
Nanoparticles—Plant Interaction: What We Know, Where We Are?


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Apoplast
2.1. Cell Wall Pore Size
2.2. Documented Entrance of NP into Apoplast
2.3. Postulated Mechanism That Allows NP to Cross the Cell Wall
3. Symplast
3.1. Data Indicating a Symplasmic Translocation of NP through PD
3.2. Possible Explanation of NP Movement through PD
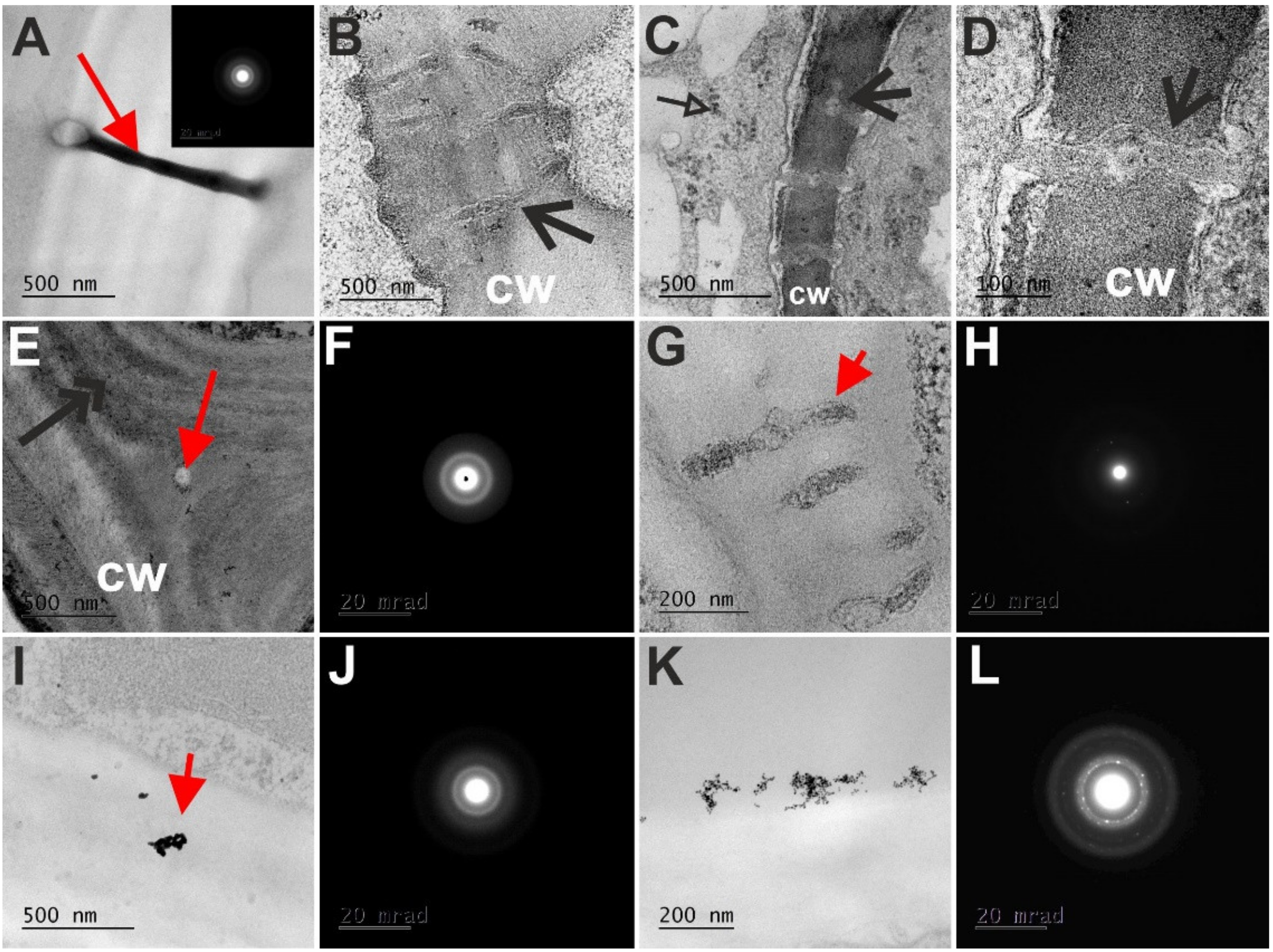
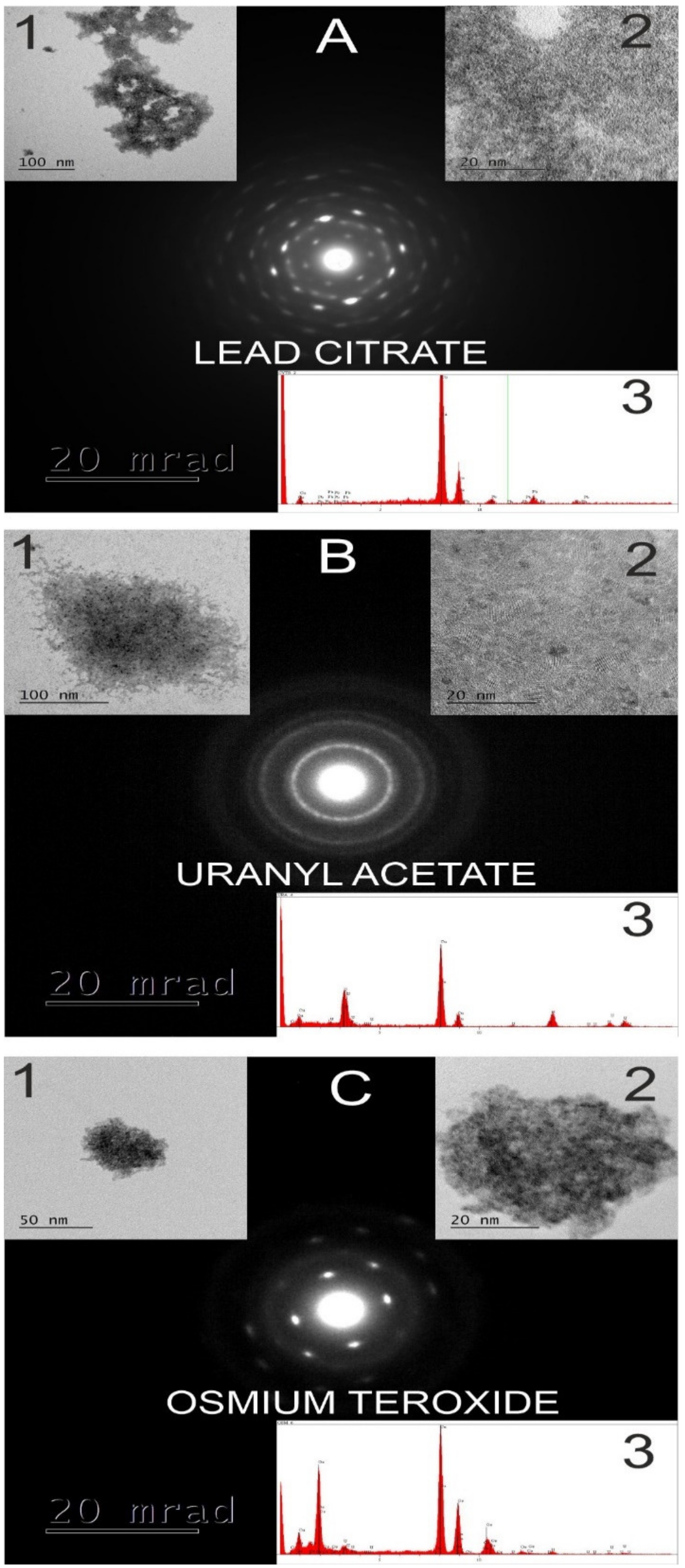
4. Transmission Electron Microscope (TEM) Analysis of the NP in the Symplast and Apoplast
5. Conclusions and Prospectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feynman, R. There’s plenty of room at the bottom. California Institute of Technology. J. Eng. Sci. 1960, 4, 23–36. [Google Scholar]
- Boholm, M.; Arvidsson, R. A definition framework for the terms nanomaterial and nanoparticle. NanoEthics 2016, 10, 25–40. [Google Scholar] [CrossRef]
- Buzea, C.; Pacheco, I. Nanomaterials and their classification. In EMR/ESR/EPR Spectroscopy for Characterization of Nanomaterials. Advanced Structured Materials; Shukla, A., Ed.; Springer: New Delhi, India, 2017; Volume 62, pp. 3–45. [Google Scholar] [CrossRef]
- Janković, N.Z.; Plata, D.L. Engineered nanomaterials in the context of global element cycles. Environ. Sci. Nano. 2019, 6, 2697–2711. [Google Scholar] [CrossRef] [Green Version]
- Arruda, S.C.C.; Silva, A.L.D.; Galazzi, R.M.; Azevedo, R.A.; Arruda, M.A.Z. Nanoparticles applied to plant science: A review. Talanta 2015, 131, 693–705. [Google Scholar] [CrossRef]
- Liu, W.; Zeb, A.; Lian, J.; Wu, J.; Xiong, H.; Tang, J.; Zheng, S. Interactions of Metal-based and Metal-oxide-based Nanoparticles (MBNPs and MONPs) with Crop Plants: A Critical Review of Research Progress and Prospects. Environ. Rev. 2020, 28, 294–310. [Google Scholar] [CrossRef]
- Nikam, A.V.; Prasad, B.L.V.; Kulkarni, A.A. Wet chemical synthesis of metal oxide nanoparticles: A review. CrystEngComm 2018, 20, 5091–5107. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Herth, S. Plant nanotoxicology. Trends Plant Sci. 2011, 16, 583–589. [Google Scholar] [CrossRef]
- Millán-Chiu, B.E.; Rodriguez-Torres, M.P.; Loske, A.M. Nanotoxicology in plants. In Green Nanoparticles. Nanotechnology in the Life Sciences; Patra, J., Fraceto, L., Das, G., Campos, E., Eds.; Springer: Cham, Switzerland, 2020; pp. 43–76. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; de Silva, K.; Nedosekin, D.A.; Dervishi, E.; Biris, A.S.; Shashkov, E.V.; Galanzha, E.I.; Zharov, V.P. Complex genetic, photothermal and photoacoustic analysis of nanoparticle-plant interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 1028–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of metal and metal oxide nanoparticles on plant: A critical review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Hossain, Z.; Mustafa, G.; Komatsu, S. Plant responses to nanoparticle stress. Int. J. Mol. Sci. 2015, 16, 26644–26653. [Google Scholar] [CrossRef] [Green Version]
