

Differential Anti-Tumor Effects of IFN-Inducible Chemokines CXCL9, CXCL10, and CXCL11 on a Mouse Squamous Cell Carcinoma Cell Line

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Lentiviral Expression Vectors
2.3. Generation of Cell Lines Stably Expressing the IFN-Inducible Chemokines
2.4. Cell Proliferation Assay
2.5. Transplantation of a Cell Line Stably Expressing IFN-Inducible Chemokines into Nude Mice
2.6. IHC Analysis
2.7. Statistical Analyses
3. Results
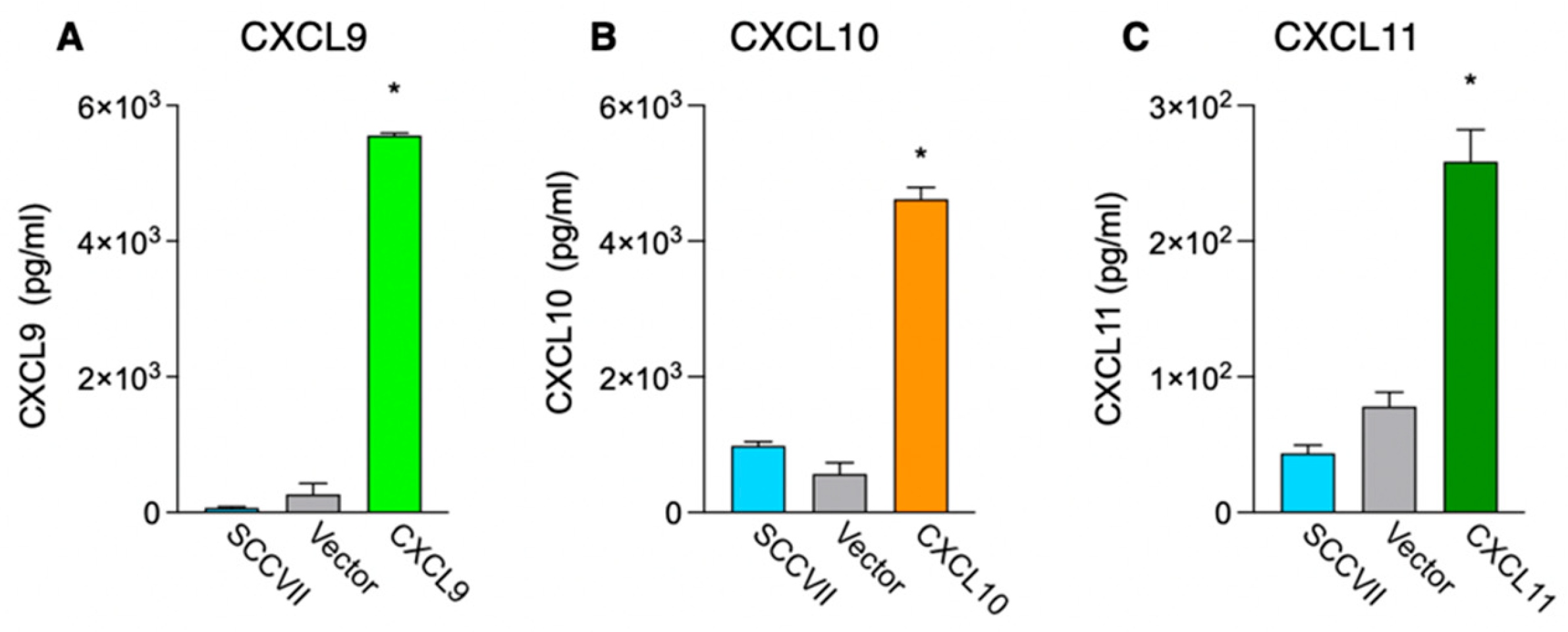
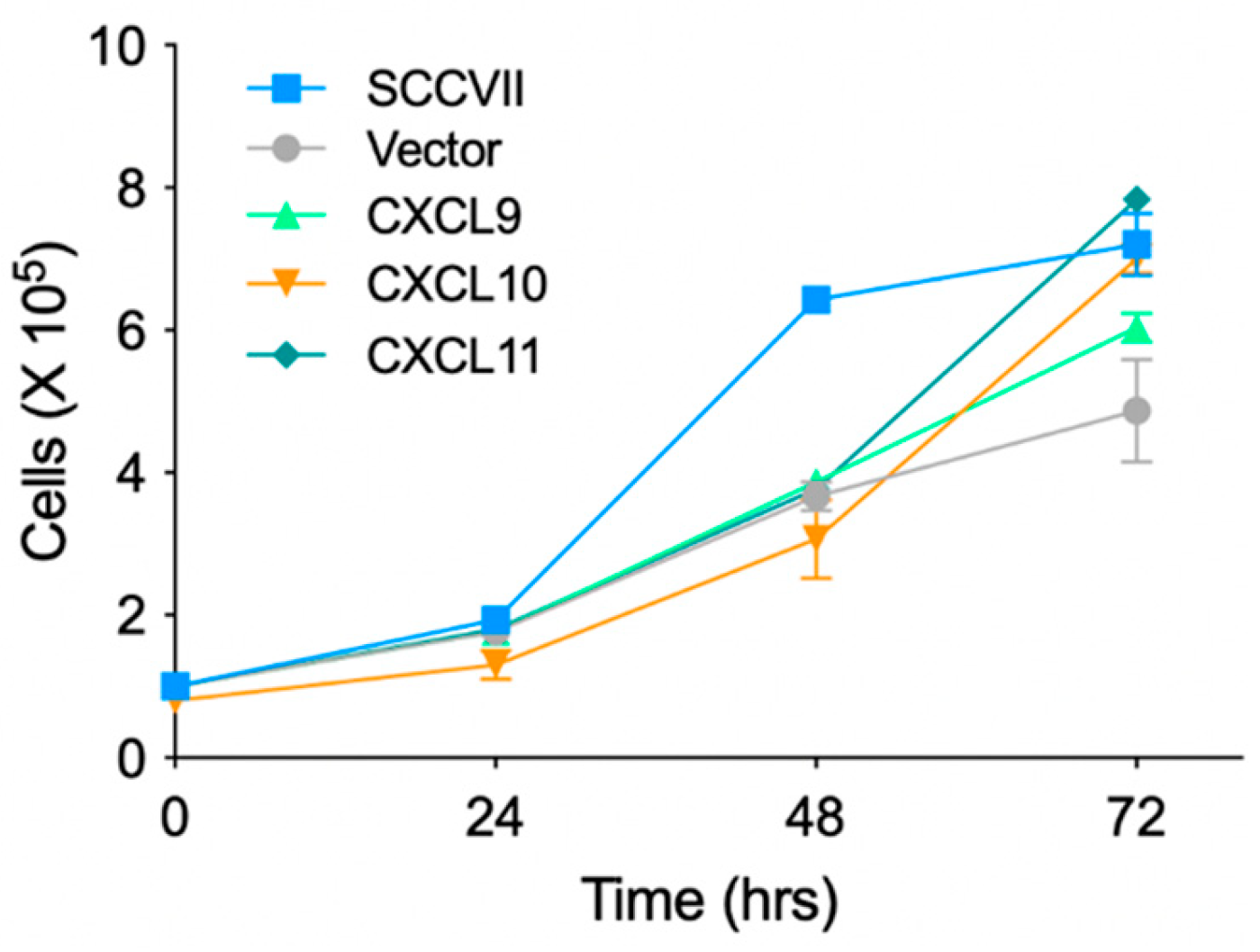
3.1. Generation of Cell Lines Stably Expressing IFN-Inducible Chemokines and Investigation of Their Proliferation In Vitro
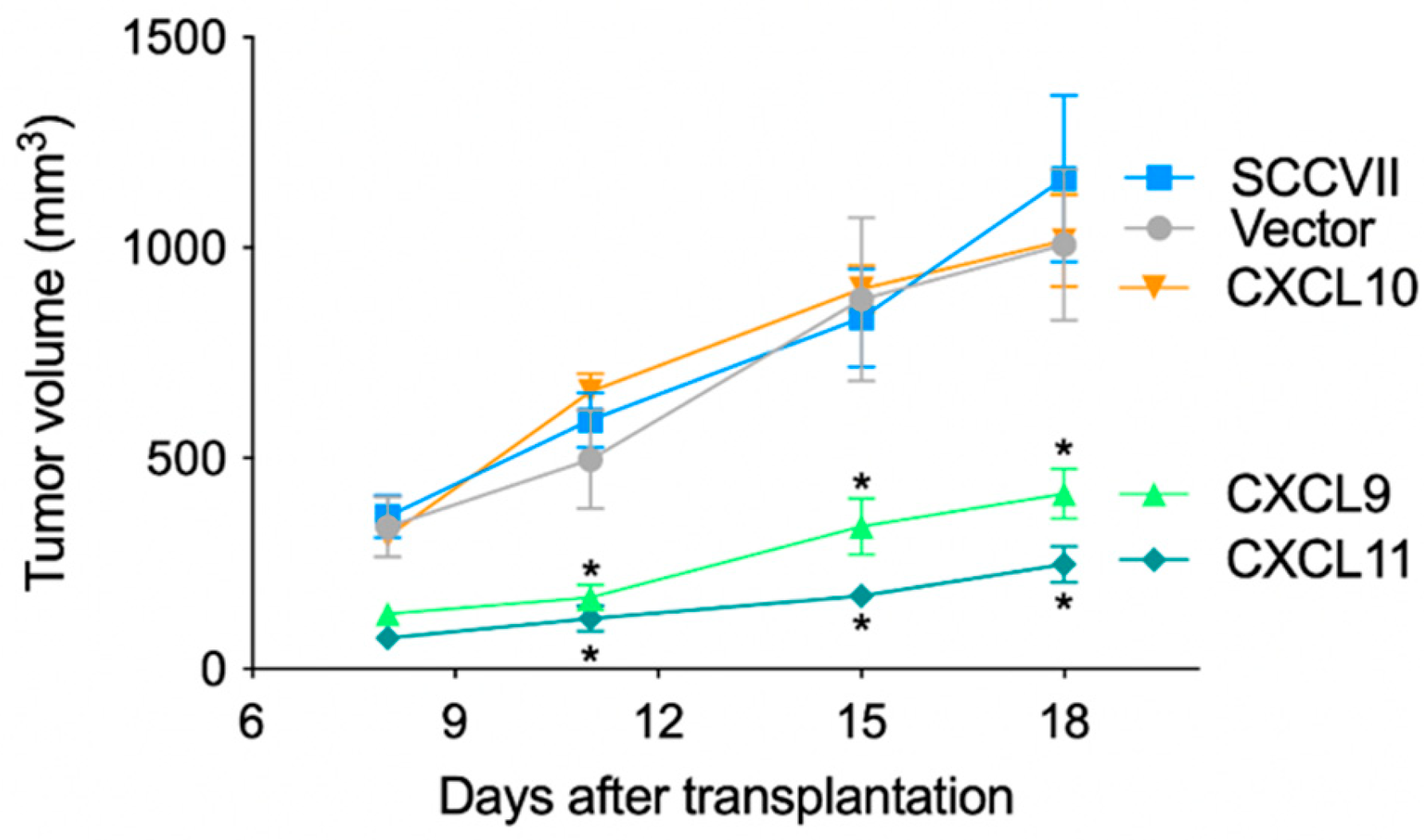
3.2. Anti-Tumor Effects of IFN-Inducible Chemokines in Nude Mice
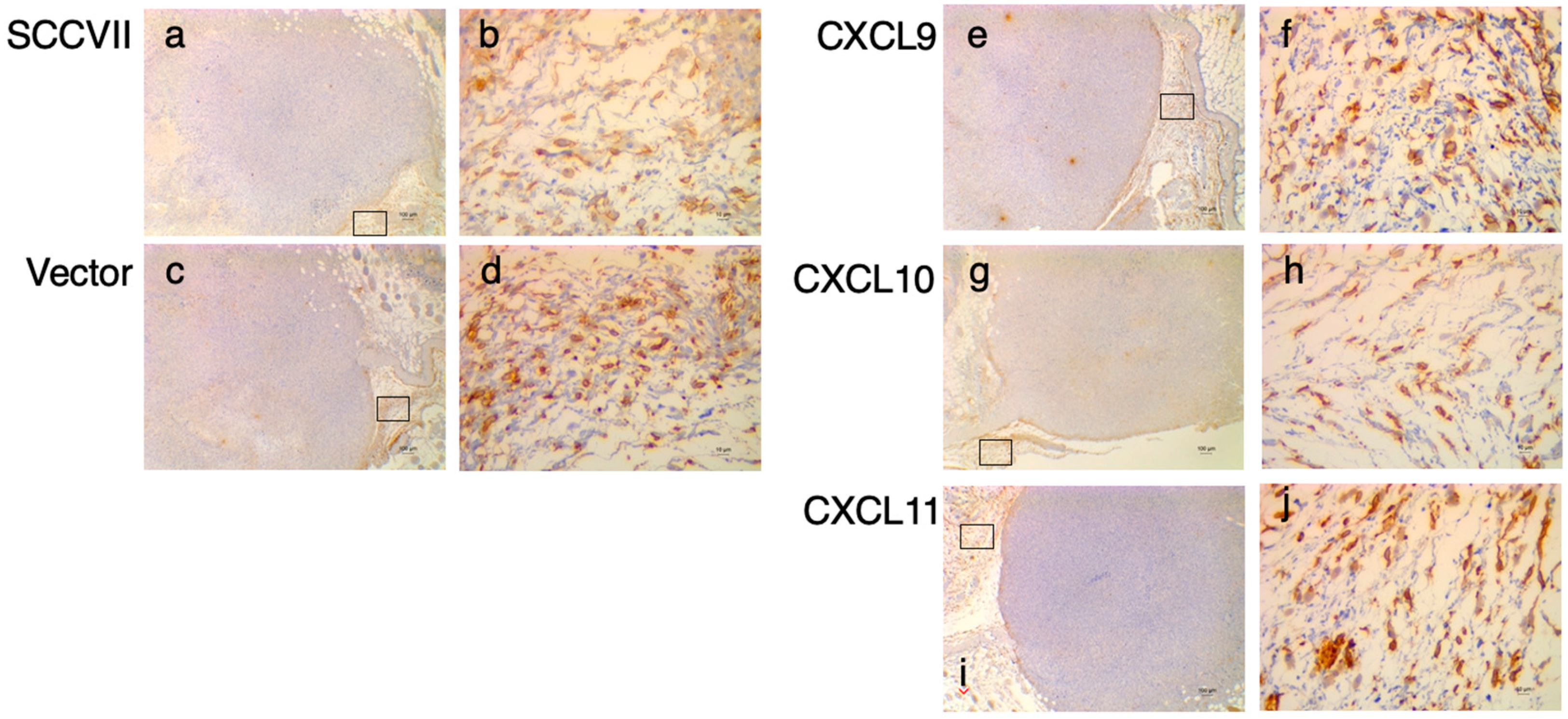
3.3. IHC Study of DPP4 in Tumor Tissues Expressing the IFN-Inducible Chemokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and Chemokine Receptors: Positioning Cells for Host Defense and Immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef]
- Schulz, O.; Hammerschmidt, S.I.; Moschovakis, G.L.; Förster, R. Chemokines and Chemokine Receptors in Lymphoid Tissue Dynamics. Annu. Rev. Immunol. 2016, 34, 203–242. [Google Scholar] [CrossRef]
- Farber, J.M. A Macrophage mRNA Selectively Induced by Gamma-Interferon Encodes a Member of the Platelet Factor 4 Family of Cytokines. Proc. Natl. Acad. Sci. USA 1990, 87, 5238–5242. [Google Scholar] [CrossRef]
- Luster, A.D.; Unkeless, J.C.; Ravetch, J.V. Gamma-Interferon Transcriptionally Regulates an Early-Response Gene Containing Homology to Platelet Proteins. Nature 1985, 315, 672–676. [Google Scholar] [CrossRef]
- Ohmori, Y.; Hamilton, T.A. A Macrophage LPS-Inducible Early Gene Encodes the Murine Homologue of IP-10. Biochem. Biophys. Res. Commun. 1990, 168, 1261–1267. [Google Scholar] [CrossRef]
