
Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors


Department of Life Sciences, University of Modena and Reggio Emilia, Via G. Campi 103/287, 41125 Modena, Italy
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(16), 7177; https://doi.org/10.3390/app11167177
Submission received: 14 July 2021
/
Revised: 28 July 2021
/
Accepted: 30 July 2021
/
Published: 4 August 2021
(This article belongs to the Special Issue State-of-Art of Microbial Concerns in Food Safety)
Abstract
:The aim of this study was to evaluate and characterize the bacterial load present in twenty-four Ready-To-Eat (RTE) sandwiches, purchased at refrigerated vending machines and supermarkets in the province of Modena (Italy). We isolated 54 bacterial strains, including pathogens of interest in food safety, such as Listeria, Staphylococcus, Enterococcus, Yersinia, Aeromonas and Acinetobacter spp. Phenotypic tests have been performed on these pathogens to detect the presence of virulence factors, such as gelatinase production and hemolytic capability. To test their antibiotic resistance features, the minimum inhibitory concentration (MIC) against eight commonly used antibiotics (Amikacin, Ciprofloxacin, Ampicillin, Oxacillin, Imipenem, Tetracycline, Erythromycin and Vancomycin) was also evaluated. The results showed that among the 54 isolates, fifty percent (50%) belonged to harmless microorganisms (Leuconostoc and Lactococcus), whereas the remaining fifty percent (50%) included pathogenic bacteria (Listeria ivanovii, Listeria monocytogenes, Staphylococcus aureus, Yersinia, and Citrobacter spp.), species responsible for pathologies often difficult to treat due to the presence of antibiotic resistance features. This study demonstrates the importance of thorough controls, both during the production and marketing of RTE food like sandwiches, to avoid reaching the infectious load and the onset of pathologies, particularly dangerous for old and immunocompromised patients.
1. Introduction
Ready-to-eat food (RTE) is a type of product that must not be cooked or reheated before serving. This includes salads, vegetables, fruits, cooked meats, smoked fish, desserts, sandwiches, cheese and food that are previously cooked in order to be later served cold. The trend towards consumption of minimally processed, ready-to-eat, refrigerated and frozen food products poses new problems to the identification and management of bacterial risks for consumers, mainly due to the inability to maintain a temperature-controlled environment. Even if properly stored, RTE foods can contain high risk ingredients that allow pathogens to grow and multiply. Recent modifications in food production and processing practices and the ever-changing eating habits of the consumers, who appreciate the readiness of RTE foods, have affected the incidence of pathogens that can multiply in such foods. This typology of foods presents more safety and quality complications, because of their susceptibility to bacterial contamination, for the absence of the adequate preservatives and storage temperature during the food chain. These products are usually characterized by a long shelf life at refrigerated temperature and they can be contaminated with spoilage bacteria and foodborne pathogens during all stages of the production process. Like many types of RTE food, sandwiches are tasty and easy to eat, especially for workers who are always in a hurry and eating meals away from home. Sandwiches are a popular RTE food typology that can contain some high-risk ingredients, such as raw vegetables, eggs and salad dressings, and whose preparation usually involves manual handling, which increases the number of potential contamination sources. Many pathogenic bacteria like Listeria, Staphylococcus, Shigella, Yersinia spp., ubiquitous bacteria found in many environments like soil or water, can come into contact with these raw or fresh ingredients [1]. Many human pathogens can also be transmitted through bare-hand handlings, like S. aureus, which can be found on human skin and hair, or other pathogens colonizing the intestinal tract. The multi-ingredients used for the production and the sandwich fillings like cheese, chicken, eggs, ham, salad, sauce, and various types of fresh produce contribute to the microbial contamination and have been associated with foodborne illness outbreaks, the last reported in the UK in 2019, where hospitalized patients died after the consumption of sandwiches contaminated by Listeria monocytogenes [2]. The RTE foods preservation techniques are based on low temperatures, which do not prevent some psychrotrophic bacteria from multiplying at refrigeration temperatures, such as L. monocytogenes, a ubiquitous Gram-positive genus that can be isolated from a wide range of food products [3]. This psychrotrophic pathogen may be already present in raw materials, but the risks associated with the consumption of RTE foods [4] could also be due to their presence in processing plant equipment and cross-contamination during the food storage at refrigeration temperature [5]. L. monocytogenes is the most important specie responsible for food-borne outbreaks and severe illness, listeriosis, mainly in elderly consumers, immunocompromised individuals and pregnant women [6], even if other species like L. ivanovii and L. seeligeri have been found in sporadic cases. L. ivanovii has been isolated, although rarely, from infected humans, indicating a pathogenic potential for humans [7]. Although, L. seeligeri carries a virulence gene cluster that is not expressed, it has been associated with rare cases of infections in humans [8,9].
The objective of the present investigation was to examine the microbiological quality of 24 ready-to-eat sandwiches, to evaluate the risk for consumers due to the presence of pathogenic strains, and the presence in the isolates of antibiotic resistance and other virulence factors.
2. Materials and Methods
2.1. Sampling
Twenty-four sandwiches in modified atmosphere packaging have been purchased on different days from automatic distributors and in supermarkets in the province of Modena, processed within 4 h of the collection and stored at 4 °C for the duration of the experiment. The sandwiches contained different ingredient combinations: Tuna and tomato, ham and cheese, tomato and mozzarella cheese, tuna and eggs, turkey and vegetables, shrimp and pink sauce, cooked ham, raw ham, smoked cheese and tomatoes, cooked ham and mushrooms, tuna and onions, cooked ham and artichokes, raw ham and eggplants. The ingredients and the expiry date were recorded, to ensure that the samples were within their shelf-life period at the end of the study.
2.2. Microbiological Analysis
Sandwiches samples (25 g) were homogenized using a Stomacher (Lab Blender, Seward, London, UK) for 1 min in sterile plastic bags, with 225 mL buffered peptone water added (Oxoid, Milan, Italy). The homogenate samples and the appropriate dilutions were spread on selective and non-selective plates and incubated at 37 °C for 24–48 h. Tryptic Soy Agar (TSA) was used as a non-selective medium for the total bacterial count, while as selective plates the following media were used: MacConkey Agar (MK) for the isolation of Enterobacteriaceae, Listeria Palcam Agar Base (PAL), for the isolation of species belonging to the genus Listeria, Mannitol Salt Agar (MSA), for the isolation of Staphylococcus spp., Kanamycin Aesculin Azide Agar Base (KA) for Enterococcus spp. (all media from Oxoid, Milan, Italy). After incubation, colonies grown on selective plates were individually cultured to proceed with the identification and biological characterization. All bacterial counts were recorded as CFU/g.
