
Morphological and Molecular Alterations Induced by Lead in Embryos and Larvae of Danio rerio

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Substance
2.2. Zebrafish Husbandry and Maintenance
2.3. Exposure Conditions
2.4. Statistical Analyses
3. Results
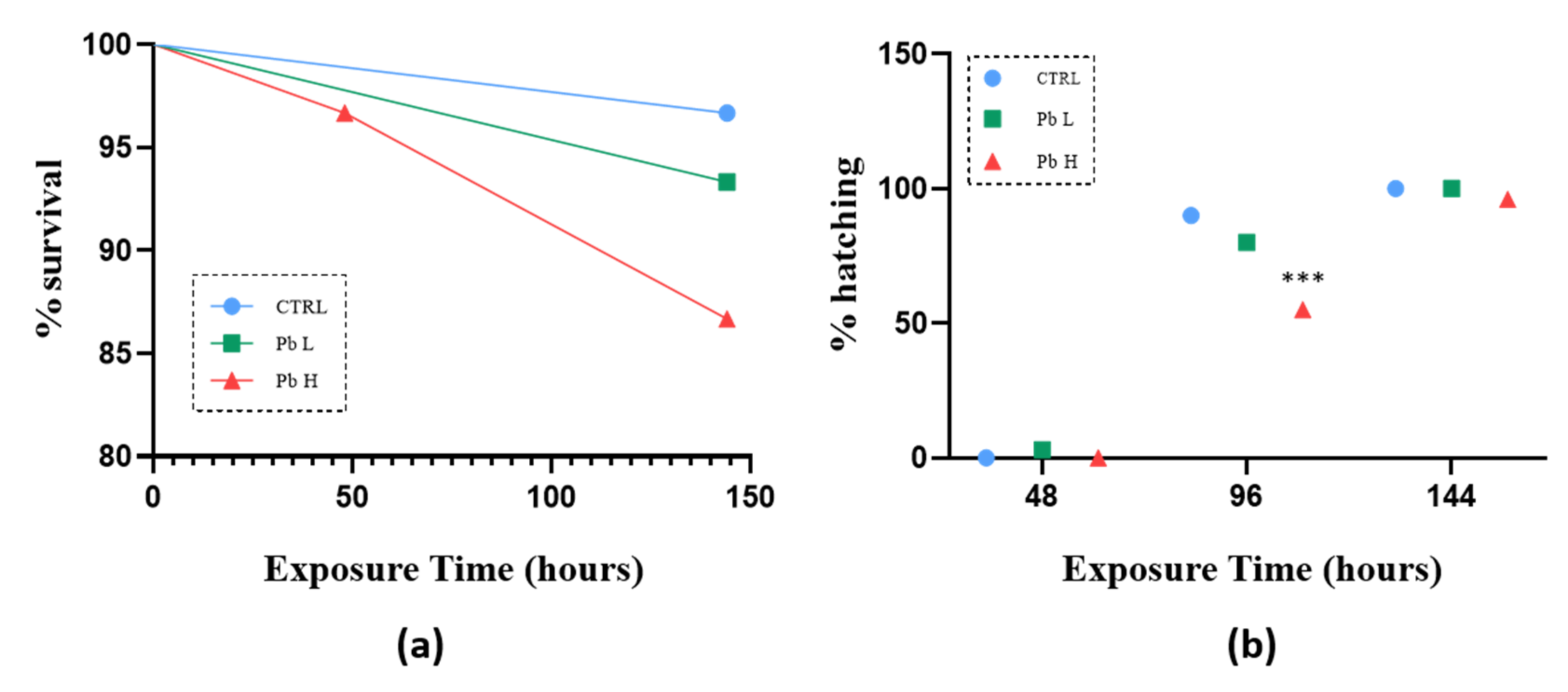
3.1. Survivor and Hatching Rate
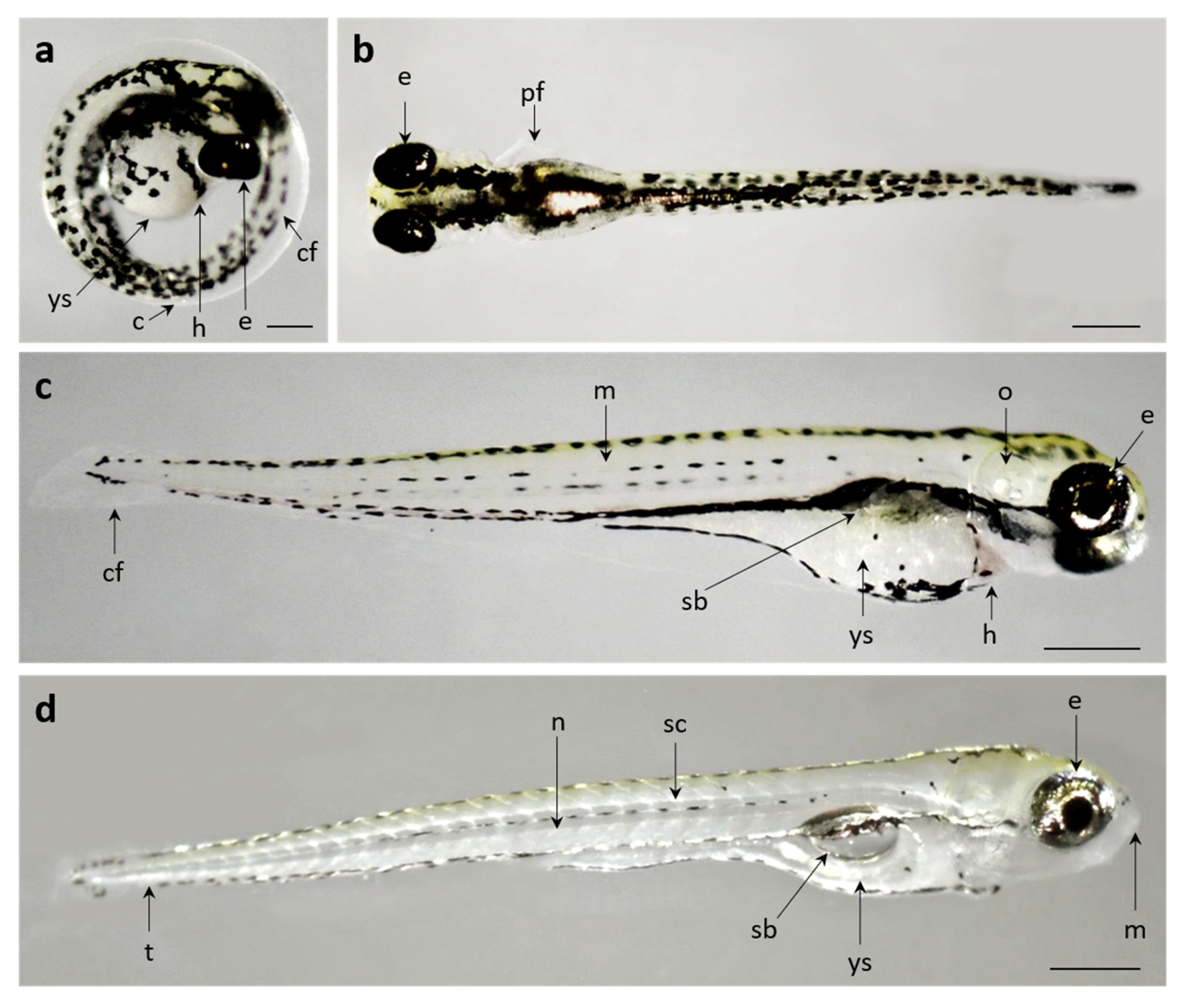
3.2. Developmental Toxicity
3.2.1. Control Group
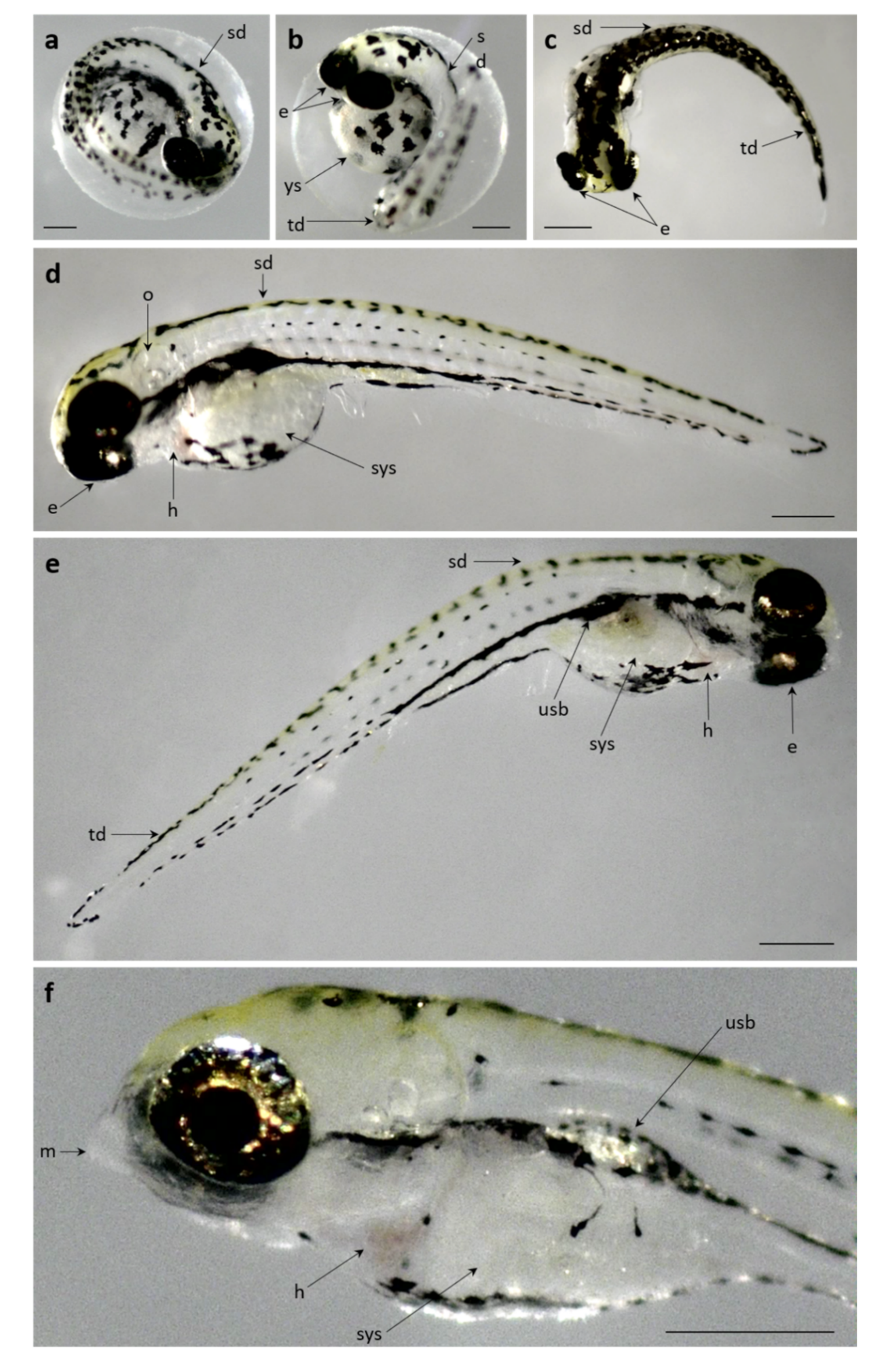
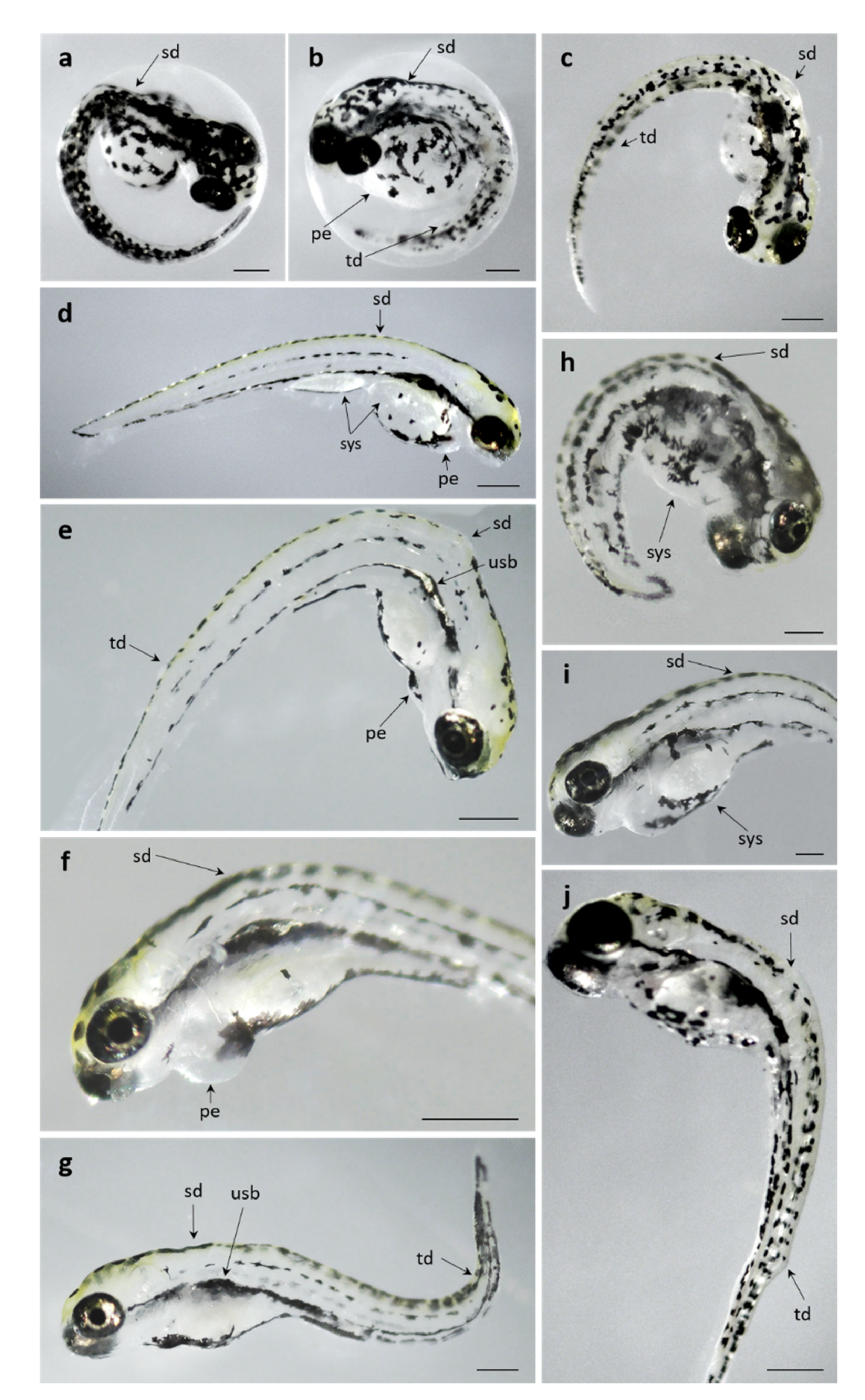
3.2.2. Pb Exposed Groups
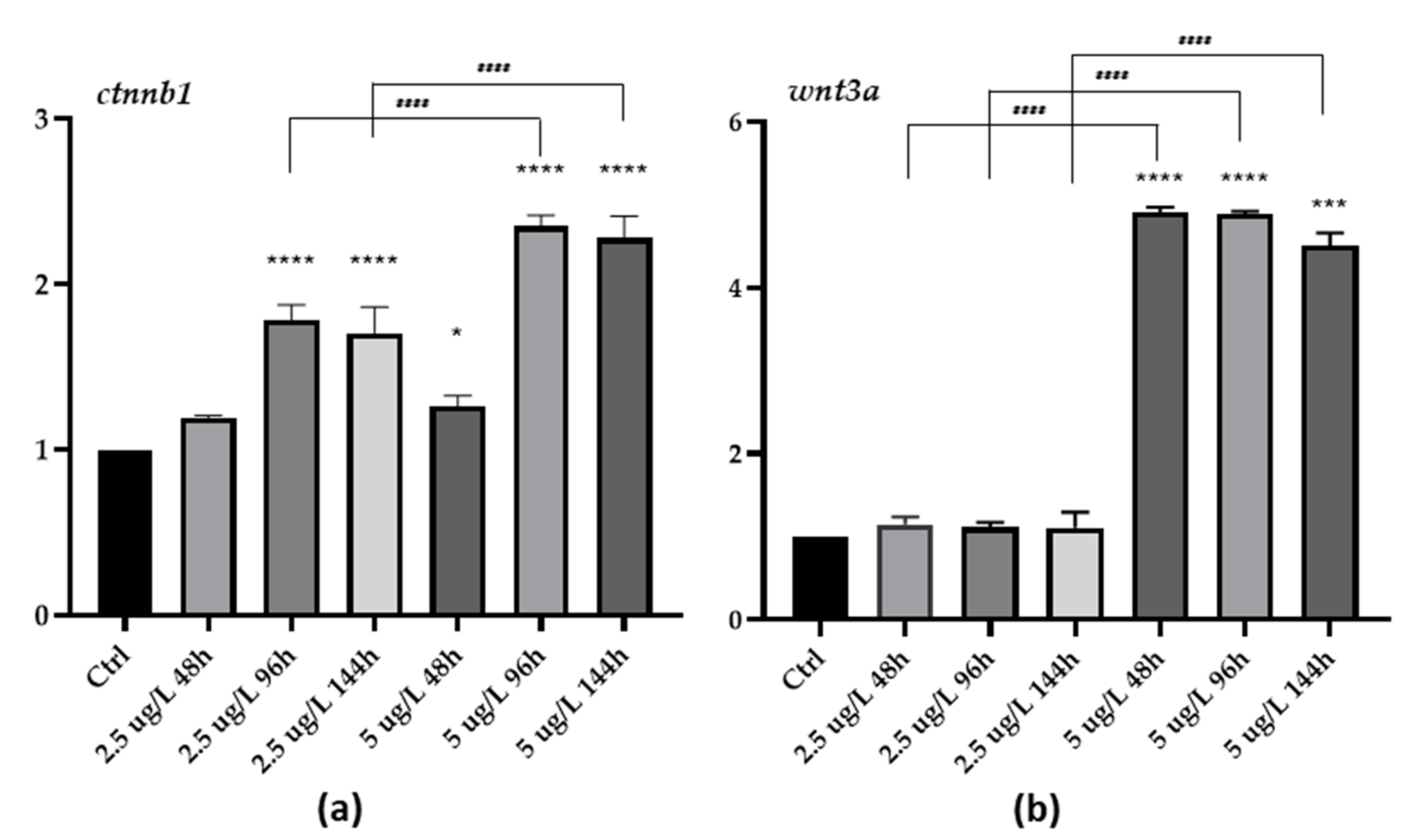
3.3. Gene Expression
4. Discussion
4.1. Morphological Modifications
4.2. Gene-Expression
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rahman, Z.; Singh, V.P. The relative impact of toxic heavy metals (THMs) (arsenic (As), cadmium (Cd), chromium (Cr)(VI), mercury (Hg), and lead (Pb) on the total environment: An overview. Environ. Monit. Assess. 2019, 191, 1–21. [Google Scholar] [CrossRef]
- Pratush, A.; Kumar, A.; Hu, Z. Adverse effect of heavy metals (As, Pb, Hg, and Cr) on health and their bioremediation strategies: A review. Int. Microbiol. 2018, 21, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Sall, M.L.; Diaw, A.K.D.; Gningue-Sall, D.; Efremova Aaron, S.; Aaron, J.J. Toxic heavy metals: Impact on the environment and human health, and treatment with conducting organic polymers, a review. Environ. Sci. Pollut. Res. 2020, 27, 29927–29942. [Google Scholar] [CrossRef]
- Su, C.; Lu, Y.; Johnson, A.C.; Shi, Y.; Zhang, M.; Zhang, Y.; Juergens, M.D.; Jin, X. Which metal represents the greatest risk to freshwater ecosystem in Bohai Region of China? Ecosyst. Health Sustain. 2017, 3, e01260. [Google Scholar] [CrossRef]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A review of toxicity and mechanisms of individual and mixtures of heavy metals in the environment. Environ. Sci. Pollut. Res. 2016, 23, 8244–8259. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.G.; Lin, Q.; Huang, H.H.; Wang, L.G.; Ning, J.J.; Du, F.Y. Heavy metals in fish tissues/stomach contents in four marine wild commercially valuable fish species from the western continental shelf of South China Sea. Mar. Pollut. Bull. 2017, 114, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Cabral-Pinto, M.M.S.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; Khan, S.A.; et al. Lead toxicity: Health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health 2020, 17, 2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Choi, H.; Hwang, U.K.; Kang, J.C.; Kang, Y.J.; Kim, K.I.; Kim, J.H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Phar. 2019, 68, 101–108. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B.; Li, N.; Ji, X.; Liu, K.; Jin, M. Zebrafish neurobehavioral phenomics applied as the behavioral warning methods for fingerprinting endocrine disrupting effect by lead exposure at environmentally relevant level. Chemosphere 2019, 231, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.K.; Parvin, E.; Islam, M.M.; Akter, M.S.; Khan, S.; Al-Mamun, M.H. Lead-and cadmium-induced histopathological changes in gill, kidney and liver tissue of freshwater climbing perch Anabas testudineus (Bloch, 1792). Chem. Ecol. 2014, 30, 532–540. [Google Scholar] [CrossRef]