- Vanguri, P.; Farber, J.M. Identification of CRG-2. An Interferon-Inducible mRNA Predicted to Encode a Murine Monokine. J. Biol. Chem. 1990, 265, 15049–15057. [Google Scholar] [CrossRef] [PubMed]
- Cole, K.E.; Strick, C.A.; Paradis, T.J.; Ogborne, K.T.; Loetscher, M.; Gladue, R.P.; Lin, W.; Boyd, J.G.; Moser, B.; Wood, D.E.; et al. Interferon-Inducible T Cell Alpha Chemoattractant (I-TAC): A Novel non-ELR CXC Chemokine with Potent Activity on Activated T Cells through Selective High Affinity Binding to CXCR3. J. Exp. Med. 1998, 187, 2009–2021. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Erdel, M.; Duba, H.C.; Werner, E.R.; Werner-Felmayer, G. Cloning, Genomic Sequence, and Chromosome Mapping of Scyb11, the Murine Homologue of SCYB11 (Alias betaR1/H174/SCYB9B/I-TAC/IP-9/CXCL11). Cytogenet. Cell Genet. 2000, 88, 278–282. [Google Scholar] [CrossRef]
- Singh, A.K.; Arya, R.K.; Trivedi, A.K.; Sanyal, S.; Baral, R.; Dormond, O.; Briscoe, D.M.; Datta, D. Chemokine Receptor Trio: CXCR3, CXCR4 and CXCR7 Crosstalk via CXCL11 and CXCL12. Cytokine Growth Factor Rev. 2013, 24, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Loetscher, M.; Gerber, B.; Loetscher, P.; Jones, S.A.; Piali, L.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. Chemokine Receptor Specific for IP-10 and Mig: Structure, Function, and Expression in Activated T-Lymphocytes. J. Exp. Med. 1996, 184, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Groom, J.R.; Luster, A.D. CXCR3 Ligands: Redundant, Collaborative and Antagonistic Functions. Immunol. Cell Biol. 2011, 89, 207–215. [Google Scholar] [CrossRef]
- Angiolillo, A.L.; Sgadari, C.; Taub, D.D.; Liao, F.; Farber, J.M.; Maheshwari, S.; Kleinman, H.K.; Reaman, G.H.; Tosato, G. Human Interferon-Inducible protein 10 Is a Potent Inhibitor of Angiogenesis In Vivo. J. Exp. Med. 1995, 182, 155–162. [Google Scholar] [CrossRef]
