
A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies

Department of Environmental Studies, University of Portland, Portland, OR 97203, USA
*
Author to whom correspondence should be addressed.
Environments 2023, 10(5), 76; https://doi.org/10.3390/environments10050076
Submission received: 22 March 2023
/
Revised: 14 April 2023
/
Accepted: 30 April 2023
/
Published: 2 May 2023
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:We view the history of the Columbia River Basin through a resilience lens from the point of view of salmonids, as keystone species for the river basin ecosystems and social systems. We rely on the concept of multiple stable states as depicted in a stability landscape, as a scientific theory, but equally as a metaphor and a mental model. Using evidence-based plausibility arguments concerning the existence, creation, and potential critical transitions between regimes, we describe change over centuries. We argue that a critical transition occurred taking the state of the system from its historic regime into a novel regime stabilized by new social feedbacks and institutional configurations. By using a state space defined by four variables used in policy deliberations for salmon recovery we tie our results to historical and contemporary management issues. Knowledge of (a) which regime is currently occupied and (b) which critical transitions between regimes are possible are both crucial to effective policy formation. We draw distinctions between positions held by federal agencies, tribal agencies, and civil society organizations as to the current state of affairs and policy recommendations, raising questions about the appropriate use of decision support systems in the public process for decision making.
1. Introduction
Salmon have played a central role for the peoples and ecosystems of the Columbia River Basin (CRB) for thousands of years. The Pacific Northwest of the U.S. has been characterized by writer Timothy Egan as “any place a salmon can get to”, but the role of these crucial species extends beyond defining geography, as salmon occupy central roles in ecological, social, cultural, and economic systems. However, these keystones of the Pacific Northwest are in serious decline. Estimates of historical levels of annual returns range from 10 to 16 million fish annually, but these numbers have dwindled to between 750 thousand and 3 million individuals per year, with at least 80% of these originating from hatcheries [1]—an important consideration as it was the genetic and life history diversity of the 6 salmonid species that lent the salmon metapopulation its original resilience [2]. The National Oceanic and Atmospheric Administration (NOAA) [3] estimates that 29% of nearly 1400 historical populations of Pacific salmon have become extinct since Euro-American contact. Among the remaining populations, over one-third are listed as threatened or endangered under the U.S. Endangered Species Act. Exacerbating the sense of urgency are predictions by climatologists that the next few decades will bring significant changes in northwestern stream flow and temperature, placing further stress on already depleted runs of salmon. Climate change impacts will be felt especially in the low-elevation basins of northeast Oregon, the north Washington Cascades, and in north-central Idaho. Juvenile salmon will have to migrate to the ocean earlier through lower tributary rivers, and adults returning to spawn will face the challenge of lower and warmer summer flows, and more scouring winter floods will threaten their eggs [4]. The path to salmon recovery may require the removal of some of the 31 hydroelectric dams that provide power for industrial and residential users throughout the Pacific Northwest and beyond to the west coast, an action that would radically change the regional economy and perhaps contravene an international treaty. Governments of two countries, six states, several dozen Native American nations in the U.S. and First Nations in Canada—some federally recognized and others not—and hundreds of counties and municipalities have a stake in reaching management solutions palatable to their constituencies. In spite of the most extensive and expensive ecological restoration effort in U.S., if not world, history, little if any progress has been made in restoring the salmon and the ecosystem on which they depend. Even so, after all the decades and all the billions of dollars, the region is alive with innovation, motivation, and determination to restore the ecological, social, and cultural place known as “Salmon Nation” [5].
2. Methods
In this section, we give an outline of the resilience history of the Columbia River Basin from the point of view of the salmon-centered ecosystem—a specified resilience [6] approach. The outcome of this description is a proposed stability landscape for the social-ecological system specified by the salmonid species and encompassing the social, ecological, economic, and governance systems involved in their welfare.
In describing a regime in a social-ecological system, we begin by proposing a collection of state variables—the central factors that can be used to define the state space, specify the current state of the system, describe the fundamental structure, feedbacks, and processes that constitute the overall system, and characterize the identities of the regimes it contains. Since 1875, when U.S. Commissioner of Fish and Fisheries Spencer Baird wrote a strongly worded report about the precipitous decline in Columbia River salmon and identified dams, habitat change, and overharvest as critical issues [7], four key factors known as the 4 Hs are now commonly recognized as central to the welfare of salmon: harvest, habitat, hydro (the system of hydroelectric dams on the river), and the additional post-1875 factor of hatcheries. We will adopt these 4 Hs as the state variables for our discussion, both due to their comprehensive descriptive power and for the connection they afford to the history of recovery planning. It is likely impossible to determine whether it is objectively true or not that the current state of the system is completely determined by the 4 Hs. Nonetheless, this assumption is made by the vast majority of the stakeholders participating in recovery planning. At a minimum, these four variables define the planning space for salmon recovery in the Columbia River Basin.
There are a number of ways regimes in a complex social-ecological system can be specified. We take the approach that a regime is characterized by three elements: its identity [8]; the stabilizing factors, both stabilizing feedbacks and institutions [9], that underlie its resilience; and the thresholds and accelerating feedbacks [10] that describe its relationship to adjacent regimes. We will use these elements to make arguments for the plausibility of four regimes: historic, refugia, techno, and remnant.
As we will be considering the history of salmon in the CRB from a resilience perspective, we will need to specify at different times which regime the system state currently occupies and whether it has made a shift, a critical transition, to another regime. Here we take an approach using the concept of regime identity as well as a consideration of which stabilizing feedbacks and institutions are dominant at the time [11].
Since the foundational work on multiple stable states in forest ecosystems [12], changes in relatively slow controlling variables have been identified as indicators, even drivers, of critical transitions in complex adaptive systems. As described by Carpenter et al. [13], slow variables “control the boundaries of the domain of attraction” and play a prominent role in “defining the underlying structure of the system”. In mathematical models [14], the variables are often separated, according to relative rates of change, into fast and slow categories where the fast variables play the role of state variables, and the slow variables are often treated as parameters. Variations in certain slow variables may change the resilience, configuration, and number of basins of attraction, and can induce critical transitions from one regime to another. Following other authors (e.g., [15]), we will call a slow variable associated with changes in the number of basins of attraction a critical slow variable, since they represent “the critical objects of monitoring, as they are the key indicators of ecosystem resilience” [16]. We argue below that life history diversity in the salmonid metapopulation is such a critical slow variable.
Prior to contact with European populations, the Columbia River Basin supported diverse and abundant seasonal runs of a variety of anadromous salmonids returning from the sea to spawn in either the mainstem river or tributaries. For thousands of years these populations, characteristically returning to their natal stream to breed, made up a dynamic series of genetically distinct subpopulations that utilized at some stage in their life history virtually all the habitats in time and space that the river system afforded, and which provided a major influx of nutrients of marine origin into the inland northwest. The term life history specifies one of these geographic and temporally distinct breeding populations while the term metapopulation refers to this modular network of all the life histories in the basin.
We use the term life history diversity to refer to the diversity of the metapopulation as a whole. Much like the composite state variables, the 4 Hs, life history diversity is a composite variable. We distinguish three components of life history diversity: the genetic diversity within and among the collection of separate runs; the modular network structure and complexity of the geographic distribution of their natal streams; and the temporal diversity of the timing of their in- and out-migrations to the ocean. For the purposes of this paper, the network structure and complexity of life history diversity are particularly crucial. It is important to note that the network structure itself has a temporal dimension. Connectivity and barriers to movement ebb and increase with seasonal changes in stream flow rates, with consequent changes in stream levels.
The geographic specificity of these life histories can be remarkably precise [17]—for example, the natal stream of one salmon life history is restricted to 200 m of stream between Multnomah Falls and the Columbia River. However, salmon do not always return to their natal streams, and this is a key to their resilience. At a rate of about 1%, they stray to other breeding populations, giving the larger network of life histories the ability to repopulate in response to a localized extirpation and to increase the genetic diversity within a given life history. This network structure of tightly knit subgroups connected to each other via weak links is often called a modular system and is cited as an important source of resilience in complex adaptive systems generally [18], and in linked social and ecological systems in particular [19,20,21,22]. Ali Kharrazi and coauthors note [23] “While modularity, redundancy, and diversity impact the resiliency of a system, less is known of the joint impact of these system level properties. Tradeoffs between modularity, diversity, and redundancy is an important research frontier for social-ecological systems”. Lichatowich describes for salmonids [24],
“In undisturbed rivers, each salmon population is composed of a bundle of several life histories, or several alternative survival strategies. Unlike the salmon raised in a hatchery environment with its feedlot regime, the salmon in a natural population in a healthy river do not all do the same thing in the same place at the same time. They follow different pathways through the time and space dimensions of their habitat. As the riverscape changes due to natural disturbances (fires, floods, droughts, and so on) some of the salmon’s life histories are survival peaks while others drop into troughs. This diverse array of life histories diminishes the risk of catastrophic mortality and loss of an entire population in a naturally changing environment.”
It is obvious that the image of a metapopulation distributed across a terrain with weak genetic connections, and life histories that are survival peaks with troughs between them, corresponds in many ways to Sewall Wright’s shifting balance theory of evolution [25,26,27]. The local adaptations of different salmon life histories spawning in local reaches of a shifting river system provide a context highly generative for evolutionary adaptations. According to Wright, a species divided into many local units largely breeding internally but with occasional outcrossing (both of which for salmon have temporal as well as spatial dimensions) will undergo intergroup selection that rapidly increases the average adaptiveness of the local populations. Wright believed that this circumstance provides the most effective situation for trial and error among genetic combinations. All of these perspectives should be borne in mind when we discuss the advent of hatcheries, whose operations violated all of the conditions that Wright held to be valuable for generating highly adaptive populations.
While the modular structure of the salmon metapopulation is fundamental to its resilience, the complexity of that system is also relevant to the existence of thresholds. Much as in the better-known case of regime shifts due to a decreased connectivity in spatially specified terrestrial ecosystems [28,29], recent research supports a similar phenomenon in riverine networks. Here patchiness also occurs but has a distinct network characterization in the very different dendritic topology of aquatic systems [30]. Recent modeling [31,32,33] suggests that loss of topological complexity [34] in a dendritic ecological network is associated with an increased risk of extinction and nonlinear effects on the stability of fish metapopulations. Salmon are particularly vulnerable to a reduction in the complexity of their “dendritic landscapes, where fragmentation typically results in smaller and more variably-sized fragments, thus reducing metapopulation persistence relative to linear systems” [35]. Modeling of dendritic networks reveals that the “interplay between connectivity and dispersal influences metapopulation persistence” [36]. In our case, this highlights the critical influence of a loss of complexity in the riverine system, which we will call dendritic complexity, combined with the loss of the connectivity required for natural repopulation through straying, to produce increased risks of extinctions. Letcher et al. [37] connect the themes of modularity, life history, and complexity in a modeling study of brook trout population dynamics, finding that as connectivity in a network decreases, local extirpation risks increase, further concluding that local extirpations can trigger network-wide extinctions.
In addition, due to the well-established dependence of the surrounding terrestrial systems on salmon for nutrient flows [38,39,40], a loss of dendritic complexity in the salmon metapopulation could well be associated with regime shifts in those ecosystems. Within the aquatic environment, Gresh et al. [41] estimate that “6–7% of the marine-derived nitrogen and phosphorous once delivered to the rivers of the Pacific Northwest is currently reaching those streams” and argue that this “has contributed to the downward spiral of salmonid abundance and diversity in general, further diminishing the possibility of salmon population recovery to self-sustaining levels.” The nutrients that salmon brought to the inland Pacific Northwest once provided necessary materials for the growth of the forests of tall trees, that in turn shaded and cooled the streams and rivers producing excellent conditions for salmon reproduction. In addition to their function as a keystone species for aquatic ecosystems, salmon have been shown to play a similar role for wildlife in terrestrial ecosystems [42]. A previous study [43] demonstrated the striking effects of the decline of salmonid nutrient subsidies for terrestrial plant biodiversity. This study also concluded that understanding “how salmon affect terrestrial ecosystems is central to conservation plans that aim to better integrate ecosystem values into resource management.” The dendritic complexity of the salmon-occupied network plays a crucial role as “the occurrence of runs in very small streams has important consequences for wildlife biology”. Indeed, local extirpation of eagle populations due to the loss of nutrients from salmon has been observed as far upstream as Montana [44]. Thus, loss of dendritic complexity in salmon-occupied riverine networks may be associated with thresholds for cascading effects quite broadly.