- Ahmed, Q.; Khan, D.; Elahi, N. Concentrations of heavy metals (Fe, Mn, Zn, Cd, Pb, and Cu) in muscles, liver and gills of adult Sardinella Albella (Valenciennes, 1847) from Gwadar water of Balochistan, Pakistan. FUUAST J. Biol. 2014, 4, 195–204. [Google Scholar]
- Regulation, E.C. No 1907/2006 Of the European Parliament and of the Council of 18 December 2006 Concerning the Registration. Evaluation, Authorisation and Restriction of Chemicals (REACH), Establishing a European Chemicals Agency. Amend. Dir. 2006, 45, 1–849. [Google Scholar]
- Obeng-Gyasi, E. Sources of lead exposure in various countries. Rev. Environ. Health 2019, 34, 25–34. [Google Scholar] [CrossRef]
- Giri, S.S.; Yun, S.; Jun, J.W.; Kim, H.J.; Kim, S.G.; Kang, J.W.; Kim, S.J.; Han, S.J.; Sukumaran, V.; Park, S.C. Therapeutic effect of intestinal autochthonous Lactobacillus reuteri P16 against waterborne lead toxicity in Cyprinus carpio. Front. Immunol. 2018, 9, 1824. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.M.; Ali, M.L.; Islam, M.S.; Rahman, M.Z. Preliminary assessment of heavy metals in water and sediment of Karnaphuli River, Bangladesh. Environ. Nanotechnol. Monit. Manage. 2016, 5, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Bhuyan, M.S.; Bakar, M.A.; Rashed-Un-Nabi, M.; Senapathi, V.; Chung, S.Y.; Islam, M.S. Monitoring and assessment of heavy metal contamination in surface water and sediment of the Old Brahmaputra River, Bangladesh. Appl. Water Sci. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, P.; Andreji, J.; Dvořáková-Líšková, Z.; Vejsada, P. Assessment of selected heavy metals pollution in water, sediments and fish in the basin Dyje, Czech Republic. Neuroendocrinol. Lett. 2014, 35, 101–109. [Google Scholar]
- Kim, H.J.; Mahboob, S.; Viayaraghavan, P.; Al-Ghanim, K.A.; Al-Misned, F.; Kim, Y.O.; Ahmed, Z. Determination of toxic effects of lead acetate on different sizes of zebrafish (Danio rerio) in soft and hard water. J. King Saud Univ. Sci. 2020, 32, 1390–1394. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. The lead accumulation and hematological findings in juvenile rock fish Sebastes schlegelii exposed to the dietary lead (II) concentrations. Ecotox. Environ. Safe. 2015, 115, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Aldavood, S.J.; Abbott, L.C.; Evans, Z.R.; Griffin, D.J.; Lee, M.D.; Quintero-Arevalo, N.M.; Villalobos, A.R. Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) embryos. Water 2020, 12, 3005. [Google Scholar] [CrossRef]