2.3. Identification of Isolates
The identification of the isolates was obtained using the EnteroPluri-Test (Liofilchem, Italy) to identify microorganisms belonging to the Enterobacteriaceae family and the API® strips (Biomerieux, Marcy l’Etoile, France) for the other genera. Subsequently, the bacterial identification was definitively confirmed by matrix-assisted laser desorption ionization (MALDI) time-of-flight mass spectrometry (TOF/MS). All the strains were stored in phosphate-buffered saline (PBS; 8 g NaCl, 0.2 g KCl, 2.9 g Na2HPO4·12H2O, 0.2 g KH2PO4 with 1L of distilled water) supplemented with 30% (vol/vol) glycerine at −80 °C.
2.4. Evaluation of Virulence Factors: Hemolysin and Gelatinase Production
Hemolysin and gelatinase production were evaluated in all 54 isolates by spotting the plates added with the specific media with 10 μL of suspension, cultured in Tryptic Soy Broth (Oxoid, SpA, Milan, Italy) at 30 °C for 48 h. For the hemolysin production, all the strains were cultured on blood agar plates containing 5% of defibrinated horse blood (bioMérieux, Florence, Italy). After incubation at 37 °C for 24 h, the haemolytic activity was determined by observing a clear zone of haemolysis (β-haemolysis), a partial and greening haemolysis zone (α-haemolysis) or no activity (γ-haemolysis) around the spots. Gelatinase production was assessed by inoculation of the strains in a Nutrient Broth containing 10% gelatin. Positive gelatinase was recorded as degradation of the gelatin to liquid. Given that some microorganisms produce low amounts of gelatinase, all negative results were further incubated for up to 15 days to observe any delayed positive reactions.
2.5. Determination of the Minimum Inhibitory Concentration (MIC)
The MICs of 30 out of 54 isolated and identified strains have only been evaluated, chosen on the basis of their ability to cause food poisoning. Following the guidelines of the EUCAST (European Committee on Antimicrobial Susceptibility Testing), the MICs were obtained for eight antibiotics: Amikacin, Ciprofloxacin, Ampicillin, Oxacillin, Imipenem, Tetracycline, Erythromycin and Vancomycin (Table 1). The evaluation was carried out using the agar dilution method by adding double dilutions of antibiotics to the Mueller-Hinton agar medium (Oxoid, SpA, Milan, Italy). The results were then compared with the breakpoints from EUCAST 2021 [10].
3. Results
3.1. Quantification and Identification of the Isolates
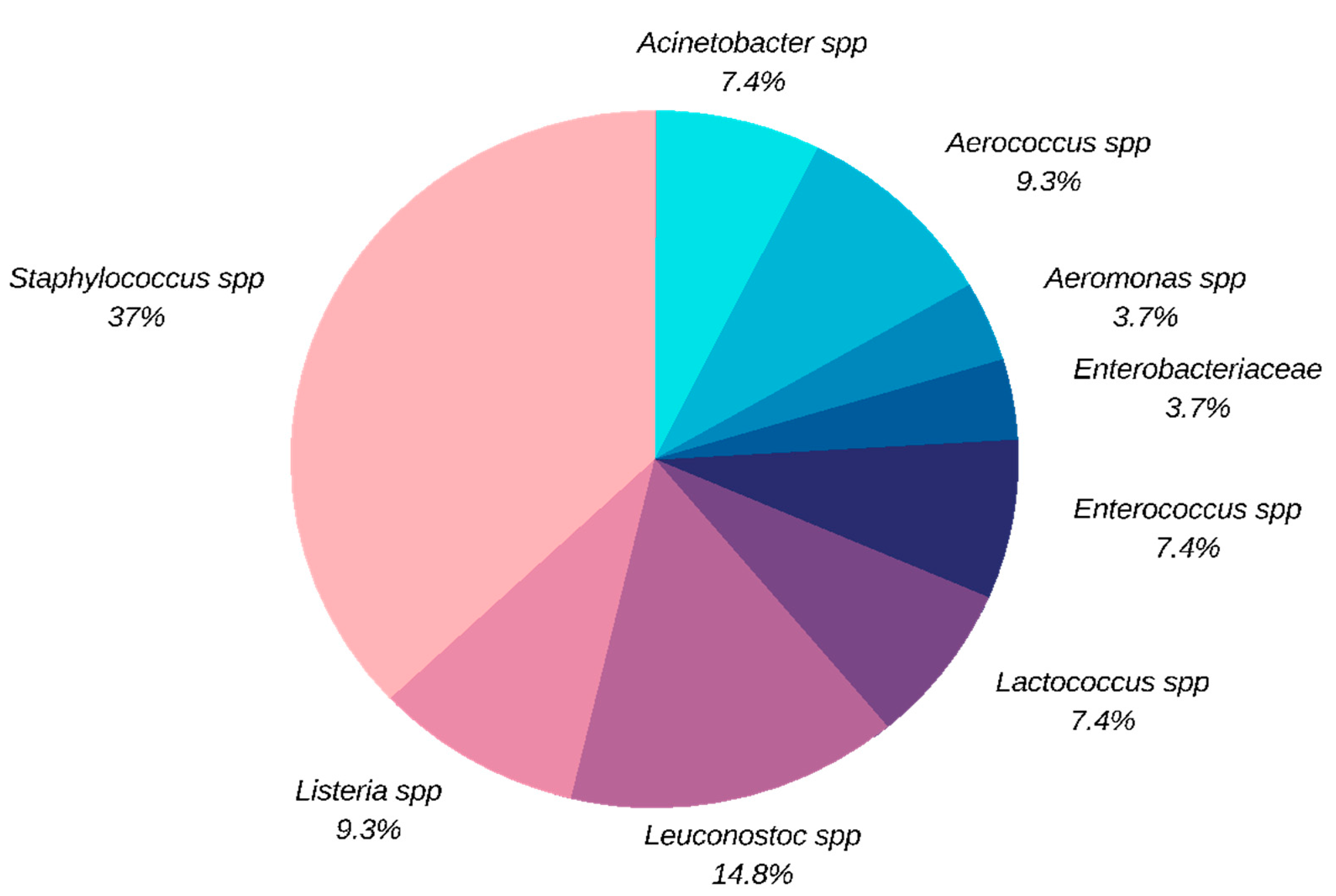
A total bacterial count from 104 CFU/g to 106 CFU/g was found on the non-selective TSA medium for the most processed sandwiches produced with the greatest amount of ingredients and the most manipulated ones (meats, sauces and vegetables of different types). Fifty-four strains were isolated from both samples and ingredients on selective media MSA, KA, PAL, MK with bacterial load ranging from 102 to 105 CFU/g, 102 to 104 CFU/g, 103 to 104 CFU/g, and 102 to 103 CFU/g, respectively. Of the 54 strains examined, twenty (37.04%) belonged to the genera Staphylococcus, eight (14.81%) to Leuconostoc, five to (9.26%) Listeria, five (9.26%) to Aerococcus, four (7.41%) to Lactococcus, four (7.41%) to Enterococcus, four (7.41%) to Acinetobacter, two (3.70%) to Aeromonas, one (1.85%) to Citrobacter, one (1.85%) to Yersinia (the last two referred together in the figure as Enterobacteriaceae) (Figure 1).