3. Regimes and Critical Transitions
3.1. The Historic Regime
The intricacy of what we will call the historic regime (see Figure 1) of salmon in the Columbia River Basin has been researched as part of ongoing efforts to plan for salmon recovery efforts. Salmon populations are often described by the timing of their entry into freshwater on their return from the ocean to spawn (e.g., spring chinook, summer chinook, fall chinook). The division of the species into multiple runs allows subpopulations to take advantage of upper tributary habitats and migration corridors that vary dramatically in a seasonal fashion. High early spring flows can make passage possible over falls that would pose an impassible barrier at the lower water levels prevalent later in the season. Some runs of fish migrate for a considerable time before spawning, with the adults remaining above river areas that they could no longer pass until their spawning season occurs. As Brannon et al. [45] describe:
“Life history forms, therefore, are genetically predisposed to strategies that optimize survival under the restraints of local stream environments. When referring to life history strategy, we are describing a behavioral repertoire that has proven successful for a particular population of the species under given environmental constraints. For example, fall Chinook that utilize a stream for incubation and early rearing that is unsuitable later in the summer or winter will acquire a short-term pattern of residence and disperse downstream before unsuitable conditions arise…”
According to Brannon et al., the residence time of juveniles in their natal stream system, prior to outmigration to the ocean, is variable. For example, ocean-type juvenile chinook leave freshwater their first year to migrate to the ocean, and stream-type chinook remain in freshwater for a year prior to outmigration. Different seasonal runs of chinook tend to dominantly demonstrate one or the other of these life histories (spring chinook are generally stream-type and fall chinook are generally ocean-type) but enough variability exists to add a layer of complexity and, therefore, ecological resilience in the face of a fluctuating marine environment.
Temperature considerations are crucial as well, with different life stages of species of salmonids having critical temperatures that vary by 5 to 6 degrees C [46] with temperatures as low as 10 degrees C representing the upper optimal limit for spawning and incubation. Since different stream systems have different temperature profiles across a season and longitudinally from year to year [47], the passage of life history stages through seasonal temperature progressions becomes an intricate evolutionary dance.
One approach to describing historical population structures defines strata as groups of local populations sharing the same ecological zone, and with different run timings. For example, McElhaney et al. note [48] that in the Cascade ecological zone of the Lower Columbia River, the fall chinook stratum is believed to have comprised 9 historical populations and overall chinook salmon in the lower Columbia River, to the point where Bonneville Dam now stands, are believed to have comprised 31 local populations with a significant degree of genetic isolation between them. Overall, salmonids and steelhead, not including the now virtually extirpated sockeye salmon, are believed to have comprised 82 local populations between the Bonneville Dam site and the mouth of the river. Recovery planning has proceeded on the basis that the historical population structure provided a degree of resilience in the face of environmental challenges. As also noted by McElhaney et al.:
“We approached the stratum risk criteria from the perspective of the historical template. The approach is based on the principle that the historical population structure of the strata produced a relatively low risk of extinction, and the closer the population structure is to that historical structure, the lower its extinction risk. Conversely a population structure that deviates greatly from the historical structure would be considered at high risk.”
Genetic analyses of existing fish on the basis of isozymes and DNA variability support this almost baroque degree of population subdivision [49].
Prior to human interventions, the salmon metapopulation largely maintained its identity, characterized by high levels of life history diversity punctuated by some quite dramatic disturbances, for millions of years. Local events such as landslides or volcanic activity would temporarily diminish the salmon populations in specific reaches of the river system, but even a small number of fish, straying from their natal stream when they returned from the ocean to breed, could re-colonize a denuded area. Larger events might have blocked passage in the main channel for a time, such as the Hood River lahar of 100,000 years ago which would have interrupted the river’s flow very significantly. This lahar from Mt. Hood probably blocked passage entirely for several years. This is described as follows:
“About 100,000 years ago, a large portion of the volcano’s north flank and summit collapsed. The resulting debris avalanche transformed into a lahar that swept down the Hood River valley. At the river’s mouth, where the town of Hood River now stands, the lahar was 400 feet deep. The lahar crossed the Columbia River and surged up the White Salmon River valley on the Washington side. Since that time lava has filled in the scar left by the debris avalanche”.[50]
The recovery of the salmon-centered ecosystem from system-wide events such as this—and even more dramatic ones farther in the past—gives testimony to the remarkable resilience provided by the modular system [51].
Archeological evidence shows that 8000 or 9000 years ago, the region’s Native Americans were eating primarily land mammals that they hunted, although in some sites salmon were already very important [52]. The development of smoking and drying as techniques for salmon preservation approximately 3000 years ago changed that, as dense populations began to settle where salmon were seasonally abundant but now could offer a food source throughout the year thanks to the new technological advances. Doubtless annual fish return levels were fluctuating because of the effects of ENSO and the Pacific Decadal Oscillation on ocean conditions, in terms of ambient temperature conditions, on the abundance of food, and also on stream flows in the Columbia Basin [53]. At times there was doubtless scarcity, perhaps especially in spring when dried salmon supplies might be exhausted, but such events would have strengthened the native population’s emphasis on conservation rather than the reverse. As Joseph Taylor [54] summarized:
“Indians were seasoned rational fishers. They developed a culture and economy that meshed well with nature. Respect, propitiation, utility, and territoriality reinforced a coherent, moderate strategy of exploitation. The synergistic result was a culturally specific form of conservation. No single mechanism entirely mitigated the potential for destruction, but in combination they worked wonders. Fishing techniques could wreak havoc on salmon runs, and cultural routes to prestige encouraged heavy consumption; but cultural forms of respect moderated harvest, and the technical limitations of food preservation capped storage capacity.”
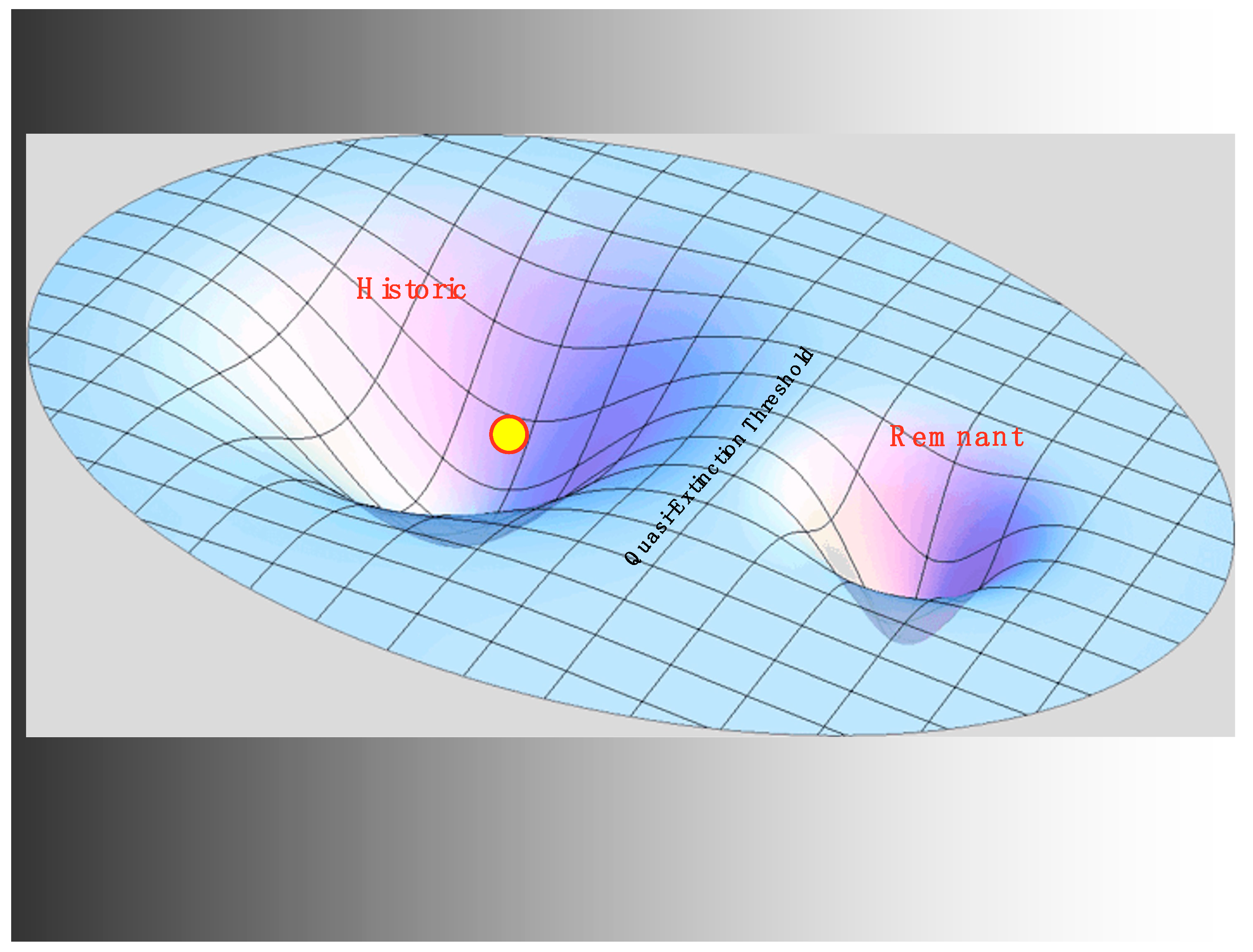
In terms of the stability landscape, this overall situation might be summarized as having an extremely resilient regime, characterized by an extensive modular distribution of fluctuating but generally abundant populations of various runs of salmonid species. The resilience of the regime is produced by this modular metapopulation as well as a robust collection of balancing social-ecological feedbacks associated with cultural norms in the Native American communities occupying the basin. We will designate this basin of attraction as the historic regime. As O’Connor [55] summarizes:
“The recent ages of some of the geologic events that formed these features show quite emphatically that the pre-1805 landscape—including the land, people, and ecosystems—was not static but one of drastic and dramatic change… The large volcanic eruptions of Mount Hood in about 500 and again in the 1780s, as well as the large Mount St. Helens eruptions of about 1480, almost certainly had large effects on the Columbia River and the ecosystems and occupants that depended on it… Large sediment pulses into the river from volcanic eruptions are analogous to, but far larger than, sedimentation caused by land-use practices. The huge Missoula Floods of the last ice age reshaped and locally dredged the Columbia River channel…Yet, the tremendous natural resources sustaining the Native American populations at the time of Lewis and Clark attest to the resilience of the Columbia ecosystem in the face of such huge disturbances, even ones of just a few decades or centuries before.”
Climatic fluctuations on the land, oscillating ocean conditions, self-regulating Native American harvests of salmon, and natural events that blocked and unblocked tributary streams at various times were all contained within what might be envisioned as a social-ecologically founded system of two regimes, the deep historic regime that typically contained the momentary state of the system, and a remnant regime, characterized by very low life history diversity and resilience provided by the stabilizing factor known as the Allee effect [56], with the two regimes separated by a threshold we call the quasi-extinction threshold. The multiple geographic breeding areas, multiple species and temporally diverse runs of salmonids in the region, a combination of dispersion, connectedness, and life history diversity [57] all served to build high levels of resilience and resistance that favored the system state remaining in the historic regime or returning to it after external conditions recovered from major disturbances. Multiple years spent maturing at sea, with some degree of variability among members of each cohort, buffered the system against bad ocean conditions due to the fluctuations of the Pacific Decadal Oscillation. Multiple breeding tributaries meant that if a wildfire, landslide, or volcanic event cut one tributary off from the sea for a time, or made it inhospitable to salmonids, there were always recolonizers available among the small percentage of strays that return from the ocean to someplace other than their natal stream. Biological diversity combined with an appropriately complex network structure produces stability, and, in this instance, both geographic and biological diversity in salmonid population reproduction made for a system with great potential to endure.
3.2. Habitat, Harvest, and a Preview of the Refugia Regime
The arrival of a large influx of settlers, largely from Europe, and their new economic activities, rapidly began to change this picture of dynamic stability. We can describe what happened in terms of the 4 Hs, recognized today as the major drivers forcing regional salmonid declines. These are worth considering individually, in roughly the order they became important.
The first H, habitat, began to be modified almost immediately by activities as diverse as logging, mining, raising livestock, and river channelization. Early logging was mainly to clear areas for agriculture, without consideration that river food webs respond strongly to riparian zone alterations. Commercial logging rapidly developed into a very large industry by the end of the 1800s. The conversion of natural resources to a form of currency managed from outside the region made decisions to overexploit the forests of the northwest seem rational from a distance. The environmental consequences were invisible, and maximization of short-term profit seemed a rational choice in the absence of any connection between the decision maker and the land. When distant markets determine the rate of resource exploitation, the immediate stabilizing constraints of strong balancing feedback loops are absent [58]. Floating logs downstream to sawmills, which could scour away the river bottoms in good salmon breeding areas, not only involved the destructive movements of the very large log rafts, but often involved the construction of splash dams; wooden structures that allowed the water level of a stream to be raised until it was time to release a log raft downstream, and which were a significant barrier to salmon migration.