- Osman, A.G.; Wuertz, S.; Mekkawy, I.A.; Exner, H.J.; Kirschbaum, F. Lead induced malformations in embryos of the African catfish Clarias gariepinus (Burchell, 1822). Environ. Toxicol. 2007, 22, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Viganò, L.; Casatta, N.; Farkas, A.; Mascolo, G.; Roscioli, C.; Stefani, F.; Vitelli, M.; Olivo, F.; Clerici, L.; Robles, P.; et al. Embryo/larval toxicity and transcriptional effects in zebrafish (Danio rerio) exposed to endocrine active riverbed sediments. Environ. Sci. Pollut. Res. 2020, 27, 10729–10747. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, J.C. Effects of sub-chronic exposure to lead (Pb) and ascorbic acid in juvenile rockfish: Antioxidant responses, MT gene expression, and neurotransmitters. Chemosphere 2017, 171, 520–527. [Google Scholar] [CrossRef]
- Lee, J.; Freeman, J.L. Zebrafish as a model for investigating developmental lead (Pb) neurotoxicity as a risk factor in adult neurodegenerative disease: A mini-review. Neurotoxicology 2014, 43, 57–64. [Google Scholar] [CrossRef]
- Xia, J.; Lu, L.; Jin, C.; Wang, S.; Zhou, J.; Ni, Y.; Fu, Z.; Jin, Y. Effects of short term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish. Comp. Biochem. Physiol. C: Toxicol. Pharmacol. 2018, 209, 1–8. [Google Scholar] [CrossRef]
- Shampa, J.; Batham, D. Acute toxicity and biochemical studies of lead nitrate on the liver and kidney of fresh water fish Mystus cavasius. J. Glob. Biosci. 2016, 5, 4590–4597. [Google Scholar]
- Zhang, H.; Liu, Y.; Liu, R.; Liu, C.; Chen, Y. Molecular mechanism of lead-induced superoxide dismutase inactivation in zebrafish livers. J. Phys. Chem. B 2014, 118, 14820–14826. [Google Scholar] [CrossRef]
- Ishaque, A.; Ishaque, S.; Arif, A.; Abbas, H.G. Toxic effects of lead on fish and human. Biol. Clin. Sci. Res. J. 2020, 2020, e045. [Google Scholar] [CrossRef]
- Dai, J.; Zhang, L.; Du, X.; Zhang, P.; Li, W.; Guo, X.; Li, Y. Effect of lead on antioxidant ability and immune responses of Crucian carp. Biol. Trace Elem. Res. 2018, 186, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.; Chakraborty, S.; Sengupta, M. Lead toxicity on non-specific immune mechanisms of freshwater fish Channa punctatus. Aquat. Toxicol. 2014, 152, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.B.R.; Nagae, M.Y.; Zaia, C.T.B.V.; Zaia, D.A.M. Acute morphological and physiological effects of lead in the neotropical fish Prochilodus lineatus. Braz. J. Biol. 2004, 64, 797–807. [Google Scholar] [CrossRef]
- Yin, J.; Wang, A.P.; Li, W.F.; Shi, R.; Jin, H.T.; Wei, J.F. Time-response characteristic and potential biomarker identification of heavy metal induced toxicity in zebrafish. Fish. Shellfish Immun. 2018, 72, 309–317. [Google Scholar] [CrossRef]