- Shang, H.; Guo, H.; Ma, C.; Li, C.; Chefetz, B.; Polubesova, T.; Xing, B. Maize (Zea mays L.) root exudates modify the surface chemistry of CuO nanoparticles: Altered aggregation, dissolution and toxicity. Sci. Total Environ. 2019, 690, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Tarrahi, R.; Mahjouri, S.; Khataee, A. A review on in vivo and in vitro nanotoxicological studies in plants: A headlight for future targets. Ecotoxicol. Environ. Saf. 2021, 208, 111697. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Suskova, S.; Mandzhieva, S.; Tsitsuashvili, V.; Chapligin, V.; Fedorenko, A. Effects of Copper Nanoparticles (CuO NPs) on Crop Plants: A Mini Review. BioNanoScience 2018, 8, 36–42. [Google Scholar] [CrossRef]
- Zhou, P.; Adeel, M.; Shakoor, N.; Guo, M.; Hao, Y.; Azeem, I.; Li, M.; Liu, M.; Rui, Y. Application of Nanoparticles Alleviates Heavy Metals Stress and Promotes Plant Growth: An Overview. Nanomaterials 2021, 11, 26. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanzari, I.; Leone, A.; Ambrosone, A. Nanotechnology in plant science: To make a long story short. Front. Bioeng. Biotechnol. 2019, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Schwab, F.; Zhai, G.; Kern, M.; Turner, A.; Schnoor, J.L.; Wiesner, M.R. Barriers, pathways and processes for uptake, translocation and accumulation of nanomaterials in plants–Critical review. Nanotoxicology 2016, 10, 257–278. [Google Scholar] [CrossRef]
- Salmén, L. Micromechanical understanding of the cell-wall structure. Comptes Rendus Biol. 2004, 327, 873–880. [Google Scholar] [CrossRef]
- Rondeau-Mouro, C.; Defer, D.; Leboeuf, E.; Lahaye, M. Assessment of cell wall porosity in Arabidopsis thaliana by NMR spectroscopy. Int. J. Biol. Macromol. 2008, 42, 83–92. [Google Scholar] [CrossRef]
- Carpita, N.; Sabularsed, D.; Montezinos, D.; Delmer, D.P. Determination of the pore size of cell walls of living plant cells. Science 1979, 205, 1144–1147. [Google Scholar] [CrossRef]
- Fleischer, A.; O’Neill, M.A.; Ehwald, R. The pore size of non-graminaceous plant cell walls is rapidly decreased by borate ester cross-linking of the pectic polysaccharide rhamnogalacturonan II. Plant Physiol. 1999, 121, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Money, N.P. Measurement of pore size in the hyphal cell wall of Achlya bisexualis. Exp. Mycol. 1990, 14, 234–242. [Google Scholar] [CrossRef]
- Flournoy, D.S.; Kirk, T.K.; Highley, T.L. Wood decay by brown-rot fungi: Changes in pore structure and cell wall volume. Holzforschung 1991, 45, 383–388. [Google Scholar] [CrossRef]
- Berestovsky, G.N.; Ternovsky, V.I.; Kataev, A.A. Through pore diameter in the cell wall of Chara corallina. J. Exp. Bot. 2001, 52, 1173–1177. [Google Scholar] [CrossRef]