- Lasagni, L.; Francalanci, M.; Annunziato, F.; Lazzeri, E.; Giannini, S.; Cosmi, L.; Sagrinati, C.; Mazzinghi, B.; Orlando, C.; Maggi, E.; et al. An Alternatively Spliced Variant of CXCR3 Mediates the Inhibition of Endothelial Cell Growth Induced by IP-10, Mig, and I-TAC, and Acts as Functional Receptor for Platelet Factor 4. J. Exp. Med. 2003, 197, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, J.E.; Addison, C.A.; Burdick, M.D.; Kunkel, S.L.; Strieter, R.M. Identification and Partial Characterization of a Variant of Human CXCR3 Generated by Posttranscriptional Exon Skipping. J. Immunol. 2004, 173, 6234–6240. [Google Scholar] [CrossRef] [PubMed]
- Arenberg, D.A.; Kunkel, S.L.; Polverini, P.J.; Morris, S.B.; Burdick, M.D.; Glass, M.C.; Taub, D.T.; Iannettoni, M.D.; Whyte, R.I.; Strieter, R.M. Interferon-Gamma-Inducible protein 10 (IP-10) Is an Angiostatic Factor That Inhibits Human Non-Small Cell Lung Cancer (NSCLC) Tumorigenesis and Spontaneous Metastases. J. Exp. Med. 1996, 184, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Sgadari, C.; Angiolillo, A.L.; Cherney, B.W.; Pike, S.E.; Farber, J.M.; Koniaris, L.G.; Vanguri, P.; Burd, P.R.; Sheikh, N.; Gupta, G.; et al. Interferon-Inducible protein-10 Identified as a Mediator of Tumor Necrosis In Vivo. Proc. Natl. Acad. Sci. USA 1996, 93, 13791–13796. [Google Scholar] [CrossRef]
- Hensbergen, P.J.; Wijnands, P.G.; Schreurs, M.W.; Scheper, R.J.; Willemze, R.; Tensen, C.P. The CXCR3 Targeting Chemokine CXCL11 Has Potent Antitumor Activity In Vivo Involving Attraction of CD8+ T Lymphocytes but Not Inhibition of Angiogenesis. J. Immunother. 2005, 28, 343–351. [Google Scholar] [CrossRef]
- Yang, X.; Chu, Y.; Wang, Y.; Zhang, R.; Xiong, S. Targeted In Vivo Expression of IFN-Gamma-Inducible protein 10 Induces Specific Antitumor Activity. J. Leukoc. Biol. 2006, 80, 1434–1444. [Google Scholar] [CrossRef]