“Severe scouring from splash damming was one of the earliest reported forms of widespread anthropogenic disturbance in streams of the Pacific Northwest, USA. Splash damming was a common method of log transport in western Oregon from the 1880s through the 1950s. Before being released in large freshets to downstream lumber mills, water and logs were stored in reservoirs behind splash dams. Further protocol called for dynamiting downstream obstacles such as large boulders and natural logjams… [resulting in] significantly more bedrock and fewer deep pools in splashed reaches… three times fewer pieces of key large wood were found in splashed reaches… Many of the in-channel variables that demonstrated significant differences are regarded as indicators of salmon habitat quality… splash-dam legacy effects still persist on evaluated stream reaches 50–130 years after the practice ceased…”[59]
From a resilience perspective, splash dams contributed to interference with indigenous salmon management practices and helped trigger local transformations [60], in the face of which the Columbia River Basin nonetheless maintained its resilience through the systemic modularity [61] provided by undammed tributaries. It is also possible that the release of water from splash dams sometimes worked as a flood pulse positive to river ecosystem diversity and productivity [62]. The geographic connectedness of tributaries with spawning areas, so long as some remained untransformed by splash dams, provided the strays, or colonists, from nearby spawning areas needed to repopulate the tributary once a splash dam was removed and the structure of the river bottom and presence of large woody debris that provides shelter for migrating salmonids might slowly recover over time.
The elimination of side channels and braided channels progressed as complex river systems prone to flooding were altered to increase the area available for agriculture and reduce flooding of the developing riverside communities (the Willamette River, an important tributary, for example, lost roughly 80% of its structure due to channel simplification) [63], and even gold mining that used hydraulic cannons occurred that sometimes relied heavily upon Chinese labor for the most arduous work [64]. This process broke off large portions of the riverbank so that they could be sorted for any gold content. Both splash dams and mining impacted specific stretches of the river system, and from the perspective of salmon populations reduced the geographical extent and dendritic complexity of the system, rather than changing the basic nature of the Columbia River Basin.
Although contemporary innovations such as payment for ecosystem services represent an expression of increased levels of adaptability [65] in the “habitat arena,” historical habitat impacts took place in a period of low adaptability. Indeed, there are few if any signs of innovations that built adaptive capacity in early logging operations, splash dam construction, side channel elimination, or even later developments such as the riparian deployment of synthetic organic pesticides [66].
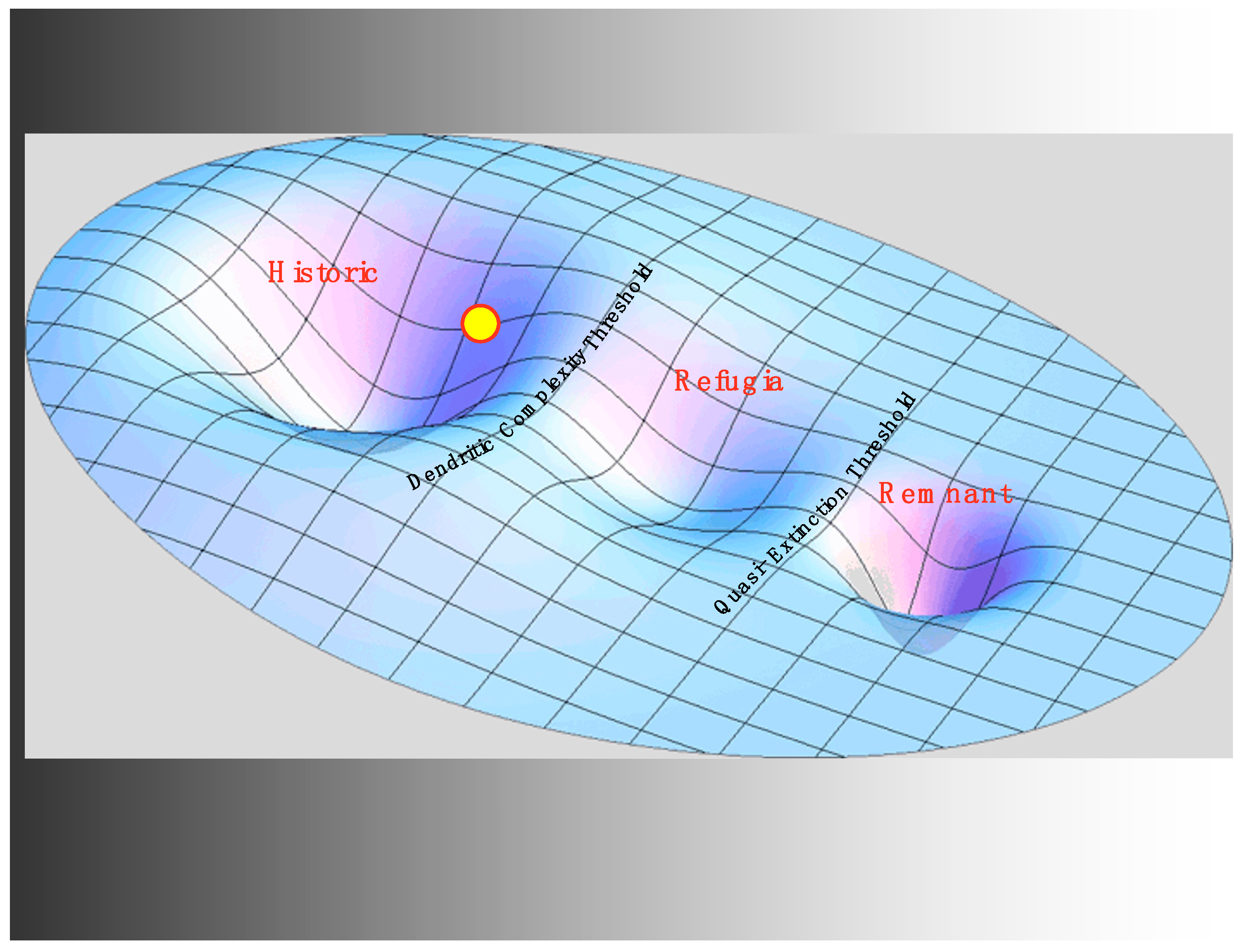
Although habitat modifications were locally extreme, it would be an exaggeration to say that the current state of the system left the historic regime immediately when large-scale habitat modification by settlers began. However, it would not be wrong to view this as a premonition of a simplified system with less connectedness, reduced dendritic complexity, fewer tributaries, and less life history diversity, which we will name the refugia regime. This new possibility began to have a shadowy existence in the face of human habitat modifications, and it became more visible on the stability landscape as more tributaries were lost to mining or splash dams or other activities, and the life history diversity previously contributed by those tributaries was lost. However, as yet, there were no strong stabilizing processes or institutions in place to provide resilience for this nascent regime. We will justify below the existence of the refugia regime (see Figure 2), when these processes appeared, but for now, we observe that habitat modifications reduced the resilience of the historic regime producing persistent directional change driving the state of the system in the direction of the remnant regime.
Changes in the second H, harvest, began to reduce the resilience of the historic regime as the use of technology increasingly replaced the Native American practices used by Tribes and First Nations for millennia. Novel, exogenously derived technologies included new forms of dipnets (large submerged baskets that could contain up to 300 fish), seines as large as 100 feet long and 12 feet deep, fences, and weirs. However, it was the advent of canneries and the resulting connection to global fish markets that had by far the greatest impact. Canneries essentially monetized the fish and changed them from what was primarily a source of sustenance and cultural identity to native peoples and later settlers, into a cash crop for large-scale export to the rest of the world. The development of railroads in the northwest and across the continent was a related technological innovation providing reliable and inexpensive transportation for large quantities of goods to be exported from the northwest to the populous markets of the east coast, and beyond to Great Britain and even Australia.
Hume and Hapgood began to can salmon in 1866. The last spike on the transcontinental railroad was driven in 1869; that year, Hapwood, Hume and Company canned 272,000 pounds of salmon. The year 1870 saw 14 canneries and 25,000,000 pounds of salmon canned, and 1884 saw 37 canneries and 40,000,000 pounds of salmon canned [67]. Although this total poundage approximates that of the fish taken by the Native Americans, cannery operations initially concentrated on the most economically valuable fish, chinook and sockeye, captured between April and July. The canneries also concentrated, spatially, in the lowest 40 miles of the Columbia River, in order to can the fish when they first entered the river and still had all of their stored body fat increasing their nutritional value.
The consequence of this disturbance to life history diversity was that the decline of salmon outpaced even the production demand of the industrial system. As the most valuable species and runs diminished due to overfishing (and overcanning) the cannery operators and fishermen extended their operations to less valuable runs and species. However, this only delayed a final decline in catch. Salmon canning declined on the Columbia River and the industry moved north to the Puget Sound and ultimately British Columbia and Alaska, leaving greatly lessened fish populations in the Columbia River Basin in its wake [68]. As this drama played out, various governmental regulatory bodies and processes too intricate to fully describe here were involved, but none of this was effective in preventing the diminution of salmonids in the Columbia River Basin. As described in a history prepared on behalf of the U.S. Army Corps of Engineers:
“Concern for the fishing industry prompted sporadic and ineffectual conservation measures. As early as 1877, the legislature of Washington Territory declared a closed season—and the next year Oregon responded with similar measures. State governments also curtailed the use of certain types of fishing gear on the Columbia River. During the late 19th century Washington and Oregon prohibited fish traps, weirs, seines, and nets placed two-thirds of the way across freshwater streams, creeks, or lakes, if they prevented the passage of fish. Purse seines, which consisted of a long curtain of webbing drawn by a boat, were prohibited on the Columbia River in 1917 and in the coastal waters of both Washington and Oregon in 1922. Fishwheels, as noted, were banned in 1926 and 1934. However, many of these restraints were rarely enforced. Although Oregon created a Board of Fish Commissioners in 1887 and Washington established a Fish Commission in 1890, neither organization possessed sufficient funds to police the river and catch offenders. Hence, early restrictions on fishing did little to stop the decline of the salmon runs”.[69]
As also described in the Army Corps of Engineers’ history, in 1926 anglers and gillnetters combined forces to achieve the first ban on fishwheels, traps, and seines in Oregon waters, and Washington State did so in 1934. This was, however, a limited reduction of fishing pressure and was almost too late. By that time the distribution and life history diversity of salmonids was greatly reduced. The current state of the system was very precarious although still in the recoverable historic conditions had the first two Hs had their impacts stopped.
To summarize, the changes in the first two state variables, habitat and harvest, systematically reduced the resilience of the historic regime. The state of the system remained in the historic regime, as a simplified network of relatively undamaged tributaries persisted and the stabilizing feedback provided by the intact, but reduced, metapopulations of salmon provided a reduced level of resilience for this simplified network of tributaries. However, with the reduced resilience of the historic regime, the state of the system had become more vulnerable to a critical transition to the remnant regime.
3.3. Hatcheries and the Techno Regime
The impacts of the third H, hatcheries, began to be felt over 100 years ago as the commercial interests dependent on salmon for their livelihood began to recognize that declining salmon returns presented an unsustainable picture. Salmon peaked in their tonnage canned in the Columbia River Basin in 1883 when 40,911,000 lbs were utilized. Five years later in 1888, the poundage utilized had already declined to 24,211,005 lbs, a drop of 40.8% from the high point [70]. The cannery operators, concerned about declining numbers of highly prized spring chinook salmon, banded together to finance the first salmon hatchery in 1877 on the Clackamas River. As Richard White explains:
“Hatcheries sought to wed technology and biology, to merge factorylike production with natural production. The canners, many fishermen, and many experts on the fisheries came to regard nature as inefficient. H. A. Jones, the federal official who first studied the decline of the Columbia fisheries, produced a rather typical piece of nineteenth-century arithmetic. He calculated that a $10,000 investment in a single hatchery and operating expenses of $3000 a year would produce 5 million mature fish a year, which, when canned, would yield $5 million. This was more than twice the fish and twice the income then being produced on the Columbia. “These figures border on the marvelous”, Jones admitted, but encapsulated in such marvelous numbers was a hope that an inefficient nature, with a helping managerial hand, could solve both a biological and a social crisis”.[71]
Hatcheries are essentially nothing less than fish factories, they were consistent with a laissez-faire attitude towards economic development of the region in general. By providing young fish through an industrial process detached from the spawning tributaries and migration channel, hatcheries made it possible for other economic activities such as water withdrawal for irrigation, logging, grazing of livestock, etc., to continue at an unfettered pace. Politically, hatcheries gave a way to prevent conflicts between different economic sectors, as hatcheries essentially traded concrete ponds for habitat. Hatcheries failed to live up to the unrealistic and unscientific dreams they were initially founded upon [72]. While they did provide more fish to be captured they also diluted the genetics of local populations due to careless mixing of fish across the region and escaped hatchery fish breeding with wild fish, they promoted diseases due to their dense housing, they degraded water quality locally due to their effluents [73], but they gave politicians something positive to point to as economic development in the region and the consequences of hatcheries accelerated.
From the point of view of resilience, it is important to note two novel feedback loops created by the introduction of hatcheries. In the first loop, as native runs decrease, high demand for commercially available fish drives the building of more hatcheries to augment natural production to meet commercial demand. However, flooding the system with hatchery fish can be described as a form of genetic pollution, acting to replace a diverse metapopulation of life histories with a genetically, temporally, and spatially restricted network of populations. This reduction in native fish, in part due to direct competition for resources with hatchery fish, further raises the perceived need to augment with hatchery releases and the cycle continues. This amplifying feedback drives a process that reduces the resilience of the historic regime as it diminishes life history diversity, the foundation of that resilience, and yet another persistent force, driving the system state toward a threshold appears.