- Gao, X.; Zhuang, S.; Jin, J.; Cao, P. Bound water content and pore size distribution in swollen cell walls determined by NMR technology. BioResources 2015, 10, 8208–8224. [Google Scholar] [CrossRef]
- Milewska-Hendel, A.; Zubko, M.; Karcz, J.; Stróż, D.; Kurczyńska, E. Fate of neutral-charged gold nanoparticles in the roots of the Hordeum vulgare L. cultivar Karat. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujino, T.; Itoh, T. Changes in pectin structure during epidermal cell elongation in pea (Pisum sativum) and its implications for cell wall architecture. Plant Cell Physiol. 1998, 39, 1315–1323. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Christie, P.; Zhang, S. Uptake, translocation and transformation of metal-based nanoparticles in plants: Recent advances and methodological challenges. Environ. Sci. Nano. 2019, 6, 41–59. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.Á.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2018, 53, 185–201. [Google Scholar] [CrossRef]
- Zhang, W.; Dan, Y.; Shib, H.; Ma, X. Elucidating the mechanisms for plant uptake and in-planta speciation of cerium in radish (Raphanus sativus L.) treated with cerium oxide nanoparticles. J. Environ. Chem. Eng. 2017, 5, 572–577. [Google Scholar] [CrossRef] [Green Version]
- Ullah, S.; Adeel, M.; Zain, M.; Rizwan, M.; Irshad, M.K.; Jilani, G.; Rui, Y. Physiological and biochemical response of wheat (Triticum aestivum) to TiO2 nanoparticles in phosphorous amended soil: A full life cycle study. J. Environ. Manage. 2020, 263, 110365. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Qayyum, M.F.; Ok, Y.S.; Adrees, M.; Ibrahim, M.; Abbas, F. Effect of metal and metal oxide nanoparticles on growth and physiology of globally important food crops: A critical review. J. Hazard. Mater. 2017, 322, 2–16. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef]
- Abbas, Q.; Yousaf, B.; Ali, M.U.; Munir, M.A.M.; El-Naggar, A.; Rinklebe, J.; Naushad, M. Transformation pathways and fate of engineered nanoparticles (ENPs) in distinct interactive environmental compartments: A review. Environ. Int. 2020, 138, 105646. [Google Scholar] [CrossRef] [PubMed]
- Iannone, M.F.; Groppa, M.D.; Zawoznik, M.S.; Coral, D.F.; van Raap, M.B.F.; Benavides, M.P. Magnetite nanoparticles coated with citric acid are not phytotoxic and stimulate soybean and alfalfa growth. Ecotoxicol. Environ. Saf. 2021, 211, 111942. [Google Scholar] [CrossRef] [PubMed]
- Thuesombat, P.; Hannongbua, S.; Akasit, S.; Chadchawan, S. Effect of silver nanoparticles on rice (Oryza sativa L. cv. KDML 105) seed germination and seedling growth. Ecotoxicol. Environ. Saf. 2014, 104, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Tombuloglu, H.; Slimani, Y.; Tombuloglu, G.; Korkmaz, A.D.; Baykal, A.; Almessiere, M.; Ercan, I. Impact of superparamagnetic iron oxide nanoparticles (SPIONs) and ionic iron on physiology of summer squash (Cucurbita pepo): A comparative study. Plant Physiol. Biochem. 2019, 139, 56–65. [Google Scholar] [CrossRef]