Staphylococcus is the most represented genus, with bacterial load ranging from 102 to 105 CFU/g. Notably, S. aureus was found on 14 samples and in 8 of these it was the only contaminant. Regarding to the other Gram-positive strains, Enterococcus was the second isolated genus, found in three samples, and represented by E. faecium (2 strains), E. avium (1 strain) and E. durans (1 strain), followed by five Aerococcus viridans isolates. Leuconostoc and Lactococcus were also found, both genera used as commercial starters in the bakery sectors. Among the Gram-negative strains, in three samples bacterial loads between 103 and 104 CFU/g were found on MacConkey Agar (MK). Aeromonas hydrophila was isolated from one sample, and it was also present in another one together with Citrobacter spp., whereas in a third sample the simultaneous presence of Yersinia enterocolitica and Acinetobacter lwoffii was observed. Last, four Listeria ivanovii subsp. londoniensis and one Listeria welshimeri were isolated from three sandwiches, with bacterial loads around 102 CFU/g for all isolates.
3.2. Determination of the Minimum Inhibitory Concentration
As reported in Table 1, all E. faecium strains were resistant to Vancomycin (8 µg/mL), while only one of these presented a resistance to Ampicillin and Erythromycin (16 µg/mL). E. avium and E. durans, following EUCAST guidelines, were tested only against Ampicillin and Vancomycin, to which they were sensitive (Table 2).
All 13 S. aureus strains were sensitive to Amikacina. Only S. aureus S29 and S41 were sensitive to Ciprofloxacin, while the other 11 strains showed a “Susceptible, increased exposure” (MIC value at the breakpoint). Only one isolate (S. aureus S26) is endowed with a multi-resistance to Erythromycin (2 µg/mL) and Oxacillin (4 µg/mL). It is important to note that most isolates presented a “Susceptible, increased exposure” profile (Table 3).
In relation to the psychrotrophic bacteria, the five Listeria isolates (Table 4) were resistant to Erythromycin. In relation to Tetracycline, all L. ivanovii londoniensis showed a “Susceptible, increased exposure” profile, while L. welshimeri was resistant to the compound, with MIC value of 4 µg/mL. Four strains (3 strains of L. ivanovii spp. londoniensis with a value of 2 µg/mL and L. welshimeri with value of 4 µg/mL) was resistant to Ampicillin. The two strains of A. hydrophila were evaluated only against Imipenem to which they were resistant, with values of 2 µg/mL. Whereas Y. enterocolitica showed a “Susceptible, increased exposure” profile for Ciprofloxacin and Tetracycline, with value of 0.25 µg/mL, and 4 µg/mL, respectively (Table 5).
Lastly, all the Acinetobacter species are sensitive to both the antibiotic Amikacin and Ciprofloxacin, and the only isolate belonging to the genus Citrobacter tested against Amikacin showed a resistance profile with a value of 16 µg/mL (Table 6).