- Gorbachev, A.V.; Kobayashi, H.; Kudo, D.; Tannenbaum, C.S.; Finke, J.H.; Shu, S.; Farber, J.M.; Fairchild, R.L. CXC Chemokine Ligand 9/Monokine Induced by IFN-Gamma Production by Tumor Cells Is Critical for T Cell-Mediated Suppression of Cutaneous Tumors. J. Immunol. 2007, 178, 2278–2286. [Google Scholar] [CrossRef]
- Kondo, T.; Nakazawa, H.; Ito, F.; Hashimoto, Y.; Osaka, Y.; Futatsuyama, K.; Toma, H.; Tanabe, K. Favorable Prognosis of Renal Cell Carcinoma with Increased Expression of Chemokines Associated with a Th1-Type Immune Response. Cancer Sci. 2006, 97, 780–786. [Google Scholar] [CrossRef]
- Sato, E.; Fujimoto, J.; Toyoki, H.; Sakaguchi, H.; Alam, S.M.; Jahan, I.; Tamaya, T. Expression of IP-10 Related to Angiogenesis in Uterine Cervical Cancers. Br. J. Cancer 2007, 96, 1735–1739. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, X.; Han, X.; Li, Z.; Zhu, Q.; Yan, J.; Yu, S.; Jin, Z.; Wang, Z.; Zheng, Q.; et al. The Chemokine CXCL9 Expression Is Associated with Better Prognosis for Colorectal Carcinoma Patients. Biomed. Pharmacother. 2016, 78, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Kistner, L.; Doll, D.; Holtorf, A.; Nitsche, U.; Janssen, K.P. Interferon-Inducible CXC-Chemokines Are Crucial Immune Modulators and Survival Predictors in Colorectal Cancer. Oncotarget 2017, 8, 89998–90012. [Google Scholar] [CrossRef]
- Sato, Y.; Motoyama, S.; Nanjo, H.; Wakita, A.; Yoshino, K.; Sasaki, T.; Nagaki, Y.; Liu, J.; Imai, K.; Saito, H.; et al. CXCL10 Expression Status Is Prognostic in Patients with Advanced Thoracic Esophageal Squamous Cell Carcinoma. Ann. Surg. Oncol. 2016, 23, 936–942. [Google Scholar] [CrossRef]
- Dangaj, D.; Bruand, M.; Grimm, A.J.; Ronet, C.; Barras, D.; Duttagupta, P.A.; Lanitis, E.; Duraiswamy, J.; Tanyi, J.L.; Benencia, F.; et al. Cooperation between Constitutive and Inducible Chemokines Enables T Cell Engraftment and Immune Attack in Solid Tumors. Cancer Cell 2019, 35, 885–900.e10. [Google Scholar] [CrossRef]