- Molnár, A.; Papp, M.; Kovács, D.Z.; Bélteky, P.; Oláh, D.; Feigl, G.; Szőllősi, R.; Rázga, Z.; Őrdőg, A.; Erdei, L.; et al. Nitro-oxidative signalling induced by chemically synthetized zinc oxide nanoparticles (ZnO NPs) in Brassica species. Chemosphere 2020, 251, 126419. [Google Scholar] [CrossRef]
- Albenne, C.; Canut, H.; Hoffmann, L.; Jamet, E. Plant Cell Wall Proteins: A Large Body of Data, but What about Runaways? Proteomes 2014, 2, 224–242. [Google Scholar] [CrossRef] [Green Version]
- Navarro, E.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.J.; Sigg, L. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef] [Green Version]
- Kurepa, J.; Paunesku, T.; Vogt, S.; Arora, H.; Rabatic, B.M.; Lu, J.; Wanzer, M.B.; Woloschak, G.E.; Smalle, J.A. Uptake and distribution of ultrasmall anatase TiO2 Alizarin red S nanoconjugates in Arabidopsis thaliana. Nano Lett. 2010, 10, 2296–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaskowiak, J.; Kwasniewska, J.; Milewska-Hendel, A.; Kurczynska, E.U.; Szurman-Zubrzycka, M.; Szarejko, I. Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots. Int. J. Mol. Sci. 2019, 20, 3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mccann, M.C.; Shi, J.; Roberts, K.; Carpita, N.C. Changes in Pectin Structure and Localization during the Growth of Unadapted and Nacl-Adapted Tobacco Cells. Plant J. 1994, 5, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Schmohl, N.; Horst, W.J. Cell wall pectin content modulates aluminium sensitivity of Zea mays (L.) cells grown in suspension culture. Plant Cell Environ. 2000, 23, 735–742. [Google Scholar] [CrossRef]
- Eticha, D.; Stass, A.; Horst, W.J. Localization of aluminium in the maize root apex: Can morin detect cell wall-bound aluminium? J. Exp. Bot. 2005, 56, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.L.; Li, Y.Y.; Zhang, Y.J.; Zhang, S.S.; Wu, Y.R.; Wu, P.; Zheng, S.J. Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex. Plant Physiol. 2008, 146, 602–611. [Google Scholar] [CrossRef]
- Milewska-Hendel, A.; Chmura, D.; Wyrwał, K.; Kurczyńska, E.U.; Kompała-Bąba, A.; Jagodziński, A.M.; Woźniak, G. Cell wall epitopes in grasses of different novel ecosystem habitats on post-industrial sites. Land Degrad. Dev. 2021, 32, 1680–1694. [Google Scholar] [CrossRef]
- Shi, H.; Kim, Y.; Guo, Y.; Stevenson, B.; Zhu, J.K. The Arabidopsis SOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion. Plant Cell. 2003, 15, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Iraki, N.M.; Singh, N.; Bressan, R.A.; Carpita, N.C. Cell walls of tobacco cells and changes in composition associated with reduced growth upon adaptation to water and saline stress. Plant Physiol. 1989, 91, 48–53. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.; Darvill, A.G.; Fry, S.C.; Albersheim, P. Structure and function of the primary cell walls of plants. Ann. Rev. Biochem. 1984, 53, 625–663. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Nguema-Ona, E.E.; Vicre-Gibouin, M.; Sorensen, I.; Willats, W.G.T.; Driouich, A. Arabinose-rich polymers as an evolutionary strategy to plasticize resurrection plant cell walls against desiccation. Planta 2013, 237, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Zemke-White, W.L.; Clements, K.D.; Harris, P.J. Acid lysis of macroalgae by marine herbivorous fishes: Effects of acid pH on cell wall porosity. J. Exp. Mar. Biol. Ecol. 2000, 245, 57–68. [Google Scholar] [CrossRef]
- Nie, G.; Zhao, J.; He, R.; Tang, Y. CuO Nanoparticle Exposure Impairs the Root Tip Cell Walls of Arabidopsis thaliana Seedlings. Water Air. Soil Pollut. 2020, 231, 1–11. [Google Scholar] [CrossRef]
- Xiao, Z.; Yue, L.; Wang, C.H.; Chen, F.; Ding, Y.; Liu, Y.; Cao, X.; Chen, Z.; Rasmann, S.; Wang, Z. Downregulation of the photosynthetic machinery and carbon storage signaling pathways mediate La2O3 nanoparticle toxicity on radish taproot formation. J. Hazard. Mater. 2021, 411, 124971. [Google Scholar] [CrossRef]
- Huang, X.; Li, Y.; Chen, K.; Chen, H.; Wang, F.; Han, X.; Yuan, R. NOM mitigates the phytotoxicity of AgNPs by regulating rice physiology, root cell wall components and root morphology. Environ. Pollut. 2020, 260, 113942. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, D.; Seo, S.M.; Kim, D. Physiological effects of zero-valent iron nanoparticles in rhizosphere on edible crop, Medicago sativa (Alfalfa), grown in soil. Ecotoxicology 2019, 28, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, R.V.; Kumari, S.; Choudhary, R.C.; Sharma, S.S.; Pal, A.; Raliya, R.; Saharan, V. Salicylic acid functionalized chitosan nanoparticle: A sustainable biostimulant for plant. Int. J. Biol. Macromol. 2019, 123, 59–69. [Google Scholar] [CrossRef]
- Singh, S.; Vishwakarma, K.; Singh, S.; Sharmab, S.; Kishor, N.; Vivek, D.; Singhe, K.; Liu, A.; Tripathi, D.K.; Chauhan, D.K. Understanding the plant and nanoparticle interface at transcriptomic and proteomic level: A concentric overview. Plant Gene. 2017, 11, 265–272. [Google Scholar] [CrossRef]
- Rico, C.M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Chemistry, Biochemistry of Nanoparticles and Their Role in Antioxidant Defense System in Plants. In Nanotechnology and Plant Sciences; Siddiqui, M., Al-Whaibi, M., Mohammad, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Ma, X.; Yan, J. Plant uptake and accumulation of engineered metallic nanoparticles from lab to field conditions. Curr. Opin. Environ. Sci. Health. 2018, 6, 16–20. [Google Scholar] [CrossRef]
- Dai, Y.; Zhao, J.; Liu, X.; Yu, X.; Jiang, Z.; Bu, Y.; Xing, B. Transformation and species identification of CuO nanoparticles in plant cells (Nicotiana tabacum). Environ. Sci. Nano. 2019, 6, 2724–2735. [Google Scholar] [CrossRef]