3.3. Phenotypical Virulence Factors
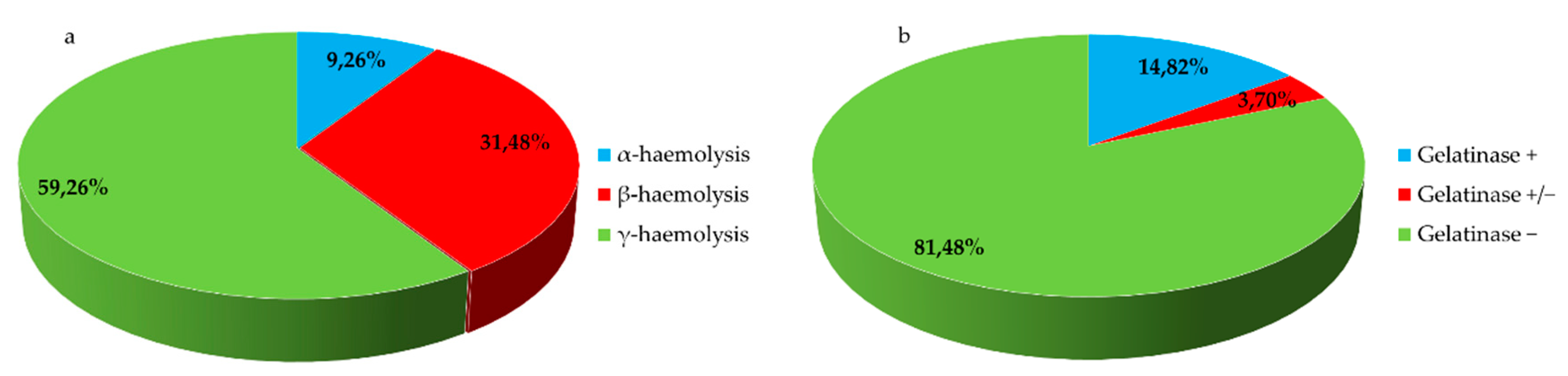
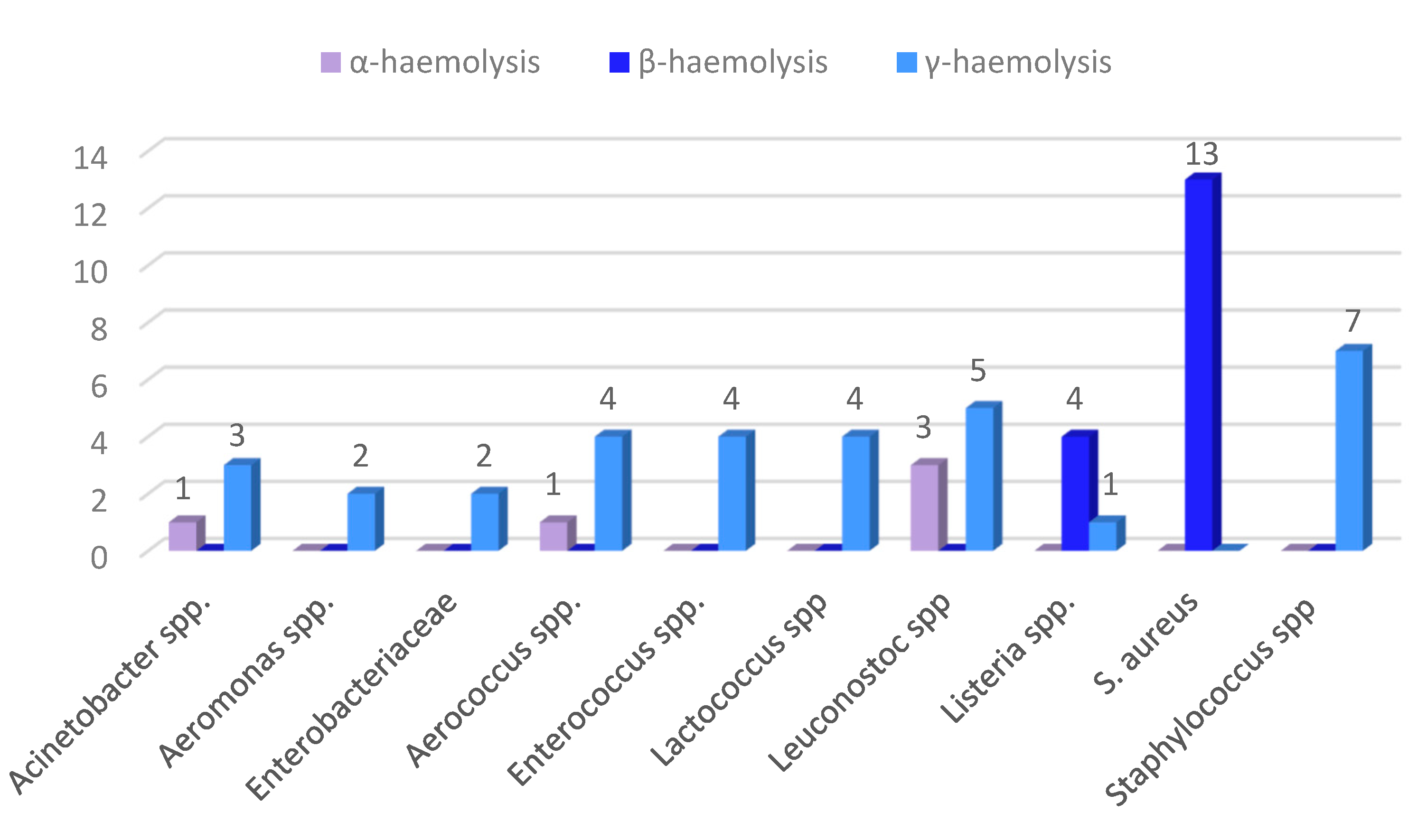
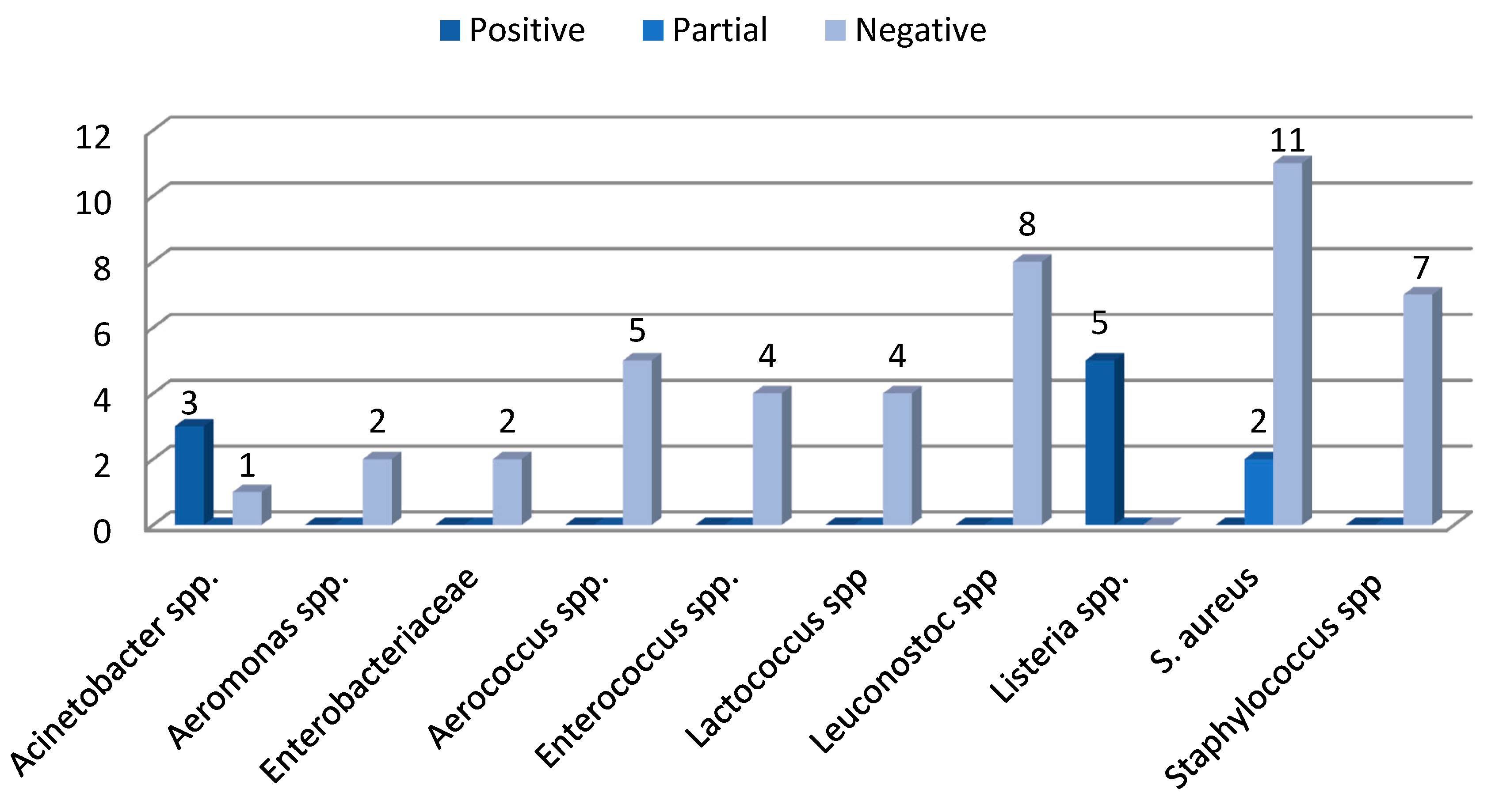
The results of the virulence tests (Figure 2a,b) showed that hemolytic and gelatinase activities were present in 40,74% and 18,52% of the isolates, respectively. Five strains (9.26%) showed α-hemolytic activity, seventeen strains (31.48%) β-hemolytic activity and in thirty-two strains (59.26%) this feature was not present. All S. aureus and 4 out 5 Listeria spp. (L. ivanovii) are endowed with β-hemolytic virulence factor (Figure 3). All Listeria and 3 out 4 Acinetobacter spp. could hydrolyze the gelatin, keeping the culture medium liquid even at temperatures below the solidification point. Two strains (Staphylococcus aureus 28S and Staphylococcus aureus 37S) showed a partial gelatinase capacity, corresponding to an incomplete solidification of the media at low temperatures (Figure 4).
4. Discussion
Microbiological contamination is distributed throughout the food processing chain, from its production to consumption. The risks are greater in foods, which do not require further heating before consumption, such as ready to eat (RTE) products. Moreover, low hygiene practices during handling, cross- contamination and poor refrigerated temperature control may be responsible for exposure to pathogens bacteria through the food chain. The European Food Safety Authority believes that up to 90% of listeriosis cases occurring from 2008 to 2015 are due to the consumption of RTE foods, such as smoked fish, preserved meat and cheeses, and also vegetables, such as ready to eat salads. A third of the cases was due to the proliferation of Listeria spp., Staphylococcus spp., Shigella, Y. enterocolitica and other pathogenic bacteria found in foods incorrectly stored in the refrigerated vending machine [25,26,27].
The present study showed that among the 54 isolates, 25 strains (46.3%) belonged to harmless microorganisms, including Leuconostoc and Lactococcus, lactobacilli used as starters in fermentation processes of cheese or bread, whereas the remaining 27 strains (53.7%) belonged to pathogenic bacteria (13 S. aureus, 5 Listeria spp., 4 Enterococcus spp., 4 Acinetobacter spp., one Y. enterocolitica, one A. hydrophila, and one Citrobacter spp.). The pathogens present in some samples were frequently found with a microbial load exceeding the limits of the EC Regulation 2073/2005 [28], as in the case of the eight samples contaminated with S. aureus only. In other studies S. aureus was found in RTE products, especially in cold served salads and sandwiches [29,30], tracing its presence in ingredients, such as vegetables and/or fruit and products of animal origin (meat, sausages) [31]. S. aureus, by the production of a thermostable toxin, is the species most frequently responsible for food poisoning (the third pathogen for food-borne diseases). S. aureus poisoning is a pathology of a modest clinical entity but has a strong socio-economic impact as it is highly widespread. In the present investigation some samples were contaminated by two or more pathogenic bacteria at the same time. Sample 3, a product characterized by many components and therefore subjected to multiple manipulations, showed the presence not only of S. aureus, but also of L. ivanovii and Citrobacter spp. In sample 6, a shrimp-based product, both Y. enterocolitica and L. ivanovii were isolated. L. ivanovii was recently associated with sporadic human infections, although it is usually linked to animal infections, in particular ruminants, and this microorganism is known as a milk and cheese contaminant [32]. Citrobacter species, belonging to the Enterobacteriaceae family, are considered opportunistic pathogens for humans, but for their wide distribution in the environment and intestinal tracts, they have the capability to be transferred from the farm to fresh produce destined for consumption, thereby constituting a public health risk [33]. Yersiniosis is the third most commonly reported zoonosis in Europe, mainly for the consumption of pig meat and products thereof [25]. In some European countries Y. enterocolitica, the dominating species among human diseases is a more significant concern than Salmonella because it can grow at refrigeration temperature [34]. As a psychrotrophic microorganism, a significant health threat is posed by refrigerated products where this pathogen, as well as Listeria and Aeromonas, can proliferate. A. hydrophila is a microorganism found in water, considered the most important infectious reservoir [35], and consequently, it is mainly isolated from seafood and vegetables. It rarely causes food outbreaks, the most recent reported in Sweden and Norway [36]. Acinetobacter is rarely associated with diarrheal disease but A. baumannii, specie found in four sandwiches, possess pathogenetic features, including antibiotic resistance, and could represent a risk for humans, linked to the consumption of contaminated foods (dairy products, raw fruit and vegetables) [37]. Enterococcus spp. have been recognized as emerging human pathogens in recent years, always endowed with antibiotic resistance and other virulence factors [38,39]. They seem to also be involved in the production of biogenic amines in some fermented foods [40]. The intoxication, caused by the ingestion of these compounds, can cause the onset of various symptoms, such as headache, vomiting up to the manifestation of severe allergic reactions.