- Ruiz-Garcia, E.; Scott, V.; Machavoine, C.; Bidart, J.M.; Lacroix, L.; Delaloge, S.; Andre, F. Gene Expression Profiling Identifies Fibronectin 1 and CXCL9 as Candidate Biomarkers for Breast Cancer Screening. Br. J. Cancer 2010, 102, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Ejaeidi, A.A.; Craft, B.S.; Puneky, L.V.; Lewis, R.E.; Cruse, J.M. Hormone Receptor-Independent CXCL10 Production Is Associated with the Regulation of Cellular Factors Linked to Breast Cancer Progression and Metastasis. Exp. Mol. Pathol. 2015, 99, 163–172. [Google Scholar] [CrossRef]
- Chang, K.P.; Wu, C.C.; Fang, K.H.; Tsai, C.Y.; Chang, Y.L.; Liu, S.C.; Kao, H.K. Serum Levels of Chemokine (C-X-C Motif) Ligand 9 (CXCL9) Are Associated with Tumor Progression and Treatment Outcome in Patients with Oral Cavity Squamous Cell Carcinoma. Oral Oncol. 2013, 49, 802–807. [Google Scholar] [CrossRef]
- Hsin, L.J.; Kao, H.K.; Chen, I.H.; Tsang, N.M.; Hsu, C.L.; Liu, S.C.; Chang, Y.S.; Chang, K.P. Serum CXCL9 Levels Are Associated with Tumor Progression and Treatment Outcome in Patients with Nasopharyngeal Carcinoma. PLoS ONE 2013, 8, e80052. [Google Scholar] [CrossRef]
- Yang, B.; Dong, K.; Guo, P.; Guo, P.; Jie, G.; Zhang, G.; Li, T. Identification of Key Biomarkers and Potential Molecular Mechanisms in Oral Squamous Cell Carcinoma by Bioinformatics Analysis. J. Comput. Biol. 2020, 27, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Rainczuk, A.; Rao, J.R.; Gathercole, J.L.; Fairweather, N.J.; Chu, S.; Masadah, R.; Jobling, T.W.; Deb-Choudhury, S.; Dyer, J.; Stephens, A.N. Evidence for the antagonistic form of CXC-motif chemokine CXCL10 in serous epithelial ovarian tumours. Int. J. Cancer 2014, 134, 530–541. [Google Scholar] [CrossRef]
- Shiels, M.S.; Katki, H.A.; Hildesheim, A.; Pfeiffer, R.M.; Engels, E.A.; Williams, M.; Kemp, T.J.; Caporaso, N.E.; Pinto, L.A.; Chaturvedi, A.K. Circulating Inflammation Markers, Risk of Lung Cancer, and Utility for Risk Stratification. J. Natl. Cancer Inst. 2015, 107, djv199. [Google Scholar] [CrossRef]
