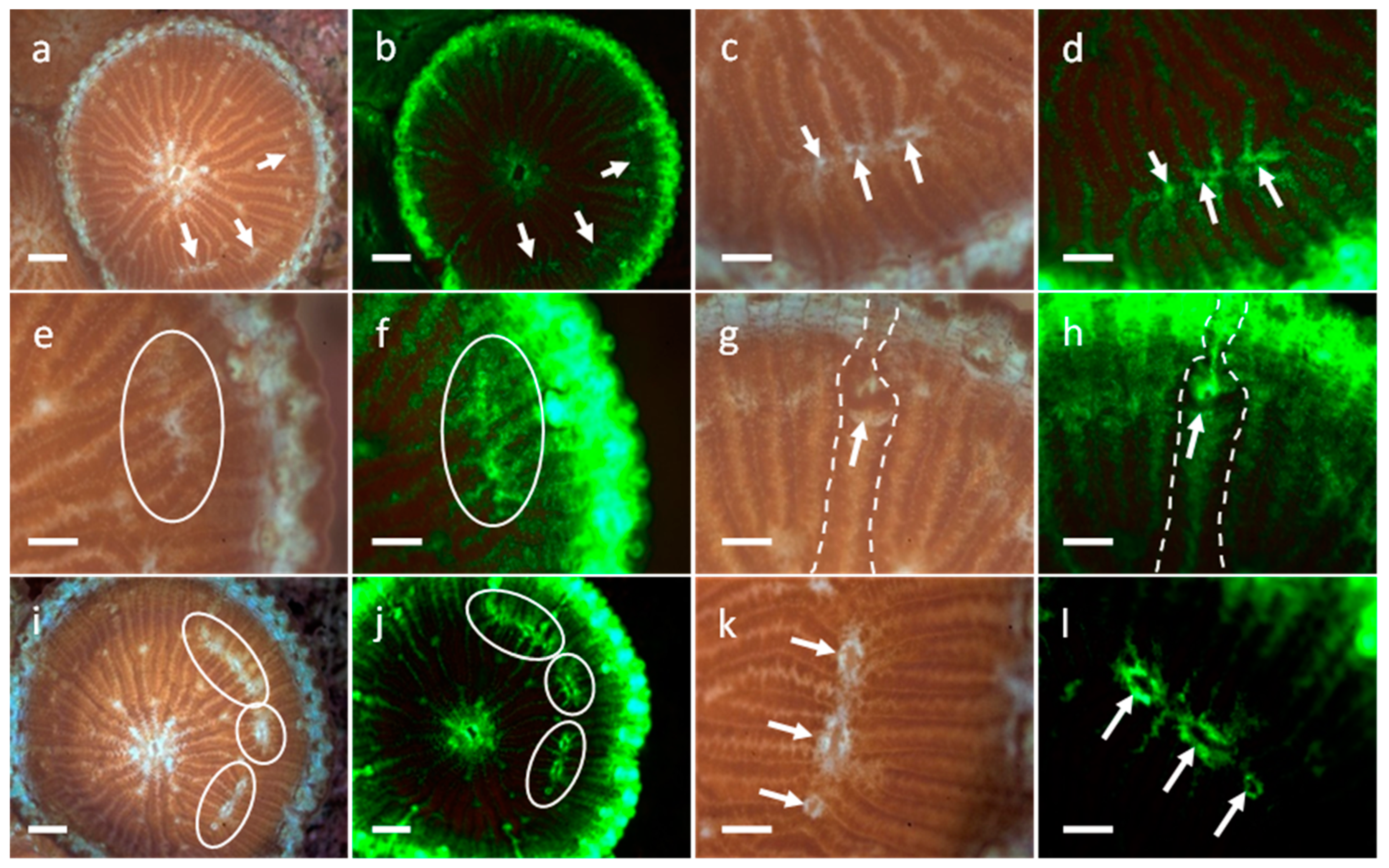
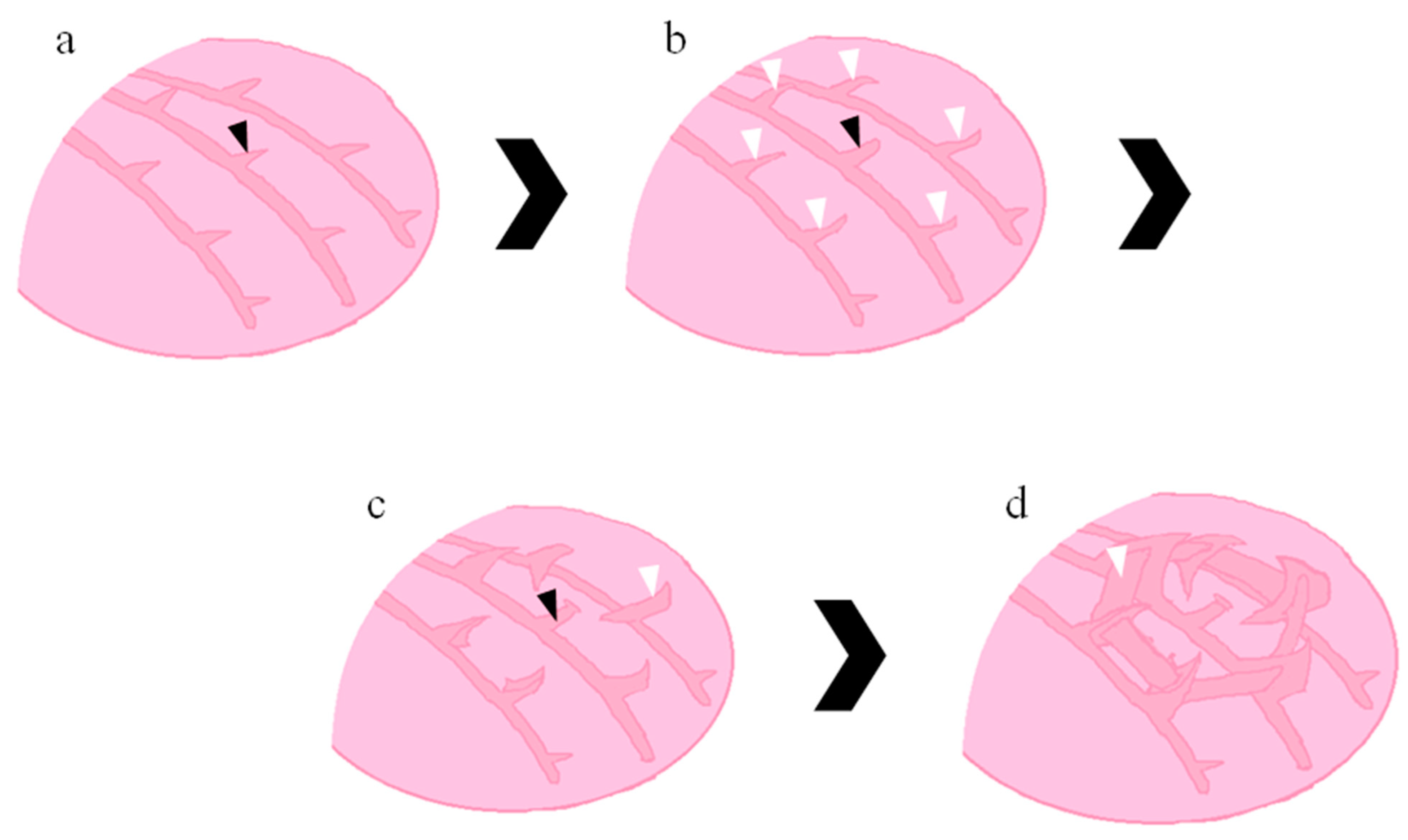
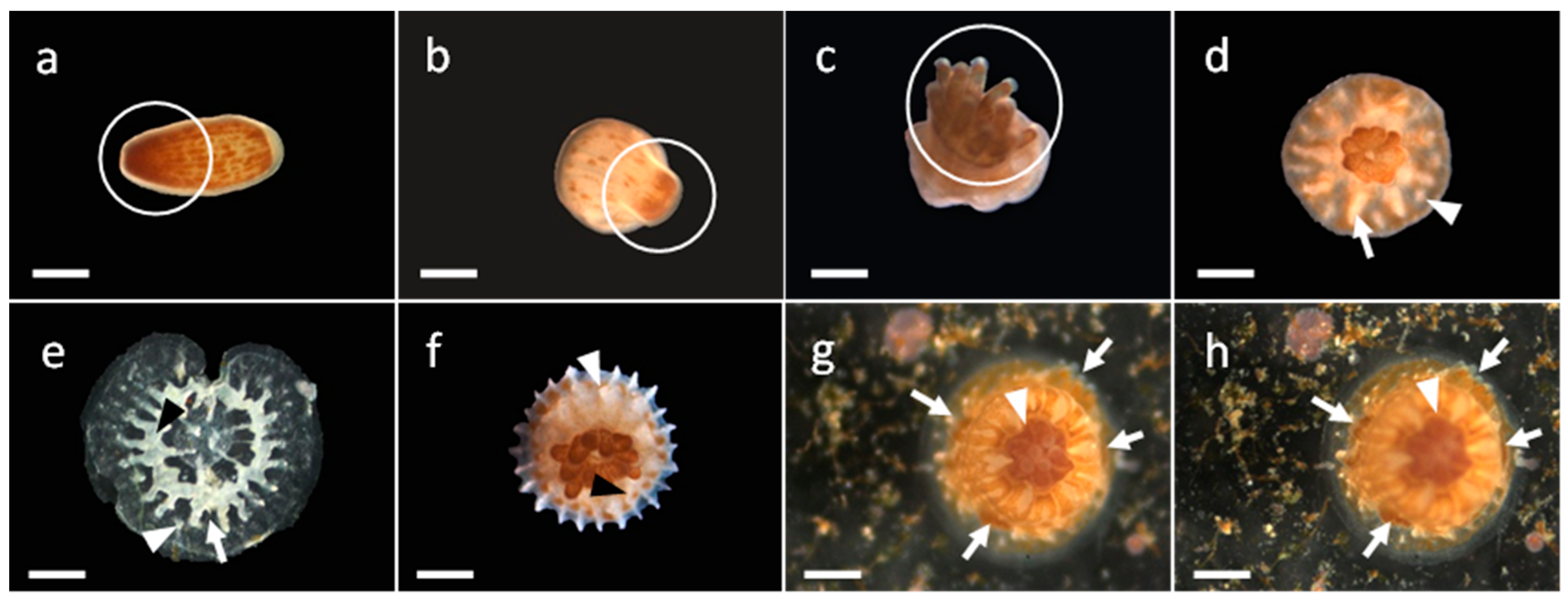
3.1. Development of G. fascicularis Spats
3.2. Development of M. elephantotus Spats
3.3. Development of S. caliendrum Spats
3.4. Development of P. verrucosa Spats
3.5. Settlement and Post-Settlement Survival Rates of Coral Larvae and Spats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Edwards, A.J.; Guest, J.R.; Heyward, A.J.; Villanueva, R.D.; Baria, M.V.; Bollozos, I.S.; Golbuu, Y. Direct seeding of mass-cultured coral larvae is not an effective option for reef rehabilitation. Mar. Ecol. Prog. Ser. 2015, 525, 105–116. [Google Scholar] [CrossRef]
- Randall, C.J.; Negri, A.P.; Quigley, K.M.; Foster, T.; Ricardo, G.F.; Webster, N.S.; Bay, L.K.; Harrison, P.L.; Babcock, R.C.; Heyward, A.J. Sexual production of corals for reef restoration in the Anthropocene. Mar. Ecol. Prog. Ser. 2020, 635, 203–232. [Google Scholar] [CrossRef]
- Shikina, S.; Chiu, Y.-L.; Chung, Y.-J.; Chen, C.-J.; Lee, Y.-H.; Chang, C.-F. Oocytes express an endogenous red fluorescent protein in a stony coral, Euphyllia ancora: A potential involvement in coral oogenesis. Sci. Rep. 2016, 6, 25868. [Google Scholar] [CrossRef] [PubMed]
- Portune, K.J.; Voolstra, C.R.; Medina, M.; Szmant, A.M. Development and heat stress-induced transcriptomic changes during embryogenesis of the scleractinian coral Acropora palmata. Mar. Genom. 2010, 3, 51–62. [Google Scholar] [CrossRef]
- Heyward, A.J.; Negri, A.P. Plasticity of larval pre-competency in response to temperature: Observations on multiple broadcast spawning coral species. Coral Reefs 2010, 29, 631–636. [Google Scholar] [CrossRef]