In relation to the virulence traits, this study has highlighted that all the L. ivanovii and three out of four A. baumannii were endowed with hemolytic and gelatinase activity. Lastly, 13 isolates showed a profile of antibiotic resistance. The half presented a multiple resistance profile among the antibiotic-resistant strains, in particular, three out of four Listeria and the only two resistant Enterococcus strains. Most of the MICs evaluated (19 out 30, 63.3%) showed a “Susceptible, increased exposure” profile (MIC value at the breakpoint); in particular, all S. aureus, L. ivanovii and Y. enterocolitica strains. The isolation of antibiotics resistant and of “Susceptible, increased exposure” strains is of concern not only for the consequent therapeutic use difficulties, but also because this characteristic can be transferred by conjugation to different species within the food microbial community [41,42]. Antibiotic resistance is encoded by genes usually located on mobile genetic elements that may transfer to human pathogens in retail food microflora. Conditions with a “Susceptible, increased exposure” could allow the coexistence in the microbial community of both very resistant strains that grow poorly in the absence of antibiotics and less susceptible strains that grow well in the absence of antibiotics [43].
5. Conclusions
The variety of pathogens isolated in the present study is a source of health concern, as the consumption of RTE foods, has continuously increased, consistent with the consumers’ ever-changing eating habits.
For this type of product, the preservation techniques are based primarily on the use of low temperatures, which do not prevent the growth of some psychrotrophic bacteria, and given this RTE food is eaten without cooking, there is also no control of mesophilic pathogens. Therefore, there is a need to increase the level of attention and awareness, following good hygiene practices throughout the food chain (from farm to fork), notably by producers, to prevent pathogens contamination during preparation, handling and storage. Among the preventive activities able to significantly reduce the health risk is the addition of natural antimicrobial compounds (essential oils, plant extracts, or bacteriocins) to reduce the amounts of preservatives added in RTE products [44,45]. Natural antimicrobials show an antioxidant and antimicrobial activity against a wide range of food pathogens and spoilage bacteria, in order to extend RTE sandwiches shelf-life without altering the sensory properties [46]. The increasing importance attributed to the quality and safety of food has prompted the European Union in 2004 to draw up a series of regulations, which entered into force in Italy in 2006, known as the “Hygiene Package”. The EC regulation 852/2004 [47] is responsible for ensuring the hygiene and safety of food products “from farm to table”, thus, from the production area (farm) up to consumers (table). It sets both general and specific hygiene requirements to be applied to personnel, means of transport and places where food is processed and sold. It also requires the implementation of the HACCP (Hazard Analysis and Critical Control Points) system, applying the principle of self-control which incentives food business operators to adopt various measures to monitor the healthiness of the finished product. In conclusion, our results show that sandwiches could be a health risk, in particular for susceptible individuals, as they can be a vehicle for the transmission of food-borne pathogens. The quantification and identification of bacterial strains isolated from the various sandwiches show how products with more components, and consequently more manipulated are the most contaminated ones. In particular, the isolation of S. aureus from a lot of samples leads us to believe that food handling is the most likely cause of bacterial contamination, followed by contamination caused by an inaccurate washing of the vegetables present in the food [29], as evidenced by the presence of other species like Listeria, Enterococcus, Enterobacteriaceae and bacteria belonging to the genera Acinetobacter, Aerococcus and Aeromonas. These results underline, once again, the need for better hygienic conditions during food processing, together with the choice of quality raw products. Given the presence of possible hygienic deficiencies in the production chain of some products examined, careful controls are of fundamental importance, both during the food processing and marketing, to avoid the onset of infections difficult to treat, especially in weaker subjects.
Author Contributions
Conceptualization, P.M. and R.I.; methodology R.I.; software, R.I. and C.C.; investigation, S.C. and R.I.; data curation, S.C. and C.C.; writing—original draft preparation, S.C.; writing—review and editing, P.M and R.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Callejón, R.M.; Rodríguez-Naranjo, M.I.; Ubeda, C.; Hornedo-Ortega, R.; Garcia-Parrilla, M.C.; Troncoso, A.M. Reported foodborne outbreaks due to fresh produce in the United States and European Union: Trends and causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef]
- Listeriosis in England and Wales: Summary for 2019. Available online: https://www.gov.uk/government/publications/listeria-monocytogenes-surveillance-reports/listeriosis-in-england-and-wales-summary-for-2019 (accessed on 13 July 2021).
- Guerra, M.M.; Mclauchlin, J.; Bernardo, F.A. Listeria in ready-to-eat and unprocessed foods produced in Portugal. Food Microbiol. 2001, 18, 423–429. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez,, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Scientific Opinion on the Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Tompkin, R.B. Control of Listeria monocytogenes in the food-processing environment. J Food Prot. 2002, 65, 709–725. [Google Scholar] [CrossRef]
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillet, C.; Join-Lambert, O.; Le Monnier, A.; Leclercq, A.; Mechaï, F.; Mamzer-Bruneel, M.F.; Bielecka, M.K.; Scortti, M.; Disson, O.; Berche, P.; et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis. 2010, 16, 136–138. [Google Scholar] [CrossRef]
- Hofer, E.; Ribeiro, R.; Feitosa, D.P. Species and serovars of the genus Listeria isolated from different sources in Brazil from 1971 to 1997. Mem. Inst. Oswaldo Cruz. 2000, 95, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Snapir, Y.M.; Vaisbein, E.; Nassar, F. Low virulence but potentially fatal outcome-Listeria ivanovii. Eur. J. Intern. Med. 2006, 17, 286–287. [Google Scholar] [CrossRef] [PubMed]
- EUCAST 2021. European Committee on Antimicrobial Susceptibility Testing European Committee on Antimicrobial Susceptibility Testing (EUCAST) (2021) Clinical-Breakpoints; Version 11.0; Växjö, Sweden. Wiley: Hoboken, NJ, USA, 2021; Available online: https://eucast.org/clinical_breakpoints (accessed on 13 July 2021).