- Khodake, S.P.; Borale, R.P. Lead nitrate induced histochemical alteration in the liver of freshwater fish Mystus bleekeri (Day). Int. J. Life Sci. 2018, 6, 131–136. [Google Scholar]
- Macirella, R.; Sesti, S.; Bernabò, I.; Tripepi, M.; Godbert, N.; Brunelli, E. Lead toxicity in seawater teleosts: A morphofunctional and ultrastructural study on the gills of the Ornate wrasse (Thalassoma pavo L.). Aquat. Toxicol. 2019, 211, 193–201. [Google Scholar] [CrossRef]
- Paul, S.; Mandal, A.; Bhattacharjee, P.; Chakraborty, S.; Paul, R.; Mukhopadhyay, B.K. Evaluation of water quality and toxicity after exposure of lead nitrate in fresh water fish, major source of water pollution. Egypt. J. Aquat. Res. 2019, 45, 345–351. [Google Scholar] [CrossRef]
- Mokhtar, D.M.; Abd-Elhafeez, H.H. Histological changes in selected organs of Oreochromis niloticus exposed to doses of lead acetate. J. Life Sci. Biomed. 2013, 3, 256–263. [Google Scholar]
- Devi, R.; Banerjee, T.K. Toxicopathological impact of sub-lethal concentration of lead nitrate on the aerial respiratory organs of “murrel” Channa striata (Bloch, Pisces). Iran. J. Environ. Health Sci. Eng. 2007, 4, 249–256. [Google Scholar]
- Bambino, K.; Chu, J. Zebrafish in toxicology and environmental health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar] [CrossRef] [Green Version]
- Shankar, P.; Dashner-Titus, E.J.; Truong, L.; Hayward, K.; Hudson, L.G.; Tanguay, R.L. Developmental toxicity in zebrafish (Danio rerio) exposed to uranium: A comparison with lead, cadmium, and iron. Environ. Pollut. 2021, 269, 116097. [Google Scholar] [CrossRef]
- Dai, Y.J.; Jia, Y.F.; Chen, N.; Bian, W.P.; Li, Q.K.; Ma, Y.B.; Chen, Y.L.; Pei, D.S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.; Glaberman, S. The zebrafish (Danio rerio) model in toxicity testing. In An. Introduction to Interdisciplinary Toxicology; from Molecules to Man, 1st ed.; Pope, C.N., Liu, J., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 525–532. [Google Scholar] [CrossRef]
- Xu, X.; Weber, D.; Burge, R.; VanAmberg, K. Neurobehavioral impairments produced by developmental lead exposure persisted for generations in zebrafish (Danio rerio). Neurotoxicology 2016, 52, 176–185. [Google Scholar] [CrossRef]
- Sanchez, O.F.; Lee, J.; Yu King Hing, N.; Kim, S.E.; Freeman, J.L.; Yuan, C. Lead (Pb) exposure reduces global DNA methylation level by non-competitive inhibition and alteration of dnmt expression. Metallomics 2017, 9, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Fan, C.; Chen, X.; Liu, J.; Wang, B.; Huang, Z.; Zhang, Y.; Meng, X.; Zou, F. Effects of cadmium, manganese, and lead on locomotor activity and neurexin 2a expression in zebrafish. Environ. Toxicol. Chem. 2017, 36, 2147–2154. [Google Scholar] [CrossRef]