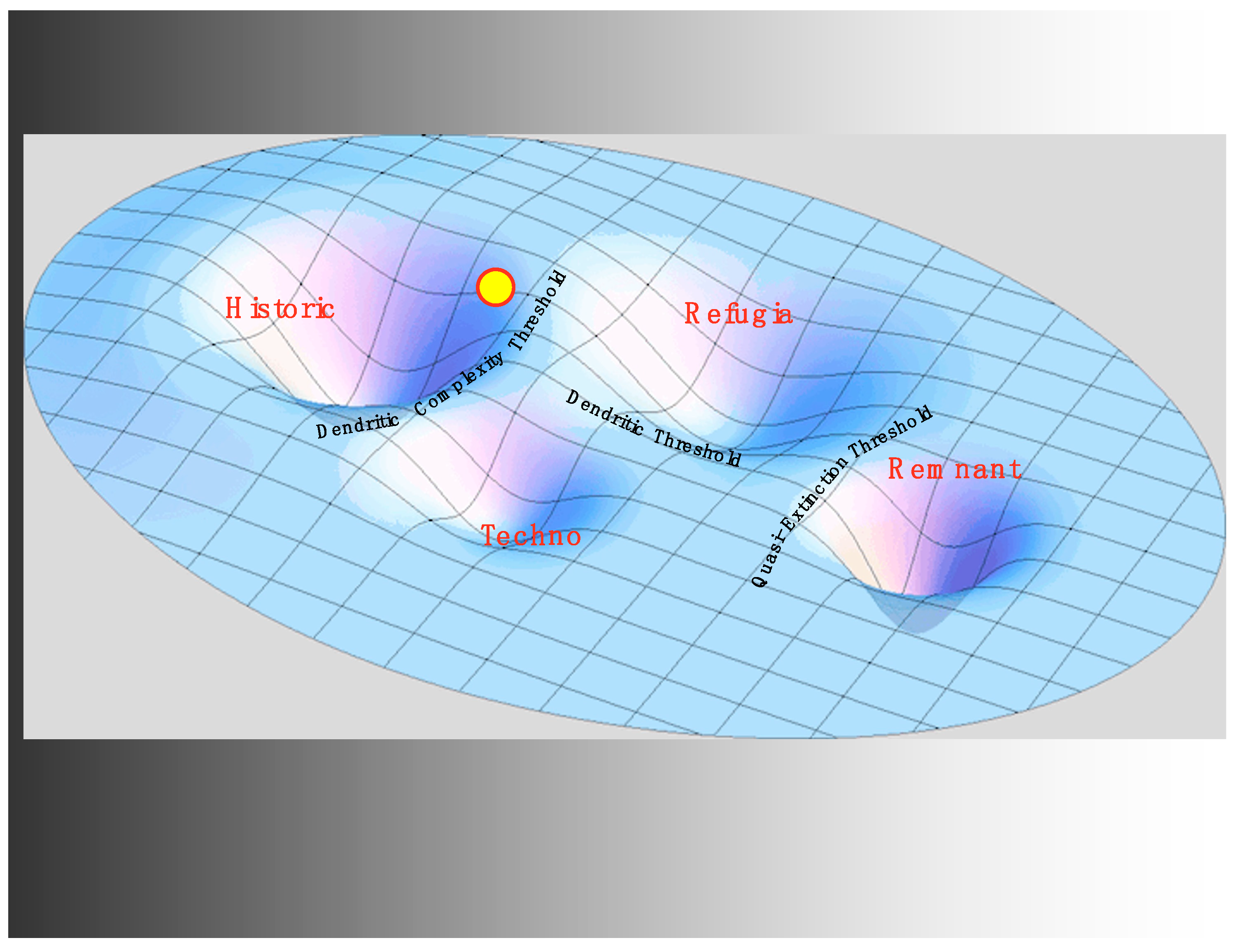
The second novel feedback created by the introduction of hatcheries is a stabilizing feedback in which increased hatchery fish in the river system produce a greater supply of salmon for fishing, and therefore reduce the perceived need for more habitat restoration to support productivity of wild fish. Conversely, increased activism for habitat restoration increases the productivity of native fish, thus reducing the need for hatcheries. Indeed, this feedback is built into federal laws such as the federal Mitchell Act that initially tied funding for hatcheries to fisheries user fees but was amended to generate funding to compensate for habitat degradation in the upper basin with hatchery production in the lower basin. Powerful stabilizing feedback such as this can form a primary source of stabilization for a regime but which regime? We propose that a new regime, the techno regime (see Figure 3), as yet unoccupied by the state of the system, is stabilized by this feedback and the surrounding institutions that support it. This regime is characterized by a river system that can be reasonably well supplied with salmon (in a quantitative sense). While this techno regime would have a substantially reduced level of life history diversity due to hatchery operations, the contribution of additional fish would generate another stabilizing feedback due to support from the sport fishing community. There is ample evidence that the state of the system is not in this regime, but it remains a cautionary possibility. As we will see, it also represents a tempting so-called solution.
In spite of the impact of changes in the state variables (the first three Hs) the current state of the system has remained in the historic regime. The resilience of that regime has certainly been reduced over the course of the past century, making critical transitions to other regimes more likely, but its resilience has remained high enough to prevent a critical transition. Particularly crucial for maintaining that resilience is the modular structure of the remaining wild salmonid metapopulation in the Columbia River Basin, which allowed for recovery from even severe but localized impacts. However, after changes in the remaining state variable, the 4th H, a critical transition will occur and the system state will move to a newly stabilized regime, the refugia regime.
3.4. Hydro and the Critical Transition to the Refugia Regime
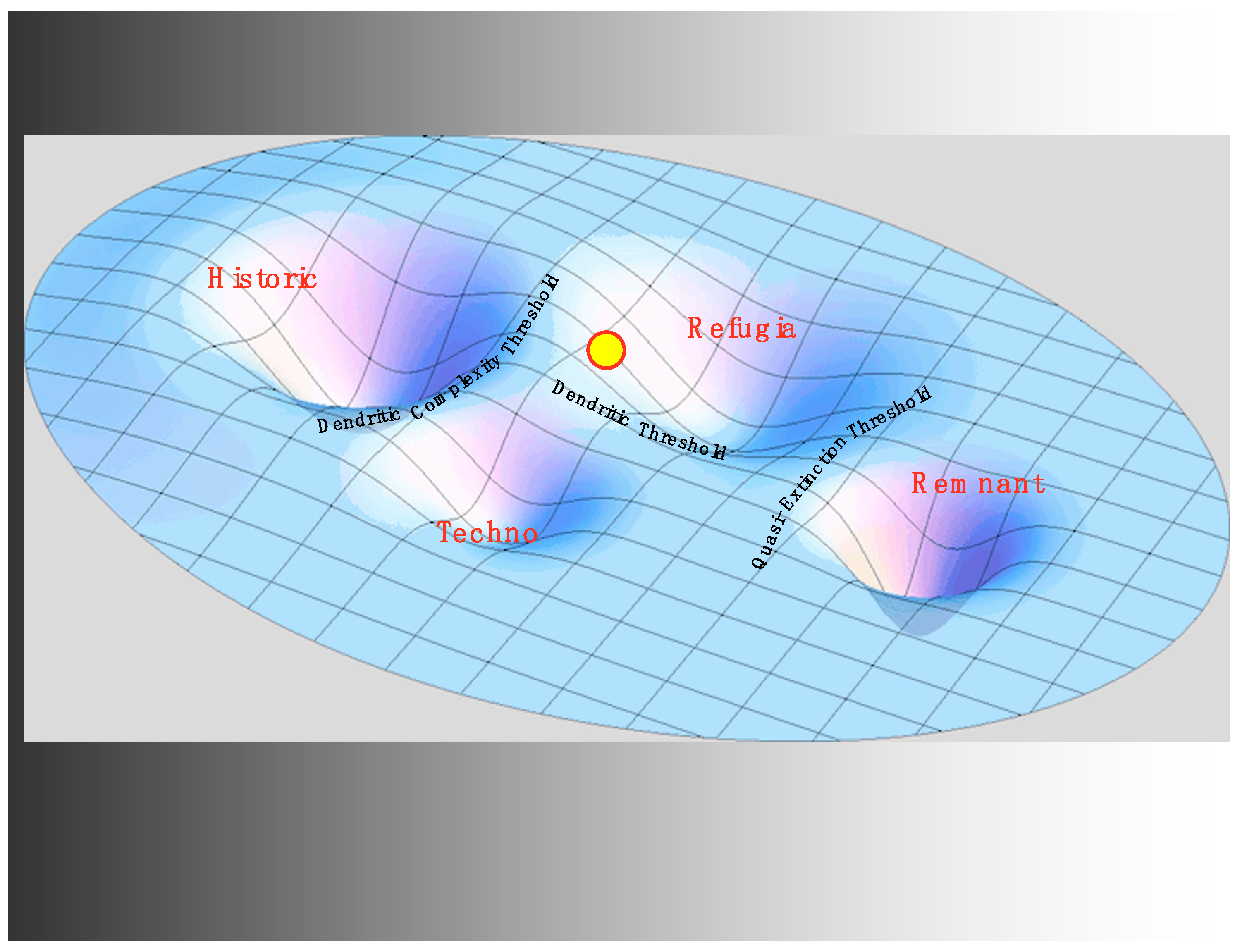
With all the impacts on life history diversity of habitat disruption, harvesting practices, and hatchery development, we see the state of the system in 1930 as still within the historic regime, but precariously close to a threshold. We will argue below that, with the construction of the system of mainstem hydroelectric dams on the Columbia (the 4th H) beginning in the 1930s, a critical transition became inevitable (see Figure 4). However, a critical transition to where? eEvidence is provided below that the transition was to the refugia regime, newly stabilized by feedbacks and institutions emerging from the rise of the U.S. conservation movement and the mandates arising from the economic depression and subsequent institutional shifts.
Many dams have been built on the Columbia River and its tributaries; presently over 270 hydropower dams over one-tenth megawatt in size and 200 for other purposes are in place. The large mainstem dam projects were catapulted into construction by exogenous factors—a global depression that made large-scale public spending on dams and other public works projects a priority. As the Northwest Power and Conservation Council reports [74]:
“For Roosevelt, the huge public works projects that would put people to work and raise America from the Depression were projects of opportunity…Puget Sound Power and Light Company completed Rock Island Dam in 1933… It was the first dam on the mainstem Columbia River, and it was not a multiple-purpose dam. Its sole purpose was to generate electricity. Construction began at Grand Coulee and Bonneville, both multiple-purpose dams, in 1933; Bonneville was completed in 1938 and Grand Coulee in 1941. The last major dams … were Lower Granite on the Snake River and Libby on the Kootenai, both in 1975. There are 14 dams on the mainstem Columbia from Mica to Bonneville, and five on the lower Snake from Hells Canyon Dam to Ice Harbor Dam… Collectively, Columbia River Basin dams have a total nameplate capacity of 36,682.2 megawatts and produce, on average, 16,604.42 megawatts of electricity…”
Some regulatory efforts were made early on to keep dam construction from harming salmonid populations, but these generally took the form of ineffectual or unenforced laws. In a statement of the importance of population diversity and dendritic complexity, contemporary in all but vocabulary, the Oregon Fish Commission quoted its work conducted over eleven years with the United States Bureau of Fisheries in its report of 1931 [75]:
“Knowing further that each race [stock] is self-propagating, it becomes perfectly apparent that all parts of the salmon run in the Columbia River must be given adequate protection if the run as a whole is to be maintained. The protection of only one or two portions of the run will not be sufficient, inasmuch as certain races will be left entirely unprotected.”
William Lang, a historian of the dams of the Columbia River system, identifies dam construction beginning in the 1930s as transformative to the river system when he writes [76]:
“The transformation of the Columbia River began years before 1949… In the spring of 1933… President Franklin D. Roosevelt… directed federal monies at two huge public works projects’ Bonneville Dam near Portland and Grand Coulee Dam in north-central Washington. Roosevelt used public works to address fundamental economic issues by creating employment and stimulating regional development in three of the nation’s river basins, the Tennessee, the Missouri, and the Columbia. What the New Deal began on these rivers changed them dramatically and refashioned the regions they drained. Grand Coulee and Bonneville dams blockaded the Columbia, the second largest river by volume of flow in the United States and the river with the greatest hydroelectric potential on the face of the globe. Roosevelt’s action put the river on a new course, literally and figuratively.”