- Hayward, D.C.; Grasso, L.C.; Saint, R.; Miller, D.J.; Ball, E.E. The organizer in evolution–gastrulation and organizer gene expression highlight the importance of Brachyury during development of the coral, Acropora millepora. Dev. Biol. 2015, 399, 337–347. [Google Scholar] [CrossRef]
- Tebben, J.; Motti, C.A.; Siboni, N.; Tapiolas, D.M.; Negri, A.P.; Schupp, P.J.; Kitamura, M.; Hatta, M.; Steinberg, P.D.; Harder, T. Chemical mediation of coral larval settlement by crustose coralline algae. Sci. Rep. 2015, 5, srep10803. [Google Scholar] [CrossRef]
- Pollock, F.J.; Katz, S.M.; van de Water, J.A.; Davies, S.W.; Hein, M.; Torda, G.; Matz, M.V.; Beltran, V.H.; Buerger, P.; Puill-Stephan, E.; et al. Coral larvae for restoration and research: A large-scale method for rearing Acropora millepora larvae, inducing settlement, and establishing symbiosis. PeerJ 2017, 5, e3732. [Google Scholar] [CrossRef]
- Humanes, A.; Noonan, S.H.C.; Willis, B.L.; Fabricius, K.E.; Negri, A.P. Cumulative effects of nutrient enrichment and elevated temperature compromise the early life history stages of the coral Acropora tenuis. PLoS ONE 2016, 11, e0161616. [Google Scholar] [CrossRef]
- Airi, V.; Gizzi, F.; Falini, G.; Levy, O.; Dubinsky, Z.; Goffredo, S. Reproductive efficiency of a Mediterranean endemic zooxanthellate coral decreases with increasing temperature along a wide latitudinal gradient. PLoS ONE 2014, 9, e91792. [Google Scholar] [CrossRef]