- Davis, B.D. Mechanism of bactericidal action of aminoglycosides. Microbiol. Rev. 1987, 51, 341–350. [Google Scholar] [CrossRef]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.T. Wirkmechanismus der Chinolone [Mechanism of action of quinolones]. Infection 1986, 14, S3–S15. [Google Scholar] [CrossRef]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of quinolone action and resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef]
- Wilkowske, C.J. The penicillins. Mayo Clin. Proc. 1977, 52, 616–624. [Google Scholar] [PubMed]
- Peechakara, B.V.; Basit, H.; Gupta, M. Ampicillin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Frère, J.M. Mechanism of action of beta-lactam antibiotics at the molecular level. Biochem. Pharmacol. 1977, 26, 2203–2210. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Laskin, A.I.; Last, J.A. Tetracyclines. Antibiot. Chemother. 1971, 17, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Shutter, M.C.; Akhondi, H. Tetracycline. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Tanaka, S.; Otaka, T.; Kaji, A. Further studies on the mechanism of erythromycin action. Biochim. Biophys. Acta 1973, 331, 128–140. [Google Scholar] [CrossRef]
- Vázquez-Laslop, N.; Mankin, A.S. How Macrolide Antibiotics Work. Trends Biochem. Sci. 2018, 43, 668–684. [Google Scholar] [CrossRef]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved glycopeptide antibacterial drugs: Mechanism of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Preuss, C.V.; Bernice, F. Vancomycin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- European Food Safety Authority, European Centre for Disease Prevention and Control. The European Union Summary report on trends and sources of zoonoses, zoonotic agents and foodborne outbreaks in 2013. EFSA J. 2015, 13, 3991. [Google Scholar]
- El-Shenawy, M.; El-Shenawy, M.; Mañes, J.; Soriano, J.M. Listeria spp. in street-vended Ready-to-Eat foods. Interdiscip. Perspect. Infect. Dis. 2011, 2011, 968031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossu, F.; Spanu, C.; Deidda, S.; Mura, E.; Casti, D.; Pala, C.; Lamon, S.; Spanu, V.; Ibba, M.; Marrocu, E.; et al. Listeria Spp. and Listeria monocytogenes contamination in Ready-To-Eat sandwiches collected from vending machines. Ital. J. Food Saf. 2016, 5, 5500. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. 2005, 338, 1–26. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32005R2073 (accessed on 13 July 2021).
- Jang, H.G.; Kim, N.H.; Choi, Y.M.; Rhee, M.S. Microbiological quality and risk factors related to sandwiches served in bakeries, cafe’s and sandwich bars in South Korea. J. Food Protect. 2013, 76, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Soares, K.; Moura, A.T.; García-Díez, J.; Oliveira, I.; Esteves, A.; Saraiva, C. Evaluation of hygienic quality of food served in Universities canteens of northem Portugal. Indian J. Microbiol. 2020, 60, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Fijałkowski, K.; Peitler, D.; Karakulska, J. Staphylococci isolated from ready-to-eat meat—Identification, antibiotic resistance and toxin gene profile. Int. J. Food Microbiol. 2016, 238, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Ordóñez, A.; Leong, D.; Morgan, C.A.; Hill, C.; Gahan, C.G.; Jordan, K. Occurrence, persistence, and virulence potential of Listeria ivanovii in foods and food processing environments in the Republic of Ireland. Biomed. Res. Int. 2015, 2015, 350526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwu, C.D.; Okoh, A.I. Preharvest transmission routes of fresh produce associated bacterial pathogens with outbreak potentials: A Review. Int. J. Environ. Res. Public Health 2019, 16, 4407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guern, A.S.; Martin, L.; Savin, C.; Carniel, E. Yersiniosis in France: Overview and potential sources of infection. Int. J. Infect. Dis. 2016, 46, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. The significance of mesophilic Aeromonas spp. in minimally processed Ready-to-Eat seafood. Microorganisms 2019, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, A.M.; Nascimento, J.D. Acinetobacter: An underrated foodborne pathogen? J. Infect. Dev. Ctries. 2017, 11, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selleck, E.M.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Bondi, M.; Laukova, A.; de Niederhausern, S.; Messi, P.; Papadopoulou, C.; Economou, V. Controversial aspects displayed by Enterococci: Probiotics or pathogens? Biomed. Res. Int. 2020, 2020, 9816185. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Enterococci from foods. FEMS Microbiol. Rev. 2002, 26, 163–171. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public. Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Lin, Y.T.; Wan, T.W.; Wang, D.Y.; Lin, H.Y.; Lin, C.Y.; Chen, Y.C.; Teng, L.J. Distribution of antibiotic resistance genes among Staphylococcus species isolated from ready-to-eat foods. J. Food Drug. Anal. 2019, 27, 841–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.; Savage, V.M.; Yeh, P.J. Intermediate levels of antibiotics may increase diversity of colony size phenotype in bacteria. Comput. Struct. Biotechnol. J. 2018, 16, 307–315. [Google Scholar] [CrossRef]
- Devlieghere, F.; Vermeiren, L.; Debevere, J. New preservation technologies: Possibilities and limitations. Int. Dairy J. 2004, 14, 273–285. [Google Scholar] [CrossRef]
- Antonio, C.M.; Abriouel, H.; López, R.L.; Omar, N.B.; Valdivia, E.; Gálvez, A. Enhanced bactericidal activity of enterocin AS-48 in combination with essential oils, natural bioactive compounds and chemical preservatives against Listeria monocytogenes in ready-to-eat salad. Food Chem. Toxicol. 2009, 47, 2216–2223. [Google Scholar] [CrossRef]
- Ben-Fadhel, Y.; Maherani, B.; Aragones, M.; Lacroix, M. Antimicrobial properties of encapsulated antimicrobial natural plant products for Ready-to-Eat carrots. Foods. 2019, 8, 535. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Regulation on the hygiene of foodstuffs, 852/2004/EC. Off. J. L 2004, 139, 54. [Google Scholar]
Figure 1.
Bacterial species detected in the selective plates.

Figure 2.
(a,b) Haemolysis and gelatinase test results for all tested strains.