Most mainstem dams on the Columbia River are roughly 100 feet in height measuring from the water upstream of the dam to the water below it. These dams pose a severe challenge to designing fish ladders that facilitate upstream migration, and even when fish passage exists factors such as temperature effects in impoundments can pose additional challenges to migration [77]. Other dams in the Columbia River system are just too tall and were designed without any attempt at ladders for fish passage. The Grand Coulee Dam rises about 350 feet above the downstream water level. Chief Joseph Dam is almost 200 feet high, and Hells Canyon Dam is over 200 feet high. When the gates closed on Grand Coulee, all upstream salmon passage, including all passage to Canada (the source of 30% of all the water in the river system), abruptly ended. Dworshak Dam rises 300 feet above the water downstream [78]. Grand Coulee, Chief Joseph, Dworshak, and Hells Canyon Dams were built with no provision for upstream passage. One way of describing impassable migration barriers is in terms of river miles:
“Fish passage ends at Chief Joseph Dam at River Mile 545 on the Columbia. Before that dam was completed, fish passage ended at Grand Coulee Dam, 51 miles upriver. In the Snake River system, fish passage ends at Hells Canyon Dam, at River Mile 247, and, on the North Fork Clearwater River, at Dworshak Dam, which is about three miles from the confluence of the North Fork with the mainstem of the Clearwater”.[79]
This description does not capture the dramatic nature of the loss of dendritic complexity of salmon habitat as the dams were constructed. Grand Coulee Dam cut off 645 river miles of upstream habitat, not counting the miles of tributaries that flowed into that stretch of the Columbia River. Adding tributary miles where salmon spawned increases this to roughly 1200 miles of river habitat lost. However this was just the first major loss of habitat, and river miles alone do not express the true loss of complexity. As Sheer and Steel [80] noted, even considering just the lower reaches of the Columbia River Basin:
“We combined existing inventories of barriers to adult fish passage in the Willamette and Lower Columbia River basins and identified 1491 anthropogenic barriers to fish passage blocking 14,931 km of streams. We quantified and compared the stream quality, land cover, and physical characteristics of lost versus currently accessible habitat by watershed, assessed the effect of barriers on the variability of accessible habitats, and investigated potential impacts of habitat reduction on endangered or threatened salmonid populations. The majority of the study watersheds have lost more than 40% of total fish stream habitat. Overall, 40% of the streams with spawning gradients suitable for steelhead (anadromous rainbow trout Oncorhynchus mykiss), 60% of streams with riparian habitat in good condition, and 30% of streams draining watersheds with all coniferous land cover are no longer accessible to anadromous fish… The differences between lost and accessible streams suggest that high-quality habitat has been disproportionately lost in many watersheds.”
Smaller tributary dams were also built, including Little Falls Dam completed in 1911 on the Spokane River at Spokane, which had a questionable fish ladder; Long Lake Dam, with no fish ladder, completed in 1915 four miles further upstream which ended salmon passage in the Spokane River; Sunbeam Dam in 1919 on the Salmon River between Stanley and Challis, Idaho which had a fish ladder but which nonetheless impeded salmon migration significantly, (a fishway was constructed around it in 1919 and the dam was blown up by person or persons unknown in 1934 to help sockeye migrate to Redfish Lake 20 miles upstream); Lewiston Dam on the Clearwater River near Lewiston, Idaho in 1927, which had a fish ladder that was inadequate and which almost ended chinook runs and which dramatically reduced steelhead runs in the Clearwater Basin, it was removed in 1973 as part of another dam project; and Brilliant Dam completed on the Kootenay River in British Columbia in 1944, which blocked salmon from migrating to the Slocan River and Slocan Lake.
Dams also degraded habitat by slowing river flows and therefore causing water temperatures to rise and inundating historically productive shallow spawning habitats. Dam construction had temporal, geographical, thermal, and food web impacts. Thus, the hydropower dams of the Columbia River Basin at first made the current state of the system in the historic regime become more precarious, by making drastic reductions in life history diversity in all three of its dimensions: genetic diversity, dendritic complexity, and temporal diversity. The hydroelectric dams were effectively a series of large amputations of parts of the river system from the perspective of the fish, and they made utilization of the connection corridors harder for much of the rest of the system. The Washington State Department of Fisheries’ publication “The Salmon Crisis” in 1951 featured a photograph of migrating salmon attempting to pass a dam on their way upstream [81] and it says, “Hundreds of miles of spawning habitat have been cut off by the construction of power dams… The Columbia and its tributaries, including the Snake River system, have a growing number of dams which are either total or partial blocks. If all the power projects now contemplated are built, most of the state’s watersheds will be largely reliant upon mechanical fish passage devises and artificial propagation to perpetuate their migratory fish… There is a vast non-productive desert behind dams…”. However, this reduction alone is not enough to claim a critical transition has likely occurred. Recently methods have been developed and employed to argue that critical transitions have occurred. For example, Tomczak et al. [82] used ecological network indicators to argue that critical transitions occurred in the Baltic ecosystem between 1976 and 2006. As well, there has been rapid development of quantitative indicators as “early warning signals” of regime shifts [83,84,85,86]. Information theory has been used to “detect interactions between salmon counts and lagged streamflows, air and water temperatures, precipitation, snowpack, climate indices, and downstream salmon counts” [87]. Given the length and complexity of this work, a comparison of these methods in regard to critical transitions and regimes will be more appropriate for a subsequent paper, especially given the extended time period of human interventions we are presently considering. Rather, we will make three plausibility arguments about transitions and regimes.
We first argue that a critical transition has occurred, without positing an alternative regime. A natural experiment has been underway since the Northwest Power Act created the Northwest Power and Conservation Council to “mitigate the effect of the hydroelectric system on fish and wildlife”. In what may well be the most expensive ecological restoration effort in history, many billions of dollars have been spent to restore native runs of salmon, to no discernible effect. The fact that this intensive and persistent effort has failed to move the system in the desired direction constitutes evidence that the state of the system is now on the other side of a threshold from the historic regime. Given the unprecedented efforts to mitigate the effects of three of the Hs, it would seem that if human activity, in terms of habitat, harvest, and hatcheries, in the region ceased, the system would not now recover to anything resembling previous levels of life history diversity [88,89].
Our second line of argumentation that a critical transition has taken place is that a shift has occurred in the dominant stabilizing feedbacks as seen empirically in other systems [90], and hence a change in the “fundamental structure and feedbacks” that partially define regime identity. As argued above, the stabilizing factors for the historic regime were dependent on high levels of life history diversity and ecologically sensitive fishing practices. While these have substantially weakened or disappeared, we posit that powerful new social-ecological feedback and institutions have replaced them.
The initial impetus for the dams came more from a desire for a stable source of irrigation water, rather than for developing a supply of electricity that (at that time) vastly exceeded the regional demand. Rather than stemming from a desire to manage nature in a way that improved upon its inefficiencies, as the original hatchery advocates sought to achieve, dam construction was originally seen as something that would mimic nature as it had been in the past.
“Interestingly, engineers viewed dams as mimicking nature since some of the largest glacial dams in the world existed in the same area during the Pleistocene epoch; in their view, they were returning it to a previous natural state… Nonetheless, increased damming harmed local wildlife, prevented fish from moving to spawning grounds, overstressed the landscape, and allowed for another and more harmful industry”.[91]
In contrast, the subsequent large-scale development of hydropower in the basin was driven primarily by a desire for economic development in the wake of the Great Depression. In contemporary times, that initial mandate has been institutionalized in several ways that serve to stabilize an alternate regime.
Following the Great Depression, the development of inexpensive and abundant electric power provided by the dams drove rapid economic development and population growth in the basin. The need to protect rapidly growing major cities from flooding (in particular Portland after the Vanport Flood of 1948) [92] has created new and powerful institutional support for a novel regime, one that leaves the dams in place and settles for a simplified river system. This is most vividly represented by the Canada–U.S. Columbia River Treaty, based on an exchange of electric power going from the U.S. hydroelectric system to Canada in exchange for protection from flooding provided by Canadian storage dams created for that purpose.
The path dependence that has developed over the last 70 years of abundant and relatively inexpensive electricity represents another novel institutional factor stabilizing a system with dams and low life history diversity. The perceived tradeoff, and hence balancing feedback, between cheap electricity and restored diversity of salmon runs is seen as a central dilemma in salmon restoration debates [93]. As evidence of the power of the institutional forces that have developed in connection with the hydro systems, we can make comparisons with other rivers in the Pacific Northwest where dam removal has occurred or been agreed to, but where the path dependency of inexpensive electricity is not a factor. As described by Blumm and Erickson [94], “nine significant dams have been removed, and four more—in the contentious Klamath Basin of Oregon and California—are slated for removal in within the next decade”. After what has been indeed decades of contention, the four Klamath Basin dams appear to be about to be removed [95]. In most of these cases, the recovery of salmon runs is a proximal cause of the dam removal and, while there has often been contentious and intense opposition, as in the Klamath Basin, the dams have come down. This points to the powerful stabilizing influence the dams provide for an alternate regime for the Columbia River Basin.
However, why is that alternate regime not the remnant regime? In other words, why are most of the remaining local populations of wild salmon not going extinct? Developing at nearly the same time as the system of dams on the river was the rise of the conservation movement in the U.S. nationally and in the Pacific Northwest regionally. The Endangered Species Act marked a key moment in turning the conservation movement into an institutional force stabilizing a regime between the historic regime and the remnant. Embedded in that law is a stabilizing feedback that increases restoration efforts in response to lowered salmonid populations and, in the interpretation of the National Oceanic and Atmospheric Administration, in response to lowered life history diversity. Regionally, the Northwest Power Act institutionalizes an impressive flow of funding, over one billion dollars annually, directly in response to the welfare of the salmon and their life history diversity. In addition to these powerful stabilizing factors, civil society through a vast array of non-profit organizations provides a similar increase in social capital in response to diminished salmon prospects. With all of these stabilizing influences in place, the refugia regime gains its resilience. This new regime is characterized by a much less extensive modular riverine system, but within that system, a high level of dendritic complexity remains, as well as substantial temporal and genetic diversity, thus a proportionally scaled-down life history diversity. Compared to the historic regime, it relies much more strongly on institutional factors and social-ecological feedback for its resilience. River flows, impeded by the dams, have been replaced with cash flows and population restoration efforts that do not make much headway toward historic population levels but which help stabilize the fish populations that remain.
Thirdly, arguing from a narrative perspective, perhaps the strongest evidence of a critical transition is that the river system has changed its identity as reflected in the writings of those intimately familiar with the river and its life. The following quote excerpts the critical argument of Richard White,
“[The Organic Machine’s] thesis is that the best way to understand the river is as an entity that has been in constant flux. Gradually human beings have modified it. They have created the illusion of conquering the river, of turning it, as the common phrase is in the Pacific Northwest, into a series of slackwater lakes. We apply social language to the river. We have raped it or killed it; but such language is deceptive. We have changed the Columbia to the detriment of some species and the benefit of others. Where once the Columbia said salmon, it now says shad and squawfish. The Columbia is not dead. The dams depend on larger natural rhythms of snowfall and snowmelt, of rain and gravity and seasons, but we have created a system where what is natural and what is human becomes harder and harder to distinguish. Each intrudes on and influences the other. The river has become an organic machine”.[96]
This is echoed by an assessment written by Lackey, Lach, and Duncan [97]:
“Many of the region’s environments have been permanently altered in ways that do not favor salmon. The Columbia Basin, for example, is now dominated by a series of mainstem and tributary reservoirs. Land use in much of the watershed has also changed in ways that no longer favor salmon…As dramatic as the environmental changes are, some fishes, especially exotics, are thriving (e.g., walleye, American shad… smallmouth bass… northern pike… and brook trout…). These exotic species are well adapted to the new environment. It would be difficult—some argue impossible—to re-create the Pacific Northwest habitats that once existed and were ideal for wild salmon. A simpler, cheaper policy option would be to manage for those fishes, typically exotics, best suited to current habitat. Such an approach, while relatively easy and cheap to accomplish, would be an explicit decision to terminate many stocks of wild salmon.”
4. Discussion
Does it make a difference, in terms of policy and practice, in which regime the state of the salmon-centered system resides? We argue that it does and that the mismatch of policy and regime status is crucial to the effectiveness of policy.
Although we have described feedback and institutional forces that give the refugia regime substantial resilience, they are tenuous since with feedback that depends on social factors that can change rapidly, the danger of movement of the system state from the refugia regime to the remnant regime remains very real. Salmon are partially oceanic organisms and there are large-scale regimes of ocean conditions such as the El Niño/Southern Oscillation and the Pacific Decadal Oscillation (ENSO and PDO) [98] as well as regional regimes such as the North Pacific Gyre Oscillation (NPGO) that fluctuate temporally and which will move directionally in the future as global climate changes [99]. These ocean conditions will also impact in-stream flows in the Columbia Basin [100]. A failure to account for fluctuating but temporarily hospitable ocean conditions contributed to the lack of recognition of the implications of habitat loss in northwest salmon management efforts [101]. Moreover, the climatic impacts felt inland and in smaller tributaries, as well as the migration channel, will synergize with oceanic regime changes and further increase the precariousness of the state of the system in the refugia regime [102]. No adjustments made in hatcheries or dams or harvest could compensate for a substantial degradation of ocean conditions, let alone one that occurred due to climate change and coincided with a diminution of salmon survival due to warming conditions inland. The forthcoming climate challenges to resiliency identified for salmon populations in California, at the southern end of the range of these fish and similarly impacted by human activities in many ways to salmon in the Pacific Northwest, provides a cautionary glimpse of a warming future [103]. Much remains to be done to understand how climate change and system resilience will interact. A framework proposed for studying climate resilience in fisheries proposes 38 attributes of importance, but notes that [104] “Overall, however, we find few studies that test resilience attributes in fisheries across all parts of the system… meaningful quantification of the attributes’ contributions to resilience remains a challenge.”