- Ricardo, G.F.; Jones, R.J.; Negri, A.P.; Stocker, R. That sinking feeling: Suspended sediments can prevent the ascent of coral egg bundles. Sci. Rep. 2016, 6, 21567. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.J.; Szmant, A.M. Elevated temperature affects development, survivorship, and settlement of the elkhorn coral, Acropora palmata (Lamarck 1816). Biol. Bull. 2009, 217, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Kuffner, I.B. Effects of ultraviolet (UV) radiation on larval settlement of the reef coral Pocillopora damicornis. Mar. Ecol. Prog. Ser. 2001, 217, 251–261. [Google Scholar] [CrossRef]
- Knutson, S.; Downs, C.A.; Richmond, R.H. Concentrations of Irgarol in selected marinas of Oahu, Hawaii and effects on settlement of coral larval. Ecotoxicology 2012, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Doropoulos, C.; Evensen, N.R.; Gómez-Lemos, L.A.; Babcock, R.C. Density-dependent coral recruitment displays divergent responses during distinct early life-history stages. R. Soc. Open Sci. 2017, 4, 170082. [Google Scholar] [CrossRef] [PubMed]
- Babcock, R.C.; Baird, A.H.; Piromvaragorn, S.; Thomson, D.P.; Willis, B.L. Identification of scleractinian coral recruits from Indo-Pacific reefs. Zool. Stud. 2002, 42, 211–226. [Google Scholar]
- Cameron, K.A.; Harrison, P.L. Density of coral larvae can influence settlement, post-settlement colony abundance and coral cover in larval restoration. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Jorissen, H.; Galand, P.E.; Bonnard, I.; Meiling, S.; Raviglione, D.; Meistertzheim, A.-L.; Hédouin, L.; Banaigs, B.; Payri, C.E.; Nugues, M.M. Coral larval settlement preferences linked to crustose coralline algae with distinct chemical and microbial signatures. Sci. Rep. 2021, 11, 14610. [Google Scholar] [CrossRef]
- Cruz, D.W.D.; Harrison, P.L. Enhancing coral recruitment through assisted mass settlement of cultured coral larvae. PLoS ONE 2020, 15, e0242847. [Google Scholar] [CrossRef]