- Proost, P.; Schutyser, E.; Menten, P.; Struyf, S.; Wuyts, A.; Opdenakker, G.; Detheux, M.; Parmentier, M.; Durinx, C.; Lambeir, A.M.; et al. Amino-Terminal Truncation of CXCR3 Agonists Impairs Receptor Signaling and Lymphocyte Chemotaxis, While Preserving Antiangiogenic Properties. Blood 2001, 98, 3554–3561. [Google Scholar] [CrossRef] [PubMed]
- Metzemaekers, M.; Van Damme, J.; Mortier, A.; Proost, P. Regulation of Chemokine Activity—A Focus on the Role of Dipeptidyl Peptidase IV/CD26. Front. Immunol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed]
- Havre, P.A.; Abe, M.; Urasaki, Y.; Ohnuma, K.; Morimoto, C.; Dang, N.H. The Role of CD26/Dipeptidyl Peptidase IV in Cancer. Front. Biosci. 2008, 13, 1634–1645. [Google Scholar] [CrossRef] [PubMed]
- Casrouge, A.; Decalf, J.; Ahloulay, M.; Lababidi, C.; Mansour, H.; Vallet-Pichard, A.; Mallet, V.; Mottez, E.; Mapes, J.; Fontanet, A.; et al. Evidence for an Antagonist Form of the Chemokine CXCL10 in Patients Chronically Infected with HCV. J. Clin. Investig. 2011, 121, 308–317. [Google Scholar] [CrossRef]
- Kawada, K.; Sonoshita, M.; Sakashita, H.; Takabayashi, A.; Yamaoka, Y.; Manabe, T.; Inaba, K.; Minato, N.; Oshima, M.; Taketo, M.M. Pivotal Role of CXCR3 in Melanoma Cell Metastasis to Lymph Nodes. Cancer Res. 2004, 64, 4010–4017. [Google Scholar] [CrossRef] [PubMed]
- Zipin-Roitman, A.; Meshel, T.; Sagi-Assif, O.; Shalmon, B.; Avivi, C.; Pfeffer, R.M.; Witz, I.P.; Ben-Baruch, A. CXCL10 Promotes Invasion-Related Properties in Human Colorectal Carcinoma Cells. Cancer Res. 2007, 67, 3396–3405. [Google Scholar] [CrossRef] [PubMed]
- Amatschek, S.; Lucas, R.; Eger, A.; Pflueger, M.; Hundsberger, H.; Knoll, C.; Grosse-Kracht, S.; Schuett, W.; Koszik, F.; Maurer, D.; et al. CXCL9 Induces Chemotaxis, Chemorepulsion and Endothelial Barrier Disruption through CXCR3-Mediated Activation of Melanoma Cells. Br. J. Cancer 2011, 104, 469–479. [Google Scholar] [CrossRef]