- Jin, M.; Ji, X.; Zhang, B.; Sheng, W.; Wang, R.; Liu, K. Synergistic effects of Pb and repeated heat pulse on developmental neurotoxicity in zebrafish. Ecotox. Environ. Safe. 2019, 172, 460–470. [Google Scholar] [CrossRef]
- Kataba, A.; Botha, T.L.; Nakayama, S.M.; Yohannes, Y.B.; Ikenaka, Y.; Wepener, V.; Ishizuka, M. Acute exposure to environmentally relevant lead levels induces oxidative stress and neurobehavioral alterations in larval zebrafish (Danio rerio). Aquat. Toxicol. 2020, 227, 105607. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.N.; Crofts, E.J.; Akemann, C.; Gurdziel, K.; Farr, R.; Baker, B.B.; Weber, D.; Baker, T.R. Developmental exposure to Pb2+ induces transgenerational changes to zebrafish brain transcriptome. Chemosphere 2020, 244, 125527. [Google Scholar] [CrossRef]
- Roy, N.M.; DeWolf, S.; Carneiro, B. Evaluation of the developmental toxicity of lead in the Danio rerio body. Aquat. Toxicol. 2015, 158, 138–148. [Google Scholar] [CrossRef]
- Tendulkar, R.; Chaudhari, K.; Fernandes, A.; Ratnaparkhi, P. An Evaluation of lead (Pb) toxicity in developing zebrafish (Danio rerio) embryos. Int. J. 2015, 3, 498–504. [Google Scholar]
- Yin, J.; Wang, A.P.; Li, W.F.; Shi, R.; Jin, H.T.; Wei, J.F. Sensitive biomarkers identification for differentiating Cd and Pb induced toxicity on zebrafish embryos. Environ. Toxicol. Pharm. 2017, 56, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Jezierska, B.; Ługowska, K.; Witeska, M. The effects of heavy metals on embryonic development of fish (a review). Fish. Physiol. Biochem. 2009, 35, 625–640. [Google Scholar] [CrossRef]
- Park, K.; Han, E.J.; Ahn, G.; Kwak, I.S. Effects of thermal stress-induced lead (Pb) toxicity on apoptotic cell death, inflammatory response, oxidative defense, and DNA methylation in zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2020, 224, 105479. [Google Scholar] [CrossRef]
- Rice, C.; Ghorai, J.K.; Zalewski, K.; Weber, D.N. Developmental lead exposure causes startle response deficits in zebrafish. Aquat. Toxicol. 2011, 105, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Sfakianakis, D.G.; Renieri, E.; Kentouri, M.; Tsatsakis, A.M. Effect of heavy metals on fish larvae deformities: A review. Environ. Res. 2015, 137, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Macirella, R.; Guardia, A.; Pellegrino, D.; Bernabò, I.; Tronci, V.; Ebbesson, L.O.; Sesti, S.; Tripepi, S.; Brunelli, E. Effects of two sublethal concentrations of mercury chloride on the morphology and metallothionein activity in the liver of Zebrafish (Danio rerio). Int. J. Mol. Sci. 2016, 17, 361. [Google Scholar] [CrossRef] [Green Version]