- Suzuki, G.; Okada, W.; Yasutake, Y.; Yamamoto, H.; Tanita, I.; Yamashita, H.; Hayashibara, T.; Komatsu, T.; Kanyama, T.; Inoue, M.; et al. Enhancing coral larval supply and seedling production using a special bundle collection system “coral larval cradle” for large-scale coral restoration. Restor. Ecol. 2020, 28, 1172–1182. [Google Scholar] [CrossRef]
- Humanes, A.; Ricardo, G.F.; Willis, B.L.; Fabricius, K.E.; Negri, A.P. Cumulative effects of suspended sediments, organic nutrients and temperature stress on early life history stages of the coral Acropora tenuis. Sci. Rep. 2017, 7, srep44101. [Google Scholar] [CrossRef] [PubMed]
- Keshavmurthy, S.; Hsu, C.M.; Kuo, C.Y.; Denis, V.; Leung, J.K.L.; Fontana, S.; Hsieh, H.J.; Tsai, W.S.; Su, W.C.; Chen, C.A. Larval development of fertilized “pseudo-gynodioecious” eggs suggests a sexual pattern of gynodioecy in Galaxea fascicularis (Scleractinia: Euphyllidae). Zool. Stud. 2012, 51, 143–149. [Google Scholar]
- Cirino, L.; Wen, Z.-H.; Hsieh, K.; Huang, C.-L.; Leong, Q.L.; Wang, L.-H.; Chen, C.-S.; Daly, J.; Tsai, S.; Lin, C. First instance of settlement by cryopreserved coral larvae in symbiotic association with dinoflagellates. Sci. Rep. 2019, 9, 18851. [Google Scholar] [CrossRef] [PubMed]
- Perrin, J.; Vielzeuf, D.; Ricolleau, A.; Dallaporta, H.; Valton, S.; Floquet, N. Block-by-block and layer-by-layer growth modes in coral skeletons. Am. Miner. 2015, 100, 681–695. [Google Scholar] [CrossRef]
- Sun, C.Y.; Stifler, C.A.; Chopdekar, R.V.; Schmidt, C.A.; Parida, G.; Schoeppler, V.; Fordyce, B.I.; Brau, J.H.; Mass, T.; Tambutté, S.; et al. From particle attachment to space-filling coral skeletons. Proc. Natl. Acad. Sci. USA 2020, 117, 30159–30170. [Google Scholar] [CrossRef] [PubMed]