Human population levels in the Pacific Northwest will certainly increase [105] leading to an unpredictable degree of increased development and habitat alteration. Much of what happens next will be based on human choices about what they are willing to pay for energy, the extent to which regulations on the use of privately owned land will be tolerated, how much salmon will be harvested and by who, what will our diet and transportation systems look like, and so forth [106]. Existing, and doubtless forthcoming, threats to the stability of the system state in the refugia regime are part of a broader policy debate. The simplified refugia regime provides substantially less protection from various changing conditions than the richer and more extensive historical regime.
If the system occupied the refugia regime and the intentions were to restore it to historic conditions, a transformative approach would be called for; incremental approaches would be insufficient. In this regard, it is interesting to consider the subbasin planning process conducted by the Northwest Power and Conservation Council (NWPCC), perhaps the largest participatory planning effort for ecological restoration in U.S. history. Central to the efforts in the second round of subbasin planning (beginning in 2000 and completed in 2005) was the use of computer decision support systems to inform decisions made by planning participants. The NWPCC adopted a particular system, the Ecosystem Diagnosis and Treatment (EDT) approach, for use by the subbasin councils to develop plans and, based on those plans, to guide funding to the individual subbasins. The planning process was described in an NWPCC document [107] as “Subbasin plans will be developed locally and in collaboration with fish and wildlife managers, local governments, interest groups and stakeholders and other state and federal land and water resources managers” and EDT is described as “The Council has developed Ecosystem Diagnosis and Treatment (EDT) to use as a subbasin planning tool and is providing planners with preliminary data sets to use for this tool. EDT is able to link habitat condition to responses in salmon… populations…”. However, EDT incorporates 45 in-stream habitat variables, an impenetrable number to a nontechnical stakeholder, and therefore preferentially empowers quantitatively oriented participants familiar with the EDT software.
Critical examination reveals that the EDT approach limits participation and focuses influence on more technologically enabled actors. By being entirely focused on in-stream variables, EDT points the discussion away from broader issues of system connectivity and its relationship to land use and decisions about economic development [108]. While the subbasin councils represented local stakeholders brought together for the purpose of planning, the adoption of EDT framed the discussion and decision-making in a way that favored incremental changes [109] from business as usual. If it is correct that the system now occupies an alternative regime from the historic, such planning methods are unlikely to return the system to anything approaching historic conditions.
However, there are policy approaches intentionally tailored to the assumption that the system is in refugia conditions. The Wild Salmon Center (WSC), a research and advocacy NGO based in Portland Oregon, has developed a plan to preserve and enhance a modular network of “salmon sanctuaries”, as the best approach to prevent further decline and prepare for future impacts such as climate change [110,111]. An implicit assumption of this approach is that the system does indeed occupy the refugia regime, that full recovery to the historic regime is currently unlikely, and that current approaches are not building needed resilience for the refugia regime. Branding their approach the “Salmon Stronghold Initiative”, the center has developed collaborative partnerships (the North America Salmon Stronghold Partnership) and introduced federal legislation (the Stronghold Conservation Act). This act was introduced in 2008 and 2011, the legislation made it as far as being reported out of committee favorably, it would represent another social-ecological institution adding resilience to the refugia regime [112]. Once again, a version of a salmon stronghold bill was introduced in 2022 [113]. Whether the political and social will for this to ever become U.S. law exists remains to be seen.
Increasing recognition of the status and importance of Native Americans in the Columbia River Basin provides another avenue of support for protection and perhaps recovery of the system. The Columbia River Inter-Tribal Fish Commission (CRITFC) has a project entitled Spirit of the Salmon: Wy-Kan-Ush-Mi Wa-Kish-Wit [114] whose goals are listed as:
- Restore anadromous fishes to the rivers and streams that support the historical, cultural, and economic practices of the tribes. (These are generally areas above Bonneville Dam.)
- Emphasize strategies that rely on natural production and healthy river systems to achieve this goal.
- Protect tribal sovereignty and treaty rights.
- Reclaim the anadromous fish resource and the environment on which it depends for future generations.
The CRITFC plan, originating in 1995 with a 2014 update, includes detailed scientifically- and culturally-based technical recommendations in 20 areas (land-use practices, water quality, watershed restoration, supplementation, reintroductions, juvenile fish migration, estuary, harvest ceilings, adult fish migration, water quality, harvest management (in-river), lamprey passage, white sturgeon, predation, mark selective fisheries, restoring fish passage, invasive species, climate change, update “lifecycle model”, and Endangered Species Act delisting). The unique legal status and expertise of the Tribes, and the twin legal principles of Trust Responsibility and Government-to-Government Consultation give the CRITFC technical recommendations a special status for helping to protect the river system. Combined this may all help maintain the system in a refugia regime rather than a remnant regime.
The CRITFC plan is based on an indigenous understanding of relationships, and given the increasing social recognition of the importance of indigenous perspectives, it may be that ultimately this helps provide a unique stabilizing social feedback helping to protect the river system and its inhabitants. Spirit of the Salmon: Wy-Kan-Ush-Mi Wa-Kish-Wit describes a creation story:
“When the Creator was preparing to bring humans onto the earth, He called a grand council of all the animal people, plant people, and everything else. In those days, the animals and plants were more like people because they could talk. He asked each one to give a gift to the humans—a gift to help them survive, since humans were pitiful and would die without help. The first to come forward was Salmon. He gave the humans his body for food. The second to give a gift was Water. She promised to be the home to the salmon. After that, everyone else gave the humans a gift, but it was special that the first to give their gifts were Salmon and Water. When the humans finally arrived, the Creator took away the animals’ power of speech and gave it to the humans. He told the humans that since the animals could no longer speak for themselves, it was a human responsibility to speak for the animals. To this day, Salmon and Water are always served first at tribal feasts to remember the story and honor the First Foods.”
The Upper Columbia United Tribes (UCUT) and First Nations in Canada work jointly to promote the reintroduction of migratory salmonids upstream of Chief Joseph and Grand Coulee dams. Their joint paper “A Vision for Salmon and Steelhead: Goals to Restore Thriving Salmon and Steelhead to the Columbia River Basin: Phase 2 Report of the Columbia Basin Partnership Task Force of the Marine Fisheries Advisory Committee” of 2020 [115] lays out a detailed strategy. This is another social force that supports, reinforces, and perhaps eventually can expand the refugia regime that currently exists.
Whether the system can ever be restored to the historic regime is another matter entirely. It may be that by building the resilience of the refugia regime, the state of the system is put in a position to take advantage of a window of opportunity to make a transition to the historic regime. Clearly, the powerful institutional stabilizing effects of the dams would need to be greatly weakened to make such a transition possible. Those effects are closely linked to the supply of inexpensive, plentiful, and low-carbon electric power production supplied by the dams. If a technological innovation were to supplant that dependency with an alternate source of power or decrease in demand, perhaps such a window of opportunity would open.
Author Contributions
All authors have read and agreed to the published version of the manuscript. The authors contributed equally to all phases of this article.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We would like to thank three anonymous reviewers whose comments allowed us to improve this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Williams, R.N. Return to the River: Restoring Salmon to the Columbia River; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2006; pp. 1–720. [Google Scholar]
- Folke, C. Resilience (Republished). Ecol. Soc. 2016, 21, 44. [Google Scholar] [CrossRef]
- Gustafson, R.G.; Waples, R.S.; Myers, J.M.; Weitkamp, L.A.; Bryant, G.J.; Johnson, O.W.; Hard, J.J. Pacific salmon extinctions: Quantifying lost and remaining diversity. Conserv. Biol. 2007, 21, 1009–1020. [Google Scholar] [CrossRef]
- Dittmer, K. Changing streamflow on Columbia basin tribal lands—Climate change and salmon. Clim. Chang. 2013, 120, 627–641. [Google Scholar] [CrossRef]
- Wolf, E.C. Salmon Nation: People, Fish, and Our Common Home, 2nd ed.; Oregon State Univeristy Press: Corvallis, OR, USA, 2003; pp. 1–92. [Google Scholar]
- Folke, F.; Carpenter, C.; Walker, W.; Scheffer, M.; Chapin, T.; Rockström, J. Resilience thinking: Integrating resilience, adaptability and transformability. Ecol. Soc. 2010, 15, 20. [Google Scholar] [CrossRef]
- Baird, S. Salmon fisheries of Oregon. The Oregonian, 3 March 1875. [Google Scholar]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social–ecological systems. Ecol. Soc. 2004, 9, 5. Available online: http://www.ecologyandsociety.org/vol9/iss2/art5/ (accessed on 27 February 2023). [CrossRef]
- Westley, F.R.; Tjornbo, O.; Schultz, L.; Olsson, P.; Folke, C.; Crona, B.; Bodin, Ö. A theory of transformative agency in linked social-ecological systems. Ecol. Soc. 2013, 18, 27. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Folke, C.; Kofinas, G.P. (Eds.) A framework for understanding change. In Principles of Ecosystem Stewardship; Springer: New York, NY, USA, 2009; pp. 3–28. [Google Scholar]
- Sendzimir, S.; Magnuszewski, M.; Flachner, F.; Balogh, B.; Molnar, G.; Sarvari, A.; Nagy, Z. Assessing the resilience of a river management regime: Informal learning in a shadow network in the Tisza River Basin. Ecol. Soc. 2008, 13, 11. Available online: http://www.ecologyandsociety.org/vol13/iss1/art11/ (accessed on 27 February 2023). [CrossRef]
- Ludwig, D.; Jones, D.; Holling, S. Qualitative Analysis of Insect Outbreak Systems: The Spruce Budworm and Forest. J. Anim. Ecol. 1978, 47, 315–332. [Google Scholar] [CrossRef]
- Carpenter, C.; Walker, W.; Anderies, A.; Abel, A. From metaphor to measurement: Resilience of what to what? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Rinaldi, S.; Scheffer, M. Geometric analysis of ecological models with slow and fast processes. Ecosystems 2000, 3, 507–521. [Google Scholar] [CrossRef]
- Smith, D.M.S.; Abel, N.; Walker, B.; Chapin, F.S., III. Drylands: Coping with Uncertainty, Thresholds, and Changes in State. In Principles of Ecosystem Stewardship; Chapin, F.S., III, Folke, C., Kofinas, G.P., Eds.; Springer: New York, NY, USA, 2009; pp. 171–195. [Google Scholar]
- Folke, F.; Carpenter, C.; Elmqvist, E.; Gunderson, G.; Holling, H.; Walker, W. Resilience and sustainable development: Building adaptive capacity in a world of transformations. Ambio 2002, 31, 437–440. [Google Scholar] [CrossRef]
- Neville, H.M.; Isaak, D.J.; Dunham, J.B.; Thurow, F.R.; Rieman, B.E. Fine-scale natal homing and localized movement as shaped by sex and spawning habitat in Chinook salmon: Insights from spatial autocorrelation analysis of individual genotypes. Mol. Ecol. 2006, 15, 4589–4602. [Google Scholar] [CrossRef]
- Webb, W.; Bodin, B. A Network Perspective on Modularity and Control of Flow in Robust Systems. In Complexity Theory for a Sustainable Future; Norburg, N., Cumming, C., Eds.; Columbia Press: New York, NY, USA, 2008; pp. 85–118. [Google Scholar]
- Levin, S.A.; Lubchenco, J. Resilience, robustness, and marine ecosystem-based management. BioScience 2008, 58, 27–32. [Google Scholar] [CrossRef]
- Walker, B.; Salt, D. Resilience Thinking: Sustaining Ecosystems and People in a Changing World; Island Press: Washington, DC, USA, 2006; 192p. [Google Scholar]
- Walker, B.; Salt, D. Resilience Practice: Building Capacity to Absorb Disturbance and Maintain Function; Island Press: Washington, DC, USA, 2012; 248p. [Google Scholar]
- Biggs, R.; Schlüter, M.; Biggs, D.; Bohensky, E.L.; BurnSilver, S.; Cundill, G.; Dakos, V.; Daw, T.M.; Evans, L.S.; Kotschy, K.; et al. Toward principles for enhancing the resilience of ecosystem services. Annu. Rev. Environ. Resour. 2012, 37, 421–448. [Google Scholar] [CrossRef]
- Kharrazi, A.; Yu, Y.; Jacob, A.; Vora, N.; Fath, B.D. Redundancy, Diversity, and Modularity in Network Resilience: Applications for International Trade and Implications for Public Policy. Curr. Res. Environ. Sustain. 2020, 2, 100006. Available online: https://www.sciencedirect.com/science/article/pii/S2666049020300049?via%3Dihub (accessed on 9 March 2023). [CrossRef]
- Lichatowitch, J. Salmon Without Rivers: A History of the Pacific Salmon Crisis; Island Press: Washington, DC, USA, 2001; 336p. [Google Scholar]
- Wright, S. Evolution in Mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef]
- Wright, S. The roles of mutation, inbreeding, crossbreeding and selection in evolution. In Proceedings of the 6th International Congress of Genetics, Ithaca, NY, USA, 24–31 August 1932; pp. 356–366. Available online: http://www.esp.org/books/6th-congress/facsimile/contents/6th-cong-p356-wright.pdf (accessed on 27 February 2023).