- Macirella, R.; Brunelli, E. Morphofunctional alterations in zebrafish (Danio rerio) gills after exposure to mercury chloride. Int. J. Mol. Sci. 2017, 18, 824. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, E.; Mauceri, A.; Maisano, M.; Bernabò, I.; Giannetto, A.; De Domenico, E.; Corapi, B.; Tripepi, S.; Fasulo, S. Ultrastructural and immunohistochemical investigation on the gills of the teleost, Thalassoma pavo L., exposed to cadmium. Acta Histochem. 2011, 113, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Doering, J.A.; Sun, J.; Beitel, S.C.; Shekh, K.; Patterson, S.; Crawford, S.; Giesy, J.P.; Wiseman, S.B.; Hecker, M. Linking oxidative stress and magnitude of compensatory responses with life-stage specific differences in sensitivity of white sturgeon (Acipenser transmontanus) to copper or cadmium. Environ. Sci. Technol. 2016, 50, 9717–9726. [Google Scholar] [CrossRef]
- Monserrat, J.M.; Martínez, P.E.; Geracitano, L.A.; Amado, L.L.; Martins, C.M.G.; Pinho, G.L.L.; Chaves, I.S.; Ferreira-Cravo, M.; Ventura-Lima, J.; Bianchini, A. Pollution biomarkers in estuarine animals: Critical review and new perspectives. Comp. Biochem. Physiol. C 2007, 146, 221–234. [Google Scholar] [CrossRef]
- Ahmad, I.; Hamid, T.; Fatima, M.; Chand, H.S.; Jain, S.K.; Athar, M.; Raisuddin, S. Induction of hepatic antioxidants in freshwater catfish (Channa punctatus Bloch) is a biomarker of paper mill effluent exposure. BBA-Gen. Subj. 2000, 1523, 37–48. [Google Scholar] [CrossRef]
- Viarengo, A.; Burlando, B.; Cavaletto, M.; Marchi, B.; Ponzano, E.; Blasco, J. Role of metallothionein against oxidative stress in the mussel Mytilus galloprovincialis. Am. J. Physiol. Regul. Integr. 1999, 277, R1612–R1619. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dynam. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Herrmann, K. Teratogenic effects of retinoic acid and related substances on the early development of the zebrafish (Brachydanio rerio) as assessed by a novel scoring system. Toxicol. In Vitro 1995, 9, 267–283. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Von Hellfeld, R.; Brotzmann, K.; Baumann, L.; Strecker, R.; Braunbeck, T. Adverse effects in the fish embryo acute toxicity (FET) test: A catalogue of unspecific morphological changes versus more specific effects in zebrafish (Danio rerio) embryos. Environ. Sci. Eur. 2020, 32, 1–18. [Google Scholar] [CrossRef]
- Osman, A.G.; Wuertz, S.; Mohammed-Geba, K. Lead-induced heat shock protein (HSP70) and metallothionein (MT) gene expression in the embryos of African catfish Clarias gariepinus (Burchell, 1822). Sci. Afr. 2019, 3, e00056. [Google Scholar] [CrossRef]
- Lukaszewicz-Hussain, A. Role of oxidative stress in organophosphate insecticide toxicity—Short review. Pestic. Biochem. Phys. 2010, 98, 145–150. [Google Scholar] [CrossRef]
- Slaninova, A.; Smutna, M.; Modra, H.; Svobodova, Z. A review: Oxidative stress in fish induced by pesticides. Neuroendocrinol. Lett. 2009, 30, 2. [Google Scholar]
- Yang, M.; Chen, J.; Zhao, J.; Meng, M. Etanercept attenuates myocardial ischemia/reperfusion injury by decreasing inflammation and oxidative stress. PLoS ONE 2014, 9, e108024. [Google Scholar] [CrossRef] [Green Version]