- Ono, K.; Masunaga, S.; Akuta, K.; Akaboshi, M.; Abe, M. Radiosensitization of SCCⅦ Tumours and Normal Tissues by Nicotinamide and Carbogen: Analysis by Micronucleus Assay. Radiother. Oncol. 1993, 28, 162–167. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, B.W., Jr.; Cope, K.A.; Johnson, C.S.; Schwartz, M.R. A New Immunocompetent Murine Model for Oral Cancer. Arch. Otolaryngol. Head Neck Surg. 1997, 123, 20–24. [Google Scholar] [CrossRef]
- Khurana, D.; Martin, E.A.; Kasperbauer, J.L.; O’Malley, B.W., Jr.; Salomao, D.R.; Chen, L.; Strome, S.E. Characterization of a Spontaneously Arising Murine Squamous Cell Carcinoma (SCC Ⅶ) as a Prerequisite for Head and Neck Cancer Immunotherapy. Head Neck 2001, 23, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Nomura, T.; Noma, H.; Yokoo, K.; Takagi, R.; Hashimoto, S.; Okamoto, M.; Sato, M.; Yu, G.; Guo, C.; et al. Effect of YM529 on a Model of Mandibular Invasion by Oral Squamous Cell Carcinoma in Mice. Clin. Cancer Res. 2005, 11, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Hartley, J.L.; Temple, G.F.; Brasch, M.A. DNA Cloning Using In Vitro Site-Specific Recombination. Genome Res. 2000, 10, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, J.; Struyf, S.; Wuyts, A.; Van Coillie, E.; Menten, P.; Schols, D.; Sozzani, S.; De Meester, I.; Proost, P. The Role of CD26/DPP IV in Chemokine Processing. Chem. Immunol. 1999, 72, 42–56. [Google Scholar] [PubMed]
- Lambeir, A.M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-Peptidase IV from Bench to Bedside: An Update on Structural Properties, Functions, and Clinical Aspects of the Enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- De Zutter, A.; Van Damme, J.; Struyf, S. The Role of Post-Translational Modifications of Chemokines by CD26 in Cancer. Cancers 2021, 13, 4247. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Skin Fibrosis. Identification and Isolation of a Dermal Lineage with Intrinsic Fibrogenic Potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef]