- Wright, S.W. The shifting balance theory and macroevolution. Annu. Rev. Genet. 1982, 16, 1–19. [Google Scholar] [CrossRef]
- Kinzig, A.P.; Ryan, P.; Etienne, M.; Allison, H.; Elmqvist, T.; Walker, B.H. Resilience and regime shifts: Assessing cascading effects. Ecol. Soc. 2006, 11, 20. Available online: http://www.ecologyandsociety.org/vol11/iss1/art20/ (accessed on 27 February 2023). [CrossRef]
- Rietkerk, M.; Dekker, S.C.; de Ruiter, P.C.; van de Koppel, J. Self-organized patchiness and catastrophic shifts in ecosystems. Science 2004, 305, 1926–1929. [Google Scholar] [CrossRef]
- Erős, E.; Olden, O.; Schick, S.; Schmera, S.; Fortin, F. Characterizing connectivity relationships in freshwaters using patch-based graphs. Landsc. Ecol. 2012, 27, 303–317. [Google Scholar] [CrossRef]
- Campbell, G.C. Structural complexity, movement bias, and metapopulation extinction risk in dendritic ecological networks. J. N. Am. Benthol. Soc. 2011, 30, 252–258. [Google Scholar] [CrossRef]
- Yeakel, J.D.; Moore, J.W.; Guimarães, P.R.; Aguiar, M.A.M. Synchronisation and stability in river metapopulation networks. Ecol. Lett. 2014, 17, 273–283. [Google Scholar] [CrossRef]
- Mari, L.; Casagrandi, R.; Bertuzzo, E.; Rinaldo, A.; Gatto, M. Metapopulation persistence and species spread in river networks. Ecol. Lett. 2014, 17, 426–434. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/ele.12242 (accessed on 27 February 2023). [CrossRef]
- Ulanowicz, R.E.; Holt, R.D.; Barfield, M. Limits on ecosystem trophic complexity: Insights from ecological network analysis. Ecol. Lett. 2014, 17, 127–136. [Google Scholar] [CrossRef]
- Waples, R.S.; Pess, G.R.; Beechie, T. Evolutionary history of Pacific salmon in dynamic environments. Evol. Appl. 2008, 1, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Fagan, F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Letcher, B.H.; Schueller, P.; Bassar, R.D.; Nislow, K.H.; Coombs, J.A.; Sakrejda, K.; Morrissey, M.; Sigourney, D.B.; Whiteley, A.R.; O’Donnell, M.J.; et al. Robust estimates of environmental effects on population vital rates: An integrated capture–recapture model of seasonal brook trout growth, survival and movement in a stream network. J. Anim. Ecol. 2015, 84, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Gende, S.M.; Edwards, R.T.; Willson, M.F.; Wipfli, M.S. Pacific Salmon in Aquatic and Terrestrial Ecosystems. BioScience 2002, 52, 917–928. [Google Scholar] [CrossRef]
- Hocking, M.D.; Reimchen, T.E. Salmon-derived nitrogen in terrestrial invertebrates from coniferous forests of the Pacific Northwest. BMC Ecol. 2002, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, C.J.; Kunze, M.D.; Murota, T.; Sibatani, A. Pacific salmon carcasses: Essential contributions of nutrients and energy for aquatic and terrestrial ecosystems. Fisheries 1999, 24, 6–15. [Google Scholar] [CrossRef]
- Gresh, T.; Lichatowich, J.; Schoonmaker, P. An estimation of historic and current levels of salmon production in the Northeast Pacific ecosystem: Evidence of a nutrient deficit in the freshwater systems of the Pacific Northwest. Fisheries 2000, 25, 15–21. [Google Scholar] [CrossRef]
- Willson, M.F.; Halupka, K.C. Anadromous fish as keystone species in vertebrate communities. Conserv. Biol. 1995, 9, 489–497. [Google Scholar] [CrossRef]
- Hocking, M.D.; Reynolds, J.D. Impacts of Salmon on Riparian Plant Diversity. Science 2011, 331, 1609–1612. [Google Scholar] [CrossRef]
- Spencer, C.N.; McClelland, B.R.; Stanford, J.A. Shrimp stocking, salmon collapse, and eagle displacement. BioScience 1991, 41, 14–21. [Google Scholar] [CrossRef]
- Brannon, E.; Powell, M.; Quinn, T.; Talbot, A. Population Structure of Columbia River Basin Chinook Salmon and Steelhead Trout; Project No. 1998-00403, BPA Report DOE/BP-08319-1; Bonneville Power Administration: Portland, OR, USA, 2002; pp. 1–181. [Google Scholar]
- Richter, A.; Kolmes, S.A. Maximum temperature limits for Chinook, coho, and chum salmon, and steelhead trout in the Pacific Northwest. Rev. Fish. Sci. 2005, 13, 23–49. [Google Scholar] [CrossRef]
- Wasowski, R.; Alexander, D.; Kolmes, S.; Banet, N.; Card, A.; Haines, M.; Kolmes, E.; Northcutt, K.; Roemer, D.; Webber, S. A multi-year longitudinal study of water quality parameters in four salmon-bearing and recreational streams on mount hood, Oregon. PLoS ONE 2013, 8, e70453. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, P.; Backman, T.; Busak, C.; Heppell, S.; Kolmes, S.; Maule, A.; Myers, J.; Rawding, D.; Shively, D.; Steel, A.; et al. Interim Report on Viability Criteria for Willamette and Lower Columbia Basin Salmonids; National Marine Fisheries Service: Woods Hole, MA, USA; Northwest Fisheries Science Center: Seattle, WA, USA, 2003; pp. 1–331. [Google Scholar]
- Myers, J.; Busack, C.; Rawding, D.; Marshall, A.; Teel, D.; Van Doornik, D.M.; Maher, M.T. Historical Population Structure of Pacific Salmonids in the Willamette River and Lower Columbia River Basins; NMFS-NWFSC-73; U.S. Department of Commerce, NOAA Technical Memorandum: Washington, DC, USA, 2006; 311p. [Google Scholar]
- Gardner, C.A.; Scott, W.E.; Major, J.J.; Pierson, T.C. Mount Hood—History and Hazards of Oregon’s Most Recently Active Volcano; U.S. Geological Survey Fact Sheet, 2000. 060-00 Online Version 1.0. Available online: http://geopubs.wr.usgs.gov/fact-sheet/fs060-00/ (accessed on 27 February 2023).
- Waples, R.; Beechie, T.; Pess, G.R. Evolutionary History, Habitat Disturbance Regimes, and Anthropogenic Changes: What Do These Mean for Resilience of Pacific Salmon Populations? Ecol. Soc. 2009, 14, 3. Available online: http://www.ecologyandsociety.org/vol14/iss1/art3/ (accessed on 16 March 2023). [CrossRef]
- Butler, V.; O’Connor, J. 9000 years of salmon fishing on the Columbia River, North America. Quat. Res. 2004, 62, 1–8. [Google Scholar] [CrossRef]
- Hamlet, A.F.; Lettenmaier, D.P. Columbia River streamflow forecasting based on ENSO and PDO climate signals. J. Water Resour. Plan. Manag. 1999, 125, 333–341. [Google Scholar] [CrossRef]
- Taylor, J.E. Making Salmon: An Environmental History of the Northwest Fisheries Crisis; University of Washington Press: Seattle, WA, USA, 2001; pp. 1–488. [Google Scholar]
- O’Connor, O. The evolving landscape of the Columbia River Gorge: Lewis and Clark and cataclysms on the Columbia. Or. Hist. Soc. 2004, 105, 390–421. [Google Scholar] [CrossRef]
- Kramer, A.M.; Berec, L.; Drake, J.M. Allee effects in ecology and evolution. J. Anim. Ecol. 2018, 87, 7–10. Available online: https://besjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2656.12777 (accessed on 8 March 2023). [CrossRef] [PubMed]
- Greene, G.; Hall, H.; Guilbault, G.; Quinn, Q. Improved viability of populations with diverse life-history portfolios. Biol. Lett. 2010, 6, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Lichatowitch, J. Salmon, People, and Place: A Biologist’s Search for Salmon Recovery; Oregon State University Press: Corvallis, OR, USA, 2013; pp. 1–272. [Google Scholar]
- Miller, R.R. Is the Past Present? Historical Splash-Dam Mapping and Stream Disturbance Detection in the Oregon Coastal Province. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2010; pp. 1–110. Available online: https://ir.library.oregonstate.edu/concern/parent/4m90f001t/file_sets/dj52w8373 (accessed on 2 March 2023).
- Atlas, W.I.; Ban, N.C.; Moore, J.W.; Tuohy, A.M.; Greening, S.; Reid, A.J.; Morven, N.; White, E.; Housty, W.G.; Housty, J.A.; et al. Indigenous Systems of Management for Culturally and Ecologically Resilient Pacific Salmon (Oncorhynchus spp.) Fisheries. BioScience 2021, 71, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Cosens, B.; Fremier, A. Assessing System Resilience and Ecosystem Services in Large River Basins: A Case Study of the Columbia River Basin. Ida. Law Rev. 2014, 51, 91–125. [Google Scholar]
- Middleton, B.A. (Ed.) The flood pulse concept in wetland restoration. In Flood Pulsing in Wetlands: Restoring the Natural Hydrological Balance; John Wiley & Sons: New York, NY, USA, 2002; pp. 1–10. [Google Scholar]
- Hulse, D.; Gregory, S.; Baker, J. Willamette River Basin Atlas; Oregon State University Press: Corvallis, OR, USA, 2002; pp. 1–178. Available online: https://oregonexplorer.info/content/willamette-river-basin-atlas?topic&ptopic (accessed on 3 March 2023).
- Rohe, R. The Chinese and Hydraulic Mining in the Far West. Min. Hist. Assoc. Annu. 1994, 1, 73–91. Available online: https://www.mininghistoryassociation.org/Journal/MHJ-v1-1994-Rohe.pdf (accessed on 8 March 2023).
- Kareiva, P.; Marvier, M.; Silliman, B. (Eds.) Uncomfortable questions and inconvenient data in conservation science. In Effective Conservation Science: Data Not Dogma; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Tierney, K.B.; Sampson, J.L.; Ross, P.S.; Sekela, M.A.; Kennedy, C.J. Salmon Olfaction is Impaired by an Environmentally Realistic Pesticide Mixture. Environ. Sci. Technol. 2008, 42, 4996–5001. [Google Scholar] [CrossRef]
- Parman, D.L. Inconstant Advocacy: The Erosion of Indian Fishing Rights in the Pacific Northwest, 1933–1956. Pac. Hist. Rev. 1984, 53, 163–189. [Google Scholar] [CrossRef]
- University of Washington Libraries Digital Collection. Salmon in the Pacific Northwest and Alaska Collection, 1890–1961; University of Washington Libraries Digital Collection: Seattle, WA, USA, 2001; Available online: https://content.lib.washington.edu/salmonweb/index.html (accessed on 7 March 2023).
- Mighetto, L.; Ebel, W.J. Saving the Salmon: A History of the U.S. Army Corps of Engineers’ Efforts to Protect Anadromous Fish on the Columbia and Snake Rivers; Historical Research Associates, The Division: Missoula, MT, USA, 1994; pp. 1–253. Available online: https://usace.contentdm.oclc.org/digital/collection/p16021coll4/id/96/ (accessed on 7 March 2023).
- Report of the Commissioner—United States Commission of Fish and Fisheries . 1895, pp. 1–1979. Available online: https://archive.org/stream/reportcommissio06statgoog/reportcommissio06statgoog_djvu.txt (accessed on 8 March 2023).
- White, R. The Organic Machine: The Remaking of the Columbia River; Hill and Wang Critical Issues/MacMillan: New York, NY, USA, 1995; pp. 1–130. [Google Scholar]
- Terui, A.; Urabe, H.; Senzaki, H.; Nishizawa, B. Intentional release of native species undermines ecological stability. Proc. Natl. Acad. Sci. USA 2023, 120, e2218044120. [Google Scholar] [CrossRef] [PubMed]
- Kolmes, S.A. Salmon Farms and Hatcheries. Environ. Sci. Policy Sustain. Dev. 2004, 46, 40–43. [Google Scholar]
- Northwest Power and Conservation Council. Dams: History and Purpose. Available online: https://www.nwcouncil.org/reports/columbia-river-history/damshistory/ (accessed on 8 March 2023).