- Maharajan, K.; Muthulakshmi, S.; Nataraj, B.; Ramesh, M.; Kadirvelu, K. Toxicity assessment of pyriproxyfen in vertebrate model zebrafish embryos (Danio rerio): A multi biomarker study. Aquat. Toxicol. 2018, 196, 132–145. [Google Scholar] [CrossRef]
- Wu, S.; Ji, G.; Liu, J.; Zhang, S.; Gong, Y.; Shi, L. TBBPA induces developmental toxicity, oxidative stress, and apoptosis in embryos and zebrafish larvae (Danio rerio). Environ. Toxicol. 2016, 31, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Quintaneiro, C.; Patrício, D.; Novais, S.C.; Soares, A.M.V.M.; Monteiro, M.S. Endocrine and physiological effects of linuron and S-metolachlor in zebrafish developing embryos. Sci. Total Environ. 2017, 586, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.G.; Mekkawy, I.A.; Verreth, J.; Kirschbaum, F. Effects of lead nitrate on the activity of metabolic enzymes during early developmental stages of the African catfish, Clarias gariepinus (Burchell, 1822). Fish. Physiol. Biochem. 2007, 33, 1–13. [Google Scholar] [CrossRef]
- Almeida, J.A.; Novelli, E.L.B.; Silva, M.D.P.; Júnior, R.A. Environmental cadmium exposure and metabolic responses of the Nile tilapia, Oreochromis niloticus. Environ. Pollut. 2001, 114, 169–175. [Google Scholar] [CrossRef]
- Almeida, J.A.; Diniz, Y.S.; Marques, S.F.G.; Faine, L.A.; Ribas, B.O.; Burneiko, R.C.; Novelli, E.L.B. The use of the oxidative stress responses as biomarkers in Nile tilapia (Oreochromis niloticus) exposed to in vivo cadmium contamination. Environ. Int. 2002, 27, 673–679. [Google Scholar] [CrossRef]
- Das, P.C.; Ayyappan, S.; Das, B.K.; Jena, J.K. Nitrite toxicity in Indian major carps: Sublethal effect on selected enzymes in fingerlings of Catla catla, Labeo rohita and Cirrhinus mrigala. Comp. Biochem. Phys. C 2004, 138, 3–10. [Google Scholar] [CrossRef]
- Oliveira, R.; Domingues, I.; Grisolia, C.K.; Soares, A.M. Effects of triclosan on zebrafish early-life stages and adults. Environ. Sci. Pollut. Res. 2009, 16, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Q. Dose-and time-related changes in aerobic metabolism, chorionic disruption, and oxidative stress in embryonic medaka (Oryzias latipes): Underlying mechanisms for silver nanoparticle developmental toxicity. Aquat. Toxicol. 2012, 124, 238–246. [Google Scholar] [CrossRef]
- Dar, O.I.; Sharma, S.; Singh, K.; Sharma, A.; Bhardwaj, R.; Kaur, A. Biomarkers for the toxicity of sublethal concentrations of triclosan to the early life stages of carps. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of β-catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [Green Version]