- Venning, F.A.; Zornhagen, K.W.; Wullkopf, L.; Sjölund, J.; Rodriguez-Cupello, C.; Kjellman, P.; Morsing, M.; Hajkarim, M.C.; Won, K.J.; Erler, J.T.; et al. Deciphering the Temporal Heterogeneity of Cancer-Associated Fibroblast Subpopulations in Breast Cancer. J. Exp. Clin. Cancer Res. 2021, 40, 175. [Google Scholar] [CrossRef]
- Barreira da Silva, R.; Laird, M.E.; Yatim, N.; Fiette, L.; Ingersoll, M.A.; Albert, M.L. Dipeptidylpeptidase 4 Inhibition Enhances Lymphocyte Trafficking, Improving Both Naturally Occurring Tumor Immunity and Immunotherapy. Nat. Immunol. 2015, 16, 850–858. [Google Scholar] [CrossRef]
- Yokoyama, W.M.; Seaman, W.E. The Ly-49 and NKR-P1 Gene Families Encoding Lectin-Like Receptors on Natural Killer Cells: The NK Gene Complex. Annu. Rev. Immunol. 1993, 11, 613–635. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, A.; Hiroi, M.; Mori, K.; Yamamoto, N.; Ohmori, Y. Differential Anti-Tumor Effects of IFN-Inducible Chemokines CXCL9, CXCL10, and CXCL11 on a Mouse Squamous Cell Carcinoma Cell Line. Med. Sci. 2023, 11, 31. https://doi.org/10.3390/medsci11020031
Matsumoto A, Hiroi M, Mori K, Yamamoto N, Ohmori Y. Differential Anti-Tumor Effects of IFN-Inducible Chemokines CXCL9, CXCL10, and CXCL11 on a Mouse Squamous Cell Carcinoma Cell Line. Medical Sciences. 2023; 11(2):31. https://doi.org/10.3390/medsci11020031
Chicago/Turabian StyleMatsumoto, Ari, Miki Hiroi, Kazumasa Mori, Nobuharu Yamamoto, and Yoshihiro Ohmori. 2023. "Differential Anti-Tumor Effects of IFN-Inducible Chemokines CXCL9, CXCL10, and CXCL11 on a Mouse Squamous Cell Carcinoma Cell Line" Medical Sciences 11, no. 2: 31. https://doi.org/10.3390/medsci11020031