- Fish Commission of the State of Oregon. Biennial Report of the Fish Commission of the State of Oregon. 1931, pp. 1–33. Available online: https://ir.library.oregonstate.edu/concern/technical_reports/r207tt794 (accessed on 8 March 2023).
- Lang, W.T. 1949: Year of Decision on the Columbia River. Columbia Mag. 2005, 19, 8–15. Available online: https://www.washingtonhistory.org/wp-content/uploads/2020/04/1949-1.pdf (accessed on 8 March 2023).
- Caudill, C.C.; Keefer, M.L.; Clabough, T.S.; Naughton, G.P.; Burke, B.J.; Peery, C.A. Indirect Effects of Impoundment on Migrating Fish: Temperature Gradients in Fish Ladders Slow Dam Passage by Adult Chinook Salmon and Steelhead. PLoS ONE 2013, 8, e85586. [Google Scholar] [CrossRef] [PubMed]
- Whitney, W.; Coutant, C.; Mundy, M.; Williams, W. Mitigation of salmon losses due to hydroelectric development. In Return to the River, Restoring Salmon to the Columbia River; Williams, R.N., Ed.; Elsevier Academic Press: New York, NY, USA, 2006; pp. 325–416. [Google Scholar]
- Northwest Power and Conservation Council. Fish Passage at Dams. Available online: https://www.nwcouncil.org/reports/columbia-river-history/fishpassage/ (accessed on 8 March 2023).
- Sheer, M.B.; Steel, E.A. Lost Watersheds: Barriers, Aquatic Habitat Connectivity, and Salmon Persistence in the Willamette and Lower Columbia River Basins. Trans. Am. Fish. Soc. 2006, 135, 1654–1669. Available online: https://www.fs.usda.gov/pnw/lwm/aem/docs/steel/2006_sheer_and_steel_lost_watersheds.pdf (accessed on 8 March 2023). [CrossRef]
- Washington Department of Fisheries. The Salmon Crisis. 1951, pp. 1–14. Available online: https://digitalcollections.lib.washington.edu/digital/collection/salmon/id/421 (accessed on 9 March 2023).
- Tomczak, M.T.; Heymans, J.J.; Yletyinen, J.; Niiranen, S.; Otto, S.A.; Blenckner, T. Ecological network indicators of ecosystem status and change in the Baltic Sea. PLoS ONE 2013, 8, e75439. [Google Scholar] [CrossRef] [PubMed]
- Brock, W.A.; Carpenter, S.R. Early Warnings of Regime Shift When the Ecosystem Structure Is Unknown. PLoS ONE 2012, 7, e45586. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef]
- Dakos, V.; Carpenter, S.R.; Brock, W.A.; Ellison, A.M.; Guttal, V.; Ives, A.R.; Kéfi, S.; Livina, V.; Seekell, D.A.; van Nes, E.H.; et al. Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PLoS ONE 2012, 7, e41010. [Google Scholar] [CrossRef]
- Biggs, R.; Carpenter, S.R.; Brock, W.A. Turning back from the brink: Detecting an impending regime shift in time to avert it. Proc. Natl. Acad. Sci. USA 2009, 106, 826–831. [Google Scholar] [CrossRef]
- Goodwell, A.; Campbell, N. An information theory-based approach to characterize drivers of upstream salmon migration. PLoS ONE 2022, 17, e0269193. [Google Scholar] [CrossRef]
- Lackey, R.T.; Lach, D.H.; Duncan, S.L. (Eds.) Chapter 3. Wild Salmon in Western North America: Forecasting the Most Likely Status in 2100. In Salmon 2100: The Future of Wild Pacific Salmon; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 57–70. [Google Scholar]
- Lackey, R.T. Saving wild salmon: A 165 year policy conundrum. In Proceedings of the Dubach Workshop: Science and Scientists in the Contemporary Policy Process, Oregon State University, Portland, OR, USA, 3–4 October 2013; Available online: http://forestpolicypub.com/wp-content/uploads/2014/01/165-Year-Salmon-Policy-Conundrum-R-T-Lackey.pdf (accessed on 16 March 2022).
- Biggs, R.; Peterson, G.D.; Rocha, J.C. The Regime Shifts Database: A framework for analyzing regime shifts in social-ecological systems. Ecol. Soc. 2018, 23, 9. [Google Scholar] [CrossRef]
- Newberry, D. The Organic Machine: A Discussion of Human Relationships with Nature. Built Environment Plus. 2017. Available online: https://builtenvironmentplus.org/organic-machine-discussion-human-relationships-nature/ (accessed on 7 March 2023).
- Geiling, N. How Oregon’s Second Largest City Vanished in a Day. Smithsonian Magazine. 2015. Available online: https://www.smithsonianmag.com/history/vanport-oregon-how-countrys-largest-housing-project-vanished-day-180954040/ (accessed on 7 March 2023).
- Blumm, M.C.; Thorson, E.J.; Smith, J.D. Practiced at the Art of Deception: The Failure of Columbia Basin Salmon Recovery Under the Endangered Species Act. Environ. Law 2006, 36, 709–810. [Google Scholar]
- Blumm, M.C.; Erickson, A. Dam Removal in the Pacific Northwest: Lessons for the Nation. Environ. Law 2012, 42, 1043–1100. [Google Scholar] [CrossRef]
- Federal Energy Regulatory Commission. Order Modifying and Approving Surrender of License and Rrmoval of Project Facilities, 181 FERC 61,122. 2022. Available online: https://www.ferc.gov/media/h-1-p-2082-063 (accessed on 7 March 2023).
- Cohen, B.R. Escaping the false binary of nature and culture through connection: Richard White’s The Organic Machine: The Remaking of the Columbia River. Organ. Environ. 2005, 18, 445–457. [Google Scholar] [CrossRef]
- Lackey, R.T.; Lach, D.H.; Duncan, S.L. (Eds.) The Challenge of Restring Wild Salmon. In Salmon 2100, The Future of Wild Pacific Salmon; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 1–11. [Google Scholar]
- Mantua, N.J.; Hare, S.R. The Pacific Decadal Oscillation. J. Oceanogr. 2002, 58, 35–44. [Google Scholar] [CrossRef]
- Di Lorenzo, E.; Schneider, N.; Cobb, K.M.; Franks PJ, S.; Chhak, K.; Miller, A.J.; McWilliams, J.C.; Bograd, S.J.; Arango, H.; Curchitser, E.; et al. North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 2008, 35, L08607. [Google Scholar] [CrossRef]
- Tootle, G.A.; Piechota, T.C.; Singh, A. Coupled oceanic-atmospheric vatiability and US streamflow. Water Resour. Res. 2005, 41, 1–11. [Google Scholar] [CrossRef]
- Bottom, D.L.; Jones, K.K.; Simenstad, C.A.; Smith, C.L. Reconnecting social and ecological resilience in salmon ecosystems. Ecol. Soc. 2009, 14, 5. Available online: http://www.ecologyandsociety.org/vol14/iss1/art5/ (accessed on 7 March 2023). [CrossRef]
- Ford, P. Declining Salmon Counts on Salmon River Flashing Red Light Warning Extinction. The Columbia Basin Bulletin, Fish & Wildlife News. 2022. Available online: https://cbbulletin.com/guest-column-declining-salmon-counts-on-salmon-river-flashing-red-light-warning-extinction/ (accessed on 7 March 2023).
- Herbold, B.; Carlson, S.M.; Henery, R.; Johnson, R.C.; Mantua, N.; McClure, M.; Moyle, P.; Sommer, T. Managing for Salmon Resilience in California’s Variable and Changing Climate. San Fr. Estuary Watershed Sci. 2018, 16, 3. [Google Scholar] [CrossRef]
- Mason, J.G.; Eurich, J.G.; Lau, J.D.; Battista, W.; Free, C.M.; Mills, K.E.; Tokunaga, K.; Zhao, L.Z.; Dickey-Collas, M.; Valle, M.; et al. Attributes of climate resilience in fisheries: From theory to practice. Fish Fish. 2022, 23, 522–544. [Google Scholar] [CrossRef]
- Hartman, G.F.; Groot, C.; Northcote, T.G. Science and Management in Sustainable Salmonid Fisheries: The Ball is Not in Our Court. In Sustainable Fisheries Management, Pacific Salmon; Knudsen, E.E., McDonald, D., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 1–752. [Google Scholar]
- Lackey, R.T. The great Pacific Northwest Salmon conundrum. Water Resour. IMPACT (Am. Water Resour. Assoc.) 2022, 24, 5–8. [Google Scholar]
- Northwest Power and Conservation Council. Guide for Subbasin Planners. 2001. Council Document Number: 2001-20. Available online: https://www.nwcouncil.org/reports/technical-guide-subbasin-planners/ (accessed on 7 March 2023).
- Hill, G.; Kolmes, S.A.; Humphreys, M.; McLain, R.; Jones, E.T. Using decision support tools in multi-stakeholder environmental planning: Restorative justice and subbasin planning in the Columbia River Basin. J. Environ. Stud. Sci. 2019, 9, 170–186. [Google Scholar] [CrossRef]
- Hill, G.; Monticino, M.; Jones, E.T.; Kolmes, S.; McLain, R. Aspirational Goals and Incremental Tools: Does forecasting exclude other frameworks for Strategic planning? In Proceedings of the UC Berkeley: Computer Professionals for Social Responsibility, Directions and Implications of Advanced Computing Conference Proceedings, Berkeley, CA, USA, 26–29 June 2008; Available online: http://www.publicsphereproject.org/events/diac08/proceedings/index.html (accessed on 7 March 2023).
- Rahr, R.; Augerot, A. A proactive sanctuary strategy to anchor and restore high-priority wild salmon ecosystems. In Salmon 2100, The Future of Wild Pacific Salmon; Lackey, R.T., Lach, D.H., Duncan, S.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2006; pp. 465–489. [Google Scholar]
- Schloss, C.A.; Lawler, J.J.; Larson, E.R.; Papendick, H.L.; Case, M.J.; Evans, D.M.; DeLap, J.H.; Langdon, J.G.R.; Hall, S.A.; McRae, B.H. Systematic Conservation Planning in the Face of Climate Change: Bet-Hedging on the Columbia Plateau. PLoS ONE 2011, 6, e28788. [Google Scholar] [CrossRef] [PubMed]
- Senate Hearing 111-833, S. 817. The Pacific Salmon Stronghold Conservation Act. The U.S. Government Publishing Office. pp. 1–94. Available online: https://www.govinfo.gov/content/pkg/CHRG-111shrg64140/html/CHRG-111shrg64140.htm (accessed on 7 March 2023).
- “Salmon Focused Investments in Sustainable Habitats Act” Is Available at Congressman Huffman’s Website. Available online: https://huffman.house.gov/imo/media/doc/Huffman%20Salmon%20FISH%20Act%20117th%20FINAL.pdf (accessed on 7 March 2023).
- Columbia River Inter-Tribal Fish Commission. Spirit of the Salmon: Wy-Kan-Ush-Mi Wa-Kish-Wit. Available online: https://plan.critfc.org/ (accessed on 14 March 2013).
- Columbia Basin Partnership Task Force (CBPTF). A Vision for Salmon and Steelhead: Goals to Restore Thriving Salmon and Steelhead to the Columbia River Basin: Phase 2 Report of the Columbia Basin Partnership Task Force of the Marine Fisheries Advisory Committee. October 2020. Available online: https://www.fisheries.noaa.gov/vision-salmon-and-steelhead-goals-restore-thriving-salmon-and-steelhead-columbia-river-basin (accessed on 16 March 2023).
Figure 1.
Precontact stability landscape. The yellow circle represents the state of the system.

Figure 2.
The refugia regime begins to appear. The yellow circle represents the state of the system.
Figure 2.
The refugia regime begins to appear. The yellow circle represents the state of the system.

Figure 3.
The techno regime is created by the introduction of hatcheries. The yellow circle represents the state of the system.
Figure 3.
The techno regime is created by the introduction of hatcheries. The yellow circle represents the state of the system.

Figure 4.
Critical transition to the refugia regime. The yellow circle represents the state of the system.
Figure 4.
Critical transition to the refugia regime. The yellow circle represents the state of the system.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hill, G.M.; Kolmes, S.A. A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies. Environments 2023, 10, 76. https://doi.org/10.3390/environments10050076
AMA Style
Hill GM, Kolmes SA. A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies. Environments. 2023; 10(5):76. https://doi.org/10.3390/environments10050076
Chicago/Turabian StyleHill, Gregory M., and Steven A. Kolmes. 2023. "A Resilience History of the Columbia River Basin and Salmonid Species: Regimes and Policies" Environments 10, no. 5: 76. https://doi.org/10.3390/environments10050076
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.