- Gumbiner, B.M.; McCrea, P.D. Catenins as mediators of the cytoplasmic functions of cadherins. J. Cell Sci. 1993, 1993, 155–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurberg, A.D.; Aires, R.; Nóvoa, A.; Rowland, J.E.; Mallo, M. Compartment-dependent activities of Wnt3a/β-catenin signaling during vertebrate axial extension. Dev. Biol. 2014, 394, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Hao, H.; Reynolds, K.; McMahon, M.; Zhou, C.J. Wnt signaling in neural crest ontogenesis and oncogenesis. Cells 2019, 8, 1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polakis, P. The many ways of Wnt in cancer. Curr. Opin. Genet. Dev. 2007, 17, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lucero, O.M.; Dawson, D.W.; Moon, R.T.; Chien, A.J. A re-evaluation of the oncogenic nature of Wnt/beta-catenin signaling inmelanoma and other cancers. Curr. Oncol. Rep. 2010, 12, 314–318. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.R.; Fagotto, F. The ins and outs of APC and betacatenin nuclear transport. EMBO Rep. 2002, 3, 834–839. [Google Scholar] [CrossRef]
- Städeli, R.; Hoffmans, R.; Basler, K. Transcription under the control of nuclear Arm/β-catenin. Curr. Biol. 2006, 16, R378–R385. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deformity rate (%) | CTRL | 2.5 µg/L | 5 µg/L | |||||||
| 48 h | 96 h | 144 h | 48 h | 96 h | 144 h | 48 h | 96 h | 144 h | ||
| Spinal deformity | 0 | 0 | 0 | 30 (a **) | 50 (a ****) | 60 (a ****) | 90 (ab ****) | 90 (a ****)(b **) | 100 (a ****)(b **) | |
| Tail deformity | 0 | 0 | 0 | 20 (a *) | 50 (a ****) | 50 (a ****) | 90 (ab ****) | 90 (a ****)(b **) | 100 (ab ****) | |
| Yolk sac swellig | 0 | 0 | 0 | 0 | 30 (a **) | 60 (a ****) | 0 | 40 (a ***) | 70 (a ****) | |
| Pericardial edema | 0 | 0 | 0 | 0 | 0 | 0 | 20 (ab *) | 50 (ab ****) | 60 (ab ****) | |
| Uninflated swim bladder | 0 | 0 | 0 | 0 | 0 | 60(a ****) | 0 | 0 | 80 (a ****) | |
| Cumulative deformity | 0 | 0 | 0 | 60 (a ****) | 90 (a ****) | 90 (a ****) | 90 (a ****)(b *) | 90 (a ****) | 100 (a ****) | |
| Score value | 0 | 0 | 0 | 12 (a *) | 35 (a ****) | 58 (a ****) | 58 (ab ****) | 74 (ab ****) | 144 (ab ****) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curcio, V.; Macirella, R.; Sesti, S.; Pellegrino, D.; Ahmed, A.I.M.; Brunelli, E. Morphological and Molecular Alterations Induced by Lead in Embryos and Larvae of Danio rerio. Appl. Sci. 2021, 11, 7464. https://doi.org/10.3390/app11167464
Curcio V, Macirella R, Sesti S, Pellegrino D, Ahmed AIM, Brunelli E. Morphological and Molecular Alterations Induced by Lead in Embryos and Larvae of Danio rerio. Applied Sciences. 2021; 11(16):7464. https://doi.org/10.3390/app11167464
Chicago/Turabian StyleCurcio, Vittoria, Rachele Macirella, Settimio Sesti, Daniela Pellegrino, Abdalmoiz I. M. Ahmed, and Elvira Brunelli. 2021. "Morphological and Molecular Alterations Induced by Lead in Embryos and Larvae of Danio rerio" Applied Sciences 11, no. 16: 7464. https://doi.org/10.3390/app11167464