- Palocci, C.; Valletta, A.; Chronopoulou, L.; Donati, L.; Bramosanti, M.; Brasili, E.; Pasqua, G. Endocytic pathways involved in PLGA nanoparticle uptake by grapevine cells and role of cell wall and membrane in size selection. Plant Cell Rep. 2017, 36, 1917–1928. [Google Scholar] [CrossRef]
- Boykov, I.N.; Shuford, E.; Zhang, B. Nanoparticle titanium dioxide affects the growth and microRNA expression of switchgrass (Panicum virgatum). Genomics 2019, 111, 450–456. [Google Scholar] [CrossRef]
- Asli, S.; Neumann, P.M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef]
- López-Moreno, M.L.; de la Rosa, G.; Hernández-Viezcas, J.Á.; Castillo-Michel, H.; Botez, C.E. Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environ. Sci. Technol. 2010, 44, 7315–7320. [Google Scholar] [CrossRef] [Green Version]
- Govea-Alcaide, E.; Masunaga, S.H.; de Souza, A.; Fajardo-Rosabal, L.; Effenberger, F.B.; Rossi, L.M.; Jardim, R.F. Tracking iron oxide nanoparticles in plant organs using magnetic measurements. J. Nanopart. Res. 2016, 18, 1–13. [Google Scholar] [CrossRef]
- Sevilem, I.; Miyashima, S.; Helariutta, Y. Cell-to-cell communication via plasmodesmata in vascular plants. Cell Adh. Migr. 2013, 7, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Marzec, M.; Kurczyńska, E.U. Importance of symplasmic communication in cell differentiation. Plant Signal. Behav. 2014, 9, e27931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, K.; Oparka, K. Imaging plasmodesmata. Protoplasma 2011, 248, 9–25. [Google Scholar] [CrossRef]
- Demchenko, K.N.; Voitsekhovskaja, O.V.; Pawlowski, K. Plasmodesmata without callose and calreticulin in higher plants–open channels for fast symplastic transport? Front. Plant Sci. 2014, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Kragler, F. Plasmodesmata: Intercellular tunnels facilitating transport of macromolecules in plants. Cell Tissue Res. 2013, 352, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Dashevskaya, S.; Kopito, R.B.; Friedman, R.; Elbaum, M.; Epel, B.L. Diffusion of anionic and neutral GFP derivatives through plasmodesmata in epidermal cells of Nicotiana benthamiana. Protoplasma 2008, 234, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Stonebloom, S.; Xu, M.; Zambryski, P.C. Plasmodesmata during development: Re-examination of the importance of primary, secondary and branched plasmodesmata structure versus function. Protoplasma 2011, 248, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Hussain, H.I.; Yi, Z.; Siegele, R.; Cresswell, T.; Kong, L.; Cahill, D.M. Uptake and cellular distribution, in four plant species of fluorescently labeled mesoporous silica nanoparticles. Plant Cell Rep. 2014, 33, 1389–1402. [Google Scholar] [CrossRef]