Figure 3.
Haemolysis test results.

Figure 4.
Gelatinase test results.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mechanism of action of antimicrobial agents used for MICs determination.
| Antimicrobial Agents | Group | Mechanism of Action | References |
|---|---|---|---|
| Amikacin | Aminoglycoside | Inhibits bacterial protein synthesis and also affects the barrier function of bacterial cell membranes. | [11,12] |
| Ciprofloxacin | Quinolone | Inhibit bacterial DNA gyrase and interfere with bacterial DNA replication. | [13,14] |
| Ampicillin | Penicillin (aminopenicillin) | Inhibition of bacterial cell wall synthesis by binding one or more of the penicillin binding proteins (PBPs). | [15,16] |
| Oxacillin | β-Lactam | Inhibits the synthesis of bacterial cell walls. | [17,18] |
| Imipenem | β-Lactam | Inhibits the synthesis of bacterial cell walls. | [17,18] |
| Tetracycline | Tetracycline | It specifically binds to the bacterial ribosomal 30S subunit at the A position, prevents the connection of aminoacyl-tRNA at this position, and inhibits protein synthesis. | [19,20] |
| Erythromycin | Macrolide | Combines with bacterial ribosomal 50S subunit to inhibit peptide acyltransferase and bacterial protein synthesis. | [21,22] |
| Vancomycin | Glycopeptide | Inhibits the synthesis of bacterial cell walls. | [23,24] |
Table 2.
MIC values for Enterococci strains and their Break Point values.
| Strains | Ciprofloxacin | Tetracycline | Ampicillin | Erythromycin | Vancomycin |
|---|---|---|---|---|---|
| E. faecium EC1 | 4 µg/mL | 1 µg/mL | 16 µg/mL | 16 µg/mL | 8 µg/mL |
| E. faecium EC2 | 4 µg/mL | 1 µg/mL | 4 µg/mL | 4 µg/mL | 8 µg/mL |
| E. avium EC16 | / | / | 1 µg/mL | / | 2 µg/mL |
| E. durans EC17 | / | / | 2 µg/mL | / | 2 µg/mL |
| BREAKPOINT | ≥8 µg/mL | ≥4 µg/mL | ≥8 µg/mL | ≥4 µg/mL | ≥4 µg/mL |
Table 3.
MIC values for Staphylococcus aureus strains and their Break Point values.
| Strains | Amikacin | Ciprofloxacin | Erythromicin | Vancomiycin | Oxacillin |
|---|---|---|---|---|---|
| S. aureus S26 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 1 µg/mL | 4 µg/mL |
| S. aureus S28 | 1 µg/mL | 1 µg/mL | 1 µg/mL | 1 µg/mL | 2 µg/mL |
| S. aureus S29 | 1 µg/mL | 0.25 µg/mL | 0.5 µg/mL | 0.5 µg/mL | 2 µg/mL |
| S. aureus S31 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S32 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 4 µg/mL |
| S. aureus S33 | 1 µg/mL | 1 µg/mL | 2 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S34 | 1 µg/mL | 1 µg/mL | 1 µg/mL | 2µg/mL | 2 µg/mL |
| S. aureus S35 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S36 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 2 µg/mL |
| S. aureus S37 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S41 | 1 µg/mL | 0.25 µg/mL | 0.5 µg/mL | 2 µg/mL | 1 µg/mL |
| S. aureus S42 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 1 µg/mL | 1 µg/mL |
| S. aureus S43 | 1 µg/mL | 1 µg/mL | 0.5 µg/mL | 2 µg/mL | 2 µg/mL |
| BREAKPOINT | ≥16 µg/mL | ≥1 µg/mL | ≥1 µg/mL | ≥2 µg/mL | ≥2 µg/mL |
Table 4.
MIC values for Listeria strains and Break Point values.
| Strains | Tetracycline | Ampicillin | Erythromycin |
|---|---|---|---|
| L. ivanovii spp. londoniensis 46L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 47L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 48L | 1 µg/mL | 2 µg/mL | 2 µg/mL |
| L. ivanovii spp. londoniensis 49L | 1 µg/mL | 0.5 µg/mL | 2 µg/mL |
| L. welshimeri 50L | 4 µg/mL | 4 µg/mL | 2 µg/mL |
| BREAKPOINT | ≥1 µg/mL | ≥1 µg/mL | ≥1 µg/mL |
Table 5.
MIC values for A. hydrophila and Y. enterocolitica strains and Break Point values.
| Strain | Imipenem | Ciprofloxacin | Tetracyclin |
|---|---|---|---|
| A. hydrophila 52EB | 2 µg/mL | ||
| A. hydrophila 59EB | 2 µg/mL | ||
| BREAKPOINT | ≥1 µg/mL | ||
| Y. enterocolitica 58EB | 0.25 µg/mL | 4 µg/mL | |
| BREAKPOINT | ≥0.25 µg/mL | ≥4 µg/mL |
Table 6.
MIC values for Acinetobacter and Citrobacter strains and Break Point values.
| Strains | Amikacin | Ciprofloxacin |
|---|---|---|
| A. Iwofii 54EB | 2 µg/mL | 0.25 µg/mL |
| A. calcoaceticus 55EB | 2 µg/mL | 0.25 µg/mL |
| A. Iwofii 56EB | 2 µg/mL | 0.25 µg/mL |
| A. Iwofii 57EB | 2 µg/mL | 0.25 µg/mL |
| BREAKPOINT | ≥8 µg/mL | ≥1 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Camellini, S.; Iseppi, R.; Condò, C.; Messi, P. Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Appl. Sci. 2021, 11, 7177. https://doi.org/10.3390/app11167177
AMA Style
Camellini S, Iseppi R, Condò C, Messi P. Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors. Applied Sciences. 2021; 11(16):7177. https://doi.org/10.3390/app11167177
Chicago/Turabian StyleCamellini, Stefania, Ramona Iseppi, Carla Condò, and Patrizia Messi. 2021. "Ready-to-Eat Sandwiches as Source of Pathogens Endowed with Antibiotic Resistance and Other Virulence Factors" Applied Sciences 11, no. 16: 7177. https://doi.org/10.3390/app11167177
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.