- Fabina, N.S.; Putnam, H.M.; Franklin, E.C.; Stat, M.; Gates, R.D. Transmission mode predicts specificity and interaction patterns in coral-Symbiodinium networks. PLoS ONE 2012, 7, e44970. [Google Scholar] [CrossRef]
- Levy, O.; Dubinsky, Z.; Achituv, Y. Photobehavior of stony corals: Responses to light spectra and intensity. J. Exp. Biol. 2003, 206, 4041–4049. [Google Scholar] [CrossRef]
- Rädecker, N.; Chen, J.E.; Pogoreutz, C.; Herrera, M.; Aranda, M.; Voolstra, C.R. Nutrient stress arrests tentacle growth in the coral model Aiptasia. Symbiosis 2019, 78, 61–64. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Galaxea fascicularis (n = 3000) | Mycedium elephantotus (n = 420) | Pocillopora verrucosa (n = 797) | Seriatopora caliendrum (n = 519) | |
|---|---|---|---|---|
| Settlement percentage (%) | 46 ± 14 | 28 ± 14 | 14 ± 16 | 100 ± 0 |
| Survival rate of 2 months Post-settlement (%) | 29 ± 23 | 10 ± 4 | 3 ± 1 | 15 ± 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.; Kang, C.-M.; Huang, C.-Y.; Li, H.-H.; Tsai, S. Study on the Development and Growth of Coral Larvae. Appl. Sci. 2022, 12, 5255. https://doi.org/10.3390/app12105255
Lin C, Kang C-M, Huang C-Y, Li H-H, Tsai S. Study on the Development and Growth of Coral Larvae. Applied Sciences. 2022; 12(10):5255. https://doi.org/10.3390/app12105255
Chicago/Turabian StyleLin, Chiahsin, Chia-Ming Kang, Chih-Yang Huang, Hsing-Hui Li, and Sujune Tsai. 2022. "Study on the Development and Growth of Coral Larvae" Applied Sciences 12, no. 10: 5255. https://doi.org/10.3390/app12105255