- Geisler-Lee, J.; Wang, Q.; Yao, Y.; Zhang, W.; Geisler, M.; Li, K.; Huang, Y.; Chen, Y.; Kolmakov, A.; Ma, X. Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology 2013, 7, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Walters, K.S.; Peate, D.W.; Alvarez, P.J.; Schnoor, J.L. Transport of gold nanoparticles through plasmodesmata and precipitation of gold ions in woody poplar. Environ. Sci. Technol. Lett. 2014, 1, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradas del Real, A.E.; Vidal, V.; Carrière, M.; Castillo-Michel, H.; Levard, C.; Chaurand, P.; Sarret, G. Silver nanoparticles and wheat roots: A complex interplay. Environ. Sci. Technol. 2017, 51, 5774–5782. [Google Scholar] [CrossRef] [Green Version]
- Spielman-Sun, E.; Lombi, E.; Donner, E.; Howard, D.; Unrine, J.M.; Lowry, G.V. Impact of surface charge on cerium oxide nanoparticle uptake and translocation by wheat (Triticum aestivum). Environ. Sci. Technol. 2017, 51, 7361–7368. [Google Scholar] [CrossRef]
- Sager, R.; Lee, J.Y. Plasmodesmata in integrated cell signalling: Insights from development and environmental signals and stresses. J. Exp. Bot. 2014, 65, 6337–6358. [Google Scholar] [CrossRef] [Green Version]
- Schulz, A. Plasmodesmal widening accompanies the short-term increase in symplasmic phloem unloading in pea root tips under osmotic stress. Protoplasma 1995, 188, 22–37. [Google Scholar] [CrossRef]
- Brandenberger, C.; Clift, M.J.D.; Gehr, P.; Muhlfeld, C.; Rothen-Rutishauser, B.; Stone, V. Intracellular imaging of nanoparticles: Is it an elemental mistake to believe what you see? Part Fibre Technol. 2010, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Lombi, E.; Scheckel, K.G.; Kempson, I.M. In situ analysis of metal(loid)s in plants: State of the art and artefacts. Environ Exp Bot. 2011, 72, 3–17. [Google Scholar] [CrossRef]
- Kennedy, C.D.; Gonsalves, F.A.N. The action of divalent zinc, cadmium, mercury, copper and lead on the trans-root potential and H+ efflux of excised roots. J. Exp. Bot. 1987, 38, 800–817. [Google Scholar] [CrossRef]
- Da Costa, M.V.J.; Sharma, P.K. Effect of copper oxide nanoparticles on growth, morphology, photosynthesis and antioxidant response in Oryza sativa. Photosynth. 2016, 54, 110–119. [Google Scholar] [CrossRef]
- Shukla, P.K.; Misra, P.; Kole, C.H. Uptake, Translocation, Accumulation, Transformation and Generational Transmission of Nanoparticles in Plants. In Plant Nanotechnology; Kole, C., Kumar, D., Khodakovskaya, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 183–218. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurczyńska, E.; Godel-Jędrychowska, K.; Sala, K.; Milewska-Hendel, A. Nanoparticles—Plant Interaction: What We Know, Where We Are? Appl. Sci. 2021, 11, 5473. https://doi.org/10.3390/app11125473
Kurczyńska E, Godel-Jędrychowska K, Sala K, Milewska-Hendel A. Nanoparticles—Plant Interaction: What We Know, Where We Are? Applied Sciences. 2021; 11(12):5473. https://doi.org/10.3390/app11125473
Chicago/Turabian StyleKurczyńska, Ewa, Kamila Godel-Jędrychowska, Katarzyna Sala, and Anna Milewska-Hendel. 2021. "Nanoparticles—Plant Interaction: What We Know, Where We Are?" Applied Sciences 11, no. 12: 5473. https://doi.org/10.3390/app11125473